
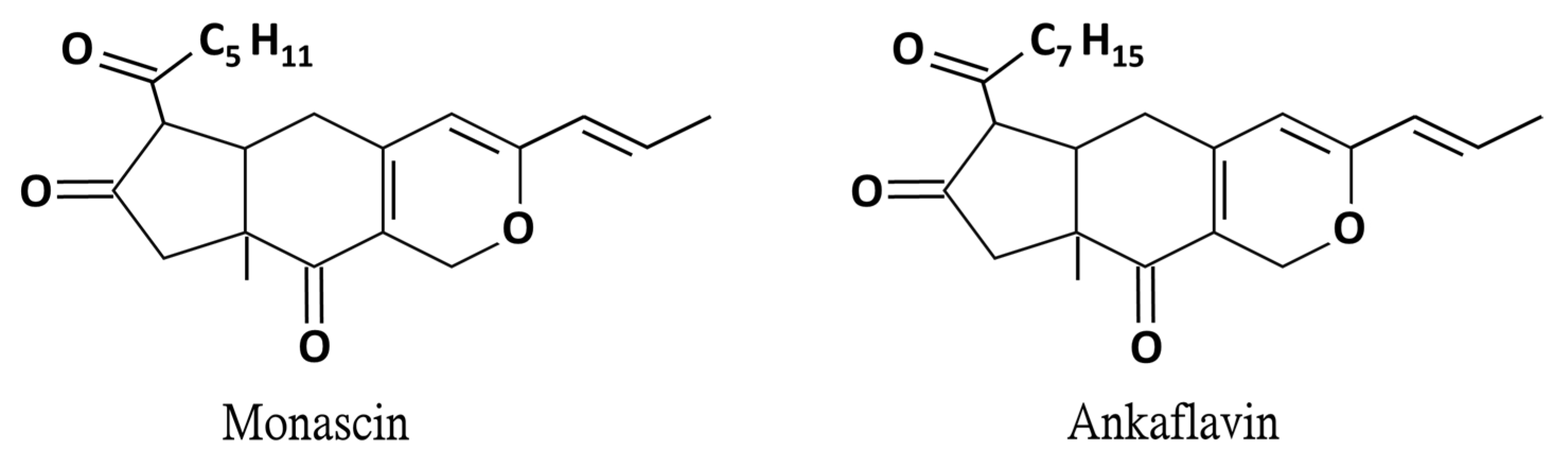
Monascin and Ankaflavin of Monascus purpureus Prevent Alcoholic Liver Disease through Regulating AMPK-Mediated Lipid Metabolism and Enhancing Both Anti-Inflammatory and Anti-Oxidative Systems



Abstract
:1. Introduction
2. Results
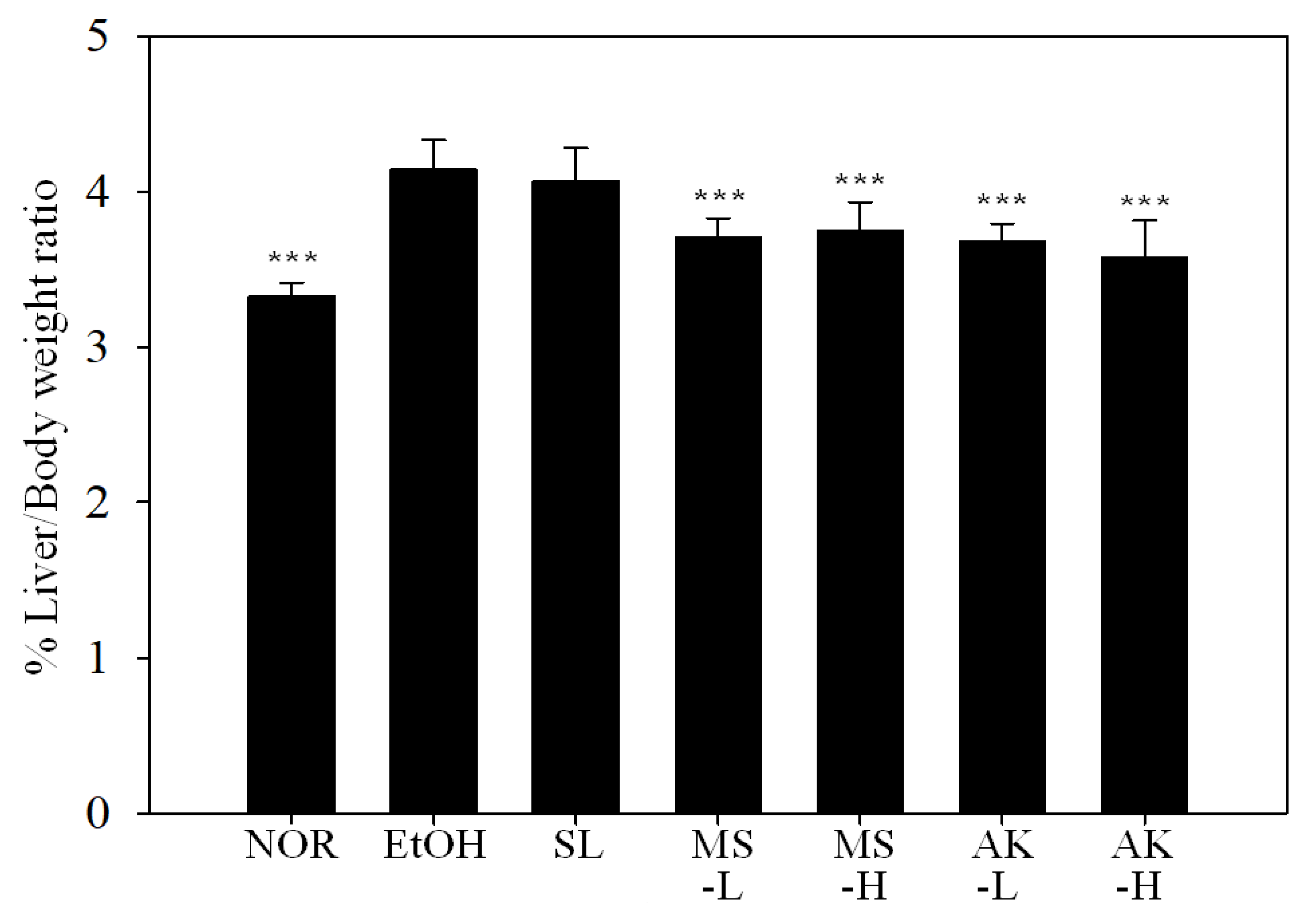
2.1. Effects of Monascin and Ankaflavin on Body Weight and Relative Liver to Body Weight Ratio in Ethanol-induced Liver Injury Mice
2.2. Effects of Monascin and Ankaflavin on Serum AST, ALT, and ALP Activities
2.3. Effects of Monascin and Ankaflavin on Serum and Hepatic TG and TC Contents
2.4. Effects of Monascin and Ankaflavin on Hepatic Pathological Changes
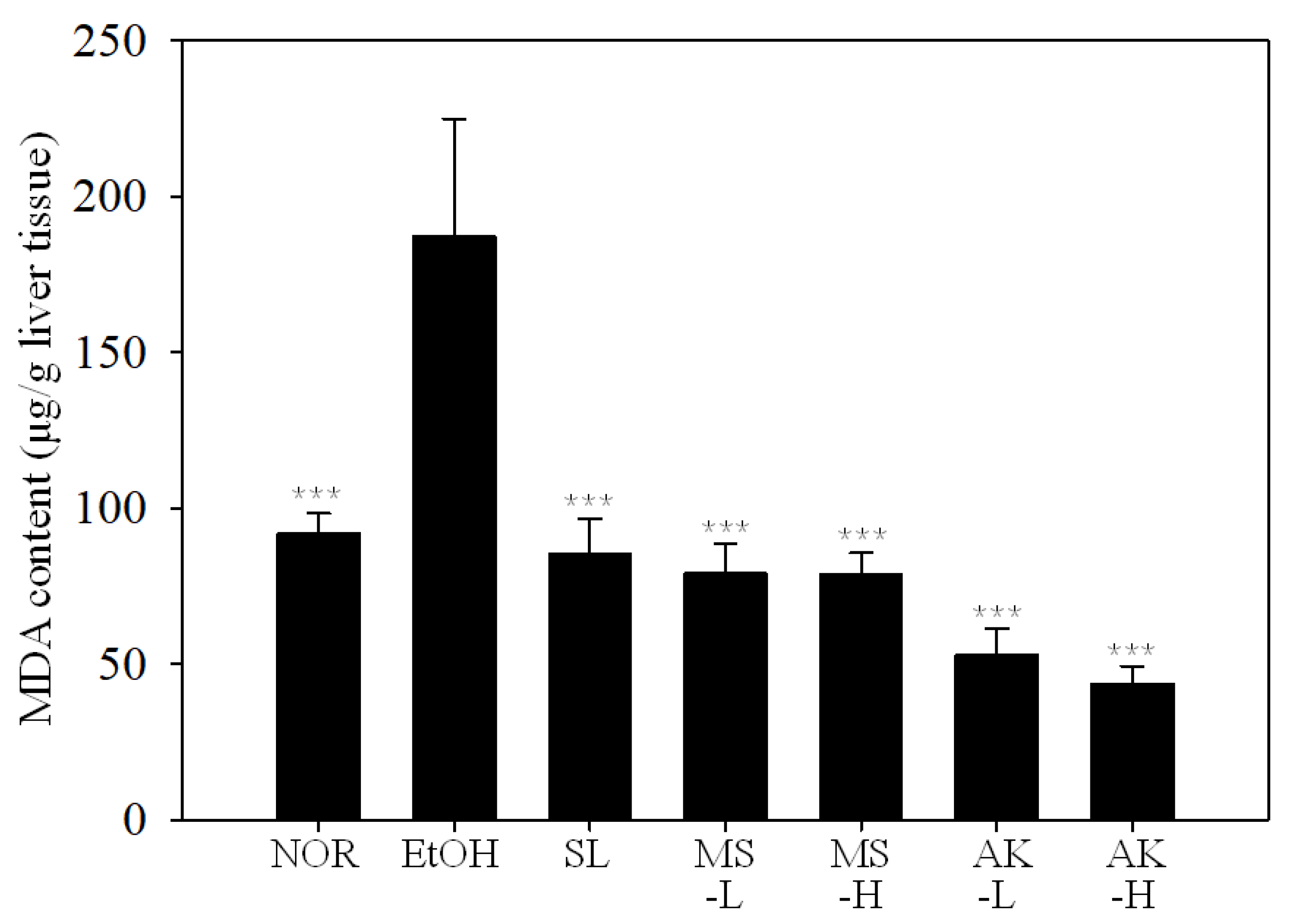
2.5. Effects of Monascin and Ankaflavin on the Lipid Peroxidation and Activities of Antioxidant Enzymes in the Liver
2.6. Effects of Monascin and Ankaflavin on the Regulation of Hepatic Inflammatory Factor Expression
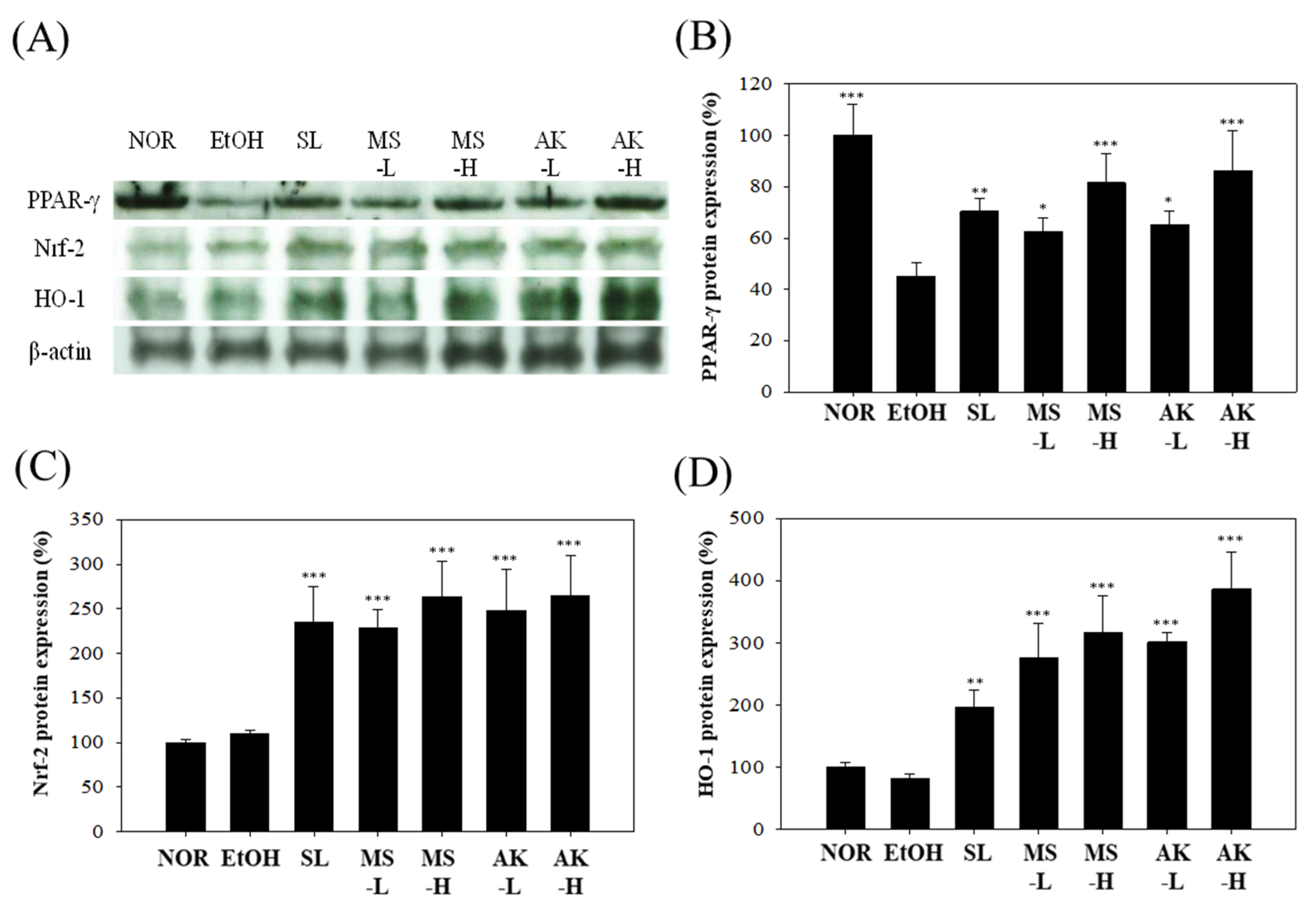
2.7. Effects of Monascin and Ankaflavin on The Regulation of Hepatic Anti-Inflammatory Factor Expression
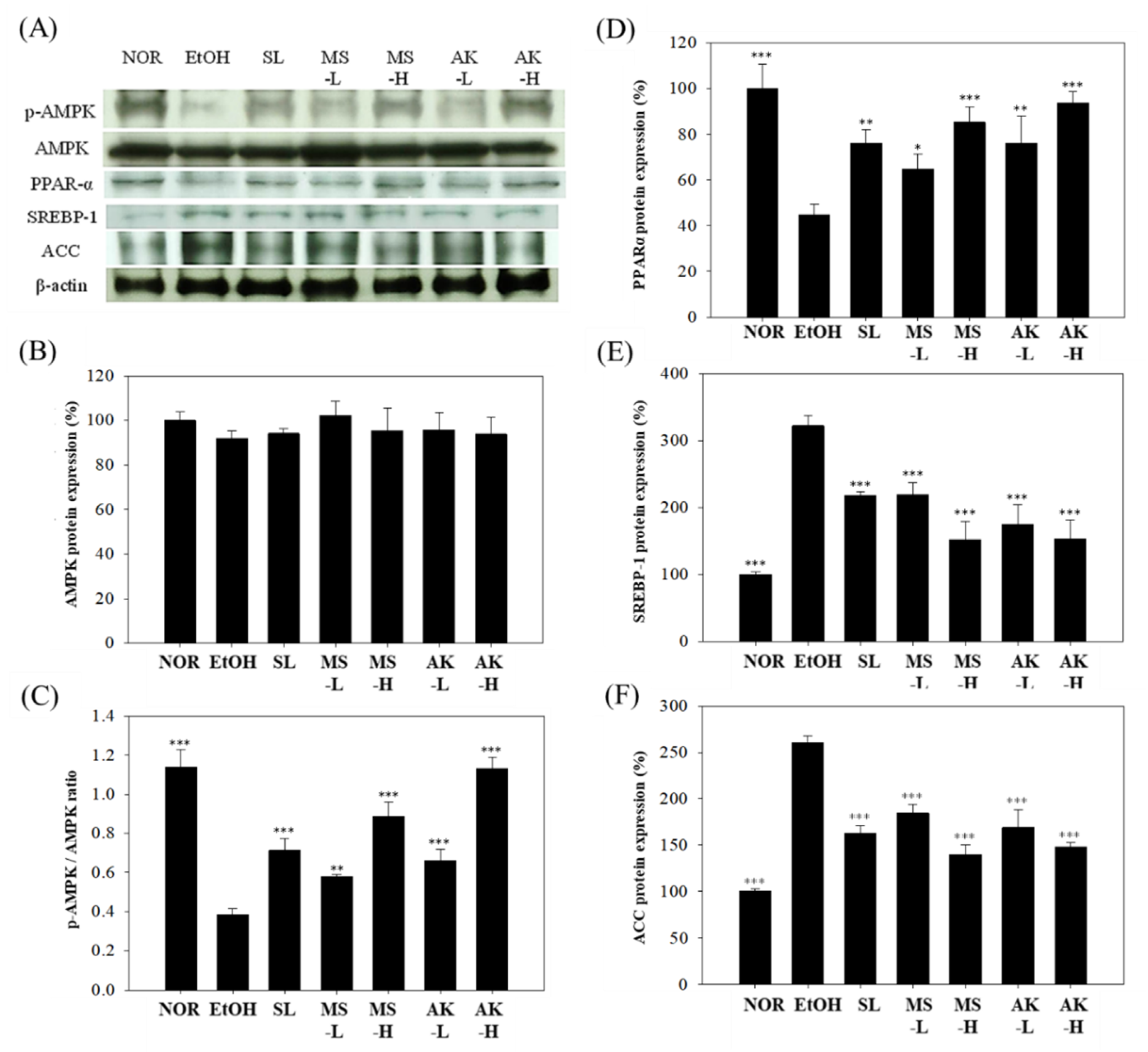
2.8. Effects of Monascin and Ankaflavin on The Regulation of Hepatic Lipid Metabolism Factor Expressions
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animal Models and Grouping
4.3. Serum Biochemistry Parameters
4.4. Liver Biochemistry Parameters
4.5. Immunoblotting
4.6. Histological Stains
4.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Malhi, H.; Gores, G.J. Cellular and molecular mechanisms of liver injury. Gastroenterology 2008, 134, 1641–1654. [Google Scholar] [CrossRef] [Green Version]
- Seitz, H.K.; Stickel, F. Molecular mechanisms of alcohol-mediated carcinogenesis. Nat. Rev. Cancer 2007, 7, 599–612. [Google Scholar] [CrossRef]
- Lumeng, L.; Crabb, D.W. Alcoholic liver disease. Curr. Opin. Gastroenterol. 2000, 16, 208–218. [Google Scholar] [CrossRef]
- Porta, E.A. Dietary modulation of oxidative stress in alcoholic liver disease in rats. J. Nutr. 1997, 127 (Suppl. 5), 912S–915S. [Google Scholar] [CrossRef] [Green Version]
- Lieber, C.S. Alcohol and the liver: Metabolism of ethanol, metabolic effects and pathogenesis of injury. Acta Med. Scandinavica. Suppl. 1985, 703, 11–55. [Google Scholar] [CrossRef]
- You, M.; Matsumoto, M.; Pacold, C.M.; Cho, W.K.; Crabb, D.W. The role of AMP-activated protein kinase in the action of ethanol in the liver. Gastroenterology 2004, 127, 1798–1808. [Google Scholar] [CrossRef]
- Sozio, M.; Crabb, D.W. Alcohol and lipid metabolism. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E10–E16. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.F.; Pan, T.M. Protective effect of Monascus-fermented red mold rice against alcoholic liver disease by attenuating oxidative stress and inflammatory response. J. Agric. Food Chem. 2011, 59, 9950–9957. [Google Scholar] [CrossRef]
- Lung, T.Y.; Liao, L.Y.; Wang, J.J.; Wei, B.L.; Huang, P.Y.; Lee, C.L. Metals of Deep Ocean Water Increase the Anti-Adipogenesis Effect of Monascus-Fermented Product via Modulating the Monascin and Ankaflavin Production. Mar. Drugs 2016, 14, 106. [Google Scholar] [CrossRef]
- Lee, C.L.; Wen, J.Y.; Hsu, Y.W.; Pan, T.M. Monascus-fermented yellow pigments monascin and ankaflavin showed antiobesity effect via the suppression of differentiation and lipogenesis in obese rats fed a high-fat diet. J. Agric. Food Chem. 2013, 61, 1493–1500. [Google Scholar] [CrossRef]
- Lee, C.L.; Hung, Y.P.; Hsu, Y.W.; Pan, T.M. Monascin and ankaflavin have more anti-atherosclerosis effect and less side effect involving increasing creatinine phosphokinase activity than monacolin K under the same dosages. J. Agric. Food Chem. 2013, 61, 143–150. [Google Scholar] [CrossRef]
- Hsu, L.C.; Hsu, Y.W.; Liang, Y.H.; Kuo, Y.H.; Pan, T.M. Anti-tumor and anti-inflammatory properties of ankaflavin and monaphilone A from Monascus purpureus NTU 568. J. Agric. Food Chem. 2011, 59, 1124–1130. [Google Scholar] [CrossRef]
- Hsu, W.H.; Lee, B.H.; Liao, T.H.; Hsu, Y.W.; Pan, T.M. Monascus-fermented metabolite monascin suppresses inflammation via PPAR-gamma regulation and JNK inactivation in THP-1 monocytes. Food Chem. Toxicol. 2012, 50, 1178–1186. [Google Scholar] [CrossRef]
- Cheng, C.F.; Pan, T.M. Monascus-fermented red mold dioscorea protects mice against alcohol-induced liver injury, whereas its metabolites ankaflavin and monascin regulate ethanol-induced peroxisome proliferator-activated receptor-gamma and sterol regulatory element-binding transcription factor-1 expression in HepG2 cells. J. Sci. Food Agric. 2018, 98, 1889–1898. [Google Scholar]
- Balasubramaniyan, V.; Kalaivani Sailaja, J.; Nalini, N. Role of leptin on alcohol-induced oxidative stress in Swiss mice. Pharmacol. Res. Off. J. Ital. Pharmacol. Soc. 2003, 47, 211–216. [Google Scholar] [CrossRef]
- Quertemont, E. Genetic polymorphism in ethanol metabolism: Acetaldehyde contribution to alcohol abuse and alcoholism. Mol. Psychiatry 2004, 9, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Altamirano, J.; Bataller, R. Alcoholic liver disease: Pathogenesis and new targets for therapy. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 491–501. [Google Scholar] [CrossRef]
- Zhao, C.Y.; Jiang, L.L.; Li, L.; Deng, Z.J.; Liang, B.L.; Li, J.M. Peroxisome proliferator activated receptor-gamma in pathogenesis of experimental fatty liver disease. World J. Gastroenterol. 2004, 10, 1329–1332. [Google Scholar] [CrossRef]
- Yan, D.; Dong, J.; Sulik, K.K.; Chen, S.Y. Induction of the Nrf2-driven antioxidant response by tert-butylhydroquinone prevents ethanol-induced apoptosis in cranial neural crest cells. Biochem. Pharmacol. 2010, 80, 144–149. [Google Scholar] [CrossRef] [Green Version]
- Lieber, C.S.; DeCarli, L.M. The feeding of alcohol in liquid diets: Two decades of applications and 1982 update. Alcohol. Clin. Exp. Res. 1982, 6, 523–531. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, H.H.; Hu, Y.Y. Effect of jianpi huoxue recipe on gut flora in rats with alcoholic fatty liver induced by Lieber-DeCarli liquid diet. Zhongguo Zhong Xi Yi Jie He Za Zhi Zhongguo Zhongxiyi Jiehe Zazhi = Chin. J. Integr. Tradit. West. Med./Zhongguo Zhong Xi Yi Jie He Xue HuiZhongguo Zhong Yi Yan Jiu Yuan Zhu Ban 2011, 31, 73–79. [Google Scholar]
- Lee, S.J.; Kim, S.Y.; Min, H. Effects of vitamin C and E supplementation on oxidative stress and liver toxicity in rats fed a low-fat ethanol diet. Nutr. Res. Pract. 2013, 7, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.J.; Koh, E.J.; Kim, C.S.; Zee, O.P.; Kwak, J.H.; Jeong, W.J.; Kim, J.H.; Lee, S.M. Agrimonia eupatoria protects against chronic ethanol-induced liver injury in rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2012, 50, 2335–2341. [Google Scholar] [CrossRef]
- Suter, P.M. Is alcohol consumption a risk factor for weight gain and obesity? Crit. Rev. Clin. Lab. Sci. 2005, 42, 197–227. [Google Scholar] [CrossRef]
- Lands, W.E.; Zakhari, S. The case of the missing calories. Am. J. Clin. Nutr. 1991, 54, 47–48. [Google Scholar] [CrossRef] [Green Version]
- Lieber, C.S. Pathogenesis and treatment of alcoholic liver disease: Progress over the last 50 years. Rocz. Akad. Med. W Bialymst. 2005, 50, 7–20. [Google Scholar]
- Cederbaum, A.I.; Lu, Y.; Wu, D. Role of oxidative stress in alcohol-induced liver injury. Arch. Toxicol. 2009, 83, 519–548. [Google Scholar] [CrossRef]
- You, M.; Crabb, D.W. Recent advances in alcoholic liver disease II. Minireview: Molecular mechanisms of alcoholic fatty liver. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G1–G6. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Yao, T.; Song, Z. Chronic alcohol consumption disrupted cholesterol homeostasis in rats: Down-regulation of low-density lipoprotein receptor and enhancement of cholesterol biosynthesis pathway in the liver. Alcohol. Clin. Exp. Res. 2010, 34, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Medina, J.; Moreno-Otero, R. Pathophysiological basis for antioxidant therapy in chronic liver disease. Drugs 2005, 65, 2445–2461. [Google Scholar] [CrossRef]
- Vitaglione, P.; Morisco, F.; Caporaso, N.; Fogliano, V. Dietary antioxidant compounds and liver health. Crit. Rev. Food Sci. Nutr. 2004, 44, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- McKim, S.E.; Gabele, E.; Isayama, F.; Lambert, J.C.; Tucker, L.M.; Wheeler, M.D.; Connor, H.D.; Mason, R.P.; Doll, M.A.; Hein, D.W.; et al. Inducible nitric oxide synthase is required in alcohol-induced liver injury: Studies with knockout mice. Gastroenterology 2003, 125, 1834–1844. [Google Scholar] [CrossRef]
- Mandrekar, P.; Szabo, G. Signalling pathways in alcohol-induced liver inflammation. J. Hepatol. 2009, 50, 1258–1266. [Google Scholar] [CrossRef] [Green Version]
- Nanji, A.A.; Miao, L.; Thomas, P.; Rahemtulla, A.; Khwaja, S.; Zhao, S.; Peters, D.; Tahan, S.R.; Dannenberg, A.J. Enhanced cyclooxygenase-2 gene expression in alcoholic liver disease in the rat. Gastroenterology 1997, 112, 943–951. [Google Scholar] [CrossRef]
- Zandi, E.; Rothwarf, D.M.; Delhase, M.; Hayakawa, M.; Karin, M. The IkappaB kinase complex (IKK) contains two kinase subunits, IKKalpha and IKKbeta, necessary for IkappaB phosphorylation and NF-kappaB activation. Cell 1997, 91, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Debril, M.B.; Renaud, J.P.; Fajas, L.; Auwerx, J. The pleiotropic functions of peroxisome proliferator-activated receptor gamma. J. Mol. Med. 2001, 79, 30–47. [Google Scholar] [CrossRef]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Farombi, E.O.; Shrotriya, S.; Na, H.K.; Kim, S.H.; Surh, Y.J. Curcumin attenuates dimethylnitrosamine-induced liver injury in rats through Nrf2-mediated induction of heme oxygenase-1. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2008, 46, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Maines, M.D.; Gibbs, P.E. 30 some years of heme oxygenase: From a "molecular wrecking ball" to a "mesmerizing" trigger of cellular events. Biochem. Biophys. Res. Commun. 2005, 338, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Andersen, D.B.; Holub, B.J. The influence of dietary inositol on glyceride composition and synthesis in livers of rats fed different fats. J. Nutr. 1976, 106, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Misra, P.; Chakrabarti, R. The role of AMP kinase in diabetes. Indian J. Med. Res. 2007, 125, 389–398. [Google Scholar]
- Alberts, A.W.; Strauss, A.W.; Hennessy, S.; Vagelos, P.R. Regulation of synthesis of hepatic fatty acid synthetase: Binding of fatty acid synthetase antibodies to polysomes. Proc. Natl. Acad. Sci. USA 1975, 72, 3956–3960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Villafranca, J.; Guillen, A.; Castro, J. Ethanol consumption impairs regulation of fatty acid metabolism by decreasing the activity of AMP-activated protein kinase in rat liver. Biochimie 2008, 90, 460–466. [Google Scholar] [CrossRef]
- Hsu, Y.W.; Hsu, L.C.; Liang, Y.H.; Kuo, Y.H.; Pan, T.M. Monaphilones A-C, three new antiproliferative azaphilone derivatives from Monascus purpureus NTU 568. J. Agric. Food Chem. 2010, 58, 8211–8216. [Google Scholar] [CrossRef] [PubMed]
- CDER. Guidance for Industry: Estimating the Maximum Safe. Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers; FDA: Silver Spring, MD, USA, 2005. [Google Scholar]
- Song, Z.; Deaciuc, I.; Song, M.; Lee, D.Y.; Liu, Y.; Ji, X.; McClain, C. Silymarin protects against acute ethanol-induced hepatotoxicity in mice. Alcohol Clin. Exp. Res. 2006, 30, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Abdella, N.; al Awadi, F.; Salman, A.; Armstrong, D. Thiobarbituric acid test as a measure of lipid peroxidation in Arab patients with NIDDM. Diabetes Res. 1990, 15, 173–177. [Google Scholar] [PubMed]
- Bihaqi, S.W.; Singh, A.P.; Tiwari, M. Supplementation of Convolvulus pluricaulis attenuates scopolamine-induced increased tau and amyloid precursor protein (AbetaPP) expression in rat brain. Indian J. Pharmacol. 2012, 44, 593–598. [Google Scholar] [CrossRef]
- Lee, C.L.; Kuo, T.F.; Wu, C.L.; Wang, J.J.; Pan, T.M. Red mold rice promotes neuroprotective sAPPalpha secretion instead of Alzheimer’s risk factors and amyloid beta expression in hyperlipidemic Abeta40-infused rats. J. Agric. Food Chem. 2010, 58, 2230–2238. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | AST (U/L) | ALT (U/L) | ALP (IU/L) |
|---|---|---|---|
| NOR | 44.9 ± 4.9 * | 17.3 ± 2.5 ** | 67.0 ± 3.1 ** |
| EtOH | 51.0 ± 9.8 | 22.1 ± 6.2 | 76.0 ± 5.5 |
| SL | 43.0 ± 4.7 ** | 18.5 ± 2.4 * | 73.3 ± 5.9 |
| MS-L | 41.6 ± 3.7 ** | 16.8 ± 2.7 ** | 70.6 ± 4.1 |
| MS-H | 39.5 ± 3.4 *** | 16.3 ± 1.9 ** | 68.6 ± 6.1 * |
| AK-L | 42.1 ± 4.3 ** | 17.0 ± 2.8 ** | 68.1 ± 4.3 ** |
| AK-H | 38.9 ± 3.2 *** | 16.8 ± 0.5 ** | 69.5 ± 6.7 * |
| Groups | Serum | Liver | ||
|---|---|---|---|---|
| TC (mg/dL) | TG (mg/dL) | TC (mg/g) | TG (mg/g) | |
| NOR | 65.62 ± 4.73 | 99.61 ± 10.39 ** | 3.41 ± 0.05 *** | 3.10 ± 0.56 *** |
| EtOH | 68.82 ± 10.40 | 119.61 ± 14.32 | 3.77 ± 0.16 | 4.86 ± 0.84 |
| SL | 63.74 ± 5.11 | 110.33 ± 8.78 * | 3.43 ± 0.14 *** | 3.01 ± 0.39 *** |
| MS-L | 58.35 ± 7.39 * | 94.85 ± 10.50 ** | 3.43 ± 0.11 *** | 2.95 ± 0.38 *** |
| MS-H | 60.72 ± 7.09 * | 88.45 ± 4.89 *** | 3.19 ± 0.08 *** | 2.52 ± 0.21 *** |
| AK-L | 59.32 ± 4.45 * | 106.48 ± 8.48 * | 3.22 ± 0.15 *** | 2.85 ± 0.53 *** |
| AK-H | 60.49 ± 5.72 * | 96.88 ± 9.56 ** | 3.10 ± 0.09 *** | 2.44 ± 0.18 *** |
| Groups | CAT Activity (U/mg Protein) | SOD Activity (U/mg Protein) | GPx Activity (U/mg Protein) | GRd Activity (U/mg Protein) |
|---|---|---|---|---|
| NOR | 0.54 ± 0.06 *** | 4.43 ± 0.65 *** | 807.02 ± 60.02 *** | 68.67 ± 6.02 *** |
| EtOH | 0.42 ± 0.07 | 2.59 ± 0.23 | 611.87 ± 53.06 | 57.76 ± 3.84 |
| SL | 0.43 ± 0.08 | 3.55 ± 0.52 ** | 666.49 ± 52.95 | 65.11 ± 4.25 ** |
| MS-L | 0.62 ± 0.03 *** | 3.12 ± 0.35 | 604.53 ± 64.00 | 63.31 ± 5.92 * |
| MS-H | 0.56 ± 0.06 *** | 3.60 ± 0.61 ** | 678.79 ± 52.78 * | 71.39 ± 4.82 *** |
| AK-L | 0.54 ± 0.04 *** | 3.36 ± 0.63 ** | 653.51 ± 37.76 | 66.05 ± 7.38 ** |
| AK-H | 0.58 ± 0.07 *** | 3.82 ± 0.68 *** | 685.66 ± 71.14 * | 73.18 ± 3.35 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, J.-R.; Hsu, Y.-W.; Pan, T.-M.; Lee, C.-L. Monascin and Ankaflavin of Monascus purpureus Prevent Alcoholic Liver Disease through Regulating AMPK-Mediated Lipid Metabolism and Enhancing Both Anti-Inflammatory and Anti-Oxidative Systems. Molecules 2021, 26, 6301. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26206301
Lai J-R, Hsu Y-W, Pan T-M, Lee C-L. Monascin and Ankaflavin of Monascus purpureus Prevent Alcoholic Liver Disease through Regulating AMPK-Mediated Lipid Metabolism and Enhancing Both Anti-Inflammatory and Anti-Oxidative Systems. Molecules. 2021; 26(20):6301. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26206301
Chicago/Turabian StyleLai, Jhao-Ru, Ya-Wen Hsu, Tzu-Ming Pan, and Chun-Lin Lee. 2021. "Monascin and Ankaflavin of Monascus purpureus Prevent Alcoholic Liver Disease through Regulating AMPK-Mediated Lipid Metabolism and Enhancing Both Anti-Inflammatory and Anti-Oxidative Systems" Molecules 26, no. 20: 6301. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26206301