
Attenuative Effects of Fluoxetine and Triticum aestivum against Aluminum-Induced Alzheimer’s Disease in Rats: The Possible Consequences on Hepatotoxicity and Nephrotoxicity

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Behavioral Test (Morris Water Maze (MWM)
2.2. Effect of Fluoxetine, Wheatgrass, or Their Combination on Alanine Transaminase (ALT), Aspartate Transaminase (AST), and Alkaline Phosphatase (ALP)
2.3. Effect of Fluoxetine, Wheatgrass, or Their Combination on Total Cholesterol (TC), Triacylglycerol (TG), and High-Density Lipoprotein (HDL)
2.4. Effect of Fluoxetine, Wheatgrass, or Their Combination on Hepatic Interleukin-6 (IL-6), Tumor Necrosis Factor-A (TNF-A), Nuclear Factor Kappa B (NF-Κb), and Caspase-3 Activity
2.5. Effect of Fluoxetine, Wheatgrass, or Their Combination on Hepatic Total Antioxidant Capacity (TAC), Superoxide Dismutase (SOD), Malondialdehyde (MDA), and Nitric Oxide (NO)
2.6. Effect of Fluoxetine, Wheatgrass, or Their Combination on Serum Creatinine and Urea
2.7. Effect of Fluoxetine, Wheatgrass, or Their Combination on Renal Total Antioxidant Capacity (TAC), Superoxide Dismutase (SOD), Malondialdehyde (MDA), and Nitric Oxide (NO)
2.8. Effect of Fluoxetine, Wheatgrass, or Their Combination on Renal Interleukin-6 (IL-6), Tumor Necrosis Factor-A (TNF-A), Nuclear Factor Kappa B (NF-Κb), and Caspase-3 Activity
2.9. Effect of Fluoxetine, Wheatgrass, or Their Combination on Cerebral Β-Catenin and Glycogen Synthase Kinase-3 Beta (GSK-3β)
2.10. Effect of Fluoxetine, Wheatgrass, or Their Combination on Cerebral Total Antioxidant Capacity (TAC), Superoxide Dismutase (SOD), and Malondialdehyde (MDA)
2.11. Effect of Fluoxetine, Wheatgrass, or Their Combination on Cerebral Neurotransmitters Dopamine (DA), Norepinephrine (NE), and Serotonin (5-HT)
2.12. Effect of Fluoxetine, Wheatgrass, or Their Combination on Cerebral Interleukin-1β (IL-1β) and Tumor Necrosis Factor-A (TNF-α)
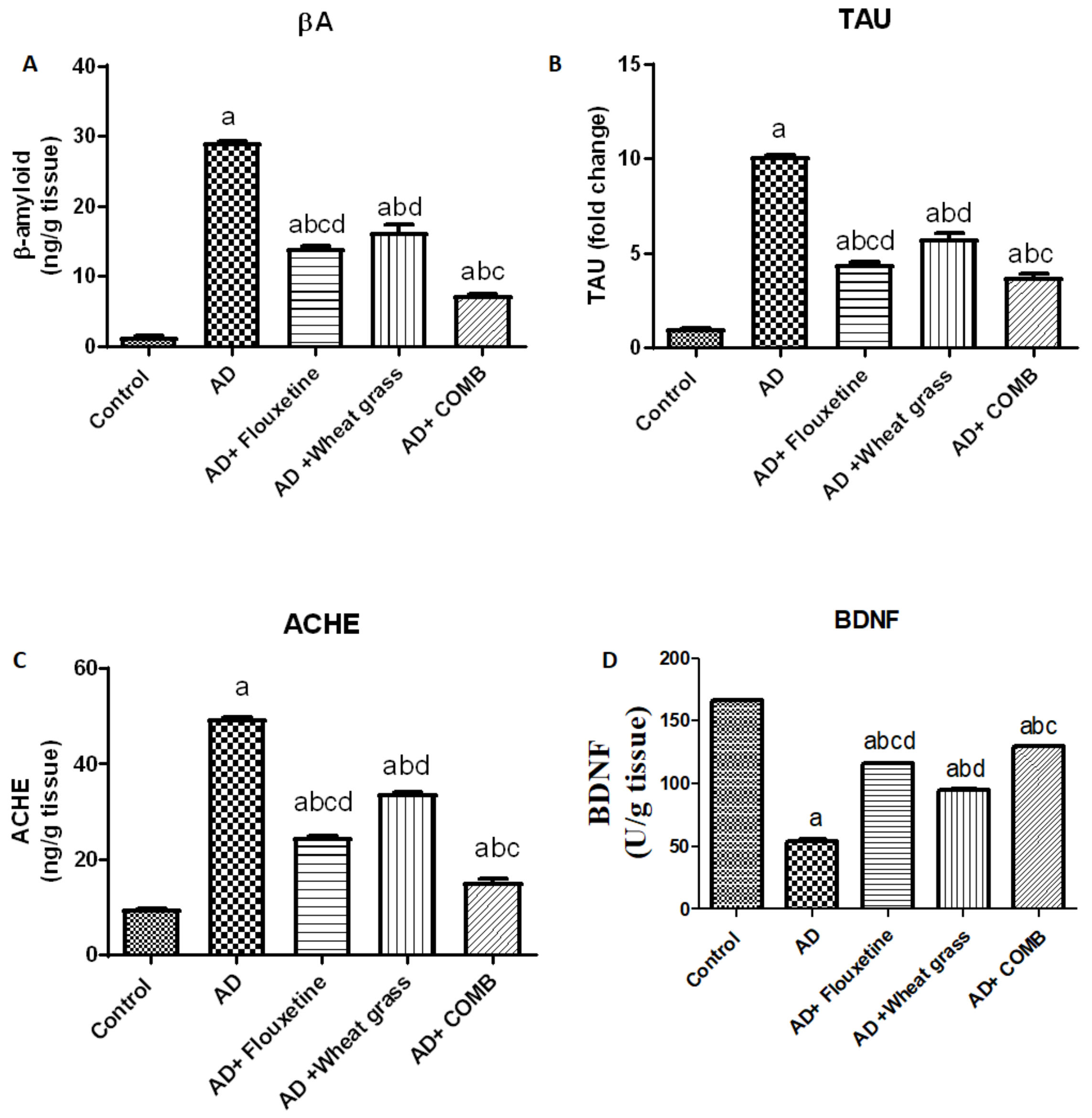
2.13. Effect of Fluoxetine, Wheatgrass, or Their Combination on Cerebral Beta-Amyloid (Aβ), Tau Protein (TAU), Acetylcholine Esterase (ACHE), and Brain-Derived Neurotrophic Factor (BDNF)
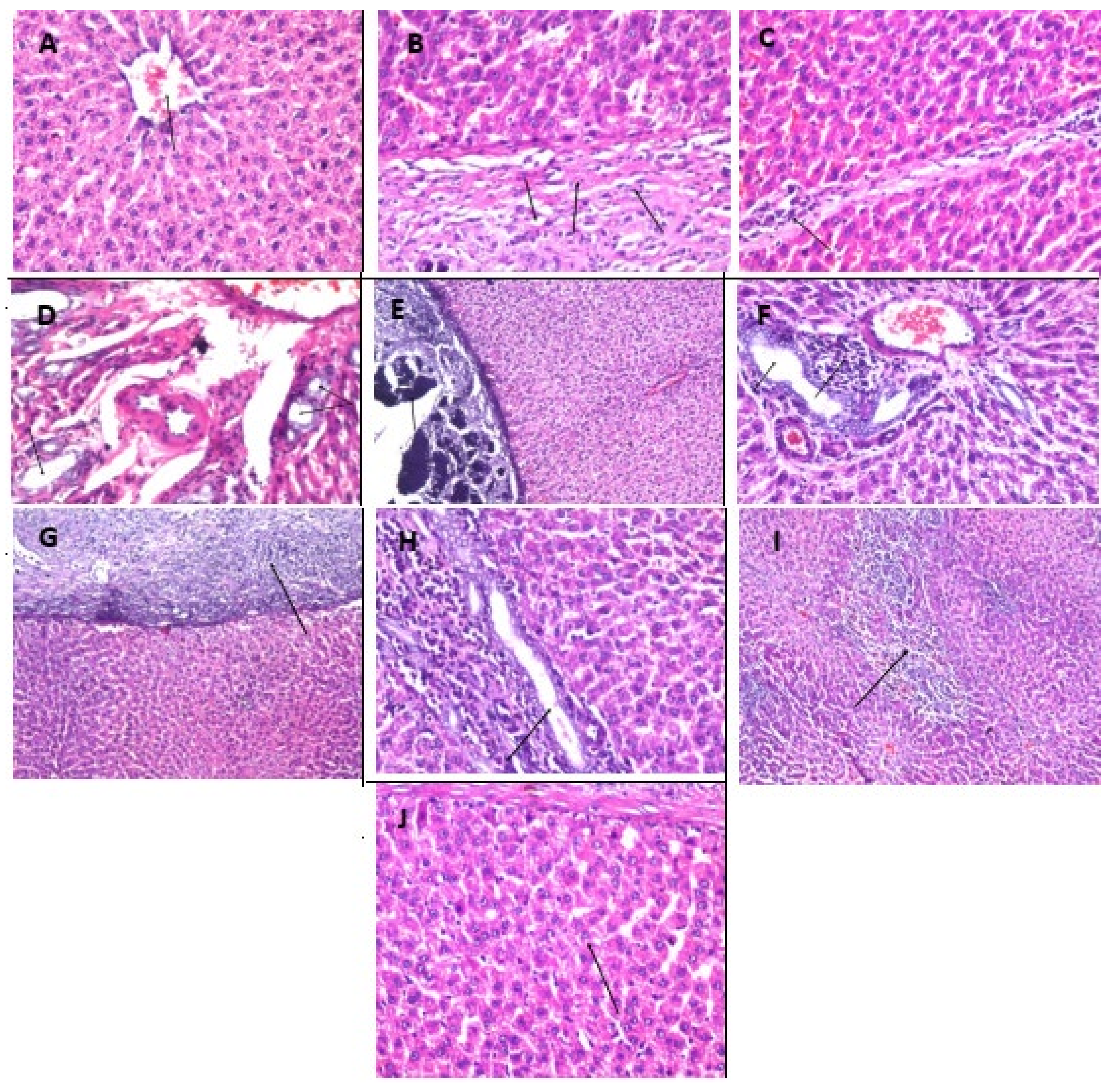
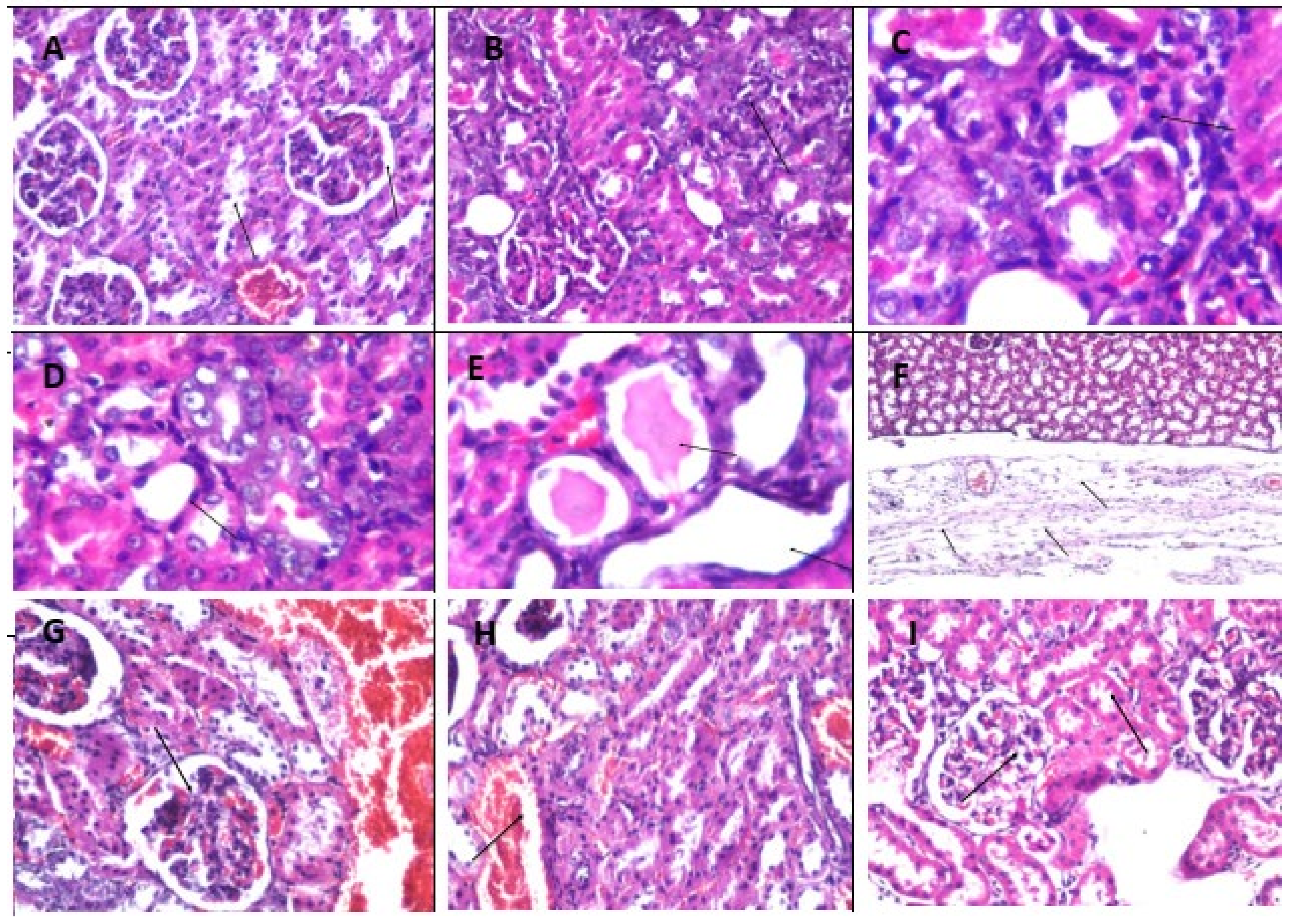
2.14. Effect of Fluoxetine, Wheatgrass, or Their Combination on Histopathological Examination of The Liver, Kidney, and Brain Tissue Specimens
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Ethical Statement
4.3. Drugs and Chemicals
4.4. Experimental Design
4.5. Behavioral Study (Morris Water Maze (MWM) Test)
4.6. Sample Preparation & Measurments
4.6.1. Estimation of Hepatic and Renal Functions
4.6.2. Estimation of Lipid Profile
4.6.3. Assessment of Apoptosis and Inflammatory Mediators
4.6.4. Brain, Hepatic, and Renal Oxidative Stress Markers
4.6.5. Determination of β-Catenin, GSK-3β in Brain Tissue
4.6.6. Determination of Brain Monoamines
4.6.7. Determination of Beta-Amyloid (Aβ) Content in Brain Tissue
4.6.8. Determination of Tau Protein (TAU) in Brain Tissue
4.6.9. Determination of ACHE in Brain Tissue
4.6.10. Determination of Brain-Derived Neurotrophic Factor (BDNF) in Brain Tissue
4.6.11. Histopathological Examination of Brain Tissue and Liver and Kidney
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- DeTure, M.A.; Dickson, D.W. The neuropathological diagnosis of Alzheimer’s disease. Mol. Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Baumgart, M.; Snyder, H.M.; Carrillo, M.C.; Fazio, S.; Kim, H.; Johns, H. Summary of the evidence on modifiable risk factors for cognitive decline and dementia: A population-based perspective. Alzheimer’s Dement. 2015, 11, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Qiu, C.; von Strauss, E.; Winblad, B.; Fratiglioni, L. APOE genotype, family history of dementia, and Alzheimer disease risk: A 6-year follow-up study. Arch. Neurol. 2004, 61, 1930–1934. [Google Scholar] [CrossRef] [Green Version]
- Xue, L.; Lou, Y.; Feng, X.; Wang, C.; Ran, Z.; Zhang, X. Prevalence of chronic kidney disease and associated factors among the Chinese population in Taian, China. BMC Nephrol. 2014, 15, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kivipelto, M.; Ngandu, T.; Fratiglioni, L.; Viitanen, M.; Kåreholt, I.; Winblad, B.; Helkala, E.-L.; Tuomilehto, J.; Soininen, H.; Nissinen, A. Obesity and vascular risk factors at midlife and the risk of dementia and Alzheimer disease. Arch. Neurol. 2005, 62, 1556–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, J.M.; Cooper, M.E. Mechanisms of diabetic complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef] [PubMed]
- Stanciu, G.D.; Bild, V.; Ababei, D.C.; Rusu, R.N.; Cobzaru, A.; Paduraru, L.; Bulea, D. Link between diabetes and Alzheimer’s disease due to the shared amyloid aggregation and deposition involving both neurodegenerative changes and neurovascular damages. J. Clin. Med. 2020, 9, 1713. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Wang, S.; Haynes, K.; Denburg, M.R.; Shin, D.B.; Gelfand, J.M. Risk of moderate to advanced kidney disease in patients with psoriasis: Population based cohort study. BMJ 2013, 347, f5961. [Google Scholar] [CrossRef] [Green Version]
- Mayer, F.; di Pucchio, A.; Lacorte, E.; Bacigalupo, I.; Marzolini, F.; Ferrante, G.; Minardi, V.; Masocco, M.; Canevelli, M.; di Fiandra, T. An estimate of attributable cases of Alzheimer disease and vascular dementia due to modifiable risk factors: The impact of primary prevention in Europe and in Italy. Dement. Geriatr. Cogn. Disord. Extra 2018, 8, 60–71. [Google Scholar] [CrossRef]
- He, Z.; Guo, J.L.; McBride, J.D.; Narasimhan, S.; Kim, H.; Changolkar, L.; Zhang, B.; Gathagan, R.J.; Yue, C.; Dengler, C. Amyloid-β plaques enhance Alzheimer’s brain tau-seeded pathologies by facilitating neuritic plaque tau aggregation. Nat. Med. 2018, 24, 29. [Google Scholar] [CrossRef]
- Ferreira-Vieira, H.T.; Guimaraes, M.I.; Silva, R.F.; Ribeiro, M.F. Alzheimer’s disease: Targeting the cholinergic system. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Hachisu, M.; Konishi, K.; Hosoi, M.; Tani, M.; Tomioka, H.; Inamoto, A.; Minami, S.; Izuno, T.; Umezawa, K.; Horiuchi, K. Beyond the hypothesis of serum anticholinergic activity in Alzheimer’s disease: Acetylcholine neuronal activity modulates brain-derived neurotrophic factor production and inflammation in the brain. Neurodegener. Dis. 2015, 15, 182–187. [Google Scholar] [CrossRef]
- Bencherif, M.; Lippiello, P.M.; Lucas, R.; Marrero, M.B. Alpha7 nicotinic receptors as novel therapeutic targets for inflammation-based diseases. Cell. Mol. Life Sci. 2011, 68, 931–949. [Google Scholar] [CrossRef] [Green Version]
- Ghiso, J.; Shayo, M.; Calero, M.; Ng, D.; Tomidokoro, Y.; Gandy, S.; Rostagno, A.; Frangione, B. Systemic catabolism of Alzheimer’s Aβ40 and Aβ42. J. Biol. Chem. 2004, 279, 45897–45908. [Google Scholar] [CrossRef] [Green Version]
- Wiseman, F.K.; Al-Janabi, T.; Hardy, J.; Karmiloff-Smith, A.; Nizetic, D.; Tybulewicz, V.L.; Fisher, E.M.; Strydom, A. A genetic cause of Alzheimer disease: Mechanistic insights from Down syndrome. Nat. Rev. Neurosci. 2015, 16, 564–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada, L.D.; Ahumada, P.; Cabrera, D.; Arab, J.P. Liver dysfunction as a novel player in Alzheimer’s progression: Looking outside the brain. Front. Aging Neurosci. 2019, 11, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanciu, G.D.; Ababei, D.C.; Bild, V.; Bild, W.; Paduraru, L.; Gutu, M.M.; Tamba, B.-I. Renal Contributions in the Pathophysiology and Neuropathological Substrates Shared by Chronic Kidney Disease and Alzheimer’s Disease. Brain Sci. 2020, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Y.; He, F.F.; Su, H.; Zhang, C.; Meng, X.F. Association between chronic kidney disease and Alzheimer’s disease: An update. Metab. Brain Dis. 2020, 35, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Mowla, A.; Mosavinasab, M.; Haghshenas, H.; Haghighi, A.B. Does serotonin augmentation have any effect on cognition and activities of daily living in Alzheimer’s dementia?: A double-blind, placebo-controlled clinical trial. J. Clin. Psychopharmacol. 2007, 27, 484–487. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Xu, H.; Zhu, S.; Wang, H.; He, J.; Zhang, H.; Guo, H.; Kong, J.; Huang, Q. Fluoxetine improves behavioral performance by suppressing the production of soluble β-amyloid in APP/PS1 mice. Curr. Alzheimer Res. 2014, 11, 672–680. [Google Scholar] [CrossRef]
- Ma, J.; Gao, Y.; Jiang, L.; Chao, F.-L.; Huang, W.; Zhou, C.-N.; Tang, W.; Zhang, L.; Huang, C.-X.; Zhang, Y. Fluoxetine attenuates the impairment of spatial learning ability and prevents neuron loss in middle-aged APPswe/PSEN1dE9 double transgenic Alzheimer’s disease mice. Oncotarget 2017, 8, 27676. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.-S.; Gao, L.-F.; Jin, L.; Wu, H.; Wang, Q.; Zhou, Y.; Fan, S.; Jiang, X.; Ke, D.; Lei, H. Fluoxetine administration during adolescence attenuates cognitive and synaptic deficits in adult 3 × TgAD mice. Neuropharmacology 2017, 126, 200–212. [Google Scholar] [CrossRef]
- Li, X.; Zhu, W.; Roh, M.-S.; Friedman, A.B.; Rosborough, K.; Jope, R.S. In vivo regulation of glycogen synthase kinase-3 β (GSK3 β) by serotonergic activity in mouse brain. Neuropsychopharmacology 2004, 29, 1426–1431. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Liang, Y.; Chen, H.; Xu, B.; Chai, C.; Xing, P. The role of fluoxetine in activating Wnt/β-catenin signaling and repressing β-amyloid production in an Alzheimer mouse model. Front. Aging Neurosci. 2018, 10, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florent-Béchard, S.; Malaplate-Armand, C.; Koziel, V.; Kriem, B.; Olivier, J.-L.; Pillot, T.; Oster, T. Towards a nutritional approach for prevention of Alzheimer’s disease: Biochemical and cellular aspects. J. Neurol. Sci. 2007, 262, 27–36. [Google Scholar] [CrossRef]
- Rimple, K.M.; Kumar, R.; Newton, A.; Reeta, H.S. Poly pharmacological effects of green blood therapy: An update. World J. Pharm. Med Res. 2016, 2, 10–21. [Google Scholar]
- Kulkarni, S.D.; Tilak, J.C.; Acharya, R.; Rajurkar, N.S.; Devasagayam, T.; Reddy, A. Evaluation of the antioxidant activity of wheatgrass (Triticum aestivum L.) as a function of growth under different conditions. Phytother. Res. An Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2006, 20, 218–227. [Google Scholar] [CrossRef]
- Barreira, J.C.; Ferreira, I.C.; Oliveira, M.B.P.; Pereira, J.A. Antioxidant activities of the extracts from chestnut flower, leaf, skins and fruit. Food Chem. 2008, 107, 1106–1113. [Google Scholar] [CrossRef]
- Rukkumani, R.; Aruna, K.; Varma, P.S.; Menon, V.P. Ferulic acid, a natural phenolic antioxidant modulates altered lipid profiles during alcohol and thermally oxidized sunflower oil induced toxicity. J. Nutraceuticals Funct. Med. Foods 2005, 4, 119–132. [Google Scholar] [CrossRef]
- Matyja, E. Aluminum enhances glutamate-mediated neurotoxicity in organotypic cultures of rat hippocampus. Folia Neuropathol. 2000, 38, 47–53. [Google Scholar]
- Cao, Z.; Wang, F.; Xiu, C.; Zhang, J.; Li, Y. Hypericum perforatum extract attenuates behavioral, biochemical, and neurochemical abnormalities in Aluminum chloride-induced Alzheimer’s disease rats. Biomed. Pharmacother. 2017, 91, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Adlard, P.A.; Bush, A.I. Metals and Alzheimer’s disease: How far have we come in the clinic? J. Alzheimer’s Dis. 2018, 62, 1369–1379. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Mucke, L. Alzheimer mechanisms and therapeutic strategies. Cell 2012, 148, 1204–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Taweel, G.M.; Ajarem, J.S.; Ahmad, M. Neurobehavioral toxic effects of perinatal oral exposure to aluminum on the developmental motor reflexes, learning, memory and brain neurotransmitters of mice offspring. Pharmacol. Biochem. Behav. 2012, 101, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-T.; Chen, R.-C.; Lu, W.-W.; Liu, S.-H.; Yang, F.-Y. Protective effects of low-intensity pulsed ultrasound on aluminum-induced cerebral damage in Alzheimer’s disease rat model. Sci. Rep. 2015, 5, 9671. [Google Scholar] [CrossRef] [Green Version]
- Foster, H.D. How aluminum causes Alzheimer’s disease: The implications for prevention and treatment of Foster’s multiple antagonist hypothesis. J. Orthomol. Med. 2000, 15, 21–51. [Google Scholar]
- García-Velázquez, L.; Arias, C. The emerging role of Wnt signaling dysregulation in the understanding and modification of age-associated diseases. Ageing Res. Rev. 2017, 37, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Piña-Crespo, J.; Li, Y. Restoring Wnt/β-catenin signaling is a promising therapeutic strategy for Alzheimer’s disease. Mol. Brain 2019, 12, 104. [Google Scholar] [CrossRef] [PubMed]
- Scali, C.; Caraci, F.; Gianfriddo, M.; Diodato, E.; Roncarati, R.; Pollio, G.; Gaviraghi, G.; Copani, A.; Nicoletti, F.; Terstappen, G.C. Inhibition of Wnt signaling, modulation of Tau phosphorylation and induction of neuronal cell death by DKK1. Neurobiol. Dis. 2006, 24, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Samadi, A.; Valderas, C.; de los Ríos, C.; Bastida, A.; Chioua, M.; González-Lafuente, L.; Colmena, I.; Gandía, L.; Romero, A.; del Barrio, L. Cholinergic and neuroprotective drugs for the treatment of Alzheimer and neuronal vascular diseases. II. Synthesis, biological assessment, and molecular modelling of new tacrine analogues from highly substituted 2-aminopyridine-3-carbonitriles. Bioorg. Med. Chem. 2011, 19, 122–133. [Google Scholar] [CrossRef]
- Ballard, C.G.; Greig, N.H.; Guillozet-Bongaarts, A.L.; Enz, A.; Darvesh, S. Cholinesterases: Roles in the brain during health and disease. Curr. Alzheimer Res. 2005, 2, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.A. Neuroinflammatory changes negatively impact on LTP: A focus on IL-1β. Brain Res. 2015, 1621, 197–204. [Google Scholar] [CrossRef]
- Balietti, M.; Giuli, C.; Conti, F. Peripheral blood brain-derived neurotrophic factor as a biomarker of Alzheimer’s disease: Are there methodological biases? Mol. Neurobiol. 2018, 55, 6661–6672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.-H.; Yu, J.-T.; Tan, L. Brain-derived neurotrophic factor in Alzheimer’s disease: Risk, mechanisms, and therapy. Mol. Neurobiol. 2015, 52, 1477–1493. [Google Scholar] [CrossRef] [PubMed]
- Giacobbo, B.L.; Doorduin, J.; Klein, H.C.; Dierckx, R.A.; Bromberg, E.; de Vries, E.F. Brain-derived neurotrophic factor in brain disorders: Focus on neuroinflammation. Mol. Neurobiol. 2019, 56, 3295–3312. [Google Scholar] [CrossRef] [Green Version]
- Beattie, E.C.; Stellwagen, D.; Morishita, W.; Bresnahan, J.C.; Ha, B.K.; Von Zastrow, M.; Beattie, M.S.; Malenka, R.C. Control of synaptic strength by glial TNFα. Science 2002, 295, 2282–2285. [Google Scholar] [CrossRef]
- Cheng, X.; Yang, L.; He, P.; Li, R.; Shen, Y. Differential activation of tumor necrosis factor receptors distinguishes between brains from Alzheimer’s disease and non-demented patients. J. Alzheimer’s Dis. 2010, 19, 621–630. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Zhang, Y.; Zhan, J.; Wang, C.; Du, X.; Yin, G.; Cao, B.; Ning, Y.; Soares, J.; Zhang, X. Burnout and cognitive impairment: Associated with serum BDNF in a Chinese Han population. Psychoneuroendocrinology 2017, 77, 236–243. [Google Scholar] [CrossRef]
- Francis, B.M.; Yang, J.; Hajderi, E.; Brown, M.E.; Michalski, B.; McLaurin, J.; Fahnestock, M.; Mount, H.T. Reduced tissue levels of noradrenaline are associated with behavioral phenotypes of the TgCRND8 mouse model of Alzheimer’s disease. Neuropsychopharmacology 2012, 37, 1934–1944. [Google Scholar] [CrossRef]
- Marien, M.R.; Colpaert, F.C.; Rosenquist, A.C. Noradrenergic mechanisms in neurodegenerative diseases: A theory. Brain Res. Rev. 2004, 45, 38–78. [Google Scholar] [CrossRef] [PubMed]
- Yousef, M.I.; Mutar, T.F.; Kamel, M.A.E.-N. Hepato-renal toxicity of oral sub-chronic exposure to aluminum oxide and/or zinc oxide nanoparticles in rats. Toxicol. Rep. 2019, 6, 336–346. [Google Scholar] [CrossRef]
- Wang, X.; Gong, J.; Gui, Z.; Hu, T.; Xu, X. Halloysite nanotubes-induced Al accumulation and oxidative damage in liver of mice after 30-day repeated oral administration. Environ. Toxicol. 2018, 33, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Al-Kahtani, M.; Morsy, K. Ameliorative effect of selenium nanoparticles against aluminum chloride-induced hepatorenal toxicity in rats. Environ. Sci. Pollut. Res. 2019, 26, 32189–32197. [Google Scholar] [CrossRef] [PubMed]
- Kellett, K.A.; Williams, J.; Vardy, E.R.; Smith, A.D.; Hooper, N.M. Plasma alkaline phosphatase is elevated in Alzheimer’s disease and inversely correlates with cognitive function. Int. J. Mol. Epidemiol. 2011, 2, 114. [Google Scholar]
- Miranda, A.S.; Cordeiro, T.M.; dos Santos Lacerda Soares, T.M.; Ferreira, R.N.; Simoes e Silva, A.C. Kidney–brain axis inflammatory cross-talk: From bench to bedside. Clin. Sci. 2017, 131, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- McAdams-DeMarco, M.A.; Bae, S.; Chu, N.; Gross, A.L.; Brown, C.H.; Oh, E.; Rosenberg, P.; Neufeld, K.J.; Varadhan, R.; Albert, M. Dementia and Alzheimer’s disease among older kidney transplant recipients. J. Am. Soc. Nephrol. 2017, 28, 1575–1583. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2294. [Google Scholar] [CrossRef] [Green Version]
- Hooijmans, C.R.; Kiliaan, A.J. Fatty acids, lipid metabolism and Alzheimer pathology. Eur. J. Pharmacol. 2008, 585, 176–196. [Google Scholar] [CrossRef]
- Durairaj, V.; Shakya, G.; Rajagopalan, R. Hepatoprotective role of wheatgrass on alcohol and ΔPUFA-induced oxidative stress in rats. J. Diet. Suppl. 2015, 12, 126–137. [Google Scholar] [CrossRef]
- Rana, S.; Kamboj, J.K.; Gandhi, V. Living life the natural way–Wheatgrass and Health. Funct. Foods Health Dis. 2011, 1, 444–456. [Google Scholar] [CrossRef]
- Eissa, H.A.; Mohamed, S.S.; Hussein, A.M. Nutritional value and impact of wheatgrass juice (Green Blood Therapy) on increasing fertility in male albino rats. Bull. Natl. Res. Cent. 2020, 44, 30. [Google Scholar] [CrossRef]
- Bhondave, P.D.; Devarshi, P.P.; Mahadik, K.R.; Harsulkar, A.M. ‘Ashvagandharishta’ prepared using yeast consortium from Woodfordia fruticosa flowers exhibit hepatoprotective effect on CCl4 induced liver damage in Wistar rats. J. Ethnopharmacol. 2014, 151, 183–190. [Google Scholar] [CrossRef]
- Galanos, C.; Freudenberg, M.A. Mechanisms of endotoxin shock and endotoxin hypersensitivity. Immunobiology 1993, 187, 346–356. [Google Scholar] [CrossRef]
- Baldwin, A.S. Regulation of cell death and autophagy by IKK and NF-κB: Critical mechanisms in immune function and cancer. Immunol. Rev. 2012, 246, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Poudel, B.; Nepali, S.; Xin, M.; Ki, H.H.; Kim, Y.H.; Kim, D.K.; Lee, Y.M. Flavonoids from Triticum aestivum inhibit adipogenesis in 3T3-L1 cells by upregulating the insig pathway. Mol. Med. Rep. 2015, 12, 3139–3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abed, K.; Yaqoob, K.; Abdoh, A.; Mohammed, S.; Pankaj, T.; Hakeem, S.; Mamoon, H. Investigation of antigenotoxic potential of wheatgrass (Triticum aestivum) powder on cyclophosphamide induced genotoxicity and oxidative stress in mice. Austin J. Pharmacol. Ther. 2017, 5, 1098. [Google Scholar]
- Olajide, O.J.; Yawson, E.O.; Gbadamosi, I.T.; Arogundade, T.T.; Lambe, E.; Obasi, K.; Lawal, I.T.; Ibrahim, A.; Ogunrinola, K.Y. Ascorbic acid ameliorates behavioural deficits and neuropathological alterations in rat model of Alzheimer’s disease. Environ. Toxicol. Pharmacol. 2017, 50, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Renshaw, P.F.; Parsegian, A.; Yang, C.K.; Novero, A.; Yoon, S.J.; Lyoo, I.K.; Cohen, B.M.; Carlezon, W.A., Jr. Lovastatin potentiates the antidepressant efficacy of fluoxetine in rats. Pharmacol. Biochem. Behav. 2009, 92, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.A.; Ahmed, H.I.; Abu-Elfotuh, K. Modeling stages mimic Alzheimer’s disease induced by different doses of aluminum in rats: Focus on progression of the disease in response to time. J. Alzheimer’s Parkinsonism Dement. 2016, 1, 1–11. [Google Scholar]
- Pazini, F.L.; Cunha, M.P.; Rosa, J.M.; Colla, A.R.; Lieberknecht, V.; Oliveira, Á.; Rodrigues, A.L.S. Creatine, similar to ketamine, counteracts depressive-like behavior induced by corticosterone via PI3K/Akt/mTOR pathway. Mol. Neurobiol. 2016, 53, 6818–6834. [Google Scholar] [CrossRef] [PubMed]
- Bitra, V.R.; Rapaka, D.; Mathala, N.; Akula, A. Effect of wheat grass powder on aluminum induced Alzheimer’s disease in Wistar rats. Asian Pac. J. Trop. 2014, 7, S278–S281. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-J.; Liu, Y.-L.; Zhong, Q.; Yu, Y.-F.; Su, H.-L.; Toque, H.A.; Dang, Y.-H.; Chen, F.; Xu, M.; Chen, T. Tetrahydropalmatine protects against methamphetamine-induced spatial learning and memory impairment in mice. Neurosci. Bull. 2012, 28, 222–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.-T.; Wang, Z.-J.; Cai, H.-Y.; Yuan, L.; Hu, M.-M.; Wu, M.-N.; Qi, J.-S. Sex differences in neuropathology and cognitive behavior in APP/PS1/tau triple-transgenic mouse model of Alzheimer’s disease. Neurosci. Bull. 2018, 34, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Tsakiris, S.; Schulpis, K.H.; Marinou, K.; Behrakis, P. Protective effect of L-cysteine and glutathione on the modulated suckling rat brain Na+, K+-ATPase and Mg2+-ATPase activities induced by the in vitro galactosaemia. Pharmacol. Res. 2004, 49, 475–479. [Google Scholar] [CrossRef]
- Welch, A.S.; Welch, B.L. Isolation, reactivity and aggression: Evidence for an involvement of brain catecholamines and serotonin. In The Physiology of Aggression and Defeat; Springer: Boston, MA, USA, 1971; pp. 91–142. [Google Scholar]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MWM | “Time Spent (sec) in Target Quadrant” | “Escape Latency (sec) for Total Four Days” |
|---|---|---|
| Control | 50.80 ± 1.43 | 64.93 ± 1.06 |
| AD | 17.00 a ± 1.14 | 147.7 a ± 2.96 |
| AD/Fluoxetine | 34.80 abcd ± 1.74 | 79.02 abc ± 0.37 |
| AD/Wheatgrass | 39.20 abd ± 0.86 | 97.03 abd ± 0.48 |
| AD/Combination | 46.20 bc ± 0.86 | 78.08 bc ± 0.60 |
| Serum | Control | AD | AD/Fluoxetine | AD/Wheatgrass | AD/Combination |
|---|---|---|---|---|---|
| ALT (U/L) | 14.13 ± 0.56 | 97.83 a ± 0.78 | 70.63 abcd ± 1.57 | 39.63 abd ± 0.59 | 30.63 abc ± 0.59 |
| AST (U/L) | 21.10 ± 1.04 | 92.03 a ± 0.73 | 53.38 abcd ± 1.45 | 42.83 abd ± 1.83 | 31.78 abc ± 0.59 |
| ALP (U/L) | 114.61 ± 0.39 | 372.80 a ± 3.05 | 198.72 abcd ± 0.97 | 186.04 abd ± 1.47 | 166.03 abc ± 7.91 |
| Creatinine (mg/dL) | 0.29 ± 0.01 | 4.03 a ± 0.21 | 2.85 abcd ± 0.03 | 1.80 abd ± 0.15 | 0.76 bc ± 0.01 |
| Urea (mg/dL) | 45.13 ± 1.89 | 90.20 a ± 0.25 | 63.56 abcd ± 0.84 | 49.36 b ± 1.81 | 45.38 b ± 1.48 |
| TC (mg/mL) | 132.72 ± 1.85 | 301.90 a ± 2.51 | 206.04 abcd ± 1.47 | 175.63 ab ± 1.57 | 171.90 ab ± 2.95 |
| TG (mg/mL) | 72.05 ± 1.42 | 135.21 a ± 1.56 | 85.06 abcd ± 1.16 | 79.40 ab ± 0.32 | 76.42 b ± 0.65 |
| HDL (mg/mL) | 63.94 ± 0.85 | 18.93 a ± 0.66 | 40.48 abd ± 0.70 | 41.81 abd ± 0.27 | 48.85 abc ± 0.67 |
| Hepatic | Control | AD | AD/Fluoxetine | AD/Wheatgrass | AD/Combination |
|---|---|---|---|---|---|
| IL-6 (pg/mg) | 31.45 ± 0.37 | 131.92 a ± 0.64 | 90.22 abcd ± 1.59 | 77.93 abd ± 0.72 | 61.62 abc ± 0.36 |
| TNF-α (pg/mg) | 32.67 ± 1.27 | 120.93 a ± 0.41 | 98.34 abcd ± 0.67 | 83.56 abd ± 0.84 | 63.03 abc ± 2.43 |
| NF-κB (pg/mg) | 1.00 ± 0.01 | 9.98 a ± 0.06 | 6.19 abcd ± 0.14 | 4.695 ab ± 0.11 | 2.38 abc ± 0.07 |
| Caspase-3 Activity (µM pNA/min/mL) | 1.98 ± 0.03 | 5.06 a ± 0.02 | 3.72 abcd ± 0.06 | 2.78 ab ± 0.03 | 2.72 ab ± 0.06 |
| SOD (U/mg) | 2.96 ± 0.04 | 0.39 a ± 0.02 | 1.01 abcd ± 0.02 | 1.65 abd ± 0.12 | 2.22 abc ± 0.08 |
| MDA (mmol/g) | 11.25 ± 0.41 | 104.91 a ± 2.70 | 88.58 abcd ± 0.76 | 52.48 abd ± 0.81 | 28.94 abc ± 1.33 |
| TAC (nmol/mg) | 27.98 ± 0.44 | 9.32 a ± 0.34 | 14.96 abcd ± 0.10 | 11.88 abd ± 0.17 | 21.28 abc ± 0.44 |
| NO (nmol/mg) | 1.63 ± 0.04 | 20.30 a ± 0.58 | 12.43 abcd ± 0.53 | 7.94 abd ± 0.16 | 6.10 abc ± 0.05 |
| Renal | Control | AD | AD/Fluoxetine | AD/Wheatgrass | AD/Combination |
|---|---|---|---|---|---|
| IL-6 (pg/mg) | 33.50 ± 2.15 | 99.06 a ± 1.23 | 66.49 abcd ± 1.55 | 56.36 ab ± 1.27 | 51.58 ab ± 0.66 |
| TNF-α (pg/mg) | 36.43 ± 0.22 | 135.6 a ± 1.41 | 84.79 abc ± 1.68 | 85.13 abd ± 1.29 | 71.91 abc ± 0.33 |
| NF-κB (pg/mg) | 1.03 ± 0.04 | 4.69 a ± 0.05 | 3.17 abc ± 0.09 | 2.99 abd ± 0.02 | 2.69 abc ± 0.05 |
| Caspase-3 Activity (µM pNA/min/mL) | 2.87 ± 0.06 | 19.36 a ± 0.27 | 7.98 abc ± 0.14 | 8.12 abd ± 0.17 | 7.09 abc ± 0.24 |
| SOD (U/mg) | 2.36 ± 0.09 | 0.28 a ± 0.01 | 0.78 abc ± 0.03 | 0.92 abd ± 0.01 | 1.35 abc ± 0.11 |
| MDA (mmol/g) | 6.68 ± 0.04 | 44.66 a ± 1.23 | 27.23 abcd ± 1.32 | 16.71 abd ± 0.78 | 11.08 abc ± 0.56 |
| TAC (nmol/mg) | 29.60 ± 0.50 | 12.3 a ± 0.21 | 16.70 abcd ± 0.32 | 18.72 abd ± 0.14 | 21.83 abc ± 0.56 |
| NO (nmol/mg) | 0.94 ± 0.03 | 11.65 a ± 0.37 | 8.17 abcd ± 0.09 | 5.12 abd ± 0.17 | 2.99 abc ± 0.03 |
| Cerebral | Control | AD | AD/Fluoxetine | AD/Wheatgrass | AD/Combination |
|---|---|---|---|---|---|
| β –Catenine (nmol/mg) | 3.19 ± 0.07 | 0.61 a ± 0.07 | 1.78 abcd ± 0.06 | 2.50 ab ± 0.03 | 3.13 b ± 0.09 |
| GSK-3β (nmol/mg) | 1.01 ± 0.01 | 10.12 a ± 0.10 | 5.78 abc ± 0.29 | 5.65 abd ± 0.21 | 3.75 abc ± 0.14 |
| DA (nmol/mg) | 68.18 ± 0.91 | 16.24 a ± 0.51 | 38.76 abcd ± 0.82 | 28.28 abd ± 0.54 | 45.14 abc ± 0.01 |
| NE (nmol/mg) | 721.1 ± 2.66 | 236.1 a ± 0.47 | 584.1 abcd ± 3.85 | 451.1 abd ± 5.05 | 591.2 abc ± 2.91 |
| 5-HT (nmol/mg) | 11.56 ± 0.08 | 4.06 a ± 0.05 | 9.03 abcd ± 0.51 | 6.85 abd ± 0.08 | 10.52 bc ± 0.31 |
| IL-1β (pg/mg) | 28.20 ± 0.43 | 117.94 a ± 1.01 | 57.57 abcd ± 2.96 | 83.33 abd ± 3.04 | 54.03 abc ± 0.86 |
| TNF-α (pg/mg) | 27.02 ± 0.04 | 212.13 a ± 4.36 | 63.18 abcd ± 0.45 | 87.53 abd ± 0.56 | 59.43 abc ± 2.03 |
| SOD (U/mg) | 3.62 ± 0.04 | 0.32 a ± 0.02 | 1.78 abcd ± 0.06 | 2.31 abd ± 0.06 | 2.78 abc ± 0.04 |
| MDA (mmol/g) | 6.46 ± 0.19 | 99.10 a ± 3.56 | 36.07 abcd ± 1.87 | 45.38 abd ± 2.02 | 31.73 abc ± 1.49 |
| TAC (nmol/mg) | 32.78 ± 0.76 | 9.15 a ± 0.45 | 18.55 abc ± 0.15 | 17.73 abd ± 1.01 | 22.32 abc ± 0.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abu-Elfotuh, K.; Ragab, G.M.; Salahuddin, A.; Jamil, L.; Abd Al Haleem, E.N. Attenuative Effects of Fluoxetine and Triticum aestivum against Aluminum-Induced Alzheimer’s Disease in Rats: The Possible Consequences on Hepatotoxicity and Nephrotoxicity. Molecules 2021, 26, 6752. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216752
Abu-Elfotuh K, Ragab GM, Salahuddin A, Jamil L, Abd Al Haleem EN. Attenuative Effects of Fluoxetine and Triticum aestivum against Aluminum-Induced Alzheimer’s Disease in Rats: The Possible Consequences on Hepatotoxicity and Nephrotoxicity. Molecules. 2021; 26(21):6752. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216752
Chicago/Turabian StyleAbu-Elfotuh, Karema, Ghada M. Ragab, Ahmad Salahuddin, Lubna Jamil, and Ekram Nemr Abd Al Haleem. 2021. "Attenuative Effects of Fluoxetine and Triticum aestivum against Aluminum-Induced Alzheimer’s Disease in Rats: The Possible Consequences on Hepatotoxicity and Nephrotoxicity" Molecules 26, no. 21: 6752. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216752