
Roles of the 2-Oxoglutarate-Dependent Dioxygenase Superfamily in the Flavonoid Pathway: A Review of the Functional Diversity of F3H, FNS I, FLS, and LDOX/ANS

 ,
,
Abstract
:1. Introduction
2. Catalytic Mechanisms and Adversity Resistance of the Four 2-OGD Members
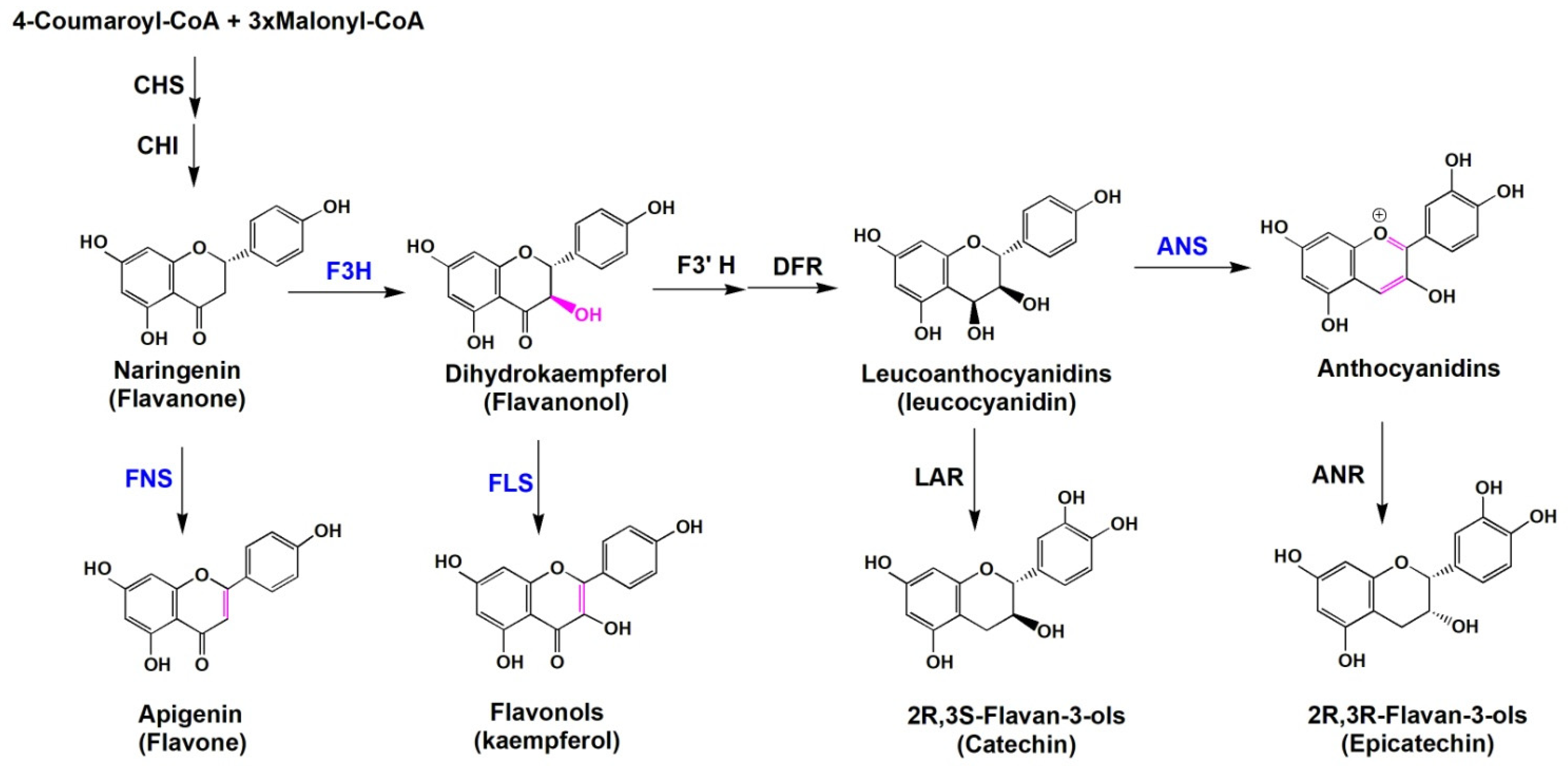
2.1. F3Hs
2.2. FNSsⅠ
2.3. FLSs
2.4. LDOXs/ANSs
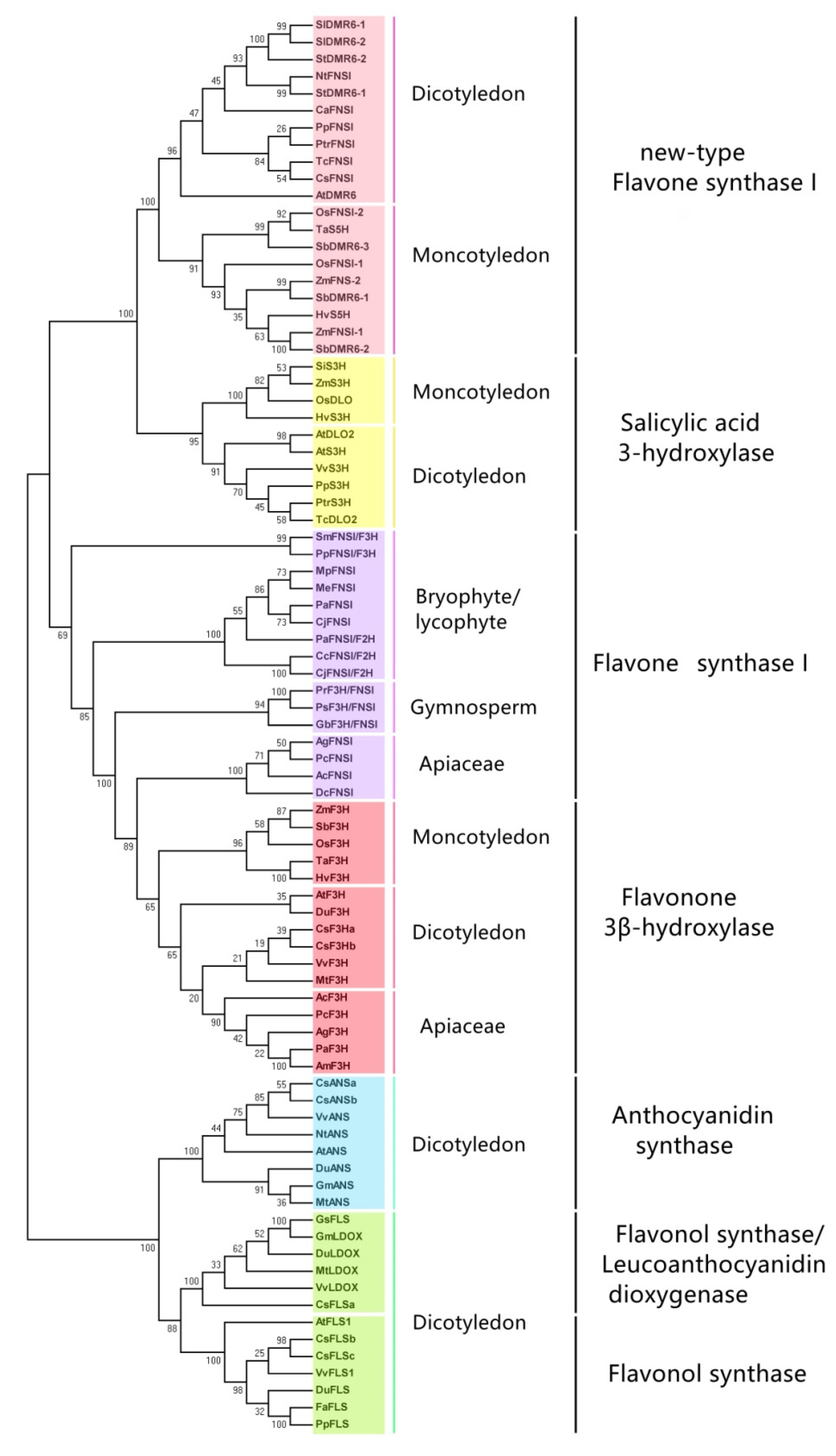
3. Phylogenetic Analysis of Plant 2-OGDs
4. Conclusions and Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
References
- Grotewold, E. The Science of Flavonoids; Springer: New York, NY, USA, 2006. [Google Scholar]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant. Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant. Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Paiva, N.L. Stress-lnduced phenylpropanoid metabolism. Plant. Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Flavonoidbiosynthesis, S.W. Acolorful model for genetics, biochemistry, cell biology, and biotechnology. Plant. Physiol. 2001, 126, 485–493. [Google Scholar]
- Dixon, R.A.; Sarnala, S. Proanthocyanidin Biosynthesis—A Matter of Protection. Plant Physiol. 2020, 184, 579–591. [Google Scholar] [CrossRef]
- Liao, Y.; Zhou, X.; Zeng, L. How does tea (Camellia sinensis) produce specialized metabolites which determine its unique quality and function: A review. Crit. Rev. Food Sci. Nutr. 2021, 1–17. [Google Scholar]
- Farrow, S.; Facchini, P. Functional diversity of 2-oxoglutarate/Fe(II)-dependent dioxygenases in plant metabolism. Front. Plant. Sci. 2014, 5, 524. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Yao, S.; Dai, X.; Yin, Q.; Liu, Y.; Jiang, X.; Wu, Y.; Qian, Y.; Pang, Y.; Gao, L.; et al. Identification of UDP-glycosyltransferases involved in the biosynthesis of astringent taste compounds in tea (Camellia sinensis). J. Exp. Botany 2016, 67, 2285–2297. [Google Scholar] [CrossRef] [Green Version]
- Stafford, H. Flavonoid evolution: An enzymic approach. Plant. Physiol. 1991, 96, 680–685. [Google Scholar] [CrossRef] [Green Version]
- Höfer, R.; Boachon, B.; Renault, H.; Gavira, C.; Miesch, L.; Iglesias, J.; Ginglinger, J.F.; Allouche, L.; Miesch, M.; Grec, S.; et al. Dual function of the cytochrome P450 CYP76 family from Arabidopsis thaliana in the metabolism of monoterpenols and phenylurea herbicides. Plant. Physiol. 2014, 166, 1149–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borg, A.; Beerens, K.; Pfeiffer, M.; Desmet, T.; Nidetzky, B. Stereo-electronic control of reaction selectivity in short-chain dehydrogenases: Decarboxylation, epimerization, and dehydration. Curr. Opin. Chem. Biol. 2021, 61, 43–52. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Emiliani, J.; Rodriguez, E.J.; Campos-Bermudez, V.A.; Grotewold, E.; Casati, P. The identification of maize and arabidopsis type i flavone synthases links flavones with hormones and biotic interactions. Plant. Physiol. 2015, 169, 1090–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravind, L.; Koonin, E. The DNA-repair protein AlkB, EGL-9, and leprecan define new families of 2-oxoglutarate- and iron-dependent dioxygenases. Genome Biol. 2001, 2, 1–8. [Google Scholar] [CrossRef]
- Martinez, S.; Hausinger, R. Catalytic Mechanisms of Fe(II)- and 2-Oxoglutarate-dependent Oxygenases. J. Biol. Chem. 2015, 290, 20702–20711. [Google Scholar] [CrossRef] [Green Version]
- Tarhonskaya, H.; Szöllössi, A.; Leung, I.K.; Bush, J.T.; Henry, L.; Chowdhury, R.; Iqbal, A.; Claridge, T.D.; Schofield, C.J.; Flashman, E. Studies on deacetoxycephalosporin C synthase support a consensus mechanism for 2-oxoglutarate dependent oxygenases. Biochemistry 2014, 53, 2483–2493. [Google Scholar] [CrossRef]
- Kawai, Y.; Ono, E.; Mizutani, M. Evolution and diversity of the 2-oxoglutarate-dependent dioxygenase superfamily in plants. Plant. J. Cell Mol. Biol. 2014, 78, 328–343. [Google Scholar] [CrossRef]
- Meza, T.J.; Moen, M.N.; Vågbø, C.B.; Krokan, H.E.; Klungland, A.; Grini, P.; Falnes, P. The DNA dioxygenase ALKBH2 protects Arabidopsis thaliana against methylation damage. Nucleic Acids Res. 2012, 40, 6620–6631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorres, K.; Raines, R. Prolyl 4-hydroxylase. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 106–124. [Google Scholar] [CrossRef]
- Martens, S.; Forkmann, G.; Matern, U.; Lukačin, R. Cloning of parsley flavone synthase I. Phytochemistry 2001, 58, 43–46. [Google Scholar] [CrossRef]
- Gebhardt, Y.; Witte, S.; Forkmann, G.; Lukačin, R.; Matern, U.; Martens, S. Molecular evolution of flavonoid dioxygenases in the family Apiaceae. Phytochemistry 2005, 66, 1273–1284. [Google Scholar] [CrossRef]
- Martens, S.; Forkmann, G.; Britsch, L.; Wellmann, F.; Matern, U.; Lukačin, R. Divergent evolution of flavonoid 2-oxoglutarate-dependent dioxygenases in parsley1. FEBS Lett. 2003, 544, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Britsch, L.; Ruhnau-Brich, B.; Forkmann, G. Molecular cloning, sequence analysis, and in vitro expression of flavanone 3 beta-hydroxylase from Petunia hybrida. J. Biol. Chem. 1992, 267, 5380–5387. [Google Scholar] [CrossRef]
- Owens, D.K.; Crosby, K.C.; Runac, J.; Howard, B.A.; Winkel, B.S. Biochemical and genetic characterization of Arabidopsis flavanone 3β-hydroxylase. Plant. Physiol. Biochem. 2008, 46, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Owens, D.K.; Alerding, A.B.; Crosby, K.C.; Bandara, A.B.; Westwood, J.H.; Winkel, B.S. Functional analysis of a predicted flavonol synthase gene family in Arabidopsis. Plant. Physiol. 2008, 147, 1046–1061. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.Y.; Sharma, S.B.; Paiva, N.L.; Ferreira, D.; Dixon, R.A. Role of anthocyanidin reductase, encoded by BANYULS in plant flavonoid biosynthesis. Science 2003, 299, 396–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, J.-I.; Tanaka, Y.; Yamazaki, M.; Saito, K. Reaction Mechanism from Leucoanthocyanidin to Anthocyanidin 3-Glucoside, a Key Reaction for Coloring in Anthocyanin Biosynthesis. J. Biol. Chem. 2001, 276, 25797–25803. [Google Scholar] [CrossRef] [Green Version]
- Forkmann, G.; Heller, W.; Grisebach, H. Anthocyanin Biosynthesis in Flowers of Matthiolaincana Flavanone 3-and Flavonoid 3′-Hydroxylases. Zeitschrift für Naturforschung C 1980, 35, 691–695. [Google Scholar] [CrossRef]
- Britsch, L.; Heller, W.; Grisebach, H. Conversion of Flavanone to Flavone, Dihydroflavonol and Flavonol with an Enzyme System from Cell Cultures of Parsley. Zeitschrift für Naturforschung 1981, 36, 742–750. [Google Scholar] [CrossRef]
- Froemel, S.; De Vlaming, P.; Stotz, G.; Wiering, H.; Forkmann, G.; Schram, A.W. Genetic and biochemical studies on the conversion of flavanones to dihydroflavonols in flowers of Petunia hybrida. Theor. Appl. Genet. 1985, 70, 561–568. [Google Scholar] [CrossRef]
- Forkmann, G.; Stotz, G. Selection and characterisation of flavanone 3-hydroxylase mutants ofDahlia, Streptocarpus, Verbena andZinnia. Planta 1984, 161, 261–265. [Google Scholar] [CrossRef]
- Lukačin, R.; Britsch, L. Identification of Strictly Conserved Histidine and Arginine Residues as Part of the Active Site in Petunia hybrida Flavanone 3β-Hydroxylase. Eur. J. Biochem. 1997, 249, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Lukačin, R.; Gröning, I.; Pieper, U.; Matern, U. Site-directed mutagenesis of the active site serine290 in flavanone 3beta-hydroxylase from Petunia hybrida. Eur. J. Biochem. 2000, 267, 853–860. [Google Scholar] [CrossRef]
- Wisman, E.; Hartmann, U.; Sagasser, M.; Baumann, E.; Palme, K.; Hahlbrock, K.; Saedler, H.; Weisshaar, B. Knock-out mutants from an En-1 mutagenized Arabidopsis thaliana population generate phenylpropanoid biosynthesis phenotypes. Proc. Natl. Acad. Sci. USA 1998, 95, 12432–12437. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Rani, A.; Kumar, S.; Sood, P.; Mahajan, M.; Yadav, S.K.; Singh, B.; Ahuja, P.S. An early gene of the flavonoid pathway, flavanone 3-hydroxylase, exhibits a positive relationship with the concentration of catechins in tea (Camellia sinensis). Tree Physiol. 2008, 28, 1349–1356. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Li, X.; Liu, Y.; Cao, B. Regulation of flavanone 3-hydroxylase gene involved in the flavonoid biosynthesis pathway in response to UV-B radiation and drought stress in the desert plant, Reaumuriasoongorica. Plant. Physiol. Biochem. 2013, 73, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, M.; Yadav, S.K. Overexpression of a tea flavanone 3-hydroxylase gene confers tolerance to salt stress and Alternaria solani in transgenic tobacco. Plant. Mol. Biol. 2014, 85, 551–573. [Google Scholar] [CrossRef]
- Li, C.; Liu, S.; Yao, X.; Wang, J.; Wang, T.; Zhang, Z.; Zhang, P.; Chen, K. PnF3H, a flavanone 3-hydroxylase from the Antarctic moss Pohlia nutans, confers tolerance to salt stress and ABA treatment in transgenic Arabidopsis. Plant. Growth Regul. 2017, 83, 489–500. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant. Biol. 2016, 16, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhao, L.; Wang, J.; Zheng, L.; Dang, Z.; Wang, Y. Cloning and functional analysis of two flavanone-3-hydroxylase genes from Reaumuriatrigyna. Acta Physiol. Plant. 2014, 36, 1221–1229. [Google Scholar] [CrossRef]
- Han, Y.; Huang, K.; Liu, Y.; Jiao, T.; Ma, G.; Qian, Y.; Wang, P.; Dai, X.; Gao, L.; Xia, T. Functional Analysis of Two Flavanone-3-Hydroxylase Genes from Camellia sinensis: A Critical Role in Flavonoid Accumulation. Genes 2017, 8, 300. [Google Scholar] [CrossRef] [Green Version]
- Li, D.-D.; Ni, R.; Wang, P.-P.; Zhang, X.-S.; Wang, P.-Y.; Zhu, T.-T.; Sun, C.-J.; Liu, C.-J.; Lou, H.-X.; Cheng, A.-X. Molecular Basis for Chemical Evolution of Flavones to Flavonols and Anthocyanins in Land Plants. Plant. Physiol. 2020, 184, 1731–1743. [Google Scholar] [CrossRef]
- Asakawa, Y.; Ludwiczuk, A.; Nagashima, F. Chemical constituents of bryophytes. Bio- and chemical diversity, biological activity, and chemosystematics. Progress in the chemistry of organic natural products. Prog. Chem. Org. Nat. Prod. 2013, 95, 1–796. [Google Scholar]
- Wang, L.; Wang, L.N.; Zhao, Y.; Lou, H.X.; Cheng, A.X. Secondary metabolites from Marchantia paleacea calluses and their allelopathic effects on Arabidopsis seed growth. Nat. Prod.Res. 2013, 27, 274–276. [Google Scholar] [CrossRef]
- Kim, J.H.; Cheon, Y.M.; Kim, B.G.; Ahn, J.H. Analysis of flavonoids and characterization of theOsFNS gene involved in flavone biosynthesis in Rice. J. Plant. Biol. 2008, 51, 97–101. [Google Scholar] [CrossRef]
- Sutter, A.; Poulton, J.; Grisebach, H. Oxidation of flavanone to flavone with cell-free extracts from young parsley leaves. Arch. Biochem. Biophys. 1975, 170, 547–556. [Google Scholar] [CrossRef]
- Britsch, L. Purification and characterization of flavone synthase I, a 2-oxoglutarate-dependent desaturase. Arch. Biochem. Biophys. 1990, 282, 152–160. [Google Scholar] [CrossRef]
- Han, X.-J.; Wu, Y.-F.; Gao, S.; Yu, H.-N.; Xu, R.-X.; Lou, H.-X.; Cheng, A.-X. Functional characterization of aPlagiochasmaappendiculatumflavone synthase I showing flavanone 2-hydroxylase activity. FEBS Lett. 2014, 588, 2307–2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, S.; Forkmann, G. Genetic control of flavone synthase II activity in flowers of Gerbera hybrids. Phytochemistry 1998, 49, 1953–1958. [Google Scholar] [CrossRef]
- Righini, S.; Rodriguez, E.J.; Berosich, C.; Grotewold, E.; Casati, P.; Ferreyra, M.L.F. Apigenin produced by maize flavone synthase I and II protects plants against UV-B-induced damage. Plant. Cell Environ. 2018, 42, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Martens, S. Mithöfer, A. Flavones and flavone synthases. Phytochemistry 2005, 66, 2399–2407. [Google Scholar] [CrossRef]
- Zhu, Y.-X.; Ge, C.; Ma, S.; Liu, X.-Y.; Liu, M.; Sun, Y.; Wang, G.-F. Maize ZmFNSI Homologs Interact with an NLR Protein to Modulate Hypersensitive Response. Int. J. Mol. Sci. 2020, 21, 2529. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhao, L.; Zhao, J.; Li, Y.; Wang, J.; Guo, R.; Gan, S.; Liu, C.J.; Zhang, K. S5H/DMR6 Encodes a Salicylic Acid 5-Hydroxylase That Fine-Tunes Salicylic Acid Homeostasis. Plant. Physiol. 2017, 175, 1082–1093. [Google Scholar] [CrossRef] [Green Version]
- Holton, T.; Brugliera, F.; Tanaka, Y. Cloning and expression of flavonol synthase from Petunia hybrida. Plant. J. 1993, 4, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Shi, Y.; Fu, Z.; Li, W.-W.; Lai, S.; Wu, Y.; Wang, Y.; Liu, Y.; Gao, L.; Xia, T. Functional characterization of three flavonol synthase genes from Camellia sinensis: Roles in flavonol accumulation. Plant. Sci. 2020, 300, 110632. [Google Scholar] [CrossRef]
- Lukačin, R.; Wellmann, F.; Britsch, L.; Martens, S.; Matern, U. Flavonol synthase from Citrus unshiu is a bifunctional dioxygenase. Phytochemistry 2003, 62, 287–292. [Google Scholar] [CrossRef]
- Prescott, A.G.; Stamford, N.P.; Wheeler, G.; Firmin, J.L. In Vitro properties of a recombinant flavonol synthase from Arabidopsis thaliana. Phytochemistry 2002, 60, 589–593. [Google Scholar] [CrossRef]
- Turnbull, J.J.; Nakajima, J.I.; Welford, R.W.; Yamazaki, M.; Saito, K.; Schofield, C.J. Mechanistic studies on three 2-oxoglutarate-dependent oxygenases of flavonoid biosynthesis: Anthocyanidin synthase, flavonol synthase, and flavanone 3beta-hydroxylase. J. Biol. Chem. 2004, 279, 1206–1216. [Google Scholar] [CrossRef] [Green Version]
- Abrahams, S.; Lee, E.; Walker, A.; Tanner, G.J.; Larkin, P.J.; Ashton, A.R. TheArabidopsis TDS4gene encodes leucoanthocyanidin dioxygenase (LDOX) and is essential for proanthocyanidin synthesis and vacuole development. Plant. J. 2003, 35, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, J.J.; Nagle, M.J.; Seibel, J.F.; Welford, R.W.; Grant, G.H.; Schofield, C.J. The C-4 stereochemistry of leucocyanidin substrates for anthocyanidin synthase affects product selectivity. Bioorganic Med. Chem. Lett. 2003, 13, 3853–3857. [Google Scholar] [CrossRef]
- Turnbull, J.J.; Sobey, W.J.; Aplin, R.T.; Hassan, A.; Firmin, J.L.; Schofield, C.J.; Prescott, A.G. Are anthocyanidins the immediate products of anthocyanidin synthase? Chem. Commun. 2000, 24, 2473–2474. [Google Scholar] [CrossRef]
- Welford, R.W.; Turnbull, J.J.; Claridge, T.D.; Prescott, A.G.; Schofield, C.J. Evidence for oxidation at C-3 of the flavonoid C-ring during anthocyanin biosynthesis. Chem. Commun. 2001, 18, 1828–1829. [Google Scholar] [CrossRef] [PubMed]
- Wellmann, F.; Griesser, M.; Schwab, W.; Martens, S.; Eisenreich, W.; Matern, U.; Lukačin, R. Anthocyanidin synthase from Gerbera hybrida catalyzes the conversion of (+)-catechin to cyanidin and a novel procyanidin. FEBS Lett. 2006, 580, 1642–1648. [Google Scholar] [CrossRef] [Green Version]
- Jun, J.H.; Xiao, X.; Rao, X.; Dixon, R.A. Proanthocyanidin subunit composition determined by functionally diverged dioxygenases. Nat. Plants 2018, 4, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, M.; Andel, A.; Huibers, R.P.; Panstruga, R.; Weisbeek, P.J.; Van den Ackerveken, G. Identification of arabidopsis loci required for susceptibility to the downy mildew pathogen Hyaloperonosporaparasitica. Mol. Plant. Microbe Interact. 2005, 18, 583–592. [Google Scholar] [CrossRef] [Green Version]
- Van Damme, M.; Huibers, R.P.; Elberse, J.; Van den Ackerveken, G. Arabidopsis DMR6 encodes a putative 2OG-Fe(II) oxygenase that is defense-associated but required for susceptibility to downy mildew. Plant. J. 2008, 54, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Thomazella, D.P.D.T.; Seong, K.; Mackelprang, R.; Dahlbeck, D.; Geng, Y.; Gill, U.S.; Qi, T.; Pham, J.; Giuseppe, P.; Lee, C.Y.; et al. Loss of function of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Fliegmann, J.; Furtwängler, K.; Malterer, G.; Cantarello, C.; Schüler, G.; Ebel, J.; Mithöfer, A. Flavone synthase II (CYP93B16) from soybean (Glycine max L.). Phytochemistry 2010, 71, 508–514. [Google Scholar] [CrossRef]
- Du, Y.; Chu, H.; Wang, M.; Chu, I.K.; Lo, C. Identification of flavone phytoalexins and a pathogen-inducible flavone synthase II gene (SbFNSII) in sorghum. J. Exp. Bot. 2009, 61, 983–994. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Zhang, Y.; Wang, G.; Hill, L.; Weng, J.-K.; Chen, X.-Y.; Xue, H.; Martin, C. A specialized flavone biosynthetic pathway has evolved in the medicinal plant, Scutellariabaicalensis. Sci. Adv. 2016, 2, e1501780. [Google Scholar] [CrossRef]
- Markham, K.R. Distribution of flavonoids in the lower plants and its evolutionary significance. In The Flavonoids; Springer: Boston, MA, USA, 1988; pp. 427–468. [Google Scholar]
- Pelletier, M.K.; Murrell, J.R.; Shirley, B.W. Characterization of Flavonol Synthase and Leucoanthocyanidin Dioxygenase Genes in Arabidopsis (Further Evidence for Differential Regulation of “Early” and “Late” Genes). Plant. Physiol. 1997, 113, 1437–1445. [Google Scholar] [CrossRef] [Green Version]
- Wellmann, F.; Lukačin, R.; Moriguchi, T.; Britsch, L.; Schiltz, E.; Matern, U. Functional expression and mutational analysis of flavonol synthase fromCitrusunshiu. JBIC J. Biol. Inorg. Chem. 2002, 269, 4134–4142. [Google Scholar] [CrossRef]
- Ferreyra, M.L.F.; Rius, S.; Emiliani, J.; Pourcel, L.; Feller, A.; Morohashi, K.; Casati, P.; Grotewold, E. Cloning and characterization of a UV-B-inducible maize flavonol synthase. Plant. J. 2010, 62, 77–91. [Google Scholar] [CrossRef]
- Vu, T.T.; Jeong, C.Y.; Nguyen, H.N.; Lee, D.; Lee, S.A.; Kim, J.H.; Hong, S.-W.; Lee, H. Characterization of Brassica napus Flavonol Synthase Involved in Flavonol Biosynthesis in Brassica napus L. J. Agric. Food Chem. 2015, 63, 7819–7829. [Google Scholar] [CrossRef]
- Nielsen, K.; Deroles, S.C.; Markham, K.R.; Bradley, M.J.; Podivinsky, E.; Manson, D. Antisense flavonol synthase alters copigmentation and flower color in lisianthus. Mol. Breed. 2002, 9, 217–229. [Google Scholar] [CrossRef]
- Tian, J.; Han, Z.-Y.; Zhang, J.; Hu, Y.; Song, T.; Yao, Y. The Balance of Expression of Dihydroflavonol 4-reductase and Flavonol Synthase Regulates Flavonoid Biosynthesis and Red Foliage Coloration in Crabapples. Sci. Rep. 2015, 5, 12228. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Ning, G.; Wang, Z.; Shen, Y.; Jin, H.; Li, P.; Huang, S.; Zhao, J.; Bao, M. Disequilibrium of flavonol synthase and dihydroflavonol-4-reductase expression associated tightly to white vs. red color flower formation in plants. Front. Plant. Sci. 2015, 6, 1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Kim, D.H.; Park, B.R.; Lee, J.Y.; Lim, S.H. Molecular and functional characterization of Oryza sativa FLAVONOL SYNTHASE (OsFLS), a bifunctional dioxygenase. J. Agric. Food Chem. 2019, 26, 7399–7409. [Google Scholar] [CrossRef]
- Saito, K.; Kobayashi, M.; Gong, Z.; Tanaka, Y.; Yamazaki, M. Direct evidence for anthocyanidin synthase as a 2-oxoglutarate-dependent oxygenase: Molecular cloning and functional expression of cDNA from a red forma of Perilla frutescens. Plant J. 1999, 17, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Wilmouth, R.C.; Turnbull, J.J.; Welford, R.W.; Clifton, I.J.; Prescott, A.G.; Schofield, C.J. Structure and mechanism of anthocyanidin synthase from arabidopsis thaliana. Structure 2002, 10, 93–103. [Google Scholar] [CrossRef]
- Cheng, A.-X.; Han, X.-J.; Wu, Y.-F.; Lou, H.-X. The Function and Catalysis of 2-Oxoglutarate-Dependent Oxygenases Involved in Plant Flavonoid Biosynthesis. Int. J. Mol. Sci. 2014, 15, 1080–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebhardt, Y.H.; Witte, S.; Steuber, H.; Matern, U.; Martens, S. Evolution of Flavone Synthase I from Parsley Flavanone 3β-Hydroxylase by Site-Directed Mutagenesis. Plant. Physiol. 2007, 144, 1442–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Halitschke, R.; Yin, C.; Liu, C.J.; Gan, S.S. Salicylic acid 3-hydroxylase regulates Arabidopsis leaf longevity by mediating salicylic acid catabolism. Proc. Natl. Acad. Sci. USA 2013, 110, 14807–14812. [Google Scholar] [CrossRef] [Green Version]
- Zeilmaker, T.; Ludwig, N.R.; Elberse, J.; Seidl, M.F.; Berke, L.; Van Doorn, A.; Schuurink, R.C.; Snel, B.; Ackerveken, G.V.D. DOWNY MILDEW RESISTANT 6 and DMR6-LIKE OXYGENASE 1 are partially redundant but distinct suppressors of immunity in Arabidopsis. Plant. J. 2014, 81, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.; Lewis, N.E.; Lerman, J.A.; Lee, D.-H.; Chang, R.L.; Kim, D.; Palsson, B.O. Network Context and Selection in the Evolution to Enzyme Specificity. Science 2012, 337, 1101–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]




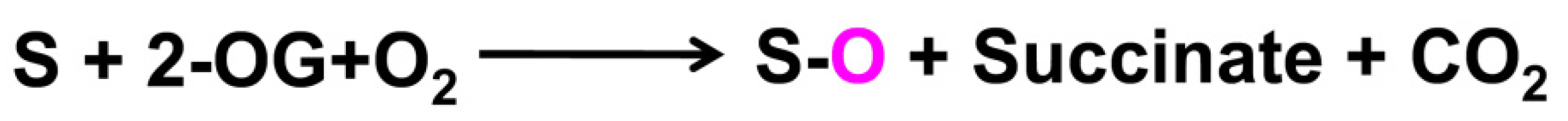{kind=link}
{kind=link}
{kind=link}
| The Members of 2-OGDs | Substrates | Corresponding Product | Verified in Vivo or in Vitro | References |
|---|---|---|---|---|
| F3H | (2S)-Flavanone | (2R, 3R)-Dihydroflavonols | in vivo and in vitro | [23,31] |
| FNS | (2S)-Flavanone | Flavones | in vivo and in vitro | [20,47] |
| (2R, 3S)-cis-DHK | Kaempferol | in vitro | [22] | |
| Salicylic Acid | 2,5-DHBA | in vivo and in vitro | [53] | |
| FLS | (2R,3R)-trans-Dihydroflavonols | Flavonols | in vivo and in vitro | [54,55] |
| (2S)-Naringenin | (2R,3S)-cis-DHK, (2R,3R)-trans-DHK, Kaempferol and Apigenin | in vitro | [56,57] | |
| (2R)-Naringenin | (2S,3S)-trans-DHK | in vitro | [56] | |
| (2R,3S,4R)-leucocyanidin | DHQ, Q and Cyanidin | in vitro | [58] | |
| ANS/LDOX | (2R,3S,4S)-leucocyanidin | (4S)-flav-2-en-3,4-diol, Cyanidin | in vivo | [27,59] |
| (2R,3S,4S)-leucocyanidin | Q (85%), DHQ, and Cyanidin | in vitro | [60,61] | |
| (2R,3S,4R)-leucocyanidin | DHQ (66%), Q (30%) and Cyanidin | in vitro | [60] | |
| (2R,3R)-trans-Dihydroflavonols | Flavonols | in vitro | [61] | |
| (2S)-Naringenin | (2R,3S)-cis-DHK, (2R,3R)-trans-DHK, Kaempferol and Apigenin | in vitro | [58] | |
| (2R)-Naringenin | (2S,3S)-trans-DHK | in vitro | [62] | |
| (+)-Catechin | Cyanidin | in vitro | [63] | |
| New-type LDOX | (2R,3S,4S)-leucocyanidin | Cyanidin and Q | in vitro | [64] |
| (+)-Catechin | Cyanidin | in vivo and in vitro | [64] | |
| (2R,3R)-trans-Dihydroflavonols | Flavonols | in vitro | [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Shi, Y.; Li, K.; Yang, D.; Liu, N.; Zhang, L.; Zhao, L.; Zhang, X.; Liu, Y.; Gao, L.; et al. Roles of the 2-Oxoglutarate-Dependent Dioxygenase Superfamily in the Flavonoid Pathway: A Review of the Functional Diversity of F3H, FNS I, FLS, and LDOX/ANS. Molecules 2021, 26, 6745. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216745
Wang Y, Shi Y, Li K, Yang D, Liu N, Zhang L, Zhao L, Zhang X, Liu Y, Gao L, et al. Roles of the 2-Oxoglutarate-Dependent Dioxygenase Superfamily in the Flavonoid Pathway: A Review of the Functional Diversity of F3H, FNS I, FLS, and LDOX/ANS. Molecules. 2021; 26(21):6745. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216745
Chicago/Turabian StyleWang, Yueyue, Yufeng Shi, Kaiyuan Li, Dong Yang, Nana Liu, Lingjie Zhang, Lei Zhao, Xinfu Zhang, Yajun Liu, Liping Gao, and et al. 2021. "Roles of the 2-Oxoglutarate-Dependent Dioxygenase Superfamily in the Flavonoid Pathway: A Review of the Functional Diversity of F3H, FNS I, FLS, and LDOX/ANS" Molecules 26, no. 21: 6745. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216745