
Luteolin-3′-O-Phosphate Inhibits Lipopolysaccharide-Induced Inflammatory Responses by Regulating NF-κB/MAPK Cascade Signaling in RAW 264.7 Cells

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
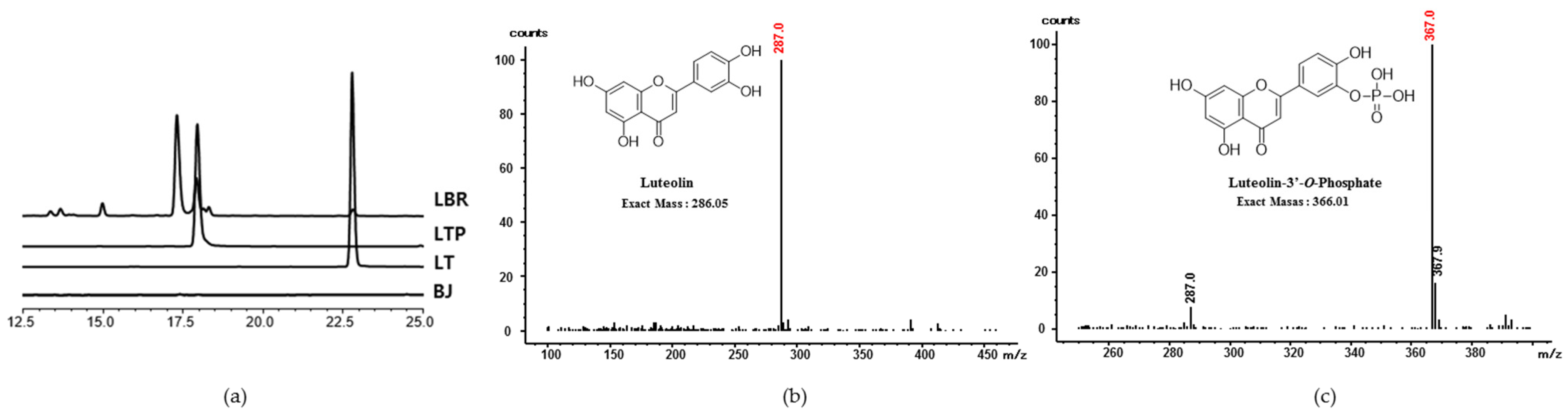
2.1. Analysis and Identification of LT Biorenovation Conversion Products
2.2. Determination of Position of Phosphoryation with NMR Results
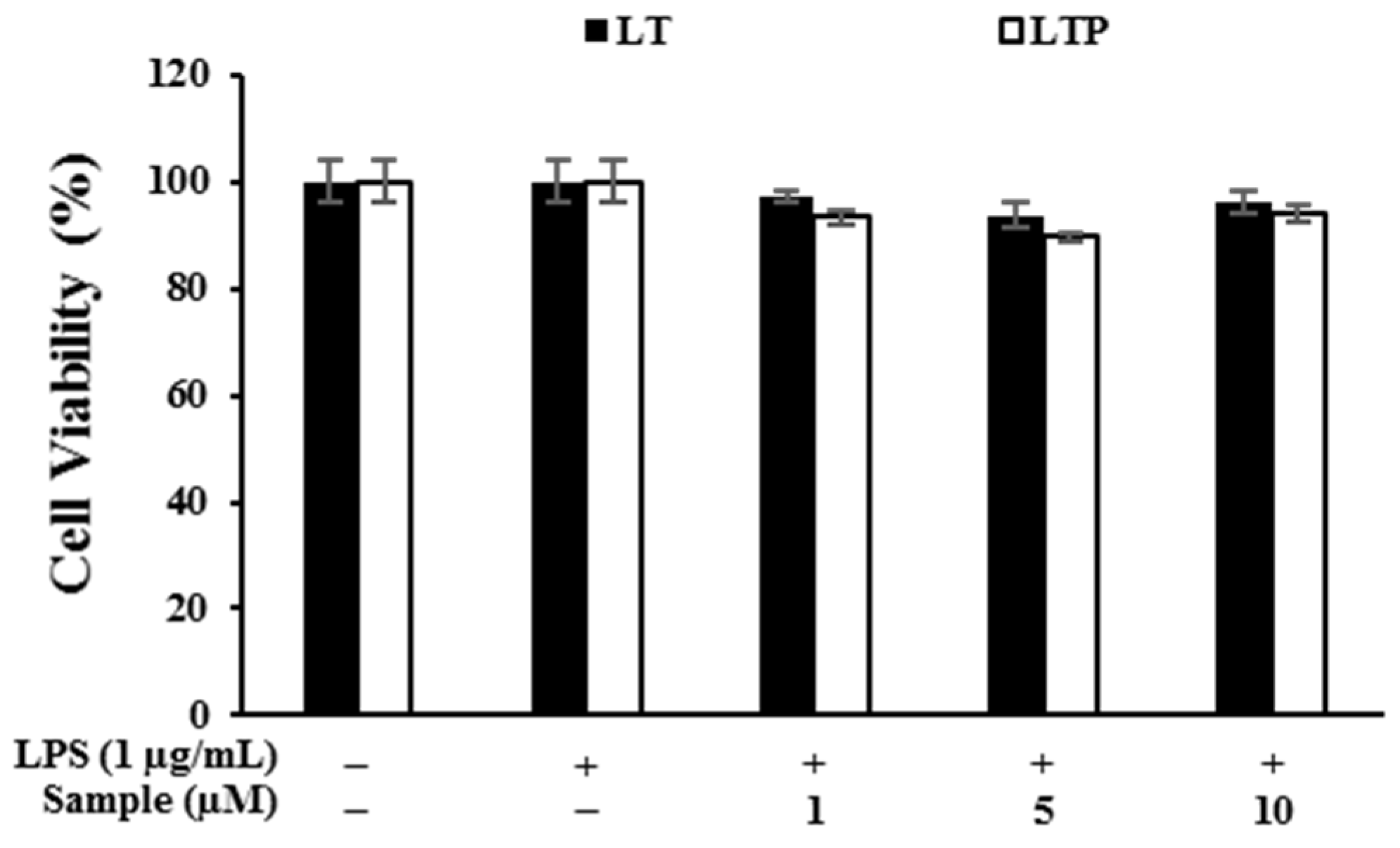
2.3. Cytotoxic Effects of Compounds on RAW 264.7 Cells
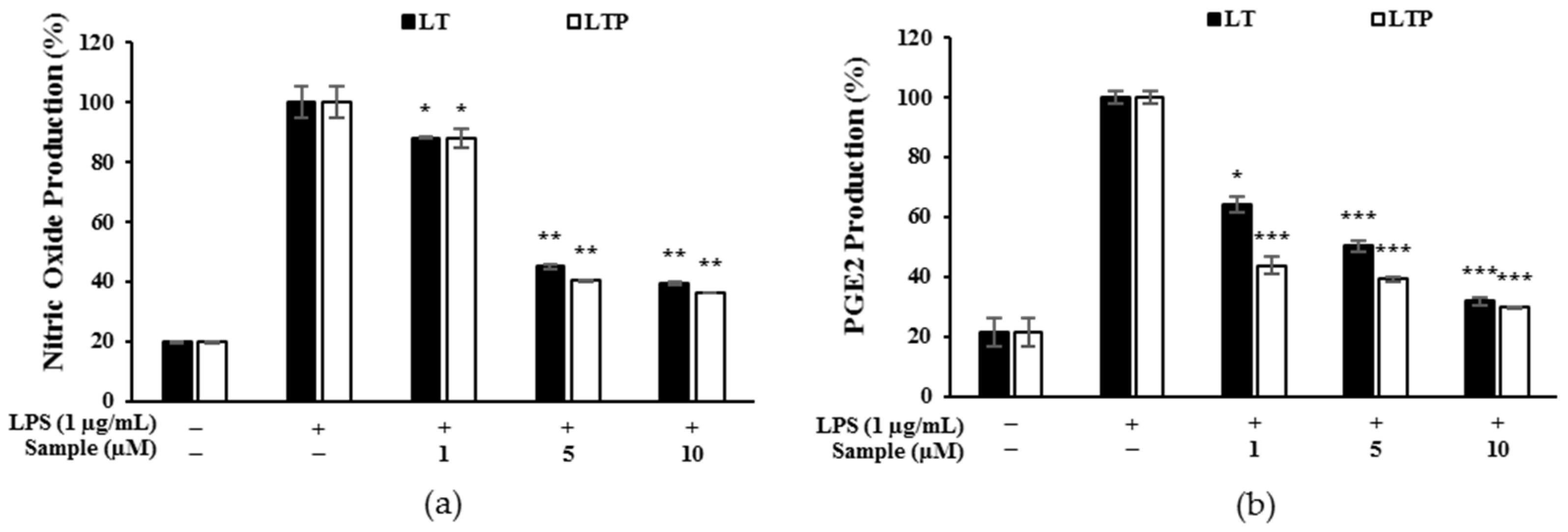
2.4. Production of NO and PGE2
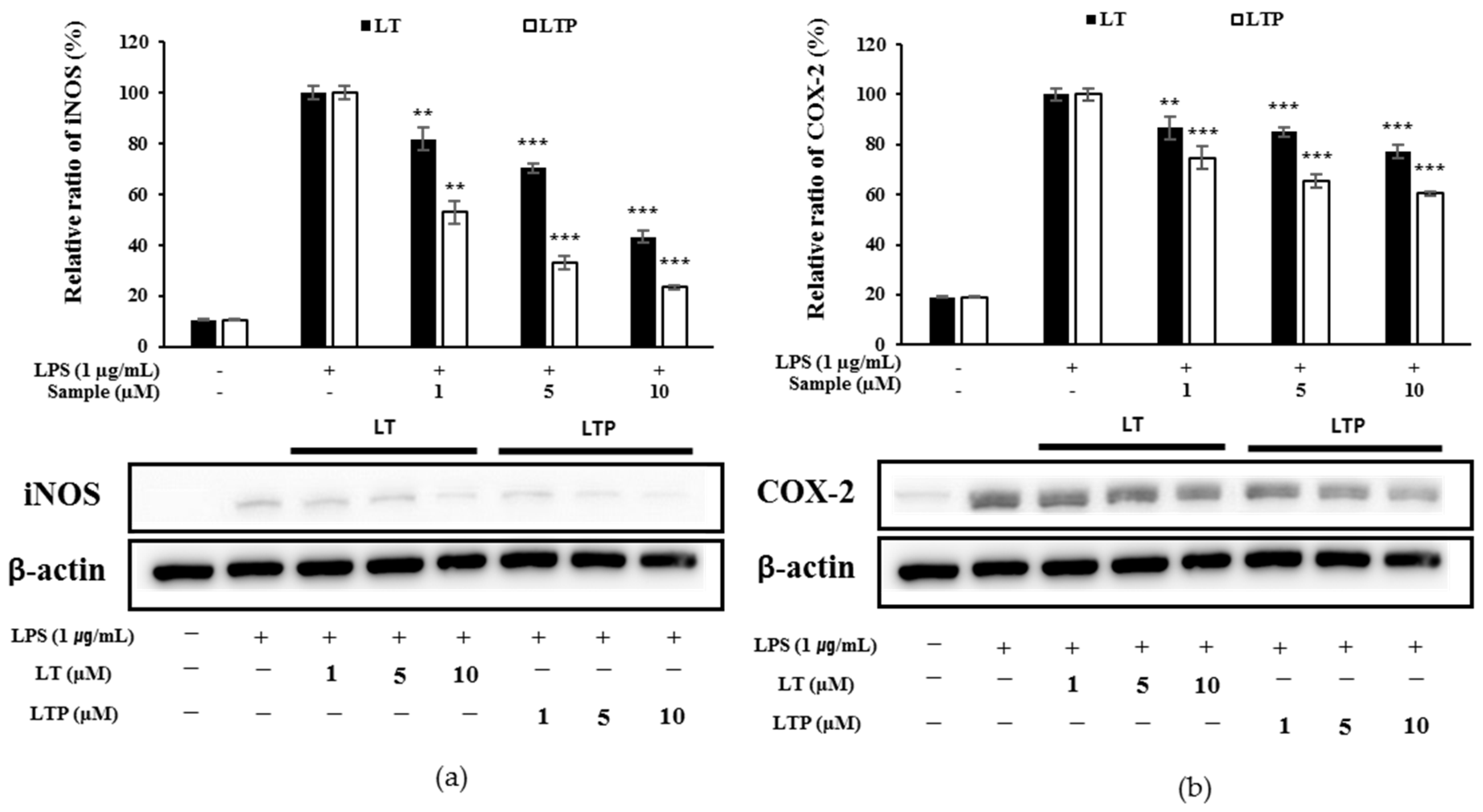
2.5. Comparison of iNOS and COX-2 Expression Inhibition
2.6. Effects of LT and LTP on LPS-Induced IL-1β, IL-6, and TNF-α Expression
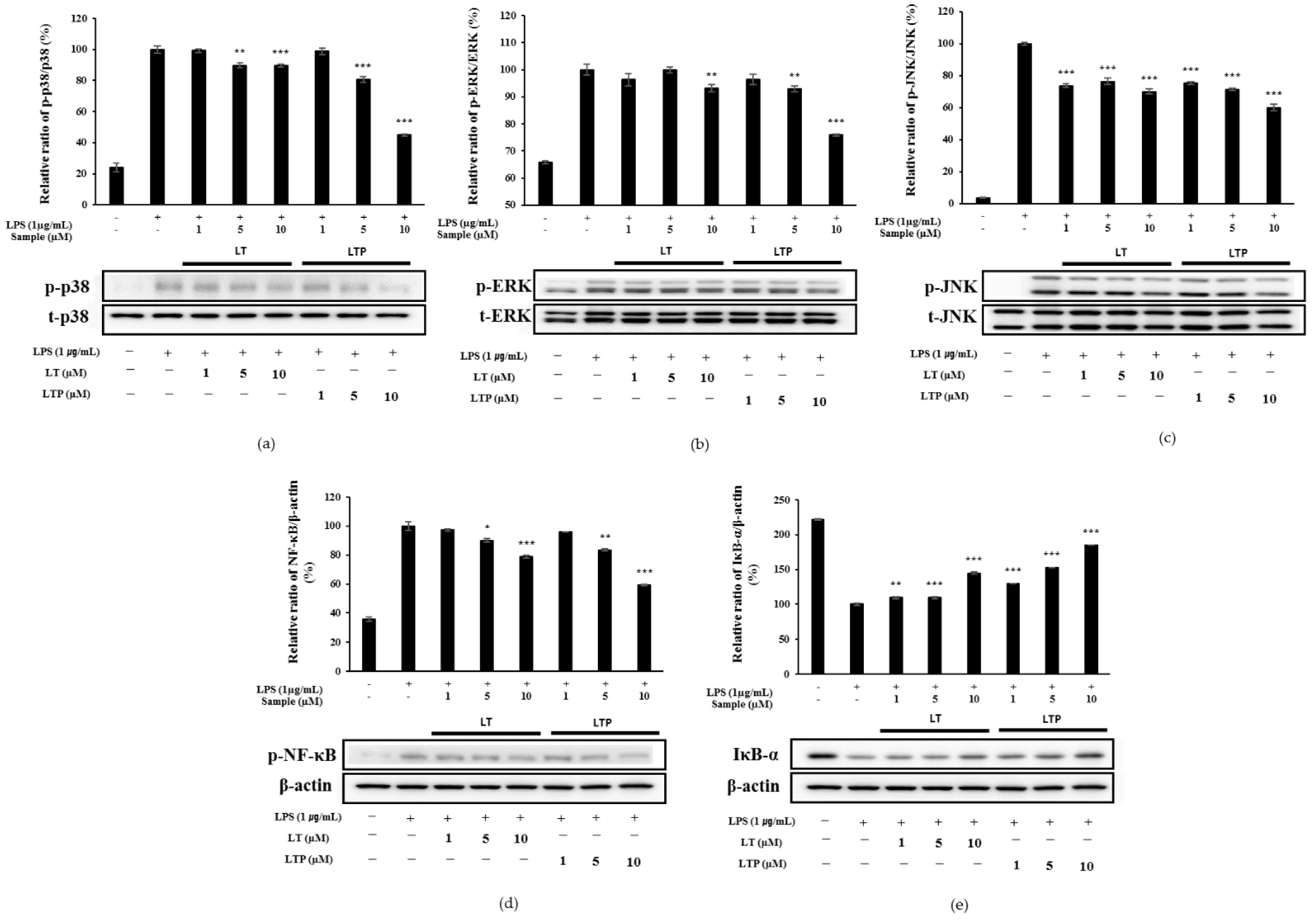
2.7. Effects of LT and LTP on the Activation of NF-κB and MAPK
3. Discussion
4. Materials and Methods
4.1. Reagents and Strains Used in Biorenovation
4.2. Biorenovation of LT
4.3. HPLC Analysis of the LT Biorenovation Product
4.4. Liquid Chromatography–Mass Spectrometry (LC–MS) and Nuclear Magnetic Resonance (NMR) of the LT Biorenovation Product
4.5. Cell Culture, Treatment, and Cell Viability Assay
4.6. Nitrite and Prostaglandin E2 (PGE2) Level Determination
4.7. Measurement of pro-Inflammatory Cytokine Production (TNF-α, IL-1β, and IL-6)
4.8. Western Blot Analyses
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Choi, H.J.; Eun, J.-S.; Park, Y.-R.; Kim, D.K.; Li, R.; Moon, W.S.; Park, J.M.; Kim, H.S.; Cho, N.-P.; Cho, S.-D.; et al. Ikarisoside A inhibits inducible nitric oxide synthase in lipopolysaccharide-stimulated RAW 264.7 cells via p38 kinase and nuclear factor-kappaB signaling pathways. Eur. J. Pharmacol. 2008, 601, 171–178. [Google Scholar] [CrossRef]
- Becker, S.; Mundandhara, S.; Devlin, R.B.; Madden, M.C. Regulation of cytokine production in human alveolar macrophages and airway epithelial cells in response to ambient air pollution particles: Further mechanistic studies. Toxicol. Appl. Pharmacol. 2005, 207, 269–275. [Google Scholar] [CrossRef]
- Moretta, A.; Marcenaro, E.; Sivori, S.; Della Chiesa, M.; Vitale, M.; Moretta, L. Early liaisons between cells of the innate immune system in inflamed peripheral tissues. Trends Immunol. 2005, 26, 668–675. [Google Scholar] [CrossRef]
- Lopes, A.J.O.; Vasconcelos, C.C.; Pereira, F.A.N.; Silva, R.H.M.; Queiroz, P.F.D.S.; Fernandes, C.V.; Garcia, J.B.S.; Ramos, R.; Da Rocha, C.Q.; Lima, S.T.D.J.R.M.; et al. Anti-Inflammatory and Antinociceptive Activity of Pollen Extract Collected by Stingless Bee Melipona fasciculata. Int. J. Mol. Sci. 2019, 20, 4512. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T.; Willoughby, D.A.; Gilroy, D.W. Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nat. Rev. Immunol. 2002, 2, 787–795. [Google Scholar] [CrossRef]
- Allavena, P.; Garlanda, C.; Borrello, M.G.; Sica, A.; Mantovani, A. Pathways connecting inflammation and cancer. Curr. Opin. Genet. Dev. 2008, 18, 3–10. [Google Scholar] [CrossRef]
- Luan, H.; Zhang, Q.; Wang, L.; Wang, C.; Zhang, M.; Xu, X.; Zhou, H.; Li, X.; Xu, Q.; He, F.; et al. OM85-BV induced the productions of IL-1β, IL-6, and TNF-α via TLR4- and TLR2-mediated ERK1/2/NF-κB Pathway in RAW264.7 Cells. J. Interferon Cytokine Res. 2014, 34, 526–536. [Google Scholar] [CrossRef]
- Sacco, R.E.; Waters, W.R.; Rudolph, K.M.; Drew, M.L. Comparative nitric oxide production by LPS-stimulated monocyte-derived macrophages from Ovis canadensis and Ovis aries. Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kaplanski, G.; Marin, V.; Montero-Julian, F.; Mantovani, A.; Farnarier, C. IL-6: A regulator of the transition from neutrophil to monocyte recruitment during inflammation. Trends Immunol. 2003, 24, 25–29. [Google Scholar] [CrossRef]
- Park, J.Y.; Pillinger, M.H.; Abramson, S.B. Prostaglandin E2 synthesis and secretion: The role of PGE2 synthases. Clin. Immunol. 2006, 199, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.S. Interleukins and tumor necrosis factor in inflammation. Crit. Rev. Clin. Lab. Sci. 1990, 28, 37–59. [Google Scholar] [CrossRef]
- Karin, M.; Ben-Neriah, Y. Phosphorylation meets ubiquitination: The control of NF-[kappa]B activity. Ann. Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef] [PubMed]
- Grilli, M.; Chiu, J.J.-S.; Lenardo, M.J. NF-kappa B and Rel: Participants in a Multiform Transcriptional Regulatory System. Int. Rev. Cytol. 1993, 143, 1–62. [Google Scholar] [CrossRef] [PubMed]
- Pateras, I.; Giaginis, C.; Tsigris, C.; Patsouris, E.; Theocharis, S. NF-κB signaling at the crossroads of inflammation and atherogenesis: Searching for new therapeutic links. Expert Opin. Ther. Targets 2014, 18, 1089–1101. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Pautz, A.; Art, J.; Hahn, S.; Nowag, S.; Voss, C.; Kleinert, H. Regulation of the expression of inducible nitric oxide synthase. Nitric Oxide 2010, 23, 75–93. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Kim, B.H.; Jang, J.O.; Lee, J.-H.; Park, Y.E.; Kim, J.-G.; Yoon, Y.-C.; Jeong, S.J.; Kwon, G.-S.; Lee, J.-B. Increased Anti-oxidative Activity and Whitening Effects of a Saposhnikovia Extract Following Bioconversion Fermentation using Lactobacillus plantarum BHN-LAB 33. J. Life Sci. 2019, 29, 1208–1217. [Google Scholar] [CrossRef]
- Choi, H.R.; Park, J.-S.; Kim, K.M.; Kim, M.S.; Ko, K.W.; Hyun, C.-G.; Ahn, J.W.; Seo, J.-H.; Kim, S.Y. Enhancing the antimicrobial effect of genistein by biotransformation in microbial system. Ind. Eng. Chem. 2018, 63, 255–261. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, T.J.; Sim, J.H.; Kim, S.Y. The Effects of Gleditsia japonica Extract Following Biorenovation on Melanogenesis-related genes in B16F10 Melanoma Cells. KSBB J. 2020, 35, 303–309. [Google Scholar] [CrossRef]
- Kim, M.S.; Park, T.J.; Lim, J.S.; Kim, S.Y. Effect of Agaricus biorenovate Extract on Collagen Synthesis and Matrix Metalloproteinase-1 Production in Human Dermal Fibroblast. KSBB J. 2019, 34, 49–53. [Google Scholar] [CrossRef]
- Middleton, E.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- García-Lafuente, A.; Guillamón, E.; Villares, A.; Rostagno, M.A.; Martínez, J.A. Flavonoids as anti-inflammatory agents: Implications in cancer and cardiovascular disease. Inflamm. Res. 2009, 58, 537–552. [Google Scholar] [CrossRef]
- Wang, S.; Cao, M.; Xu, S.; Shi, J.; Mao, X.; Yao, X.; Liu, C. Luteolin alters macrophage polarization to inhibit inflammation. Inflammation 2020, 43, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Farzaei, M.H.; Abbasabadi, Z.; Ardekani, M.R.S.; Rahimi, R.; Farzaei, F. Parsley: A review of ethnopharmacology, phytochemistry and biological activities. J. Tradit. Chin. Med. 2013, 33, 815–826. [Google Scholar] [CrossRef] [Green Version]
- Katiyar, C.; Gupta, A.; Kanjilal, S.; Katiyar, S. Drug discovery from plant sources: An integrated approach. Ayu 2012, 33, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.H.; Park, T.J.; Kang, M.S.; Kim, S.Y. Anti-inflammatory Activity of Beta vulgaris Extract Using Biorenovation in LPS-stimulated RAW 264.7 Cells. Korean Soc. Biotechnol. Bioeng. J. 2021, 36, 123–129. [Google Scholar] [CrossRef]
- Park, T.J.; Sim, J.H.; Hong, H.H.; Han, D.H.; Kim, S.Y. Anti-inflammatory Effect of Colocasia esculenta Biorenovate Extract in LPS-stimulated RAW 264.7 Cells. Korean Soc. Biotechnol. Bioeng. J. 2020, 35, 162–168. [Google Scholar] [CrossRef]
- Park, C.M.; Song, Y.S. Luteolin and luteolin-7-O-glucoside inhibit lipopolysaccharide-induced inflammatory responses through modulation of NF-κB/AP-1/PI3K-Akt signaling cascades in RAW 264.7 cells. Nutr. Res. Pract. 2013, 7, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, Y.; Lei, J.C.; Hao, Y.; Yang, Y.; Yang, C.X.; Yu, J.Q. Sensitisation of ovarian cancer cells to cisplatin by flavonoids from Scutellaria barbata. Nat. Prod. Res. 2014, 10, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.K.; Sung, S.H.; Kim, Y.C.; Kim, S.G. Inhibition of lipopolysaccharide-inducible nitric oxide synthase, TNF-α and COX-2 expression by sauchinone effects on I-κBα phosphorylation, C/EBP and AP-1 activation. Br. J. Pharmacol. 2003, 139, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.Y.; Peng, W.H.; Tsai, K.D.; Hsu, S.L. Luteolin suppresses inflammation-associated gene expression by blocking NF-kappaB and AP-1 activation pathway in mouse alveolar macrophages. Life Sci. 2007, 81, 1602–1614. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Shin, T.Y. Anti-inflammatory effect of leaves of Eriobotrya japonica correlating with attenuation of p38 MAPK, ERK, and NF-kappaB activation in mast cells. Toxicol. In Vitro 2007, 23, 1215–1219. [Google Scholar] [CrossRef]
- Israf, D.A.; Khaizurin, T.A.; Lajis, N.H.; Khozirah, S. Cardamonin inhibits COX and iNOS expression via inhibition of p65NF-kappaB nuclear translocation and Ikappa-B phosphorylation in RAW264.7 macrophage cells. Mol. Immunol. 2006, 44, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, I.T.; Won, J.H.; Jeong, S.H.; Park, E.Y.; Nam, J.H.; Choi, J.W.; Lee, K.T. Anti-inflammatory activities of ent-16alphaH, 17-hydroxy-kauran-19-oic acid isolated from the roots of Siegesbeckia pubescens are due to the inhibition of iNOS and COX-2 expression in RAW 264.7 macrophages via NF-kappaB inactivation. Eur. J. Pharmacol. 2007, 558, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Lee, E.J.; Lee, J.C.; Kim, W.K.; Kim, H.S. Anti-inflammatory effects of short chain fatty acids in IFN-gamma-stimulated RAW 264.7 murine macrophage cells: Involvement of NF-kappaB and ERK signaling pathways. Int. Immunopharmacol. 2007, 7, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.H.; Lin-Shiau, S.Y.; Lin, J.K. Suppression of nitric oxide synthase and the down-regulation of the activation of NF-kappaB in macrophages by resveratrol. Br. J. Pharmacol. 1999, 126, 673–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Z.; Dickens, M.; Raingeaud, J.; Davis, R.J.; Greenberg, M.E. Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 1995, 24, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Park, C.M.; Park, J.Y.; Noh, K.H.; Shin, J.H.; Song, Y.S. Taraxacum officinale Weber extracts inhibit LPS-induced oxidative stress and nitric oxide production via the NF-κB modulation in RAW 264.7 cells. J. Ethnopharmacol. 2011, 133, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Park, C.M.; Jin, K.S.; Lee, Y.W.; Song, Y.S. Luteolin and chicoric acid synergistically inhibited inflammatory responses via inactivation of PI3K-Akt pathway and impairment of NF-κB translocation in LPS stimulated RAW 264.7 cells. Eur. J. Pharmacol. 2011, 660, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.P.; Li, Y.C.; Chen, H.Y.; Lin, R.H.; Huang, S.S.; Chen, H.L.; Kuan, P.C.; Liao, M.F.; Chen, C.J.; Kuan, Y.H. Protective effects of luteolin against lipopolysaccharide-induced acute lung injury involves inhibition of MEK/ERK and PI3K/Akt pathways in neutrophils. Acta Pharmacol. Sin. 2010, 31, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-H.; Park, T.-J.; Park, J.-S.; Kim, M.-S.; Chi, W.-J.; Kim, S.-Y. Luteolin-3′-O-Phosphate Inhibits Lipopolysaccharide-Induced Inflammatory Responses by Regulating NF-κB/MAPK Cascade Signaling in RAW 264.7 Cells. Molecules 2021, 26, 7393. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237393
Kim J-H, Park T-J, Park J-S, Kim M-S, Chi W-J, Kim S-Y. Luteolin-3′-O-Phosphate Inhibits Lipopolysaccharide-Induced Inflammatory Responses by Regulating NF-κB/MAPK Cascade Signaling in RAW 264.7 Cells. Molecules. 2021; 26(23):7393. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237393
Chicago/Turabian StyleKim, Jung-Hwan, Tae-Jin Park, Jin-Soo Park, Min-Seon Kim, Won-Jae Chi, and Seung-Young Kim. 2021. "Luteolin-3′-O-Phosphate Inhibits Lipopolysaccharide-Induced Inflammatory Responses by Regulating NF-κB/MAPK Cascade Signaling in RAW 264.7 Cells" Molecules 26, no. 23: 7393. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237393