
Biological Response Profiling Reveals the Functional Differences of Main Alkaloids in Rhizoma Coptidis

Abstract
:1. Introduction
2. Results
2.1. HPLC Analysis of Alkaloids in Crude RC Extract
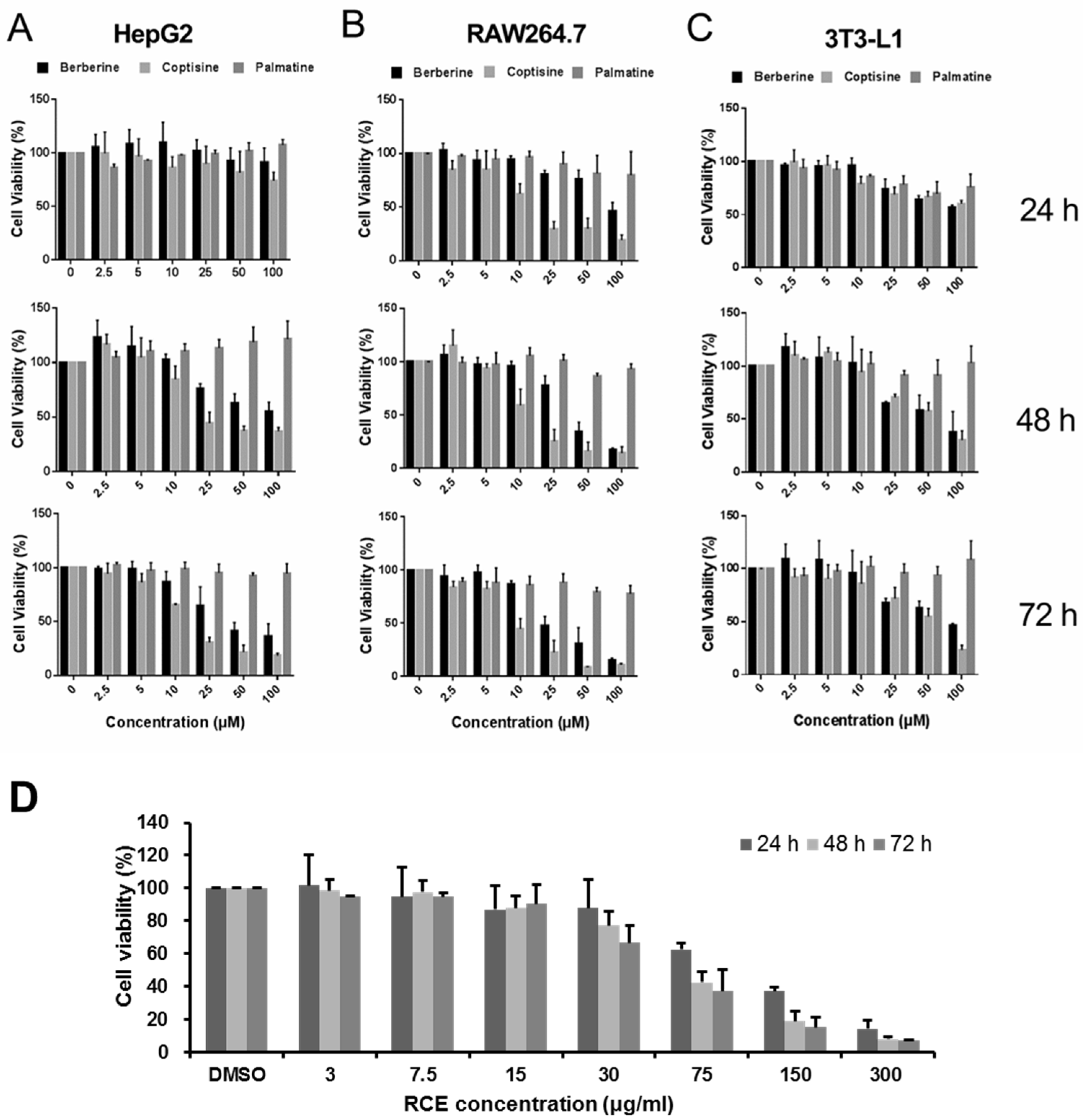
2.2. Comparison of the Effects of Alkaloids on the Proliferation of Different Cell Lines
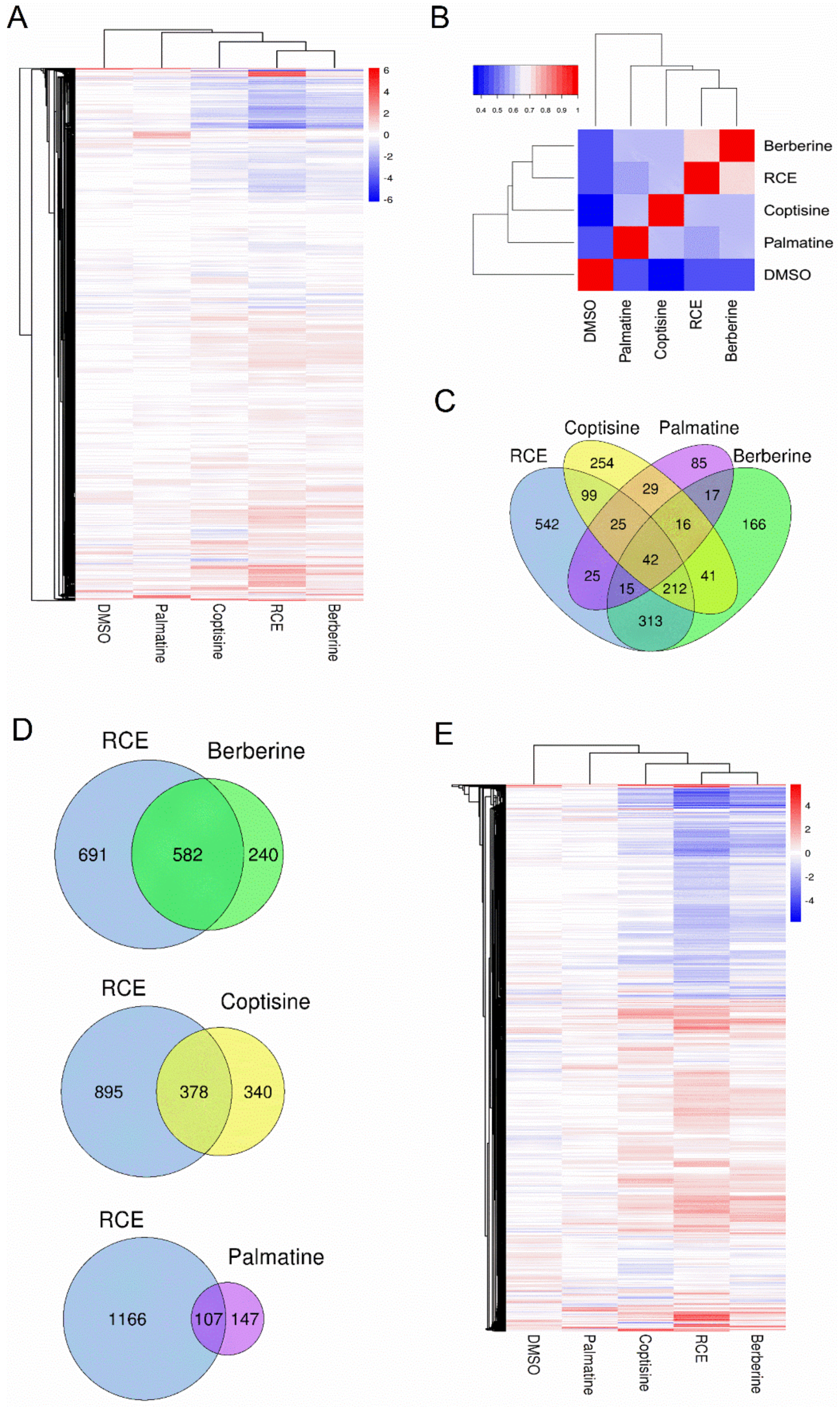
2.3. Transcriptome Analysis of RCE and Three Representative Alkaloids in RAW264.7 Cells
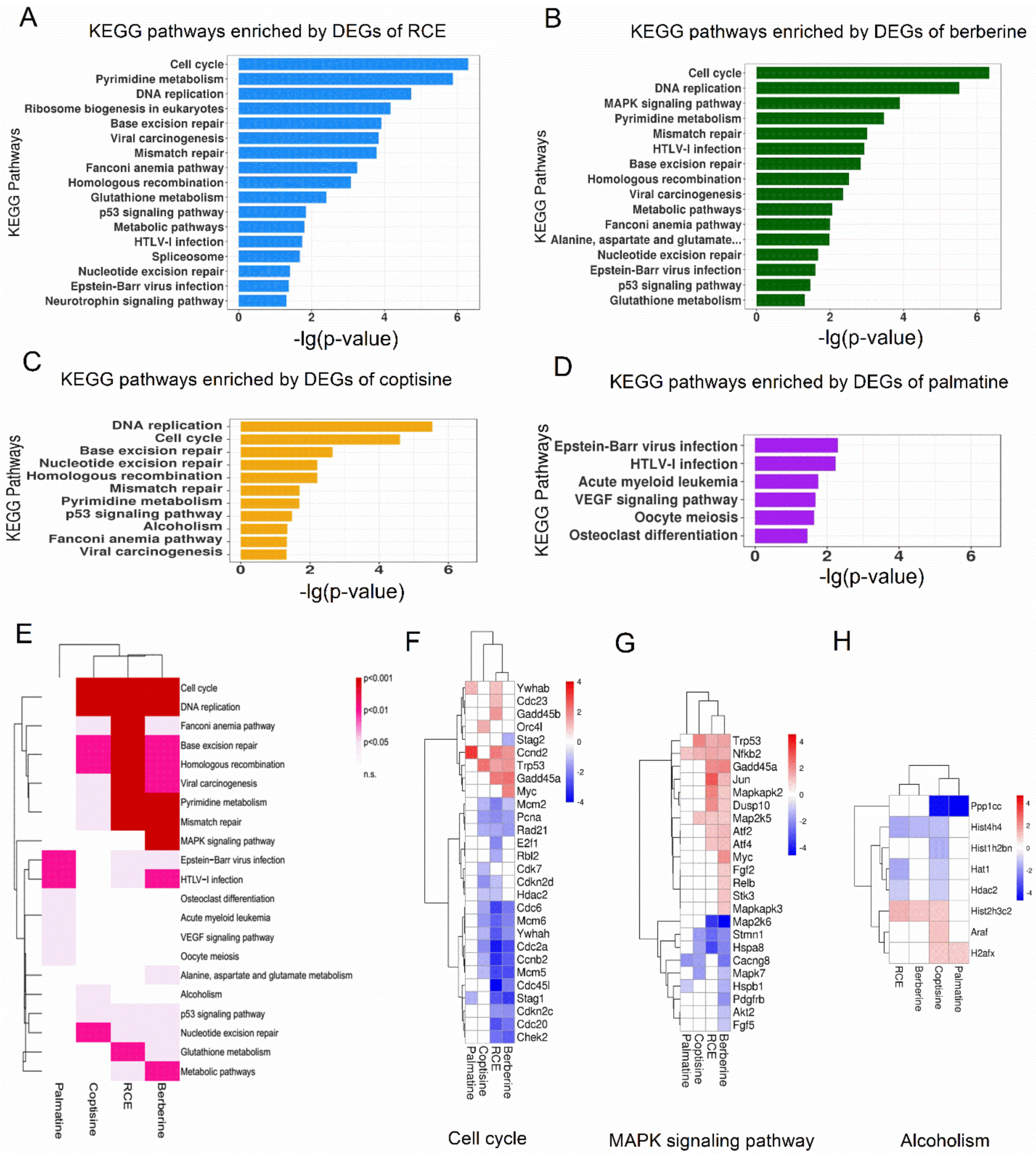
2.4. Pathway Analysis of RCE and Three Representative Alkaloids in RAW264.7 Cells
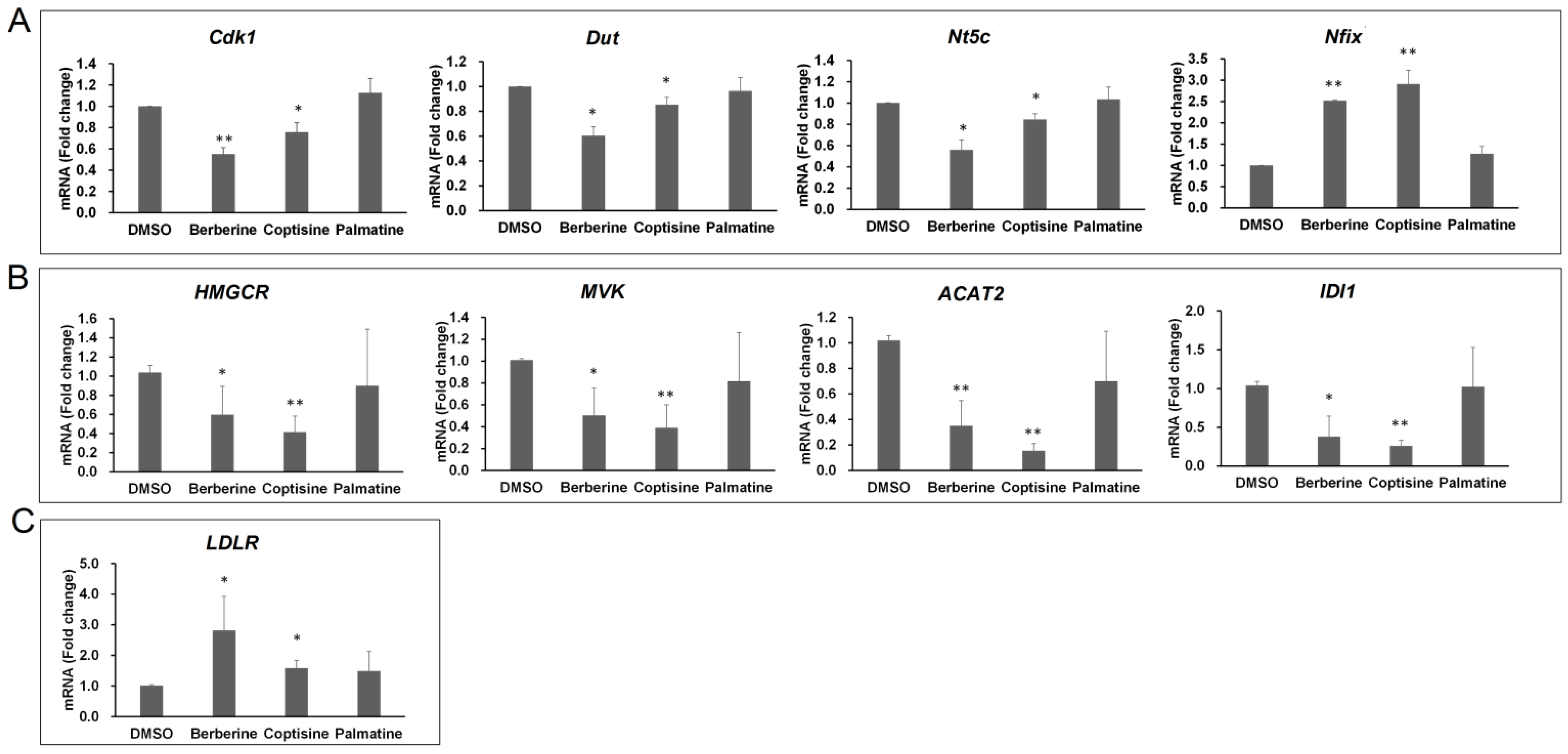
2.5. Comparative Analysis of Berberine and Coptisine on Genes Involved in Different Biological Processes
3. Discussion
4. Materials and Methods
4.1. Cells and Agents
4.2. Preparation of Crude Rhizoma Coptidis Extract (RCE)
4.3. HPLC Analysis
4.4. Cell Proliferation Assay
4.5. RNA Extraction, cDNA Synthesis, and Quantitative Real-Time PCR (qRT-PCR)
4.6. Gene Expression Profiling and Analysis
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Commission, P. The Pharmacopoeia of People’s Republic of China 2020 Edition; China Medical Science and Technology Press: Beijing, China, 2020; pp. 316–317. [Google Scholar]
- Meng, F.C.; Wu, Z.F.; Yin, Z.Q.; Lin, L.G.; Wang, R.; Zhang, Q.W. Coptidis rhizoma and its main bioactive components: Recent advances in chemical investigation, quality evaluation and pharmacological activity. Chin. Med. 2018, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, F.; Liu, J.; Lee, F.S.; Wang, X.; Yang, H. Analysis of alkaloids in Coptis chinensis Franch by accelerated solvent extraction combined with ultra performance liquid chromatographic analysis with photodiode array and tandem mass spectrometry detections. Anal. Chim. Acta 2008, 613, 184–195. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, H.; Wang, X.; Lee, F.S.; Yang, H.; Zheng, L. Analysis of major alkaloids in Rhizoma coptidis by capillary electrophoresis-electrospray-time of flight mass spectrometry with different background electrolytes. Electrophoresis 2008, 29, 2135–2147. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, L.; Lou, G.H.; Zeng, H.R.; Hu, J.; Huang, Q.W.; Peng, W.; Yang, X.B. Coptidis Rhizoma: A comprehensive review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. Pharm. Biol. 2019, 57, 193–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Tan, H.Y.; Li, L.; Yuen, M.F.; Feng, Y. Berberine and Coptidis Rhizoma as potential anticancer agents: Recent updates and future perspectives. J. Ethnopharmacol. 2015, 176, 35–48. [Google Scholar] [CrossRef]
- Feng, J.; Li, H.; Zhao, W.; Dang, H.; Wang, R.; Luo, K.; Guo, H.; Xing, W.; Cheng, J.; Song, W. Biological profiling-based systematic analysis of Rhizoma Coptidis from different growing regions and its anti-cholesterol biosynthesis activity on HepG2 cells. Mol. Pharm. 2018, 15, 2234. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mu, W.; Shang, H.; Lin, J.; Lei, X. The antihyperglycemic effects of Rhizoma Coptidis and mechanism of actions: A review of systematic reviews and pharmacological research. BioMed Res. Int. 2016, 2014, 798093. [Google Scholar] [CrossRef] [Green Version]
- Zou, K.; Li, Z.; Zhang, Y.; Zhang, H.Y.; Li, B.; Zhu, W.L.; Shi, J.Y.; Jia, Q.; Li, Y.M. Advances in the study of berberine and its derivatives: A focus on anti-inflammatory and anti-tumor effects in the digestive system. Acta Pharmacol. Sin. 2017, 38, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; Wei, J.; Abidi, P.; Lin, M.; Inaba, S.; Li, C.; Wang, Y.; Wang, Z.; Si, S.; Pan, H.; et al. Berberine is a novel cholesterol-lowering drug working through a unique mechanism distinct from statins. Nat. Med. 2004, 10, 1344–1351. [Google Scholar] [CrossRef]
- Chai, F.N.; Ma, W.Y.; Zhang, J.; Xu, H.S.; Li, Y.F.; Zhou, Q.D.; Li, X.G.; Ye, X.L. Coptisine from Rhizoma coptidis exerts an anti-cancer effect on hepatocellular carcinoma by up-regulating miR-122. Biomed. Pharmacother. 2018, 103, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, H.; Hu, B.; Yang, L.; Wang, P.; Wang, F.; Meng, X. Coptisine from Coptis chinensis inhibits production of inflammatory mediators in lipopolysaccharide-stimulated RAW 264.7 murine macrophage cells. Eur. J. Pharmacol. 2016, 780, 106–114. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Ye, X.; Wu, H.; Wang, Y.; Zou, Z.; Ning, N.; Hu, Y.; Chen, B.; Fang, X.; Li, X. The safety and anti-hypercholesterolemic effect of coptisine in Syrian golden hamsters. Lipids 2015, 50, 185–194. [Google Scholar] [CrossRef]
- Shi, L.L.; Jia, W.H.; Zhang, L.; Xu, C.Y.; Chen, X.; Yin, L.; Wang, N.Q.; Fang, L.H.; Qiang, G.F.; Yang, X.Y.; et al. Glucose consumption assay discovers coptisine with beneficial effect on diabetic mice. Eur. J. Pharmacol. 2019, 859, 172523. [Google Scholar] [CrossRef]
- Ning, N.; He, K.; Wang, Y.; Zou, Z.; Wu, H.; Li, X.; Ye, X. Hypolipidemic effect and mechanism of palmatine from Coptis chinensis in hamsters fed high-fat diet. Phytother. Res. 2015, 29, 668–673. [Google Scholar] [CrossRef]
- Yan, B.; Wang, D.; Dong, S.; Cheng, Z.; Na, L.; Sang, M.; Yang, H.; Yang, Z.; Zhang, S.; Yan, Z. Palmatine inhibits TRIF-dependent NF-kappaB pathway against inflammation induced by LPS in goat endometrial epithelial cells. Int. Immunopharmacol. 2017, 45, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Tarabasz, D.; Kukula-Koch, W. Palmatine: A review of pharmacological properties and pharmacokinetics. Phytother. Res. 2020, 34, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Rong, J.; Tilton, R.J.; Ng, K.; Liu, C.; Tam, P.; Lau, A.; Cheng, Y. Genome-wide biological response fingerprinting (BioReF) of the Chinese botanical formulation ISF-1 enables the selection of multiple marker genes as a potential metric for quality control. J. Ethnopharmacol. 2007, 113, 35–44. [Google Scholar] [CrossRef]
- Santamaria, D.; Barriere, C.; Cerqueira, A.; Hunt, S.; Tardy, C.; Newton, K.; Caceres, J.F.; Dubus, P.; Malumbres, M.; Barbacid, M. Cdk1 is sufficient to drive the mammalian cell cycle. Nature 2007, 448, 811–815. [Google Scholar] [CrossRef]
- Takatori, H.; Yamashita, T.; Honda, M.; Nishino, R.; Arai, K.; Yamashita, T.; Takamura, H.; Ohta, T.; Zen, Y.; Kaneko, S. dUTP pyrophosphatase expression correlates with a poor prognosis in hepatocellular carcinoma. Liver Int. 2010, 30, 438–446. [Google Scholar] [CrossRef]
- Galmarini, C.M.; Cros, E.; Graham, K.; Thomas, X.; Mackey, J.R.; Dumontet, C. 5′-(3′)-nucleotidase mRNA levels in blast cells are a prognostic factor in acute myeloid leukemia patients treated with cytarabine. Haematologica 2004, 89, 617–619. [Google Scholar]
- Piper, M.; Gronostajski, R.; Messina, G. Nuclear Factor One X in development and disease. Trends Cell Biol. 2019, 29, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Kou, S.; Han, B.; Wang, Y.; Huang, T.; He, K.; Han, Y.; Zhou, X.; Ye, X.; Li, X. Synergetic cholesterol-lowering effects of main alkaloids from Rhizoma Coptidis in HepG2 cells and hypercholesterolemia hamsters. Life Sci. 2016, 151, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Qian, X.; Li, J.; Cui, X.; Chen, L.; Cai, B.; Tan, S. Simultaneous determination of 11 alkaloids in crude and wine-processed Rhizoma Coptidis by HPLC-PAD. J. Chromatogr. Sci. 2015, 53, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.; Ye, X.; Wang, D.; He, K.; Yang, Y.; Liu, X.; Li, X. Safety evaluation of main alkaloids from Rhizoma Coptidis. J. Ethnopharmacol. 2013, 145, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yang, J.; Chang, Y.; Qiao, L.; Dang, H.; Luo, K.; Guo, H.; An, Y.; Ma, C.; Shao, H.; et al. Caffeine-free hawk tea lowers cholesterol by reducing free cholesterol uptake and the production of very-low-density lipoprotein. Commun. Biol. 2019, 2, 173. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | HepG2 | RAW264.7 | 3T3-L1 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Time (h) | 24 | 48 | 72 | 24 | 48 | 72 | 24 | 48 | 72 |
| Berberine | N/A | 123.4 | 47.56 | 104.8 | 41.93 | 28.27 | 120.8 | 76.45 | 77.97 |
| Coptisine | 454.7 | 34.88 | 18.10 | 16.82 | 15.38 | 10.29 | 152.9 | 53.26 | 50.63 |
| Palmatine | N/A | N/A | 24.33 × 104 | 883.4 | 7292 | 29.16 × 103 | 1043 | N/A | N/A |
| Treatment | Total DEGs Number | Upregulated Gene Number | Downregulated Gene Number |
|---|---|---|---|
| RCE | 1273 | 585 | 691 |
| Berberine | 822 | 346 | 478 |
| Coptisine | 718 | 359 | 360 |
| Palmatine | 254 | 124 | 131 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, L.; Feng, S.; Zhang, X.; Zhao, W.; Feng, J.; Ma, C.; Wang, R.; Song, W.; Cheng, J. Biological Response Profiling Reveals the Functional Differences of Main Alkaloids in Rhizoma Coptidis. Molecules 2021, 26, 7389. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237389
Xie L, Feng S, Zhang X, Zhao W, Feng J, Ma C, Wang R, Song W, Cheng J. Biological Response Profiling Reveals the Functional Differences of Main Alkaloids in Rhizoma Coptidis. Molecules. 2021; 26(23):7389. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237389
Chicago/Turabian StyleXie, Lan, Shanshan Feng, Xiaoling Zhang, Wenlong Zhao, Juan Feng, Chengmei Ma, Ruijun Wang, Weifang Song, and Jing Cheng. 2021. "Biological Response Profiling Reveals the Functional Differences of Main Alkaloids in Rhizoma Coptidis" Molecules 26, no. 23: 7389. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237389