
Bio-Guided Fractionation of Retama raetam (Forssk.) Webb & Berthel Polar Extracts

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
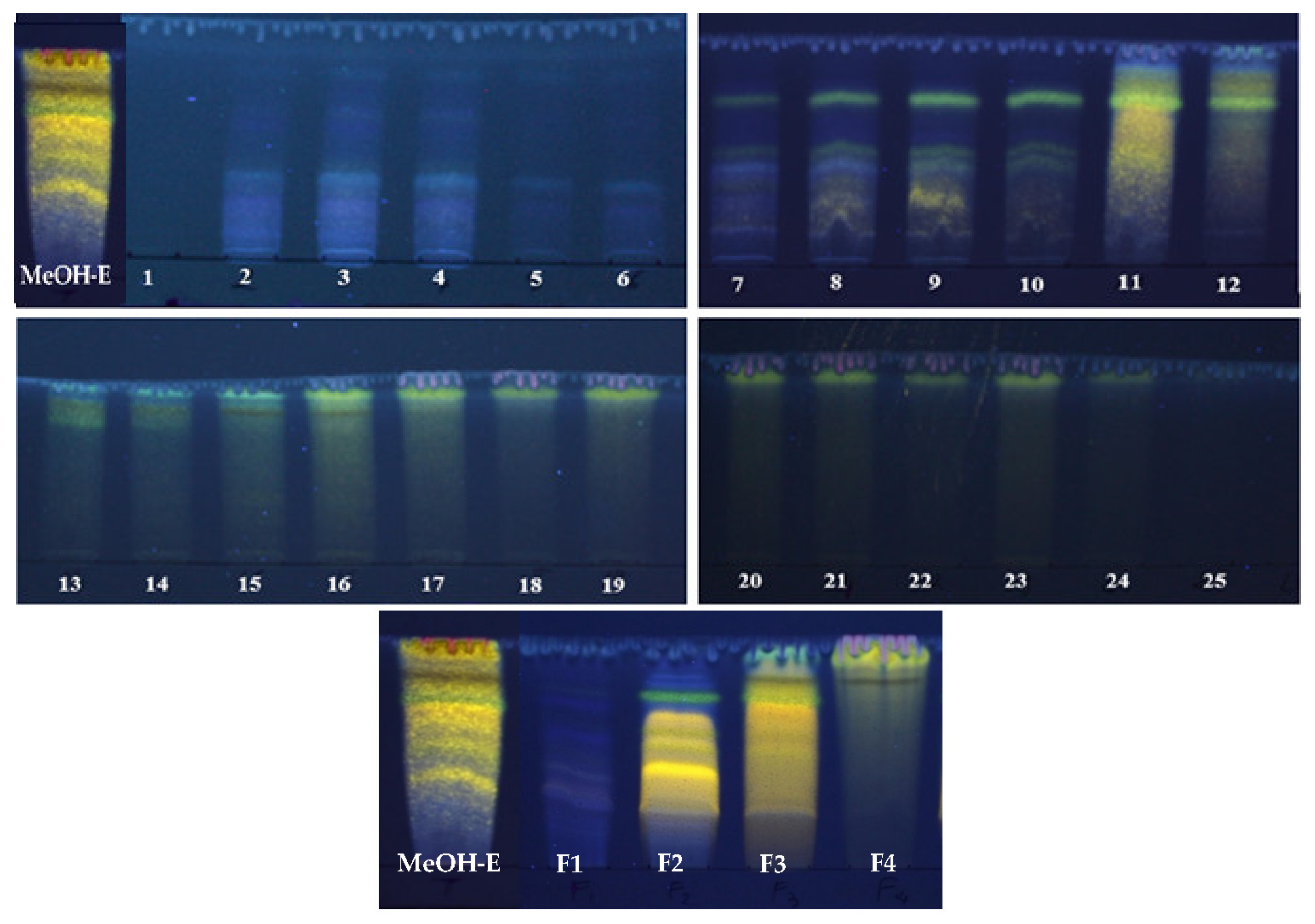
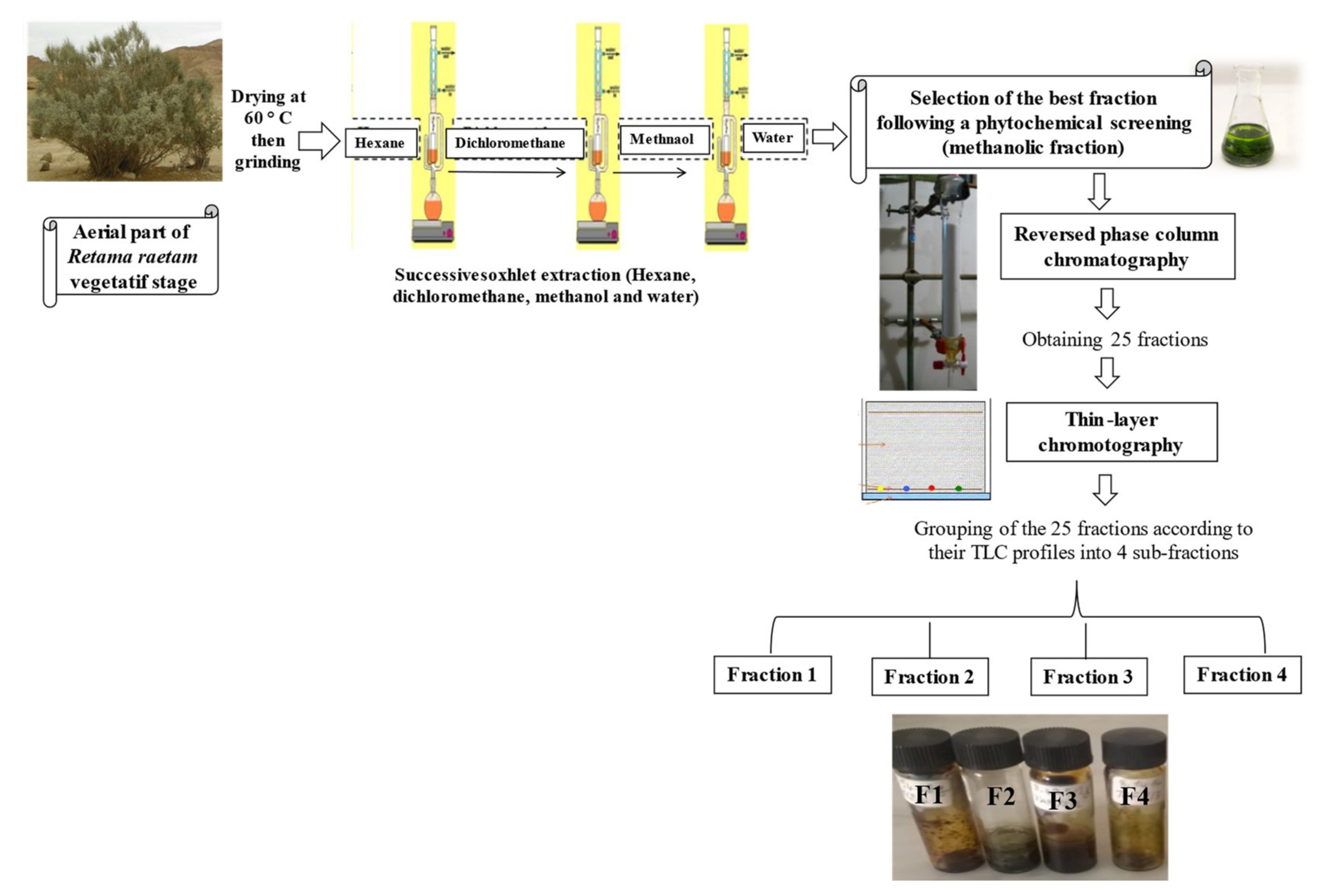
2.1. Fractionation of R. reatam Methanolic Extract and Phytochemical Composition
2.2. Antioxidant Activities
2.3. Cytotoxicity Assay
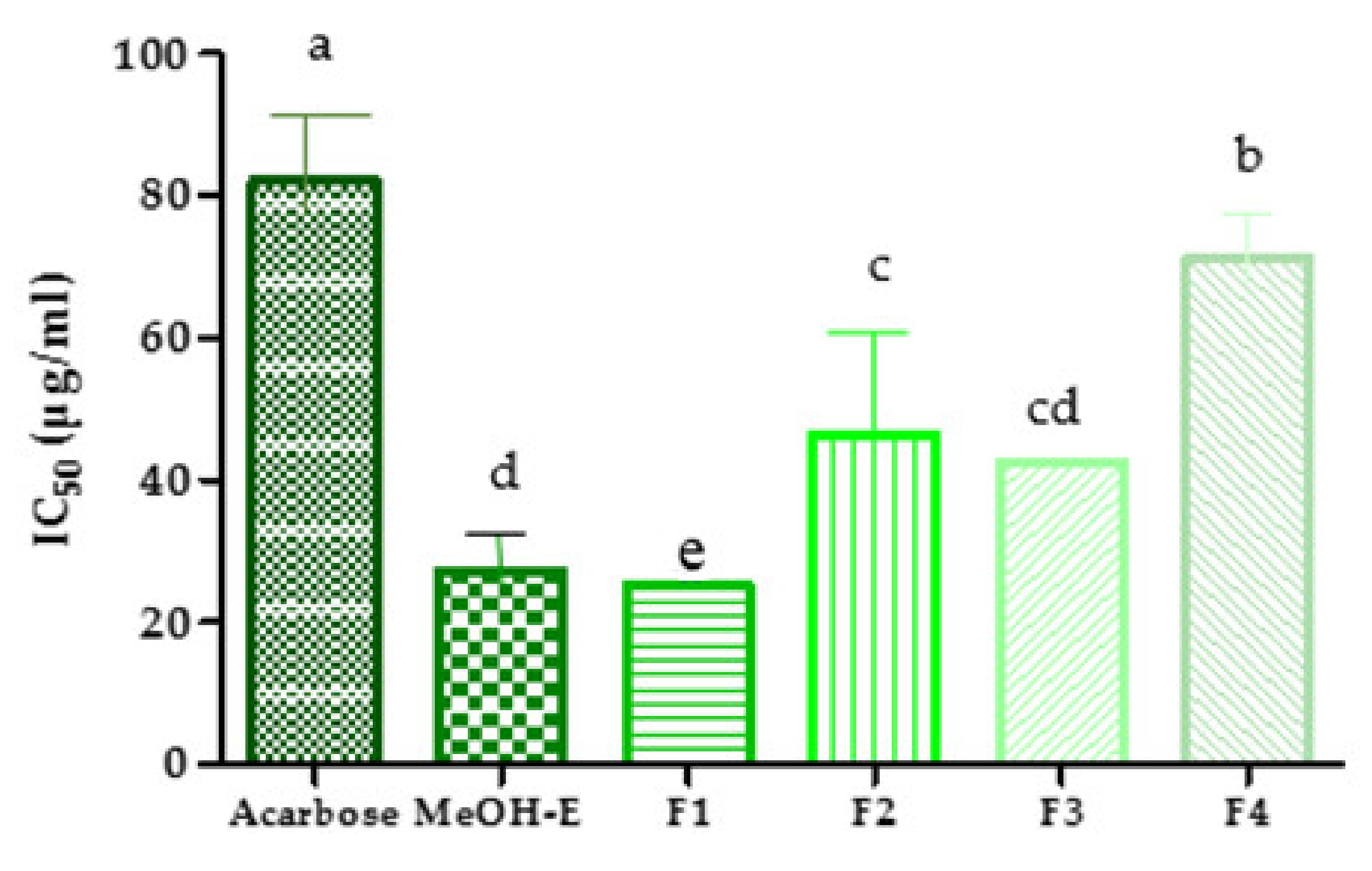
2.4. Inhibition of Pancreatic α-Amylase Activity
2.5. Antibacterial Activity
3. Materials and Methods
3.1. Plant Material and Fractionation
3.2. Phenolic Compounds
3.3. Antioxidant Activities
3.3.1. DPPH Quenching Ability
3.3.2. ORAC Assay
3.3.3. Antioxidant Cell Assay Using 2′,7′-Dichlorofluorescin-Diacetate (DCFH-DA)
3.4. Cytotoxicity Assay
3.5. Pancreatic α-Amylase Activity In Vitro Study
3.6. Antibacterial Activity
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P.C. Aromatic Plants as a Source of Bioactive Compounds. Agriculture 2012, 2, 228–243. [Google Scholar] [CrossRef] [Green Version]
- León-González, A.J.; Navarro, I.; Acero, N.; Muñoz Mingarro, D.; Martín-Cordero, C. Genus Retama: A review on traditional uses, phytochemistry, and pharmacological activities. Phytochem. Rev. 2018, 17, 701–731. [Google Scholar] [CrossRef]
- Mechergui, K.; Mahmoudi, H.; Larbi Khouja, M.; Jaouadi, W. Factors influencing seed germination of the pastoral plant Retama raetam subsp. bovei (Fabaceae): Interactive effects of fruit morphology, salinity, and osmotic stress. Biologija 2017, 63, 134–151. [Google Scholar] [CrossRef] [Green Version]
- Al-Tubuly, R.A.; Auzi, A.A.; Al-Etri-Endi, A.A.; Nahar, L.; Sarker, S.D. Effects of Retama raetam (Forssk.) Webb & Berthel. (Fabaceae) on the central nervous system in experimental animals. Arch. Biol. Sci. 2011, 63, 1015–1021. [Google Scholar]
- Omara, E.; Nada, S.A.; El-Toumy, S.A. Evaluation of hepatoprotective activity of the Retama raetam seeds on carbon tetrachloride-induced liver damage in rats. Planta Med. 2009, 75. [Google Scholar] [CrossRef]
- Maghrani, M.; Zeggwagh, N.A.; Haloui, M.; Eddouks, M. Acute diuretic effect of aqueous extract of Retama raetam in normal rats. J. Ethnopharmacol. 2005, 13, 31–35. [Google Scholar] [CrossRef]
- Maghrani, M.; Lemhadri, A.; Jouad, H.; Michel, J.B.; Eddouks., M. Effect of the desert plant Retama raetam on glycaemia in normal and streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2003, 87, 21–25. [Google Scholar] [CrossRef]
- Maghrani, M.; Lemhadri, A.; Zeggwagh, N.A.; El Amraoui, A.; Haloui, M.; Jouad, H.; Eddouks, M. Effect of Retama raetam on lipid metabolism in normal and recent-onset diabetic rats. J. Ethnopharmacol. 2004, 90, 323–329. [Google Scholar] [CrossRef]
- Edziri, H.; Ammar, S.; Groh, P.; Mahjoub, M.A.; Mastouri, M.; Gutmann, L.; Zine, M.; Laouni, M. Antimicrobial and cytotoxic activity of Marrubium alysson and Retama raetam grown in Tunisia. Pak. J. Biol. Sci. 2007, 10, 1759–1762. [Google Scholar]
- Edziri, H.; Ammar, S.; Groh, P.; Mahjoub, M.A.; Mastouri, M.; Gutmann, L.; Zine, M.; Laouni, M. Antibacterial, antifungal and cytotoxic activities of two flavonoids from Retama raetam flowers. Molecules 2012, 17, 7284–7293. [Google Scholar] [CrossRef] [Green Version]
- Djeddi, S.; Karioti, A.; Yannakopoulou, E.; Papadopoulos, K.; Chatter, R.; Helen Skaltsa, H. Analgesic and antioxidant activities of Algerian Retama raetam (Forssk.) Webb & Berthel extracts. Rec. Nat. Prod. 2013, 7, 169–176. [Google Scholar]
- El-Hamdani, N.; Filali-Ansari, N.; Rabiaa, F.; El Abbouyi, A.; El Khyari, S. Antifungal activity of the alkaloids extracts from aerial parts of Retama monosperma. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 965–971. [Google Scholar]
- Miguel, M.; Bouchmaaa, N.; Aazza, S. Antioxidant, anti-inflammatory and anti-acetylcholinesterase activities of eleven extracts of Moroccan plants. Fresenius Environ. Bull. 2014, 23, 1–14. [Google Scholar]
- Ksouri, R.; Megdiche, W.; Falleh, H.; Trabelsi, N.; Boulaaba, M.; Smaoui, A.; Abdelly, C. Influence of biological, environmental and technical factors on phenolic content and antioxidant activities of Tunisian halophytes. C. R. Biol. 2008, 331, 865–873. [Google Scholar] [CrossRef]
- Saada, M.; Falleh, H.; Jalleli, I.; Snoussi, M.; Ksouri, R. Phenolic profile, biological activities and fraction analysis of the medicinal halophyte Retama raetam. S. Afr. J. Bot. 2014, 94, 114–121. [Google Scholar]
- Saada, M.; Falleh, H.; Catarino, M.D.; Cardoso, S.M.; Ksouri, R. Plant Growth Modulates Metabolites and Biological Activities in Retama raetam (Forssk.) Webb. Molecules 2018, 23, 2177. [Google Scholar] [CrossRef] [Green Version]
- Najjaa, H.; Ben Arfa, A.; Doria, E.; Boubakri, A.; Trabelsi, N.; Falleh, H.; Tlili, H.; Neffati, M. Phenolic composition of some Tunisian medicinal plants associated with anti-proliferative effect on human breast cancer MCF-7 cells. EuroBiotech. J. 2020, 4, 104–112. [Google Scholar] [CrossRef]
- Hammouche-Mokrane, N.; León-González, A.J.; Navarro, I.; Boulila, F.; Benallaoua, S.; Martín-Cordero, C. Phytochemical Profile and Antibacterial Activity of Retama raetam and R. sphaerocarpa cladodes from Algeria. Biol. Nat. Prod. Commun. 2017, 12, 1857–1860. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.H.; Al-Rehaily, A.J.; Yousaf, M.; Ahmad, M.S.; Khan, S.I.; Khan, I.A. Two New Flavonoids from Retama raetam. Helv. Chim. Acta 2015, 98, 561–568. [Google Scholar] [CrossRef]
- Nur-E-Alam, M.; Yousaf, M.; Parveen, I.; Hafizur, R.M.; Ghani, U.; Ahmed, S.; Hameed, A.; Threadgill, M.D.; Al-Rehaily, A.J. Organic New flavonoids from the Saudi Arabian plant Retama raetam which stimulates secretion of insulin and inhibits α-glucosidase. Org. Biomol. Chem. 2019, 17, 1266–1276. [Google Scholar] [CrossRef] [Green Version]
- Azmir, J.; Zaidul, I.S.M.; Rahman, M.M.; Sharif, K.M.; Mohamed, A.; Sahena, F.; Jahurul, M.H.A. Techniques for extraction of bioactive compounds from plant materials: A review. J. Food Eng. 2013, 117, 426–436. [Google Scholar] [CrossRef]
- Kumarc, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar]
- Xu, M.J.; Wu, B.; Ding, T.; Chu, J.H.; Li, C.Y.; Zhang, J.; Wu, T.; Wu, J.; Liu, S.J.; Liu, S.L.; et al. Simultaneous characterization of prenylated flavonoids and isoflavonoids in Psoralea corylifolia L. by liquid chromatography with diode-array detection and quadrupole time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2012, 26, 2343. [Google Scholar] [CrossRef]
- Taamalli, A.; Abaza, L.; Román, D.A.; Carretero, A.S.; Gutiérrez, A.F.; Zarrouk, M.; Ben Youssef, N. Characterisation of Phenolic Compounds by HPLC–TOF/IT/MS in Buds and Open Flowers of ‘Chemlali’ Olive Cultivar. Phytochem. Anal. 2013, 24, 504–512. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Boussahel, S.; Cacciola, F.; Dahamna, S.; Mondello, L.; Saija, A.; Cimino, F.; Speciale, A.; Cristani, M. Flavonoid profile, antioxidant and antiglycation properties of Retama sphaerocarpa fruits extracts. Nat. Prod. Res. 2017, 24, 1–9. [Google Scholar] [CrossRef]
- Li, H.Q.; Luo, Y.; Qiao, C.H. The Mechanisms of Anticancer Agents by Genistein and Synthetic Derivatives of Isoflavone. Mini. Rev. Med. Chem. 2012, 12, 350–362. [Google Scholar]
- Conforti, F.; Statti, G.; Tundis, R.; Loizzo, M.R.; Bonesi, M.; Menichini, F.; Houghton, P.J. Antioxidant and cytotoxic activities of Retama raetam subsp. Gussonei. Phytother. Res. 2004, 18, 585–587. [Google Scholar] [CrossRef] [PubMed]
- Pereira, O.R.; Catarino, M.D.; Afonso, A.F.; Silva, A.M.S.; Cardoso, S.M. Salvia elegans, Salvia greggii and Salvia officinalis Decoctions: Antioxidant Activities and Inhibition of Carbohydrate and Lipid Metabolic Enzymes. Molecules 2018, 23, 3169. [Google Scholar] [CrossRef] [Green Version]
- Algandaby, M.M.; Alghamdi, H.A.; Ashour, O.M.; Abdel-Naim, A.B.; Ghareib, S.A.; Abdel-Sattar, E.A.; Hajar Abdulrahman, S. Mechanisms of the antihyperglycemic activity of Retama raetam in streptozotocin-induced diabetic rats. Food Chem. Toxicol. 2010, 48, 2448–2453. [Google Scholar] [CrossRef]
- Edziri, H.; Mastouri, M.; Ammar, S.; Matieu, M.; Patrich, G.; Hiar, R.; Mahjoub, M.A.; Ali, S.M.; Laurent, G.; Zine, M.; et al. Antimicrobial, antioxidant, and antiviral activities of Retama raetam (Forssk.) Webb flowers growing in Tunisia. World J. Microbiol. Biotechnol. 2008, 24, 2933–2940. [Google Scholar]
- Oueslati, S.; Ksouri, R.; Pichette, A.; Lavoie, S.; Girard-Lalancette, K.; Mshvildadze, V.; Abdelly, C.; Legault, J. A new flavonol glycoside from the medicinal halophyte Suaeda fruticosa. Nat. Prod. Res. 2014, 28, 960–966. [Google Scholar] [CrossRef]
- Wasli, H.; Jelali, N.; Ksouri, R.; Cardoso, S.M. Insights on the adaptation of Foeniculum vulgare Mill to iron deficiency. Appl. Sci. 2021, 11, 7072. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Saraiva, S.C.; Sobral, A.J.F.N.; Cardoso, S.M. Characterization of phenolic constituents and evaluation of antioxidant properties of leaves and stems of Eriocephalus africanus. Arab. J. Chem. 2018, 11, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Wasli, H.; Jelali, N.; Ellouzi, H.; Msaada, K.; Silva, A.M.S.; Cardoso, S.M.; Ksouri, R. Enzymatic and non-enzymatic antioxidant activities of dill (Anethum graveolens L.) in response to bicarbonate-induced oxidative damage. Agrochimica 2018, LXII, 320–335. [Google Scholar]
- Wasli, H.; Jelali, N.; Prior, R.L. Variation of polyphenolic composition, antioxidants and physiological characteristics of dill (Anethum graveolens L.) as affected by bicarbonate-induced iron deficiency conditions. Ind. Crop. Prod. 2018, 126, 466–476. [Google Scholar] [CrossRef]
- Legault, J.; Dahl, W.; Debiton, E.; Pichette, A.; Madelmont, J.C. Antitumor activity of balsam fir oil: Production of reactive oxygen species induced by alpha-humulene as possible mechanism of action. Planta Med. 2003, 69, 402–407. [Google Scholar] [PubMed]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef]
- Pereira, O.R.; Macias, R.I.R.; Domingues, M.R.M.; Marin, J.J.G.; Cardoso, S.M. Hepatoprotection of Mentha aquatica L., Lavandula dentata L. and Leonurus cardiaca L. Antioxidants 2019, 8, 267. [Google Scholar] [CrossRef] [Green Version]
- Bagamboula, M.; Uyttendaele, J.; Debevere, M. Inhibitory effect of thyme and basil essential oils, carvacrol, thymol, estragol, linalool and p-cymene towards Shigella sonnei and S. flexneri. Food Microbiol. 2003, 21, 33–42. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Yield (%) | TPC (mg GAE/g DR) | TFC (mg Catechin/g DR) |
|---|---|---|---|
| MeOH-E | - | 148.20 ± 0.74 b | 11.50 ± 2.14 b |
| F1 | 19.80 ± 0.99 c | 52.00 ± 0.05 c | 2.12 ± 0.01 d |
| F2 | 24.90 ± 0.12 b | 342.60 ± 0.04 a | 33.40 ± 0.03 a |
| F3 | 40.90 ± 0.2 a | 364.80 ± 0.03 a | 35.70 ± 0.18 a |
| F4 | 14.20 ± 0.71 d | 48.27 ± 0.06 d | 4.20 ± 0.12 c |
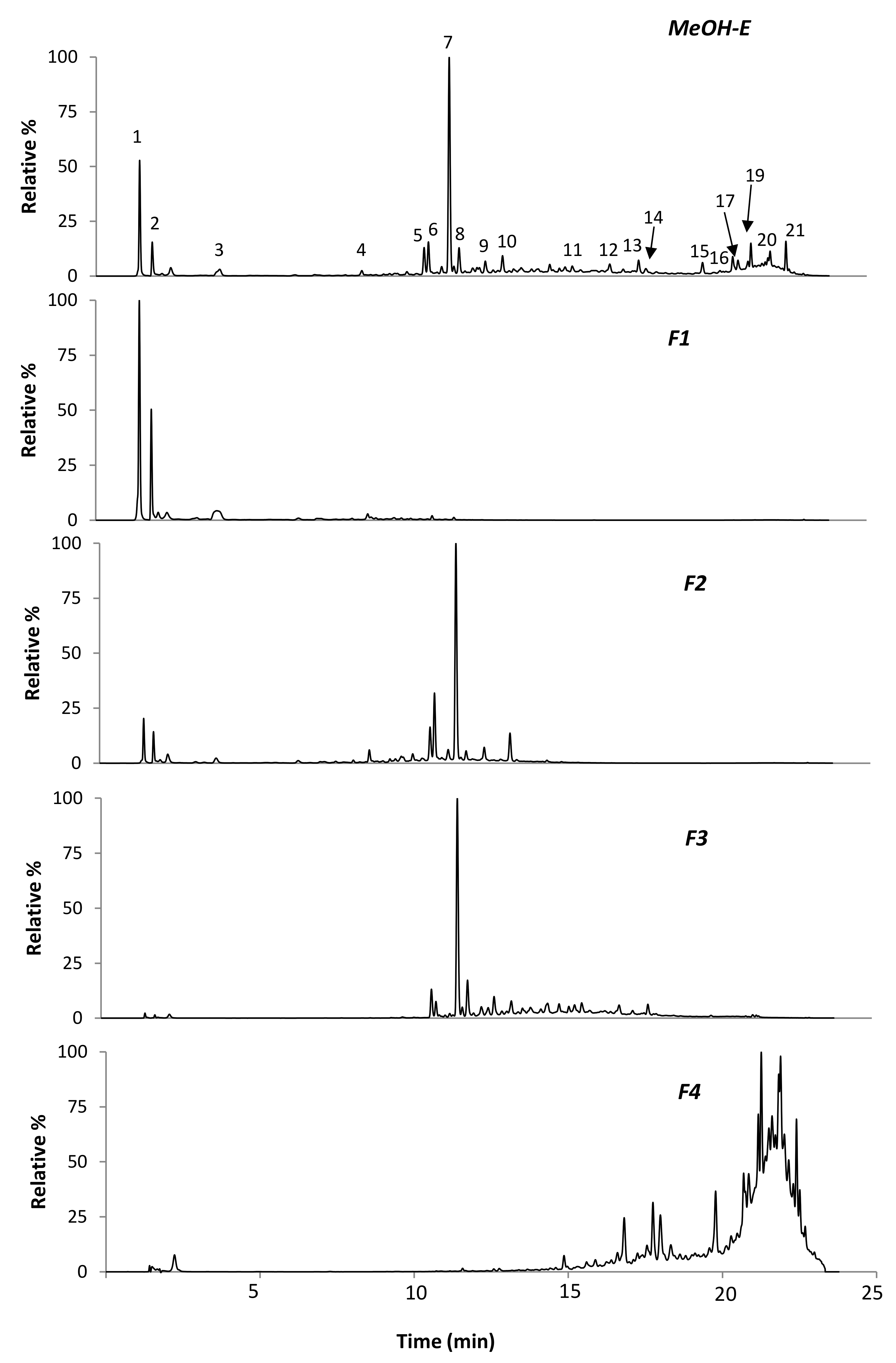
| Peak | RT (min) | λmax (nm) | m/z | ESI-MSn | Proposed Compound | MeOH-E | F1 | F2 | F3 | F4 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1.4 | 226, 304 | 191 | MS2 [191]: 127, 173, 111, 85, 93 | Quinic acid | + | + | + | - | - |
| 2 | 1.8 | 224, 302 | 191 | MS2−[191]: 111, 173 | Citric acid | + | + | + | - | - |
| 3 | 3.9 | 275 | 255 | MS2 [255]: 165, 193 | Piscidic acid | + | + | + | - | - |
| 4 | 8.8 | 260, 330sh | 593 | MS2 [593]: 473, 311 | Genistein-C-hexoside-O-hexoside | 30.6 ± 1.45 b | - | 32.4 ± 3.65 a | 23.2 ± 1.44 c | - |
| 5 | 10.6 | 255, 315sh | 445 | MS2 [445]: 283; MS3 [283]: 268 | Calycosin-O-hexoside | 32.2 ± 2.34 a | - | 11.4 ± 0.67 c | 29.4 ± 0.13 b | - |
| 6 | 10.9 | 261, 330sh | 563 | MS2 [563]: 311, 283, 341, 431 | Genistein-C-hexoside-O-pentoside | 17.39 ± 2.33 c | 0.76 ± 0.34 d | 64.08 ± 0.12 a | 33.00 ± 0.16 b | - |
| 7 | 11.6 | 261, 330sh | 431 | MS2 [431]: 311 | Genistein-C-hexoside | 157.3 ± 1.15 c | - | 207.6 ± 2.21 b | 241.6 ± 0.76 a | - |
| 8 | 11.8 | 262, 330sh | 461 | MS2 [461]: 341, 371 | Tectorigenin- C-hexoside | + | - | + | + | - |
| 9 | 12.7 | 260, 321sh | 431 | MS2 [431]: 296, 268 | Genistin | 8.79 ± 0.77 c | - | 23.2 ± 0.47 a | 23.2 ± 0.21 a | - |
| 10 | 13.2 | 262, 328sh | 575 | MS2 [575]: 431, 311, 341 | Genistein-C-hexoside-3-hydroxy-3-methylglutaroyl | 7.19 ± 1.09 c | - | 28.2 ± 0.22 a | 18.6 ± 0.17 b | - |
| 11 | 15.6 | 271, 325 | 515 | MS2 [515]: 353, 395 | Ephedroidin-O-hexoside | + | - | - | + | + |
| 12 | 16.8 | 282 | 513 | MS2 [513]: 351, 485 | ND | + | - | - | + | + |
| 13 | 17.7 | 269, 300, 325 | 513 | MS2 [559]: 351 | Retamasin β-O-hexoside | + | - | - | + | + |
| 14 | 18 | 261, 320sh | 269 | MS2 [269]: 269 | Ginestein | + | - | - | + | + |
| 15 | 19.8 | 272, 326 | 353 | MS2 [353]: 283, 282, 335 | Ephedroidin | + | - | - | - | + |
| 16 | 20.7 | 281 | 353 | MS2 [353]: 283, 297 | ND | + | - | - | - | + |
| 17 | 20.8 | 259, 300, 323 | 351 | MS2 [351]: 333, 293 | Retamasin B | + | - | - | - | + |
| 18 | 21.2 | 277 | 497 | MS2 [497]: 335 | ND | + | - | - | - | + |
| 19 | 21.3 | 271, 320 | 337 | MS2 [337]: 281, 293 | Licoflavone C | tc | - | - | - | + |
| 20 | 21.9 | 272, 321 | 365 | ND | ND | + | - | - | - | + |
| 21 | 22.4 | 275 | 295 | ND | ND | + | - | - | - | + |
| Samples | DPPH• (IC50 µg/mL) | ORAC (µmol trolox/mg) | ROS in WS1 Cells (IC50 µg/mL) |
|---|---|---|---|
| MeOH-E | 37 ± 1.85 c | 6.61 ± 0.42 d | 6.0 ± 2.0 c |
| F1 | 280 ± 1.4 a | 1.13 ± 0.25 f | 20.0 ± 2.0 a |
| F2 | 35 ± 1.75 c | 4.74 ± 0.51 e | 6.2 ± 0.6 c |
| F3 | 22 ± 1.1 d | 8.10 ± 0.25 c | 4.5 ± 0.7 d |
| F4 | 105 ± 0.52 b | 1.36 ± 0.25 f | 10.0 ± 0.8 b |
| Trolox | - | - | 0.021 ± 0.002 f |
| Quercetin | - | 21.41 ± 3.49 b | 0.090 ± 0.004 f |
| Catechin | 26.87 ± 1.99 a | 0.20 ± 0.03 e | |
| BHT | 11.50 ± 0.57 e | - | - |
| IC50 (μg/mL) | |||
|---|---|---|---|
| Samples | A549 | DLD-1 | WS1 |
| MeOH-E | 24 ± 2 e | 17.5 ± 0.2 d | 153 ± 30 b |
| F 1 | 92 ± 13 b | >200 a | 143 ± 16 c |
| F 2 | >200 a | >200 a | >200 a |
| F 3 | 34 ± 2 d | 104 ± 8 c | 106 ± 8 e |
| F 4 | 57 ± 5 c | 123 ± 8 b | 115 ± 10 d |
| Etoposide | 18.2 ± 4.0 f | 4.6 ± 0.3 e | 15.3 ± 7.0 f |
| Diameter of the Inhibition Zone Including That of the Disk (mm ± SD) | |||||||
|---|---|---|---|---|---|---|---|
| Bacterial Strains | Source No. | F1 | F2 | F3 | F4 | MeOH-E | Gentamicin |
| Gram+ | |||||||
| Bacillus cereus | ATCC 14579 | - | - | - | 15.0 ± 0.6 b | 12.0 ± 0.6 c | 28.0 ± 1.4 a |
| Staphylococcus aureus | ATCC 25923 | - | - | 9.0 ± 0.0 d | 17.0 ± 0.5 b | 15 ± 1.0 c | 32.7 ± 1.6 a |
| Gram- | |||||||
| Escherichia coli | ATCC 25922 | - | - | - | - | 17.00 ± 0.50 a | 15.00 ± 0.75 b |
| Pseudomonas aeruginosa | ATCC 27853 | - | - | - | 12.0 ± 0.0 c | 16.0 ± 0.0 b | 30.33 ± 1.51 a |
| Salmonella typhimurium | ATCC 1408 | - | 10.7 ± 0.5 d | 11.7 ± 1.1 c | - | 14.0 ± 0.5 b | 21.00 ± 1.05 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saada, M.; Wasli, H.; Jallali, I.; Kboubi, R.; Girard-Lalancette, K.; Mshvildadze, V.; Ksouri, R.; Legault, J.; Cardoso, S.M. Bio-Guided Fractionation of Retama raetam (Forssk.) Webb & Berthel Polar Extracts. Molecules 2021, 26, 5800. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195800
Saada M, Wasli H, Jallali I, Kboubi R, Girard-Lalancette K, Mshvildadze V, Ksouri R, Legault J, Cardoso SM. Bio-Guided Fractionation of Retama raetam (Forssk.) Webb & Berthel Polar Extracts. Molecules. 2021; 26(19):5800. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195800
Chicago/Turabian StyleSaada, Mariem, Hanen Wasli, Inès Jallali, Rim Kboubi, Karl Girard-Lalancette, Vakhtang Mshvildadze, Riadh Ksouri, Jean Legault, and Susana M. Cardoso. 2021. "Bio-Guided Fractionation of Retama raetam (Forssk.) Webb & Berthel Polar Extracts" Molecules 26, no. 19: 5800. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195800