
Essential Oils as a Potential Neuroprotective Remedy for Age-Related Neurodegenerative Diseases: A Review



Abstract
:1. Introduction
2. Materials and Methods
Search Strategy
3. Results
4. Discussion
4.1. The Source of EOs
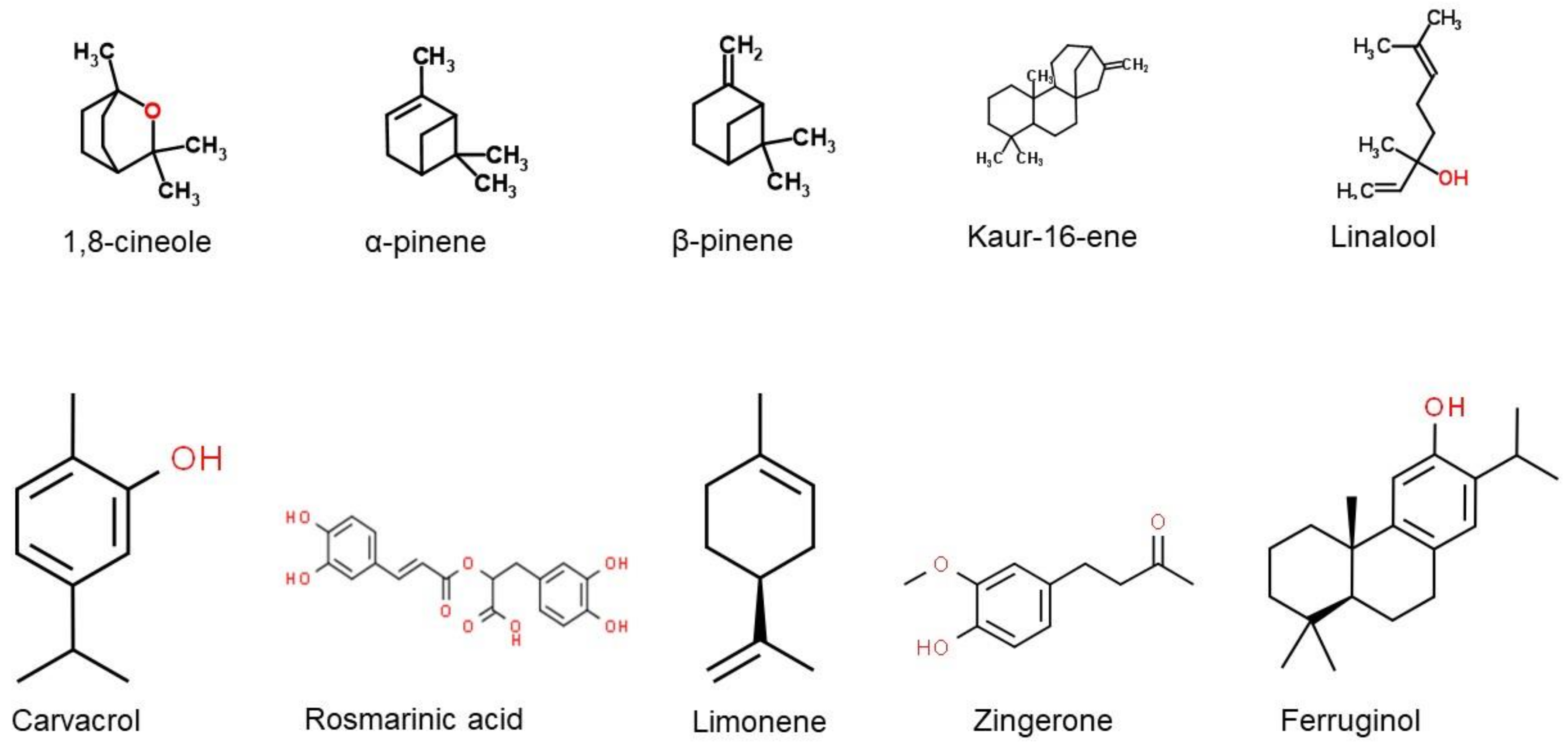
4.2. Major Component of EOs
4.3. Cholinesterase Activity in EOs
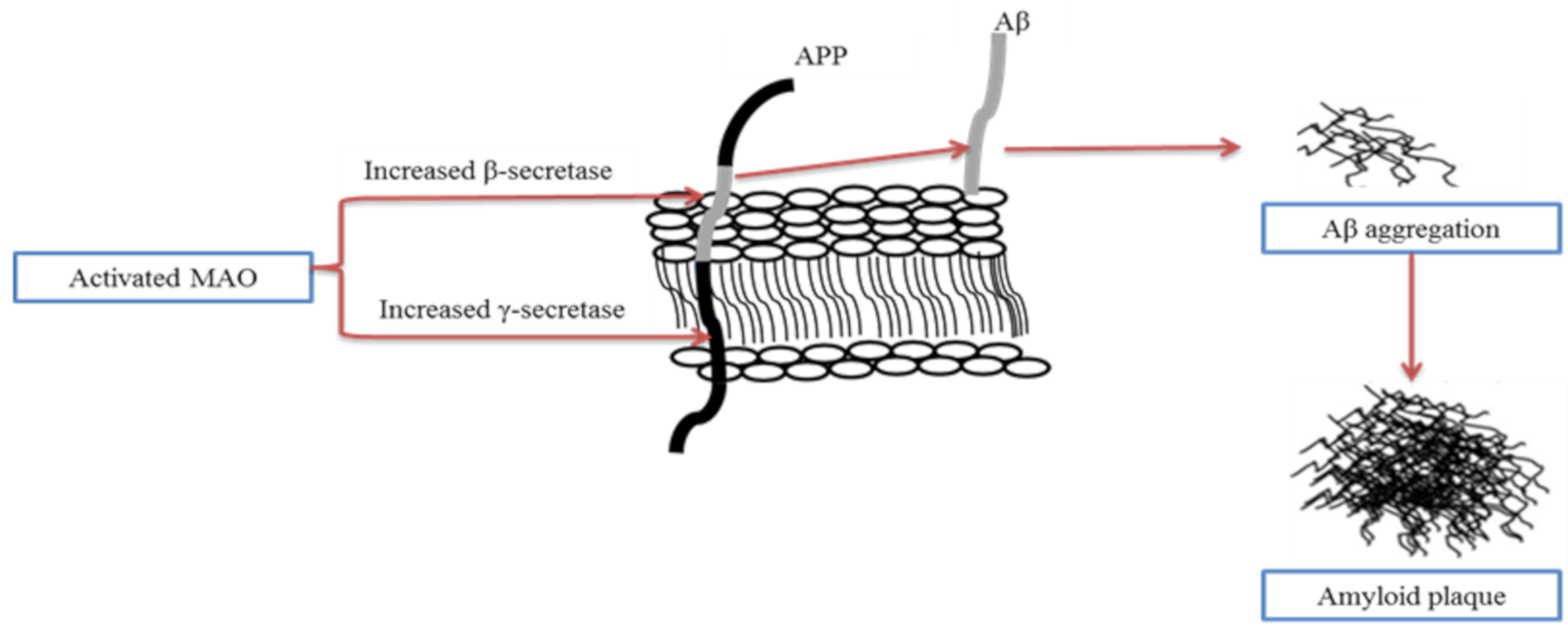
4.4. Extracellular Plaque Deposits
4.5. Current EOs on AD
4.6. Current EOs on PD
4.7. Current EOs on Other Neurodegenerative Diseases
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
List of Abbreviations
| Aβ | Amyloid-beta |
| ACh | Acetylcholine |
| AChE | Acetylcholinesterase |
| AD | Alzheimer’s disease |
| AFM | Atomic force microscope |
| ALS | Amyotrophic lateral sclerosis |
| AOPP | Advanced oxidation protein product |
| APP | Amyloid precursor protein |
| Asc | l-Ascorbate |
| α-Syn | α-Synuclein |
| ATF6 | Activating transcription factor 6 |
| BACE-1 | Beta-site APP-cleaving enzyme 1 |
| BCh | Butyrylcholine |
| BChE | Butyrylcholinesterase |
| βCD | β-cyclodextrin |
| BDNF | Brain-derived neurotropic factor |
| CA1 | Cornu ammonis |
| CAG | Cytosine–adenine–guanine |
| CAL | (E)-cinnamaldehyde |
| CAS | (E)-cinnamyl acetate |
| CAT | Catalase |
| CGC | Caenorhabditis Genetics Center |
| ChE | Cholinesterases |
| CNS | Central nervous system (human) |
| COMT | Catechol O-methyltransferase |
| COX-2 | Cyclooxygenase-2 |
| CUMS | Chronic unpredictable mild stress |
| DA | Dopamine |
| DAT | Dark avoidance test |
| DOPAC DSPE-PEG2000 | 3,4-dihydroxyphenylacetic acid Distearoylphosphatidylethanolamine with covalently linked polyethylene glycol of molecular weight 2000 |
| DTNB ELISA | 5,5′-dithiobis-(2-nitrobenzoic acid) Enzyme-linked immunosorbent assay |
| ER | Endoplasmic reticulum |
| EO | Essential oil |
| fPD FITC | Familial PD Fluorescein isothiocyanate |
| GABA | γ-aminobutyric acid |
| GC–FID | Gas chromatography–flame ionisation detector |
| GC–MS GC–ITMS | Gas chromatography–mass spectrometry Gas Chromatography/Ion Trap Mass Spectrometry |
| GC–O | Gas chromatography–olfactometry |
| GPx | Gluthathione peroxidase |
| GRP78 | Glucose regulated protein 78 |
| GSH | Reduced glutathione |
| HD HPLC–DAD HPLC–PDA | Huntington’s disease High-performance liquid chromatography–diode-array detection High-performance liquid chromatography–photodiode array detection |
| HTT | Huntingtin gene |
| HVA | Homovanillic acid |
| iNOS | Induced nitric oxide synthase |
| IRE1 | Inositol requiring enzyme 1 |
| IRE1/XBP1 | IRE1/X-Box Binding Protein 1 |
| JNK | c-Jun N-terminal kinase |
| LDH | Lactate dehydrogenase |
| L-dopa | Levodopa |
| LMN LOX | Lower motor neuron Lipoxygenase |
| MAO | Monoamine oxidase |
| MDA | Malondialdehyde |
| ME | Microemulsion |
| mHTT | Mutant HTT |
| MMP | Mitochondrial membrane potential |
| MPP+ | 1-methyl-4-phenylpyridinium |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| MWM NA–FB | Morris water maze test New micro-well plate AChE activity |
| NHPs NMR | Non-human primates Nuclear magnetic resonance |
| NO OS | Nitric oxide Oxidative stress |
| PCC | Protein carbonyl content |
| PCh | Propionylcholine |
| PD | Parkinson’s disease |
| PDI | Polydispersity index |
| PERK | Protein kinase RNA (PKR)-like ER kinase |
| p-PERK | PERK phosphorylation |
| PI3K | Phosphatidylinositol-4,5-bisphosphate 3-kinase |
| PKR | Protein kinase RNA |
| PS | Presenilin |
| RNAi | RNA interference |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| SDT | Step-down test |
| SHXW SFE SKN-1 SN | Su He Xiang Wan Supercritical fluid extraction Skinhead-1 Substantia nigra |
| SNpc | Substantia nigra pars compacta |
| SOD | Superoxide dismutase |
| sPD | Sporadic PD |
| TBARS TLC | Thiobarbituric acid-reactive substances Thin-layer chromatography |
| TNB | 5-thio-2-nitrobenzoic acid |
| TNF TTC TUNEL | Tumor necrosis factor 2,3,5-triphenyltetrazolium chloride Terminal deoxynucleotidyl transferase dUTP nick end labeling |
| UMN | Upper motor neuron |
| UPR) | Unfolded protein response |
| VMAT-2 | Vesicular monoamine transporter 2 |
| VOCs WST | Volatile organic constituents Water-soluble tetrazolium salt |
| Z-LIG | Z-ligustilide |
| 5-HT | Serotonin |
| 6-OHDA | 6-hydroxydopamine |
| 8-OHdG | Urinary 8-hydroxyguanosine |
References
- Osbourn, A.E.; Lanzotti, V. Plant-Derived Natural Products: Synthesis, Function, and Application; Springer Science & Business Media: New York, NY, USA, 2009; p. 612. [Google Scholar]
- Esposito, E.R.; Bystrek, M.V.; Klein, J.S. An Elective Course in Aromatherapy Science. Am. J. Pharm. Edu. 2014, 78, 79. [Google Scholar] [CrossRef] [Green Version]
- Robins, J.L.W. The Science and Art of Aromatherapy. J. Holist. Nurs. 1999, 17, 5–17. [Google Scholar] [CrossRef]
- Smith, C.A.; Collins, C.T.; Crowther, C.A. Aromatherapy for pain management in labour. Cochrane Database Syst. Rev. 2011. [Google Scholar] [CrossRef]
- Garcia-Salas, P.; Morales-Soto, A.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Phenolic-compound-extraction systems for fruit and vegetable samples. Molecules 2010, 15, 8813–8826. [Google Scholar] [CrossRef]
- Singer, A.C.; Crowley, D.E.; Thompson, I.P. Secondary plant metabolites in phytoremediation and biotransformation. Trends Biotechnol. 2003, 21, 123–130. [Google Scholar] [CrossRef]
- Hsieh, M.T.; Peng, W.H.; Wu, C.R.; Wang, W.H. The ameliorating effects of the cognitive-enhancing Chinese herbs on scopolamine-induced amnesia in rats. Phytother. Res. 2000, 14, 375–377. [Google Scholar] [CrossRef]
- Cioanca, O.; Mircea, C.; Hritcu, L.; Trifan, A.; Marius, M.; Aprotosoaie, A.C.; Robu, S.; Gille, E.; Hancianu, M. In vitro—In vivo correlation of the antioxidant capacity of Salviae aetheroleum essential oil. Farmacia 2015, 63, 34–39. [Google Scholar]
- Jové, M.; Portero-Otín, M.; Naudí, A.; Ferrer, I.; Pamplona, R. Metabolomics of Human Brain Aging and Age-Related Neurodegenerative Diseases. J. Neuropath. Exp. Neur. 2014, 73, 640–657. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.H.; Wang, X.; Zhu, X. Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free Radic. Biol. Med. 2013, 62, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Sarrafchi, A.; Bahmani, M.; Shirzad, H.; Rafieian-Kopaei, M. Oxidative stress and Parkinson’s disease: New hopes in treatment with herbal antioxidants. Curr. Pharm. Des. 2016, 22, 238–246. [Google Scholar] [CrossRef] [Green Version]
- González-Burgos, E.; Gómez-Serranillos, M.P. Terpene compounds in nature: A review of their potential antioxidant activity. Curr. Med. Chem. 2012, 19, 5319–5341. [Google Scholar] [CrossRef] [PubMed]
- Essa, M.; Braidy, N.; Bridge, W.; Subash, S.; Manivasagam, T.; Vijayan, R.; Al-Adawi, S.; Guillemin, G. Review of natural products on Parkinson’s disease pathology. J. Aging Res. Clin. Pract 2014, 3, 1–8. [Google Scholar]
- Karakaya, S.; Koca, M.; Yılmaz, S.V.; Yıldırım, K.; Pınar, N.M.; Demirci, B.; Brestic, M.; Sytar, O. Molecular Docking Studies of Coumarins Isolated from Extracts and Essential Oils of Zosima absinthifolia Link as Potential Inhibitors for Alzheimer’s Disease. Molecules 2019, 24, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrester, L.T.; Maayan, N.; Orrell, M.; Spector, A.E.; Buchan, L.D.; Soares-Weiser, K. Aromatherapy for dementia. Cochrane Database Syst. Rev. 2014, CD003150, 1–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Wang, K.; Lu, C.; Dong, L.; Gao, L.; Yan, M.; Aibai, S.; Yang, Y.; Liu, X. Protective effects of linalool against amyloid beta-induced cognitive deficits and damages in mice. Life Sci. 2017, 174, 21–27. [Google Scholar] [CrossRef]
- Bansal, A.; Kirschner, M.; Zu, L.; Cai, D.; Zhang, L. Coconut oil decreases expression of amyloid precursor protein (APP) and secretion of amyloid peptides through inhibition of ADP-ribosylation factor 1 (ARF1). Brain Res. 2019, 1704, 78–84. [Google Scholar] [CrossRef]
- Dalai, M.; Bhadra, S.; Chaudhary, S.; Bandyopadhyay, A.; Mukherjee, P. Anti-cholinesterase potential of Cinnamomum tamala (Buch.-Ham.) T.Nees & Eberm. leaves. Indian J. Tradit. Know. 2014, 13, 691–697. [Google Scholar]
- Sihoglu Tepe, A.; Ozaslan, M. Anti-Alzheimer, anti-diabetic, skin-whitening, and antioxidant activities of the essential oil of Cinnamomum zeylanicum. Ind. Crop. Prod. 2020, 145, 112069. [Google Scholar] [CrossRef]
- Bahadori, M.B.; Salehi, P.; Sonboli, A. Comparative study of the essential oil composition of Salvia urmiensis and its enzyme inhibitory activities linked to diabetes mellitus and Alzheimer’s disease. Int. J. Food Prop. 2017, 20, 2974–2981. [Google Scholar] [CrossRef] [Green Version]
- Cutillas, A.B.; Carrasco, A.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Salvia officinalis L. Essential Oils from Spain: Determination of Composition, Antioxidant Capacity, Antienzymatic, and Antimicrobial Bioactivities. Chem. Biodivers. 2017; 14. [Google Scholar] [CrossRef]
- Ayaz, M.; Junaid, M.; Ullah, F.; Sadiq, A.; Khan, M.A.; Ahmad, W.; Shah, M.R.; Imran, M.; Ahmad, S. Comparative chemical profiling, cholinesterase inhibitions and anti-radicals properties of essential oils from Polygonum hydropiper L: A Preliminary anti- Alzheimer’s study. Lipids Health Dis. 2015, 14, 141. [Google Scholar] [CrossRef] [Green Version]
- Ali-Shtayeh, M.S.; Abu-Zaitoun, S.Y.; Dudai, N.; Jamous, R.M. Downy Lavender Oil: A Promising Source of Antimicrobial, Antiobesity, and Anti-Alzheimer’s Disease Agents. Evid-Based Complement. Altern. Med. 2020, 2020, 5679408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soheili, M.; Khalaji, F.; Mirhashemi, M.; Salami, M. The Effect of Essential Oil of Lavandula Angustifolia on Amyloid Beta Polymerization: An In Vitro Study. Iran. J. Chem. Chem. Eng. 2018, 37, 201–207. [Google Scholar]
- Chaiyana, W.; Okonogi, S. Inhibition of cholinesterase by essential oil from food plant. Phytomedicine 2012, 19, 836–839. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, M.; Shah, I.; Ali, N.; Ali, W. In vitro acetylcholinesterase and butyrylcholinesterase inhibitory potentials of essential oil of Artemisia macrocephala. Bangladesh J. Pharm. 2015, 10, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.; Eslami, H. Preparation and study of the inhibitory effect of nano-niosomes containing essential oil from artemisia absinthium on amyloid fibril formation. Nanomed. J. 2020, 7, 243–250. [Google Scholar] [CrossRef]
- Kabuto, H.; Yamanushi, T.T. Effects of zingerone [4-(4-hydroxy-3-methoxyphenyl)-2-butanone] and eugenol [2-methoxy-4-(2-propenyl) phenol] on the pathological progress in the 6-hydroxydopamine-induced parkinson’s disease mouse model. Neurochem. Res. 2011, 36, 2244. [Google Scholar] [CrossRef]
- Alzheimer’s Association. 2019 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2019, 15, 321–387. [Google Scholar] [CrossRef]
- Reitz, C.; Mayeux, R. Alzheimer disease: Epidemiology, diagnostic criteria, risk factors and biomarkers. Biochem. Pharm. 2014, 88, 640–651. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, C.; Sarti, F.M.; Nitrini, R.; Ferreira, F.F.; Brucki, S.M.D. An assessment of direct and indirect costs of dementia in Brazil. PLoS ONE 2018, 13, e0193209. [Google Scholar] [CrossRef]
- Weintraub, S.; Wicklund, A.H.; Salmon, D.P. The neuropsychological profile of Alzheimer disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006171. [Google Scholar] [CrossRef]
- Aliev, G.; Palacios, H.H.; Walrafen, B.; Lipsitt, A.E.; Obrenovich, M.E.; Morales, L. Brain mitochondria as a primary target in the development of treatment strategies for Alzheimer disease. Int. J. Biochem. Cell Biol. 2009, 41, 1989–2004. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Pozo, A.; Mielke, M.L.; Gómez-Isla, T.; Betensky, R.A.; Growdon, J.H.; Frosch, M.P.; Hyman, B.T. Reactive glia not only associates with plaques but also parallels tangles in Alzheimer’s disease. Am. J. Pathol. 2011, 179, 1373–1384. [Google Scholar] [CrossRef] [PubMed]
- Villa, V.; Thellung, S.; Bajetto, A.; Gatta, E.; Robello, M.; Novelli, F.; Tasso, B.; Tonelli, M.; Florio, T. Novel celecoxib analogues inhibit glial production of prostaglandin E2, nitric oxide, and oxygen radicals reverting the neuroinflammatory responses induced by misfolded prion protein fragment 90–231 or lipopolysaccharide. Pharm. Res. 2016, 113, 500–514. [Google Scholar] [CrossRef] [PubMed]
- Masters, C.L.; Selkoe, D.J. Biochemistry of amyloid β-protein and amyloid deposits in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006262. [Google Scholar] [CrossRef] [PubMed]
- Nhan, H.S.; Chiang, K.; Koo, E.H. The multifaceted nature of amyloid precursor protein and its proteolytic fragments: Friends and foes. Acta. Neuropathol. 2015, 129, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campora, M.; Francesconi, V.; Schenone, S.; Tasso, B.; Tonelli, M. Journey on Naphthoquinone and Anthraquinone Derivatives: New Insights in Alzheimer’s Disease. Pharmaceuticals 2021, 14, 33. [Google Scholar] [CrossRef]
- Kocahan, S.; Doğan, Z. Mechanisms of Alzheimer’s Disease Pathogenesis and Prevention: The Brain, Neural Pathology, N-methyl-D-aspartate Receptors, Tau Protein and Other Risk Factors. Clin. Psychopharmacol. Neurosci. 2017, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Fridkin, M.; Youdim, M.B. From antioxidant chelators to site-activated multi-target chelators targeting hypoxia inducing factor, beta-amyloid, acetylcholinesterase and monoamine oxidase A/B. Mini Rev. Med. Chem. 2012, 12, 364–370. [Google Scholar] [CrossRef]
- Huang, L.; Lu, C.; Sun, Y.; Mao, F.; Luo, Z.; Su, T.; Jiang, H.; Shan, W.; Li, X. Multitarget-directed benzylideneindanone derivatives: Anti-β-amyloid (Aβ) aggregation, antioxidant, metal chelation, and monoamine oxidase B (MAO-B) inhibition properties against Alzheimer’s disease. J. Med. Chem. 2012, 55, 8483–8492. [Google Scholar] [CrossRef]
- Youdim, M.B.H.; Amit, T.; Bar-Am, O.; Weinreb, O.; Yogev-Falach, M. Implications of co-morbidity for etiology and treatment of neurodegenerative diseases with multifunctional neuroprotective-neurorescue drugs; ladostigil. Neurotox. Res. 2006, 10, 181–192. [Google Scholar] [CrossRef]
- Bar-Am, O.; Amit, T.; Weinreb, O.; Youdim, M.B.; Mandel, S. Propargylamine containing compounds as modulators of proteolytic cleavage of amyloid-beta protein precursor: Involvement of MAPK and PKC activation. J. Alzheimers Dis. 2010, 21, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Yogev-Falach, M.; Bar-Am, O.; Amit, T.; Weinreb, O.; Youdim, M.B. A multifunctional, neuroprotective drug, ladostigil (TV3326), regulates holo-APP translation and processing. FASEB J. 2006, 20, 2177–2179. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, O.; Amit, T.; Bar-Am, O.; Sagi, Y.; Mandel, S.; Youdim, M.B.H. Involvement of multiple survival signal transduction pathways in the neuroprotective, neurorescue and APP processing activity of rasagiline and its propargyl moiety. In Parkinson’s Disease and Related Disorders, 2nd ed.; Springer: Vienna, Austria, 2006; Volume 70, pp. 457–465. [Google Scholar]
- Cai, Z. Monoamine oxidase inhibitors: Promising therapeutic agents for Alzheimer’s disease (Review). Mol. Med. Rep. 2014, 9, 1533–1541. [Google Scholar] [CrossRef] [Green Version]
- Manouchehrabadi, M.; Farhadi, M.; Azizi, Z.; Torkaman-Boutorabi, A. Carvacrol Protects Against 6-Hydroxydopamine-Induced Neurotoxicity in In Vivo and In Vitro Models of Parkinson’s Disease. Neurotox Res. 2020, 37, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Fahn, S. Description of Parkinson’s disease as a clinical syndrome. Ann. N. Y. Acad. Sci. 2003, 991, 1–14. [Google Scholar] [CrossRef]
- Goedert, M. Alpha-synuclein and neurodegenerative diseases. Nat. Rev. Neurosci. 2001, 2, 492–501. [Google Scholar] [CrossRef]
- Jellinger, K.A. The pathology of Parkinson’s disease. Adv. Neurol. 2001, 86, 55–72. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Crowther, R.A.; Jakes, R.; Hasegawa, M.; Goedert, M. α-Synuclein in filamentous inclusions of Lewy bodies from Parkinson’s disease and dementia with Lewy bodies. Proc. Natl. Acad. Sci. USA 1998, 95, 6469–6473. [Google Scholar] [CrossRef] [Green Version]
- Schulz, J.B. Update on the pathogenesis of Parkinson’s disease. J. Neurol. 2008, 255, 3–7. [Google Scholar] [CrossRef]
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.; Benabid, A.L.; Sadoul, R.; Verna, J.M. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson’s disease. Prog. Neurobiol. 2001, 65, 135–172. [Google Scholar] [CrossRef]
- Wexler, N.S. Venezuelan kindreds reveal that genetic and environmental factors modulate Huntington’s disease age of onset. Proc. Natl. Acad. Sci. USA 2004, 101, 3498–3503. [Google Scholar] [PubMed] [Green Version]
- Quarrell, O.W.; Nance, M.A.; Nopoulos, P.; Paulsen, J.S.; Smith, J.A.; Squitieri, F. Managing juvenile Huntington’s disease. Neurodegener. Dis. Manag. 2013, 3, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Ramos, E.M.; Lee, J.H.; Gillis, T.; Mysore, J.S.; Hayden, M.R.; Warby, S.C.; Morrison, P.; Nance, M.; Ross, C.A.; et al. CAG repeat expansion in Huntington disease determines age at onset in a fully dominant fashion. Neurology 2012, 78, 690–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutsis, G.; Karadima, G.; Kladi, A.; Panas, M. Late-onset Huntington’s disease: Diagnostic and prognostic considerations. Parkinsonism Relat. D. 2014, 20, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.G.; Mitchell, J.D.; Lyon, M.; Moore, D.H. Riluzole for amyotrophic lateral sclerosis (ALS)/motor neuron disease (MND). Cochrane Database Syst. Rev. 2007, 7, 1–26. [Google Scholar] [CrossRef]
- Katyal, N.; Govindarajan, R. Shortcomings in the current amyotrophic lateral sclerosis trials and potential solutions for improvement. Front. Neurol. 2017, 8, 521. [Google Scholar] [CrossRef] [Green Version]
- Al-Chalabi, A.; Jones, A.; Troakes, C.; King, A.; Al-Sarraj, S.; van den Berg, L.H. The genetics and neuropathology of amyotrophic lateral sclerosis. Acta. Neuropathol. 2012, 124, 339–352. [Google Scholar] [CrossRef]
- Menke, R.A.; Körner, S.; Filippini, N.; Douaud, G.; Knight, S.; Talbot, K.; Turner, M.R. Widespread grey matter pathology dominates the longitudinal cerebral MRI and clinical landscape of amyotrophic lateral sclerosis. Brain 2014, 137, 2546–2555. [Google Scholar] [CrossRef] [Green Version]
- Hadzhieva, M.; Kirches, E.; Wilisch-Neumann, A.; Pachow, D.; Wallesch, M.; Schoenfeld, P.; Paege, I.; Vielhaber, S.; Petri, S.; Keilhoff, G. Dysregulation of iron protein expression in the G93A model of amyotrophic lateral sclerosis. Neuroscience 2013, 230, 94–101. [Google Scholar] [CrossRef]
- De Assis Lima, I.V.; Bastos, L.F.S.; Limborço-Filho, M.; Fiebich, B.L.; de Oliveira, A.C.P. Role of prostaglandins in neuroinflammatory and neurodegenerative diseases. Mediat. Inflamm. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Movahhedin, N.; Zengin, G.; Bahadori, M.B.; Sarikurkcu, C.; Bahadori, S.; Dinparast, L. Ajuga chamaecistus subsp. scoparia (Boiss.) Rech.f.: A new source of phytochemicals for antidiabetic, skin-care, and neuroprotective uses. Ind. Crop. Prod. 2016, 94, 89–96. [Google Scholar] [CrossRef]
- Karakaya, S.; Eksi, G.; Koca, M.; Demirci, B.; Kaymak, H.C.; Kaplan, M.E.; Aksakal, O. Chemical and morphological characterization of Allium tuncelianum (Amaryllidaceae) and its antioxidant and anticholinesterase potentials. Jard. Bot. Madr. 2019, 76, 85. [Google Scholar] [CrossRef]
- Abuhamdah, S.; Abuhamdah, R.; Howes, M.J.; Al-Olimat, S.; Ennaceur, A.; Chazot, P.L. Pharmacological and neuroprotective profile of an essential oil derived from leaves of Aloysia citrodora Palau. J. Pharm. Pharm. 2015, 67, 1306–1315. [Google Scholar] [CrossRef] [Green Version]
- Jyotshna; Srivastava, N.; Singh, B.; Chanda, D.; Shanker, K. Chemical composition and acetylcholinesterase inhibitory activity of Artemisia maderaspatana essential oil. Pharm. Biol. 2015, 53, 1677–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohoude, M.J.; Gbaguidi, F.; Agbani, P.; Ayedoun, M.A.; Cazaux, S.; Bouajila, J. Chemical composition and biological activities of extracts and essential oil of Boswellia dalzielii leaves. Pharm. Biol. 2017, 55, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Murata, K.; Tanaka, K.; Akiyama, R.; Noro, I.; Nishio, A.; Nakagawa, S.; Matsumura, S.; Matsuda, H. Anti-cholinesterase Activity of Crude Drugs Selected from the Ingredients of Incense Sticks and Heartwood of Chamaecyparis obtusa. Nat. Prod. Commun. 2018, 13. [Google Scholar] [CrossRef] [Green Version]
- Ademosun, A.O.; Oboh, G.; Olupona, A.J.; Oyeleye, S.I.; Adewuni, T.M.; Nwanna, E.E. Comparative Study of Chemical Composition, In Vitro Inhibition of Cholinergic and Monoaminergic Enzymes, and Antioxidant Potentials of Essential Oil from Peels and Seeds of Sweet Orange (Citrus Sinensis [L.] Osbeck) Fruits. J. Food Biochem. 2016, 40, 53–60. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Bonesi, M.; Menichini, F.; Mastellone, V.; Colica, C.; Menichini, F. Comparative Study on the Antioxidant Capacity and Cholinesterase Inhibitory Activity of Citrus aurantifolia Swingle, C. aurantium L., and C. bergamia Risso and Poit. Peel Essential Oils. J. Food Sci. 2012, 77, H40–H46. [Google Scholar] [CrossRef]
- Loizzo, M.; Ben Jemia, M.; Senatore, F.; Bruno, M.; Menichini, F.; Tundis, R. Chemistry and functional properties in prevention of neurodegenerative disorders of five Cistus species essential oils. Food Chem. Toxicol 2013, 59. [Google Scholar] [CrossRef]
- Ali-Shtayeh, M.S.; Jamous, R.M.; Abu-Zaitoun, S.Y.; Akkawi, R.J.; Kalbouneh, S.R.; Bernstein, N.; Dudai, N. Chemical profile and bioactive properties of the essential oil isolated from Clinopodium serpyllifolium (M.Bieb.) Kuntze growing in Palestine. Ind. Crop. Prod. 2018, 124, 617–625. [Google Scholar] [CrossRef]
- Lamamra, M.; Laouer, H.; Amira, S.; Erdogan Orhan, I.; Senol, F.; Demirci, B.; Akkal, S. Chemical Composition and Cholinesterase Inhibitory Activity of Different Parts of Daucus aristidis Coss. Essential Oils from Two Locations in Algeria. Rec. Nat. Prod. 2017, 11, 147–156. [Google Scholar]
- Arruda, M.; Viana, H.; Rainha, N.; Neng, N.R.; Rosa, J.S.; Nogueira, J.M.F.; Barreto, M.d.C. Anti-acetylcholinesterase and antioxidant activity of essential oils from Hedychium gardnerianum Sheppard ex Ker-Gawl. Molecules 2012, 17, 3082–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahadori, S.; Bahadori, M.B.; Zengin, G.; Maggi, F.; Dinparast, L.; Aktumsek, A. Chemical composition profile of the essential oil from Hymenocrater bituminous and its health functionality. Int. J. Food Prop. 2017, 20, S972–S980. [Google Scholar] [CrossRef]
- Bhadra, S.; Mukherjee, P.K.; Kumar, N.S.; Bandyopadhyay, A. Anticholinesterase activity of standardized extract of Illicium verum Hook. f. fruits. Fitoterapia 2011, 82, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Videira, R.; Castanheira, P.; Grãos, M.; Salgueiro, L.; Faro, C.; Cavaleiro, C. A necrodane monoterpenoid from Lavandula luisieri essential oil as a cell-permeable inhibitor of BACE-1, the β-secretase in Alzheimer’s disease. Flavour Frag J. 2013, 28, 380–388. [Google Scholar] [CrossRef]
- Costa, P.; Grosso, C.; Gonçalves, S.; Andrade, P.B.; Valentão, P.; Gabriela Bernardo-Gil, M.; Romano, A. Supercritical fluid extraction and hydrodistillation for the recovery of bioactive compounds from Lavandula viridis L’Hér. Food Chem. 2012, 135, 112–121. [Google Scholar] [CrossRef]
- Ali-Shtayeh, M.S.; Jamous, R.M.; Abu-Zaitoun, S.Y.; Khasati, A.I.; Kalbouneh, S.R. Biological Properties and Bioactive Components of Mentha spicata L. Essential Oil: Focus on Potential Benefits in the Treatment of Obesity, Alzheimer’s Disease, Dermatophytosis, and Drug-Resistant Infections. Evid-Based Complement. Altern Med. 2019, 2019, 3834265. [Google Scholar] [CrossRef] [Green Version]
- Amaral, R.; Fernandes, C.; Botas, G.; Cruz, R.; Santos, M.; Rocha, L. Anticholinesterase Activity of Essential oil from Leaves of Neomitranthes obscura (DC.) N. Silveira. Lat. Am. J. Pharm. 2014, 33, 860–863. [Google Scholar]
- Özbek, H.; Güvenalp, Z.; Ozek, T.; Sevindik, H.; Yuca, H.; Yerdelen, K.; Demirezer, L.Ö. Chemical Composition, Antioxidant and Anticholinesterase Activities of the Essential oil of Origanum rotundifolium Boiss. from Turkey. Rec. Nat. Prod. 2017, 11, 485–490. [Google Scholar] [CrossRef]
- Kawamoto, H.; Takeshita, F.; Murata, K. Inhibitory Effects of Essential Oil Extracts From Panax Plants Against β-Secretase, Cholinesterase, and Amyloid Aggregation. Nat. Prod. Commun. 2019, 14. [Google Scholar] [CrossRef]
- Bonesi, M.; Menichini, F.; Tundis, R.; Loizzo, M.; Conforti, F.; Passalacqua, N.; Statti, G.; Menichini, F. Acetylcholinesterase and butyrylcholinesterase inhibitory activity of Pinus species essential oils and their constituents. J. Enzym Inhib. Med. Ch. 2010, 25, 622–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, M.S.; da Cruz, J.N.; Gomes Silva, S.; da Costa, W.A.; de Sousa, S.H.B.; Bezerra, F.W.F.; Teixeira, E.; da Silva, N.J.N.; de Aguiar Andrade, E.H.; de Jesus Chaves Neto, A.M.; et al. Phytochemical profile, antioxidant activity, inhibition of acetylcholinesterase and interaction mechanism of the major components of the Piper divaricatum essential oil obtained by supercritical CO2. J. Supercrit Fluid 2019, 145, 74–84. [Google Scholar] [CrossRef]
- Bahadori, M.B.; Zengin, G.; Bahadori, S.; Maggi, F.; Dinparast, L. Chemical Composition of Essential Oil, Antioxidant, Antidiabetic, Anti-obesity, and Neuroprotective Properties of Prangos gaubae. Nat. Prod. Commun. 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Ullah, F.; Sadiq, A.; Ayaz, M.; Imran, M.; Ali, I.; Zeb, A.; Ullah, F.; Shah, M.R. Chemical composition, antioxidant and anticholinesterase potentials of essential oil of Rumex hastatus D. Don collected from the North West of Pakistan. BMC Complement. Altern. Med. 2016, 16, 29. [Google Scholar] [CrossRef] [Green Version]
- El Euch, S.K.; Hassine, D.B.; Cazaux, S.; Bouzouita, N.; Bouajila, J. Salvia officinalis essential oil: Chemical analysis and evaluation of anti-enzymatic and antioxidant bioactivities. S. Afr. J. Bot. 2019, 120, 253–260. [Google Scholar] [CrossRef]
- Tel, G.; Oztürk, M.; Duru, M.E.; Harmandar, M.; Topçu, G. Chemical composition of the essential oil and hexane extract of Salvia chionantha and their antioxidant and anticholinesterase activities. Food Chem. Toxicol 2010, 48, 3189–3193. [Google Scholar] [CrossRef]
- Duru, M.; Tel Çayan, G.; Öztürk, M.; Harmandar, M. Chemical Composition, Antioxidant and Anticholinesterase Activities of the Essential Oil of Salvia chrysophylla Staph. Rec. Nat. Prod. 2012, 6, 175. [Google Scholar]
- Bahadori, M.B.; Asghari, B.; Dinparast, L.; Zengin, G.; Sarikurkcu, C.; Abbas-Mohammadi, M.; Bahadori, S. Salvia nemorosa L.: A novel source of bioactive agents with functional connections. LWT 2017, 75, 42–50. [Google Scholar] [CrossRef]
- Bahadori, M.B.; Dinparast, L.; Zengin, G.; Sarikurkcu, C.; Bahadori, S.; Asghari, B.; Movahhedin, N. Functional components, antidiabetic, anti-Alzheimer’s disease, and antioxidant activities of Salvia syriaca L. Int. J. Food Prop. 2017, 20, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Öztürk, M. Anticholinesterase and antioxidant activities of Savoury (Satureja thymbra L.) with identified major terpenes of the essential oil. Food Chem. 2012, 134, 48–54. [Google Scholar] [CrossRef]
- Bahadori, M.B.; Maggi, F.; Zengin, G.; Asghari, B.; Eskandani, M. Essential oils of hedgenettles (Stachys inflata, S. lavandulifolia, and S. byzantina) have antioxidant, anti-Alzheimer, antidiabetic, and anti-obesity potential: A comparative study. Ind. Crop. Prod. 2020, 145, 112089. [Google Scholar] [CrossRef]
- Zengin, G.; Aktumsek, A.; Ceylan, R. Antioxidant Potential and Inhibition of Key Enzymes Linked to Alzheimer’s Diseases and Diabetes Mellitus by Monoterpene-Rich Essential Oil from Sideritis galatica Bornm. Endemic to Turkey. Rec. Nat. Prod. 2016, 75, 42–50. [Google Scholar]
- Temel, H.E.; Demirci, B.; Demirci, F.; Celep, F.; Kahraman, A.; Doğan, M.; Hüsnü Can Başer, K. Chemical characterization and anticholinesterase effects of essential oils derived from Salvia species. J. Essent Oil Res. 2016, 28, 322–331. [Google Scholar] [CrossRef]
- Cutillas, A.B.; Carrasco, A.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Thymus mastichina L. essential oils from Murcia (Spain): Composition and antioxidant, antienzymatic and antimicrobial bioactivities. PLoS ONE 2018, 13, e0190790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevindik, H.; Ozek, T.; Yerdelen, K.; Önal, M.; Özbek, H.; Güvenalp, Z.; Demirezer, L.Ö. Chemical Composition, Antioxidant Capacity, Acetyl- and Butyrylcholinesterase Inhibitory Activities of the Essential Oil of Thymus haussknechtii Velen. Rec. Nat. Prod. 2016, 10, 503–507. [Google Scholar]
- Costa, P.; Gonçalves, S.; Grosso, C.; Andrade, P.B.; Valentão, P.; Bernardo-Gil, M.G.; Romano, A. Chemical profiling and biological screening of Thymus lotocephalus extracts obtained by supercritical fluid extraction and hydrodistillation. Ind. Crop. Prod. 2012, 36, 246–256. [Google Scholar] [CrossRef]
- Okonogi, S.; Chaiyana, W. Enhancement of anti-cholinesterase activity of Zingiber cassumunar essential oil using a microemulsion technique. Drug Discov. 2012, 6, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Arantes, S.; Piçarra, A.; Candeias, F.; Teixeira, D.; Caldeira, A.T.; Martins, M.R. Antioxidant activity and cholinesterase inhibition studies of four flavouring herbs from Alentejo. Nat. Prod. Res. 2017, 31, 2183–2187. [Google Scholar] [CrossRef] [Green Version]
- Chaiyana, W.; Saeio, K.; Hennink, W.E.; Okonogi, S. Characterization of potent anticholinesterase plant oil based microemulsion. Int. J. Pharm. 2010, 401, 32–40. [Google Scholar] [CrossRef]
- Caputo, L.; Piccialli, I.; Ciccone, R.; de Caprariis, P.; Massa, A.; de Feo, V.; Pannaccione, A. Lavender and coriander essential oils and their main component linalool exert a protective effect against amyloid-β neurotoxicity. Phytother. Res. 2020. [Google Scholar] [CrossRef]
- Kaufmann, D.; Dogra, A.; Wink, M. Myrtenal inhibits acetylcholinesterase, a known Alzheimer target. J. Pharm. Pharm. 2011, 63, 1368–1371. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Yang, L.; Li, J. Protection against β-amyloid-induced neurotoxicity by naturally occurring Z-ligustilide through the concurrent regulation of p38 and PI3-K/Akt pathways. Neurochem. Int. 2016, 100, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Akbaba, E.; Hassan, S.; Sur, T.; Bagci, E. Memory Enhancing, Anxiolytic and Antidepressant Effects of Achillea biebersteinii (Asteraceae) Essential Oil on Scopolamine-Induced Rats. J. Essent Oil-Bear Plants 2018, 21, 825–839. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, W.; Luan, J.; Chen, D.; Liu, L.; Feng, X. Ameliorative effects of olibanum essential oil on learning and memory in Aβ1-42-induced Alzheimer’s disease mouse model. Trop. J. Pharm. Res. 2020, 19, 1643–1651. [Google Scholar]
- Bae, D.; Seol, H.; Yoon, H.G.; Na, J.R.; Oh, K.; Choi, C.Y.; Lee, D.W.; Jun, W.; Youl Lee, K.; Lee, J.; et al. Inhaled essential oil from Chamaecyparis obtuse ameliorates the impairments of cognitive function induced by injection of β-amyloid in rats. Pharm. Biol. 2012, 50, 900–910. [Google Scholar] [CrossRef]
- Liu, B.; Kou, J.; Li, F.; Huo, D.; Xu, J.; Zhou, X.; Meng, D.; Ghulam, M.; Artyom, B.; Gao, X.; et al. Lemon essential oil ameliorates age-associated cognitive dysfunction via modulating hippocampal synaptic density and inhibiting acetylcholinesterase. Aging 2020, 12, 8622–8639. [Google Scholar] [CrossRef]
- Hritcu, L.; Boiangiu, R.S.; de Morais, M.C.; de Sousa, D.P. (-)cis-Carveol, a Natural Compound, Improves β-Amyloid-Peptide 1-42-Induced Memory Impairment and Oxidative Stress in the Rat Hippocampus. Biomed. Res. Int. 2020, 2020, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hritcu, L.; Bagci, E.; Aydin, E.; Mihasan, M. Antiamnesic and Antioxidants Effects of Ferulago angulata Essential Oil Against Scopolamine-Induced Memory Impairment in Laboratory Rats. Neurochem. Res. 2015, 40, 1799–1809. [Google Scholar] [CrossRef]
- Hritcu, L.; Cioanca, O.; Hancianu, M. Effects of lavender oil inhalation on improving scopolamine-induced spatial memory impairment in laboratory rats. Phytomedicine 2012, 19, 529–534. [Google Scholar] [CrossRef]
- Ayuob, N.N.; El Wahab, M.G.A.; Ali, S.S.; Abdel-Tawab, H.S. Ocimum basilicum improve chronic stress-induced neurodegenerative changes in mice hippocampus. Metab. Brain Dis. 2018, 33, 795–804. [Google Scholar] [CrossRef]
- Gradinariu, V.; Cioanca, O.; Hritcu, L.; Trifan, A.; Gille, E.; Hancianu, M. Comparative efficacy of Ocimum sanctum L. and Ocimum basilicum L. essential oils against amyloid beta (1–42)-induced anxiety and depression in laboratory rats. PhytoChem. Rev. 2014, 14, 1–9. [Google Scholar] [CrossRef]
- Aydin, E.; Hritcu, L.; Dogan, G.; Hayta, S.; Bagci, E. The Effects of Inhaled Pimpinella peregrina Essential Oil on Scopolamine-Induced Memory Impairment, Anxiety, and Depression in Laboratory Rats. Mol. Neurobiol. 2016, 53, 6557–6567. [Google Scholar] [CrossRef] [PubMed]
- Postu, P.A.; Sadiki, F.Z.; El Idrissi, M.; Cioanca, O.; Trifan, A.; Hancianu, M.; Hritcu, L. Pinus halepensis essential oil attenuates the toxic Alzheimer’s amyloid beta (1-42)-induced memory impairment and oxidative stress in the rat hippocampus. Biomed. Pharm. 2019, 112, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Li, H.; Dong, J.; Yang, W.; Liu, T.; Wang, Y.; Wang, X.; Wang, M.; Zhi, D. Rose Essential Oil Delayed Alzheimer’s Disease-Like Symptoms by SKN-1 Pathway in C. elegans. J. Agric. Food Chem. 2017, 65, 8855–8865. [Google Scholar] [CrossRef]
- Satou, T.; Hanashima, Y.; Mizutani, I.; Koike, K. The effect of inhalation of essential oil from Rosmarinus officinalis on scopolamine-induced Alzheimer’s type dementia model mice. Flavour Frag J. 2018, 33, 230–234. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, J.-h.; Zhang, Y.; Han, C.; Li, J. Anti-Alzheimer’s disease effect of essential oil from aerial parts of Salvia miltiorrhiza Bge. Int. J. Clin. Exp. Med. 2018, 11, 641–652. [Google Scholar]
- Xu, M.; Zhang, X.; Ren, F.; Yan, T.; Wu, B.; Bi, K.; Bi, W.; Jia, Y. Essential oil of Schisandra chinensis ameliorates cognitive decline in mice by alleviating inflammation. Food Funct. 2019, 10, 5827–5842. [Google Scholar] [CrossRef]
- Hong, Y.K.; Park, S.H.; Lee, S.; Hwang, S.; Lee, M.J.; Kim, D.; Lee, J.H.; Han, S.Y.; Kim, S.T.; Kim, Y.K.; et al. Neuroprotective effect of SuHeXiang Wan in Drosophila models of Alzheimer’s disease. J. Ethnopharmacol. 2011, 134, 1028–1032. [Google Scholar] [CrossRef]
- Sadiki, F.Z.; Idrissi, M.E.; Cioanca, O.; Trifan, A.; Hancianu, M.; Hritcu, L.; Postu, P.A. Tetraclinis articulata essential oil mitigates cognitive deficits and brain oxidative stress in an Alzheimer’s disease amyloidosis model. Phytomedicine 2019, 56, 57–63. [Google Scholar] [CrossRef]
- Eskandari-Roozbahani, N.; Shomali, T.; Taherianfard, M. Neuroprotective Effect of Zataria Multiflora Essential Oil on Rats With Alzheimer Disease: A Mechanistic Study. Basic Clin. Neurosci. 2019, 10, 85–97. [Google Scholar] [CrossRef]
- Ahmadi, M.; Taherianfard, M.; Shomali, T. Zataria multiflora could improve hippocampal tau protein and TNF(α) levels and cognitive behavior defects in a rat model of Alzheimer’s disease. Avicenna J. Phytomed. 2019, 9, 465–473. [Google Scholar] [PubMed]
- Medhat, D.; El-Mezayen, H.A.; El-Naggar, M.E.; Farrag, A.R.; Abdelgawad, M.E.; Hussein, J.; Kamal, M.H. Evaluation of urinary 8-hydroxy-2-deoxyguanosine level in experimental Alzheimer’s disease: Impact of carvacrol nanoparticles. Mol. Biol. Rep. 2019, 46, 4517–4527. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Chen, Y.B.; Chen, D.F.; Lai, X.P.; Liu, D.H.; Deng, R.D.; Zhou, J.H.; Zhang, S.X.; Li, Y.W.; Lii, H.; et al. β-Asarone inhibits neuronal apoptosis via the CaMKII/CREB/Bcl-2 signaling pathway in an in vitro model and AβPP/PS1 mice. J. Alzheimers Dis. 2013, 33, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Cioanca, O.; Hancianu, M.; Mircea, C.; Trifan, A.; Hritcu, L. Essential oils from Apiaceae as valuable resources in neurological disorders: Foeniculi vulgare aetheroleum. Ind. Crop. Prod. 2016, 88. [Google Scholar] [CrossRef]
- Xu, P.; Wang, K.; Lu, C.; Dong, L.; Gao, L.; Yan, M.; Aibai, S.; Liu, X. Protective effect of lavender oil on scopolamine induced cognitive deficits in mice and H2O2 induced cytotoxicity in PC12 cells. J. Ethnopharmacol. 2016, 193, 408–415. [Google Scholar] [CrossRef]
- Xu, P.; Wang, K.; Lu, C.; Dong, L.; Gao, L.; Yan, M.; Aibai, S.; Yang, Y.; Liu, X. The Protective Effect of Lavender Essential Oil and Its Main Component Linalool against the Cognitive Deficits Induced by D-Galactose and Aluminum Trichloride in Mice. Evid-Based Complement. Altern. Med. 2017, 2017, 7426538. [Google Scholar] [CrossRef]
- Videira, R.; Castanheira, P.; Grãos, M.; Resende, R.; Salgueiro, L.; Faro, C.; Cavaleiro, C. Dose-dependent inhibition of BACE-1 by the monoterpenoid 2,3,4,4-tetramethyl-5-methylenecyclopent-2-enone in cellular and mouse models of Alzheimer’s disease. J. Nat. Prod. 2014, 77, 1275–1279. [Google Scholar] [CrossRef]
- Zouari-Bouassida, K.; Trigui, M.; Makni, S.; Jlaiel, L.; Tounsi, S. Seasonal Variation in Essential Oils Composition and the Biological and Pharmaceutical Protective Effects of Mentha longifolia Leaves Grown in Tunisia. Biomed. Res. Int. 2018, 2018, 7856517. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.; Hur, J.; Jeong, H.J.; Koo, B.S.; Pak, S.C. SuHeXiang Wan essential oil alleviates amyloid beta induced memory impairment through inhibition of tau protein phosphorylation in mice. Am. J. Chin. Med. 2011, 39, 917–932. [Google Scholar] [CrossRef]
- Yen, P.L.; Cheng, S.S.; Wei, C.C.; Lina, H.Y.; Liao, V.H.; Chang, S.T. Antioxidant Activities and Reduced Amyloid-β Toxicity of 7-Hydroxycalamenene Isolated from the Essential Oil of Zelkova serrata Heartwood. Nat. Prod. Commun. 2016, 11, 1357–1362. [Google Scholar] [CrossRef] [Green Version]
- Fathy, M.M.; Eid, H.; Hussein, M.A.; Ahmed, H.H.; Hussein, A. The role of Zingiber officinale in the treatment of Alzheimer’s disease: In-vitro and in-vivo evidences. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 735–749. [Google Scholar]
- Ghajarbeygi, P.; Hajhoseini, A.; Hosseini, M.S.; Sharifan, A. An In Vitro and In Vivo Cholinesterase Inhibitory Activity of Pistacia khinjuk and Allium sativum Essential Oils. J. Pharm. 2019, 22, 231–238. [Google Scholar] [CrossRef]
- Demirezer, L.; Gürbüz, P.; Kelicen Uğur, E.P.; Bodur, M.; Özenver, N.; Uz, A.; Güvenalp, Z. Molecular docking and ex vivo and in vitro anticholinesterase activity studies of Salvia sp. and highlighted rosmarinic acid. Turk. J. Med. Sci. 2015, 45, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Adefegha, A.; Oboh, G.; Odubanjo, O.; Ogunsuyi, O. Comparative study on the antioxidative activities, anticholinesterase properties and essential oil composition of Clove (Syzygium aromaticum) bud and Ethiopian pepper (Xylopia aethiopica). La Rivista Italiana Delle Sostanze Grasse 2016, 92, 257–268. [Google Scholar]
- Ramazani, E.; YazdFazeli, M.; Emami, S.A.; Mohtashami, L.; Javadi, B.; Asili, J.; Tayarani-Najaran, Z. Protective effects of Cinnamomum verum, Cinnamomum cassia and cinnamaldehyde against 6-OHDA-induced apoptosis in PC12 cells. Mol. Biol. Rep. 2020, 7, 2437–2445. [Google Scholar] [CrossRef] [PubMed]
- Klein-Júnior, L.C.; dos Santos Passos, C.; Tasso de Souza, T.J.; Gobbi de Bitencourt, F.; Salton, J.; de Loreto Bordignon, S.A.; Henriques, A.T. The monoamine oxidase inhibitory activity of essential oils obtained from Eryngium species and their chemical composition. Pharm. Biol. 2016, 54, 1071–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morshedi, D.; Nasouti, M. Essential Oils May Lead α-Synuclein towards Toxic Fibrils Formation. Parkinsons Dis. 2016, 2016, 6219249. [Google Scholar] [CrossRef] [Green Version]
- Morshedi, D.; Aliakbari, F.; Tayaranian-Marvian, A.; Fassihi, A.; Pan-Montojo, F.; Pérez-Sánchez, H. Cuminaldehyde as the major component of Cuminum cyminum, a natural aldehyde with inhibitory effect on alpha-synuclein fibrillation and cytotoxicity. J. Food Sci. 2015, 80, H2336–H2345. [Google Scholar] [CrossRef]
- Ning, B.; Deng, M.; Zhang, Q.; Wang, N.; Fang, Y. β-Asarone inhibits IRE1/XBP1 endoplasmic reticulum stress pathway in 6-OHDA-induced parkinsonian rats. Neurochem. Res. 2016, 41, 2097–2101. [Google Scholar] [CrossRef]
- Ning, B.; Zhang, Q.; Wang, N.; Deng, M.; Fang, Y. β-Asarone regulates ER stress and autophagy via inhibition of the PERK/CHOP/Bcl-2/Beclin-1 Pathway in 6-OHDA-induced parkinsonian rats. Neurochem. Res. 2019, 44, 1159–1166. [Google Scholar] [CrossRef]
- Beserra-Filho, J.I.; de Macêdo, A.M.; Leão, A.H.; Bispo, J.M.M.; Santos, J.R.; de Oliveira-Melo, A.J.; Menezes, P.D.P.; Duarte, M.C.; de Souza Araújo, A.A.; Silva, R.H. Eplingiella fruticosa leaf essential oil complexed with β-cyclodextrin produces a superior neuroprotective and behavioral profile in a mice model of Parkinson’s disease. Food Chem. Toxicol. 2019, 124, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Nemati, M.; Hemmati, A.A.; Najafzadeh, H.; Mansouri, M.T.; Khodayar, M.J. Evaluation of the Effects of Foeniculum vulgare Essence on Behavioral-Motor Disorders of Parkinson’s Disease induced by Reserpine in Ovariectomized and Non Ovariectomized Rats. Jundishapur J. Nat. Pharm. Prod. 2018, 13, e67391. [Google Scholar] [CrossRef]
- Issa, M.Y.; Ezzat, M.I.; Sayed, R.H.; Elbaz, E.M.; Omar, F.A.; Mohsen, E. Neuroprotective effects of Pulicaria undulata essential oil in rotenone model of parkinson’s disease in rats: Insights into its anti-inflammatory and anti-oxidant effects. S. Afr. J. Bot. 2020, 132, 289–298. [Google Scholar] [CrossRef]
- Liu, Q.; Sang Heon, K.; Yung-Wei, S.; Sok Cheon, P.; Wonwoong, L.; Jongki, H.; Jaehwan, J.; Kyoung Sang, C.; Songhee, J.; Byung-Soo, K. Neuroprotective effects of Suhexiang Wan on the in vitro and in vivo models of Parkinson’s disease. J. Tradit. Chin. Med. 2019, 39, 800–808. [Google Scholar]
- Nikolova, G.; Karamalakova, Y.; Kovacheva, N.; Stanev, S.; Zheleva, A.; Gadjeva, V. Protective effect of two essential oils isolated from Rosa damascena Mill. and Lavandula angustifolia Mill, and two classic antioxidants against L-dopa oxidative toxicity induced in healthy mice. Regul. Toxicol. Pharm. 2016, 81, 1–7. [Google Scholar] [CrossRef]
- Nikolova, G.; Karamalakova, Y.; Gadjeva, V. Reducing oxidative toxicity of L-dopa in combination with two different antioxidants: An essential oil isolated from Rosa Damascena Mill., and vitamin C. Toxicol. Rep. 2019, 6, 267–271. [Google Scholar] [CrossRef]
- Costa, P.; Gonçalves, S.; Valentão, P.; Andrade, P.B.; Almeida, C.; Nogueira, J.M.; Romano, A. Metabolic profile and biological activities of Lavandula pedunculata subsp. lusitanica (Chaytor) Franco: Studies on the essential oil and polar extracts. Food Chem. 2013, 141, 2501–2506. [Google Scholar] [CrossRef]
- Elmann, A.; Mordechay, S.; Rindner, M.; Ravid, U. Anti-neuroinflammatory effects of geranium oil in microglial cells. J. Funct. Foods 2010, 2, 17–22. [Google Scholar] [CrossRef]
- Montalván, M.; Peñafiel, M.A.; Ramírez, J.; Cumbicus, N.; Bec, N.; Larroque, C.; Bicchi, C.; Gilardoni, G. Chemical Composition, Enantiomeric Distribution, and Sensory Evaluation of the Essential Oils Distilled from the Ecuadorian Species Myrcianthes myrsinoides (Kunth) Grifo and Myrcia mollis (Kunth) DC. (Myrtaceae). Plants 2019, 8, 511. [Google Scholar] [CrossRef] [Green Version]
- Bagci, E.; Aydin, E.; Ungureanu, E.; Hritcu, L. Anthriscus nemorosa essential oil inhalation prevents memory impairment, anxiety and depression in scopolamine-treated rats. Biomed. Pharm. 2016, 84, 1313–1320. [Google Scholar] [CrossRef]
- Bassam, A.-S. Antibacterial activities of some plant extracts utilized in popular medicine in Palestine. Turk. J. Biol. 2004, 28, 99–102. [Google Scholar]
- Rios, J.-L.; Recio, M. Medicinal plants and antimicrobial activity. J. Ethnopharmacol. 2005, 100, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Donelian, A.; Carlson, L.H.C.; Lopes, T.J.; Machado, R.A.F. Comparison of extraction of patchouli (Pogostemon cablin) essential oil with supercritical CO2 and by steam distillation. J. Supercrit. Fluids 2009, 48, 15–20. [Google Scholar] [CrossRef]
- Irshad, M.; Subhani, M.A.; Ali, S.; Hussain, A. Biological Importance of Essential Oils. In Essential Oils-Oils of Nature; El-Shemy, H.A., Ed.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef] [Green Version]
- Natural Living Ideas. The 10 Most Popular Essential Oils & 174 Magical Ways To Use Them. Available online: https://www.naturallivingideas.com/most-popular-essential-oils/ (accessed on 12 December 2020).
- Sadlon, A.E.; Lamson, D.W. Immune-modifying and antimicrobial effects of Eucalyptus oil and simple inhalation devices. Altern. Med. Rev. 2010, 15, 33–47. [Google Scholar]
- ESCOP. Alchemillae Herba, The Scientific Foundation for Herbal Medicinal Products. Available online: www.escop.com (accessed on 12 December 2020).
- Juergens, U.R. Anti-inflammatory properties of the monoterpene 1.8-cineole: Current evidence for co-medication in inflammatory airway diseases. Drug Res. 2014, 64, 638–646. [Google Scholar] [CrossRef]
- CSID:2656. 1,8-cineole. Available online: http://www.chemspider.com/Chemical-Structure.2656.html (accessed on 15 January 2021).
- CSID:6402. α-pinene. Available online: http://www.chemspider.com/Chemical-Structure.6402.html (accessed on 15 January 2021).
- CSID:14198. β-pinene. Available online: http://www.chemspider.com/Chemical-Structure.14198.html (accessed on 15 January 2021).
- CSID:28952. Zingerone. Available online: http://www.chemspider.com/Chemical-Structure.28952.html (accessed on 15 January 2021).
- CSID:388386. Limonene. Available online: http://www.chemspider.com/Chemical-Structure.388386.html (accessed on 15 January 2021).
- CSID:390582. Ferruginol. Available online: http://www.chemspider.com/Chemical-Structure.390582.html (accessed on 15 January 2021).
- CSID:454172. Kaur-16-ene. Available online: http://www.chemspider.com/Chemical-Structure.454172.html (accessed on 15 January 2021).
- CSID:4474888. Rosmarinic acid. Available online: http://www.chemspider.com/Chemical-Structure.4474888.html (accessed on 15 January 2021).
- CSID:13849981. Linalool. Available online: http://www.chemspider.com/Chemical-Structure.13849981.html (accessed on 15 January 2021).
- CSID:21105867. Carvacrol. Available online: http://www.chemspider.com/Chemical-Structure.21105867.html (accessed on 15 January 2021).
- Dhanasekaran, S.; Perumal, P.; Palayan, M. In-vitro Screening for acetylcholinesterase enzyme inhibition potential and antioxidant activity of extracts of Ipomoea aquatica Forsk: Therapeutic lead for Alzheimer’s disease. J. Appl. Pharm. Sci. 2015, 5, 012–016. [Google Scholar] [CrossRef] [Green Version]
- Silver, A. The Biology of Cholinesterases, North-Holland Publishing Company: Amsterdam, The Netherlands, 1974; 1–596.
- Wilson, B.; Philip, W. Cholinesterase inhibition. In Encyclopedia of Toxicology; Elsevier: New York, NY, USA, 2005; pp. 588–599. [Google Scholar]
- Wilson, B.W. CHAPTER 48-Cholinesterases. In Handbook of Pesticide Toxicology, 2nd ed.; Krieger, R.I., Krieger, W.C., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 967–985. [Google Scholar] [CrossRef]
- Chatonnet, A.; Lockridge, O. Comparison of butyrylcholinesterase and acetylcholinesterase. Biochem. J. 1989, 260, 625–634. [Google Scholar] [CrossRef]
- Schwarz, M.; Glick, D.; Loewenstein, Y.; Soreq, H. Engineering of human cholinesterases explains and predicts diverse consequences of administration of various drugs and poisons. Pharm. Ther. 1995, 67, 283–322. [Google Scholar] [CrossRef]
- Tecles, F.; Ceron, J. Determination of whole blood cholinesterase in different animal species using specific substrates. Res. Vet. Sci. 2001, 70, 233–238. [Google Scholar] [CrossRef]
- Sramek, J.J.; Cutler, N.R. RBC cholinesterase inhibition: A useful surrogate marker for cholinesterase inhibitor activity in Alzheimer disease therapy? Alzheimer Dis. Assoc. Disord 2000, 14, 216–227. [Google Scholar] [CrossRef]
- Nicolet, Y.; Lockridge, O.; Masson, P.; Fontecilla-Camps, J.C.; Nachon, F. Crystal structure of human butyrylcholinesterase and of its complexes with substrate and products. J. Biol. Chem. 2003, 278, 41141–41147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, A.; Redman, A.M.; Jiang, X.; Lockridge, O.; Doctor, B.P. Differences in active site gorge dimensions of cholinesterases revealed by binding of inhibitors to human butyrylcholinesterase. Biochemistry 1997, 36, 14642–14651. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Colletier, J.-P.; Weik, M.; Jiang, H.; Moult, J.; Silman, I.; Sussman, J.L. Flexibility of aromatic residues in the active-site gorge of acetylcholinesterase: X-ray versus molecular dynamics. Biophys. J. 2008, 95, 2500–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dighe, S.N.; Deora, G.S.; de la Mora, E.; Nachon, F.; Chan, S.; Parat, M.-O.; Brazzolotto, X.; Ross, B.P. Discovery and Structure–Activity Relationships of a Highly Selective Butyrylcholinesterase Inhibitor by Structure-Based Virtual Screening. J. Med. Chem. 2016, 59, 7683–7689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Lu, X.; Du, C.; Liu, Y.; Wang, Y.; Hong, K.H.; Chen, Y.; Sun, H. Novel BuChE-IDO1 inhibitors from sertaconazole: Virtual screening, chemical optimization and molecular modeling studies. Bioorg. Med. Chem. Lett. 2021, 34, 127756. [Google Scholar] [CrossRef] [PubMed]
- Miles, J.A.; Kapure, J.S.; Deora, G.S.; Courageux, C.; Igert, A.; Dias, J.; McGeary, R.P.; Brazzolotto, X.; Ross, B.P. Rapid discovery of a selective butyrylcholinesterase inhibitor using structure-based virtual screening. Bioorg. Med. Chem. Lett. 2020, 30, 127609. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharm. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Pohanka, M.; Jun, D.; Kuca, K. Improvement of acetylcholinesterase-based assay for organophosphates in way of identification by reactivators. Talanta 2008, 77, 451–454. [Google Scholar] [CrossRef]
- Morizono, M.; Akinaga, Y. Studies on Tissue Cholinesterase in Domestic Animals II. Detection of Tissue Cholinesterase Isoenzymes. Mem. Fac. Agr. Kagoshima. Univ. 1981, 17, 219–234. [Google Scholar]
- Frasco, M.; Fournier, D.; Carvalho, F.; Guilhermino, L. Do metals inhibit acetylcholinesterase (AChE)? Implementation of assay conditions for the use of AChE activity as a biomarker of metal toxicity. Biomarkers 2005, 10, 360–375. [Google Scholar] [CrossRef]
- Šinko, G.; Čalić, M.; Bosak, A.; Kovarik, Z. Limitation of the Ellman method: Cholinesterase activity measurement in the presence of oximes. Anal. Biochem. 2007, 370, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Marrs, T.C. Organophosphates: History, chemistry, pharmacology. In Organophosphates and Health; Imperial College Press: London, UK, 2001; pp. 1–36. [Google Scholar]
- Michel, H.O. An electrometric method for the determination of red blood cell and plasma cholinesterase activity. J. Lab. Clin. Med. 1949, 34, 1564–1568. [Google Scholar]
- Selkoe, D.J. Alzheimer’s disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 742–760. [Google Scholar] [CrossRef] [PubMed]
- Rochette, M.; Murphy, M. Gamma-secretase: Substrates and inhibitors. Mol. Neurobiol. 2002, 26, 81–95. [Google Scholar] [CrossRef]
- Lee, V.M.; Goedert, M.; Trojanowski, J.Q. Neurodegenerative tauopathies. Annu Rev. Neurosci. 2001, 24, 1121–1159. [Google Scholar] [CrossRef]
- Satou, T.; Cummings, B.J.; Head, E.; Nielson, K.A.; Hahn, F.F.; Milgram, N.W.; Velazquez, P.; Cribbs, D.H.; Tenner, A.J.; Cotman, C.W. The progression of β-amyloid deposition in the frontal cortex of the aged canine. Brain Res. 1997, 774, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Cummings, B.J.; Head, E.; Afagh, A.J.; Milgram, N.W.; Cotman, C.W. β-amyloid accumulation correlates with cognitive dysfunction in the aged canine. Neurobiol. Learn. Mem. 1996, 66, 11–23. [Google Scholar] [CrossRef] [Green Version]
- De Felice, F.G.; Munoz, D.P. Opportunities and challenges in developing relevant animal models for Alzheimer’s disease. Ageing Res. Rev. 2016, 26, 112–114. [Google Scholar] [CrossRef]
- Casadesus, G. Handbook of Animal Models in Alzheimer’s Disease, 1st ed.; IOS Press: Amsterdam, The Netherlands, 2011; Volume 1, p. 340. [Google Scholar]
- Lecanu, L.; Papadopoulos, V. Modeling Alzheimer’s disease with non-transgenic rat models. Alzheimer’s Res. 2013, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Baser, K.H. Biological and pharmacological activities of carvacrol and carvacrol bearing essential oils. Curr Pharm. Des. 2008, 14, 3106–3119. [Google Scholar] [CrossRef]
- Jukic, M.; Politeo, O.; Maksimovic, M.; Milos, M.; Milos, M. In vitro acetylcholinesterase inhibitory properties of thymol, carvacrol and their derivatives thymoquinone and thymohydroquinone. Phytother. Res. 2007, 21, 259–261. [Google Scholar] [CrossRef] [PubMed]
- Melo, F.H.; Venâncio, E.T.; de Sousa, D.P.; de França Fonteles, M.M.; de Vasconcelos, S.M.; Viana, G.S.; de Sousa, F.C. Anxiolytic-like effect of Carvacrol (5-isopropyl-2-methylphenol) in mice: Involvement with GABAergic transmission. Fundam. Clin. Pharm. 2010, 24, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Melo, F.H.; Moura, B.A.; de Sousa, D.P.; de Vasconcelos, S.M.; Macedo, D.S.; Fonteles, M.M.; Viana, G.S.; de Sousa, F.C. Antidepressant-like effect of carvacrol (5-Isopropyl-2-methylphenol) in mice: Involvement of dopaminergic system. Fundam. Clin. Pharm. 2011, 25, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Krishtal, J.; Bragina, O.; Metsla, K.; Palumaa, P.; Tõugu, V. In situ fibrillizing amyloid-beta 1-42 induces neurite degeneration and apoptosis of differentiated SH-SY5Y cells. PLoS ONE 2017, 12, e0186636. [Google Scholar] [CrossRef] [Green Version]
- Pike, C.J.; Walencewicz-Wasserman, A.J.; Kosmoski, J.; Cribbs, D.H.; Glabe, C.G.; Cotman, C.W. Structure-activity analyses of β-amyloid peptides: Contributions of the β25–35 region to aggregation and neurotoxicity. J. Neurochem. 1995, 64, 253–265. [Google Scholar] [CrossRef]
- Kaminsky, Y.G.; Marlatt, M.W.; Smith, M.A.; Kosenko, E.A. Subcellular and metabolic examination of amyloid-β peptides in Alzheimer disease pathogenesis: Evidence for Aβ25–35. Exp. Neurol. 2010, 221, 26–37. [Google Scholar] [CrossRef]
- Gabbouj, S.; Ryhänen, S.; Marttinen, M.; Wittrahm, R.; Takalo, M.; Kemppainen, S.; Martiskainen, H.; Tanila, H.; Haapasalo, A.; Hiltunen, M.; et al. Altered Insulin Signaling in Alzheimer’s Disease Brain—Special Emphasis on PI3K-Akt Pathway. Front. Neurosci. 2019, 13. [Google Scholar] [CrossRef]
- Wu, Y.; Han, X.; Yuan, W.; Wang, X.; Meng, D.; Hu, J.; Lv, Z. Salt intervention for the diversities of essential oil composition, aroma and antioxidant activities of Kushui rose (R. setate× R. rugosa). Ind. Crop. Prod. 2020, 150, 112417. [Google Scholar] [CrossRef]
- Hsu, H.; Hsu, C. Commonly used Chinese herb formulas with illustrations, 2nd ed.; Oriental Healing Arts Institute: Long Beach, CA, USA, 1980; Volume 1, p. 671. [Google Scholar]
- Wang, M.C.; Bohmann, D.; Jasper, H. JNK signaling confers tolerance to oxidative stress and extends lifespan in Drosophila. Dev. Cell 2003, 5, 811–816. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.C.; Bohmann, D.; Jasper, H. JNK extends life span and limits growth by antagonizing cellular and organism-wide responses to insulin signaling. Cell 2005, 121, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Gulactı, T.; Ufuk, K.; Mehmet, O.; Mehmet, B.; Seda Damla, H.; Fatemeh, B.; Burcu, C.; Tuncay, D. Investigation of Anticholinesterase Activity of a Series of Salvia Extracts and the Constituents of Salvia staminea. Nat. Prod. J. 2013, 3, 3–9. [Google Scholar] [CrossRef]
- Chen, J.; Montanari, A.M.; Widmer, W.W. Two New Polymethoxylated Flavones, a Class of Compounds with Potential Anticancer Activity, Isolated from Cold Pressed Dancy Tangerine Peel Oil Solids. J. Agric. Food Chem. 1997, 45, 364–368. [Google Scholar] [CrossRef]
- Nogata, Y.; Sakamoto, K.; Shiratsuchi, H.; Ishii, T.; Yano, M.; Ohta, H. Flavonoid Composition of Fruit Tissues of Citrus Species. Biosci. Biotechnol. Biochem. 2006, 70, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avetisyan, A.; Markosian, A.; Petrosyan, M.; Sahakyan, N.; Babayan, A.; Aloyan, S.; Trchounian, A. Chemical composition and some biological activities of the essential oils from basil Ocimum different cultivars. BMC Complement. Altern. Med. 2017, 17, 60. [Google Scholar] [CrossRef] [Green Version]
- Batista, P.A.; Werner, M.F.; Oliveira, E.C.; Burgos, L.; Pereira, P.; Brum, L.F.; Story, G.M.; Santos, A.R. The antinociceptive effect of (-)-linalool in models of chronic inflammatory and neuropathic hypersensitivity in mice. J. Pain 2010, 11, 1222–1229. [Google Scholar] [CrossRef]
- Elisabetsky, E.; Marschner, J.; Souza, D.O. Effects of Linalool on glutamatergic system in the rat cerebral cortex. Neurochem. Res. 1995, 20, 461–465. [Google Scholar] [CrossRef]
- Letizia, C.S.; Cocchiara, J.; Lalko, J.; Api, A.M. Fragrance material review on linalool. Food Chem. Toxicol. 2003, 41, 943–964. [Google Scholar] [CrossRef]
- Mitić-Culafić, D.; Zegura, B.; Nikolić, B.; Vuković-Gacić, B.; Knezević-Vukcević, J.; Filipic, M. Protective effect of linalool, myrcene and eucalyptol against t-butyl hydroperoxide induced genotoxicity in bacteria and cultured human cells. Food Chem. Toxicol. 2009, 47, 260–266. [Google Scholar] [CrossRef]
- Sabogal-Guáqueta, A.M.; Osorio, E.; Cardona-Gómez, G.P. Linalool reverses neuropathological and behavioral impairments in old triple transgenic Alzheimer’s mice. Neuropharmacology 2016, 102, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Jankovic, J.; Aguilar, L.G. Current approaches to the treatment of Parkinson’s disease. Neuropsych. Dis. Treat. 2008, 4, 743–757. [Google Scholar] [CrossRef] [Green Version]
- Rao, P.V.; Gan, S.H. Cinnamon: A multifaceted medicinal plant. Evid-Based Complement. Altern. Med. 2014, 2014, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brahmachari, S.; Jana, A.; Pahan, K. Sodium benzoate, a metabolite of cinnamon and a food additive, reduces microglial and astroglial inflammatory responses. J. Immunol. 2009, 183, 5917–5927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollazadeh, H.; Hosseinzadeh, H. Cinnamon effects on metabolic syndrome: A review based on its mechanisms. Iran. J. Basic Med. Sci. 2016, 19, 1258. [Google Scholar] [PubMed]
- Liu, X.C.; Cheng, J.; Zhao, N.N.; Liu, Z.L. Insecticidal activity of essential oil of Cinnamomum cassia and its main constituent, trans-cinnamaldehyde, against the booklice, Liposcelis bostrychophila. Trop. J. Pharm. Res. 2014, 13, 1697–1702. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, S.F.; di Lorenzo, A.; Izadi, M.; Sobarzo-Sánchez, E.; Daglia, M.; Nabavi, S.M. Antibacterial effects of cinnamon: From farm to food, cosmetic and pharmaceutical industries. Nutrients 2015, 7, 7729–7748. [Google Scholar] [CrossRef] [PubMed]
- Youdim, M.B.; Bakhle, Y. Monoamine oxidase: Isoforms and inhibitors in Parkinson’s disease and depressive illness. Br. J. Pharm. 2006, 147, S287–S296. [Google Scholar] [CrossRef] [Green Version]
- Youdim, M.B.; Edmondson, D.; Tipton, K.F. The therapeutic potential of monoamine oxidase inhibitors. Nat. Rev. Neurosci. 2006, 7, 295–309. [Google Scholar] [CrossRef] [PubMed]
- McCormack, A.L.; Mak, S.K.; Shenasa, M.; Langston, W.J.; Forno, L.S.; di Monte, D.A. Pathologic modifications of α-synuclein in 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP)-treated squirrel monkeys. J. Neuropath Exp. Neur. 2008, 67, 793–802. [Google Scholar] [CrossRef]
- Ahmad, A.; Burns, C.S.; Fink, A.L.; Uversky, V.N. Peculiarities of copper binding to α-synuclein. J. Biomol. Struct. Dyn. 2012, 29, 825–842. [Google Scholar] [CrossRef]
- Mohammad-Beigi, H.; Shojaosadati, S.A.; Marvian, A.T.; Pedersen, J.N.; Klausen, L.H.; Christiansen, G.; Pedersen, J.S.; Dong, M.; Morshedi, D.; Otzen, D.E. Strong interactions with polyethylenimine-coated human serum albumin nanoparticles (PEI-HSA NPs) alter α-synuclein conformation and aggregation kinetics. Nanoscale 2015, 7, 19627–19640. [Google Scholar] [CrossRef]
- Silva, B.A.; Breydo, L.; Fink, A.L.; Uversky, V.N. Agrochemicals, α-synuclein, and Parkinson’s disease. Mol. Neurobiol. 2013, 47, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Schröder, M.; Kaufman, R.J. ER stress and the unfolded protein response. Mutat Res./Fund Mol. M. 2005, 569, 29–63. [Google Scholar]
- Ryu, E.J.; Harding, H.P.; Angelastro, J.M.; Vitolo, O.V.; Ron, D.; Greene, L.A. Endoplasmic reticulum stress and the unfolded protein response in cellular models of Parkinson’s disease. J. Neurosci. 2002, 22, 10690–10698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoozemans, J.; van Haastert, E.; Eikelenboom, P.; de Vos, R.; Rozemuller, J.; Scheper, W. Activation of the unfolded protein response in Parkinson’s disease. Biochem. Biophys. Res. Commun. 2007, 354, 707–711. [Google Scholar] [CrossRef]
- Jiang, P.; Gan, M.; Ebrahim, A.S.; Lin, W.-L.; Melrose, H.L.; Yen, S.-H.C. ER stress response plays an important role in aggregation of α-synuclein. Mol. Neurodegener. 2010, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gardner, B.M.; Pincus, D.; Gotthardt, K.; Gallagher, C.M.; Walter, P. Endoplasmic reticulum stress sensing in the unfolded protein response. Cold Spring Harb Perspect. Biol. 2013, 5, a013169. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S. Endoplasmic reticulum stress and atherosclerosis. Nat. Med. 2010, 16, 396–399. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef] [Green Version]
- Todd, D.J.; Lee, A.-H.; Glimcher, L.H. The endoplasmic reticulum stress response in immunity and autoimmunity. Nat. Rev. Immunol. 2008, 8, 663–674. [Google Scholar] [CrossRef]
- Senft, D.; Ze’ev, A.R. UPR, autophagy, and mitochondria crosstalk underlies the ER stress response. Trends Biochem. Sci. 2015, 40, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Yorimitsu, T.; Nair, U.; Yang, Z.; Klionsky, D.J. Endoplasmic reticulum stress triggers autophagy. J. Biol. Chem. 2006, 281, 30299–30304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Pattingre, S.; Sinha, S.; Bassik, M.; Levine, B. JNK1-mediated phosphorylation of Bcl-2 regulates starvation-induced autophagy. Mol. Cell 2008, 30, 678–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabuto, H.; Nishizawa, M.; Tada, M.; Higashio, C.; Shishibori, T.; Kohno, M. Zingerone [4-(4-hydroxy-3-methoxyphenyl)-2-butanone] prevents 6-hydroxydopamine-induced dopamine depression in mouse striatum and increases superoxide scavenging activity in serum. Neurochem. Res. 2005, 30, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Kabuto, H.; Tada, M.; Kohno, M. Eugenol [2-methoxy-4-(2-propenyl) phenol] prevents 6-hydroxydopamine-induced dopamine depression and lipid peroxidation inductivity in mouse striatum. Biol. Pharm. Bull. 2007, 30, 423–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siqueira-Lima, P.S.; Brito, R.G.; Araújo-Filho, H.G.; Santos, P.L.; Lucchesi, A.; Araújo, A.A.; Menezes, P.P.; Scotti, L.; Scotti, M.T.; Menezes, I.R. Anti-hyperalgesic effect of Lippia grata leaf essential oil complexed with β-cyclodextrin in a chronic musculoskeletal pain animal model: Complemented with a molecular docking and antioxidant screening. Biomed. Pharm. 2017, 91, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.A.; Guimarães, A.G.; Araújo, A.A.; Quintans-Júnior, L.J.; Quintans, J.S. New drugs or alternative therapy to blurring the symptoms of fibromyalgia—a patent review. Expert Opin. Pat. 2017, 27, 1147–1157. [Google Scholar] [CrossRef]
- Araújo-Filho, H.G.; Pereira, E.W.; Rezende, M.M.; Menezes, P.P.; Araújo, A.A.; Barreto, R.S.; Martins, A.O.; Albuquerque, T.R.; Silva, B.A.; Alcantara, I.S. D-limonene exhibits superior antihyperalgesic effects in a β-cyclodextrin-complexed form in chronic musculoskeletal pain reducing Fos protein expression on spinal cord in mice. Neuroscience 2017, 358, 158–169. [Google Scholar] [CrossRef]
- Santos, P.L.; Brito, R.G.; Quintans, J.S.S.; Araujo, A.A.S.; Menezes, I.R.A.; Brogden, N.K.; Quintans-Junior, L.J. Cyclodextrins as complexation agents to improve the anti-inflammatory drugs profile: A systematic review and meta-analysis. In Curr. Pharm. Des.; 2017; Volume 23, pp. 2096–2107. [Google Scholar]
- Brito, R.G.; Araujo, A.A.; Quintans, J.S.; Sluka, K.A.; Quintans-Junior, L.J. Enhanced analgesic activity by cyclodextrins–a systematic review and meta-analysis. Expert Opin Drug Del. 2015, 12, 1677–1688. [Google Scholar] [CrossRef]
- Andrade, A.M.; Oliveira, J.P.; Santos, A.L.; Franco, C.R.; Antoniolli, Â.R.; Estevam, C.S.; Thomazzi, S.M. Preliminary study on the anti-inflammatory and antioxidant activities of the leave extract of Hyptis fruticosa Salzm. ex Benth., Lamiaceae. Rev. Bras. Farm. 2010, 20, 962–968. [Google Scholar] [CrossRef] [Green Version]
- De Lima, A.C.B.; Paixão, M.S.; Melo, M.; de Santana, M.T.; Damascena, N.P.; Dias, A.S.; Porto, Y.C.B.S.; Fernandes, X.A.; Santos, C.C.S.; Lima, C.A.; et al. Orofacial antinociceptive effect and antioxidant properties of the hydroethanol extract of Hyptis fruticosa salmz ex Benth. J. Ethnopharmacol. 2013, 146, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Tadaiesky, M.T.; Andreatini, R.; Vital, M.A. Different effects of 7-nitroindazole in reserpine-induced hypolocomotion in two strains of mice. Eur. J. Pharm. 2006, 535, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Ben, J.; Soares, F.M.; Scherer, E.B.; Cechetti, F.; Netto, C.A.; Wyse, A.T. Running exercise effects on spatial and avoidance tasks in ovariectomized rats. Neurobiol. Learn. Mem. 2010, 94, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.L.; Zhang, Q.G.; Han, D.; Desai, B.N.; Brann, D.W. Long-term estrogen deprivation leads to elevation of Dickkopf-1 and dysregulation of Wnt/β-Catenin signaling in hippocampal CA1 neurons. Steroids 2013, 78, 624–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkaki, A.; Badavi, M.; Hoseiny, N.; Gharibnaseri, M.K.; Rahim, F. Postmenopausal effects of intrastriatal estrogen on catalepsy and pallidal electroencephalogram in an animal model of Parkinson’s disease. Neuroscience 2008, 154, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Namavar Jahromi, B.; Tartifizadeh, A.; Khabnadideh, S. Comparison of fennel and mefenamic acid for the treatment of primary dysmenorrhea. Int. J. Gynaecol. Obs. 2003, 80, 153–157. [Google Scholar] [CrossRef]
- Modaress Nejad, V.; Asadipour, M. Comparison of the effectiveness of fennel and mefenamic acid on pain intensity in dysmenorrhoea. East. Mediterr. Health J. 2006, 12, 423–427. [Google Scholar]
- Betarbet, R.; Sherer, T.B.; MacKenzie, G.; Garcia-Osuna, M.; Panov, A.V.; Greenamyre, J.T. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat. Neurosci. 2000, 3, 1301–1306. [Google Scholar] [CrossRef]
- Talpade, D.J.; Greene, J.G.; Higgins, D.S., Jr.; Greenamyre, J.T. In vivo labeling of mitochondrial complex I (NADH:ubiquinone oxidoreductase) in rat brain using [(3)H]dihydrorotenone. J. Neurochem. 2000, 75, 2611–2621. [Google Scholar] [CrossRef] [Green Version]
- Sherer, T.B.; Richardson, J.R.; Testa, C.M.; Seo, B.B.; Panov, A.V.; Yagi, T.; Matsuno-Yagi, A.; Miller, G.W.; Greenamyre, J.T. Mechanism of toxicity of pesticides acting at complex I: Relevance to environmental etiologies of Parkinson’s disease. J. Neurochem. 2007, 100, 1469–1479. [Google Scholar] [CrossRef]
- Liu, L.-L.; Yang, J.-L.; Shi, Y.-P. Phytochemicals and biological activities of Pulicaria species. Chem. Biodivers. 2010, 7, 327–349. [Google Scholar] [CrossRef]
- Lee, K.S.; Lee, J.K.; Kim, H.G.; Kim, H.R. Differential Effects of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine on Motor Behavior and Dopamine Levels at Brain Regions in Three Different Mouse Strains. Korean J. Physiol. Pharm. 2013, 17, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worwood, V.A. Aromatherapy for the Healthy Child.: More Than 300 Natural, Nontoxic, and Fragrant Essential Oil Blends, 3rd ed.; New World Library: Novato, CA, USA, 2000; p. 384. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| No. | Disease | Study | Source of Essential Oils | Ref | Method | Findings | Conclusions and/or Recommendations |
|---|---|---|---|---|---|---|---|
| 1. | ALZHEIMER’S DISEASE | In vitro | Ajuga chamaecistus subsp scoparia (Boiss.) Rech.f. | [64] |
|
|
|
| 2. | Allium tuncelianum (Amaryllidaceae) | [65] |
|
|
| ||
| 3. | Aloysia citrodora Palau | [66] |
|
|
| ||
| 4. | ALZHEIMER’S DISEASE | In vitro | Artemisia macrocephala | [26] |
|
|
|
| 5. | Artemisia maderaspatana | [67] |
|
|
| ||
| 6. | Artemisia absinthium | [27] |
|
|
| ||
| 7. | ALZHEIMER’S DISEASE | In vitro | Boswellia dalzielii | [68] |
|
|
|
| 8. | Chamaecyparis obtusa Cryptomeria japonica | [69] |
|
|
| ||
| 9. | Cinnamomum tamala (Buch.-Ham.) | [18] |
|
|
| ||
| 10. | ALZHEIMER’S DISEASE | In vitro | Cinnamomum zeylanicum | [19] |
|
|
|
| 11. | Citrus Sinensis [L.] Osbeck | [70] |
|
|
| ||
| 12. | ALZHEIMER’S DISEASE | In vitro | Citrus aurantifolia Swingle, C. aurantium L. C. bergamia Risso and Poit. | [71] |
|
|
|
| 13. | Cistus creticus Cistus salvifolius Cistus libanotis Cistus monspeliensis Cistus villo-sus | [72] |
|
|
| ||
| 14. | Clinopodium serpyllifolium (M.Bieb.) Kuntze | [73] |
|
|
| ||
| 15. | Daucus aristidis Coss. | [74] |
|
|
| ||
| 16. | ALZHEIMER’S DISEASE | In vitro | Hedychium gardnerianum,Sheppard ex Ker-Gawl | [75] |
|
|
|
| 17. | Hymenocrater bituminous | [76] |
|
|
| ||
| 18. | Illicium verum Hook.f | [77] |
|
|
| ||
| 19. | ALZHEIMER’S DISEASE | In vitro | Lavandula luisieri | [78] |
|
|
|
| 20. | Lavandula angustifolia | [24] |
|
|
| ||
| 21. | Lavandula pubescens | [23] |
|
|
| ||
| 22. | Lavandula viridis L’Her | [79] |
|
|
| ||
| 23. | ALZHEIMER’S DISEASE | In vitro | Mentha spicata L. | [80] |
|
|
|
| 24. | Neomitranthes obscura (DC.) N. Silveira | [81] |
|
|
| ||
| 25. | ALZHEIMER’S DISEASE | In vitro | Origanum rotundifolium Boiss. | [82] |
|
|
|
| 26. | Panax ginseng Panax japonicus P. notoginseng P. quinquefolius | [83] |
|
|
| ||
| 27. | Pinus sp. | [84] |
|
|
| ||
| 28. | ALZHEIMER’S DISEASE | In vitro | Piper divaricatum | [85] |
|
|
|
| 29. | Polygonum hydropiper L. | [22] |
|
|
| ||
| 30. | Prangos gaubae | [86] |
|
|
| ||
| 31. | ALZHEIMER’S DISEASE | In vitro | Rumex hastatus D. Don | [87] |
|
|
|
| 32. | Salvia officinalis | [88] |
|
|
| ||
| 33. | Salvia chionantha | [89] |
|
|
| ||
| 34. | Salvia chrysophylla Staph | [90] |
|
|
| ||
| 35. | ALZHEIMER’S DISEASE | In vitro | Salvia urmiensis | [20] |
|
|
|
| 36. | Salvia nemorosa L. | [91] |
|
|
| ||
| 37. | Salvia officinalis L. | [21] |
|
|
| ||
| 38. | ALZHEIMER’S DISEASE | In vitro | Salvia syriaca L. | [92] |
|
|
|
| 39. | Satureja thymbra L. | [93] |
|
|
| ||
| 40. | Stachys inflata, S. lavandulifolia, S. byzantina | [94] |
|
|
| ||
| 41. | ALZHEIMER’S DISEASE | In vitro | Sideritis galatica Bornm. | [95] |
|
|
|
| 42. | S. ballsiana S. cyanescens S. divaricata S. hydrangea S. kronenburgii S. macrochlamys S. nydeggeri S. pachystachys S. pseudeuphratica S. rusellii | [96] |
|
|
| ||
| 43. | Thymus mastichina L. | [97] |
|
|
| ||
| 44. | ALZHEIMER’S DISEASE | In vitro | Thymus haussknechtii Velen. | [98] |
|
|
|
| 45. | Thymus lotocephalus | [99] |
|
|
| ||
| 46. | ALZHEIMER’S DISEASE | In vitro | Zingiber cassumunar | [100] |
|
|
|
| 47. | Zosima absinthifolia Link | [14] |
|
|
| ||
| 48. | ALZHEIMER’S DISEASE | In vitro | Calamintha nepeta, Foeniculum vulgare, Mentha spicata and Thymus mastichina | [101] |
|
|
|
| 49. | Cymbopogon citratus (Gramineae), Citrus hystrix (Rutaceae) and Zingiber cassumunar (Zingiberaceae) | [102] |
|
|
| ||
| 50. | Alpinia galanga Linn., Centella asiatica Urban., Cinnamomum bejolghota (Buch. Ham.) Sweet, Citrus aurantifolia Swing, Citrus hystrix DC., Citrus maxima (Burm.) Merr., Citrus reticulata Blanco cv. Shogun, Citrus reticulata var. Fremont, Cymbopogon citratus Stapf., Eupatorium odoratum Linn,. Melissa officinalis Linn., Ocimum basilicum Linn., Ocimum canum Sims., Ocimum gratissimum Linn., Ocimum sanctum Linn., Piper sarmentosum Roxb., Polygonum odoratum Lour., Polyscias fruticosa, Harms.Zingiber cassumunar Roxb,. Zingiber officinale Rosco | [25] |
|
|
| ||
| 51. | ALZHEIMER’S DISEASE | In vitro | Lavandula angustifolia Coriandrum sativum | [103] |
|
|
|
| 52. | Artemisia annua L. (Asteraceae) Glycyrrhiza glabra L. (Fabaceae) | [104] |
|
|
| ||
| 53. | ALZHEIMER’S DISEASE | In vitro | Angelica sinensis (Oliv) Diels. Ligusticum Chuanxiong Hort | [105] |
|
|
|
| 54. | In vivo | Achillea biebersteinii (Asteraceae) | [106] |
|
|
| |
| 55. | ALZHEIMER’S DISEASE | In vivo | Boswellia serrata (Burseraceae) | [107] |
|
|
|
| 56. | Chamaecyparis obtuse | [108] |
|
|
| ||
| 57. | ALZHEIMER’S DISEASE | In vivo | Citrus limon | [109] |
|
|
|
| 58. | ALZHEIMER’S DISEASE | In vivo | Cymbopogon giganteus Illicium pachyphyllum Carum carvi | [110] |
|
|
|
| 59. | Ferulago angulata | [111] |
|
|
| ||
| 60. | ALZHEIMER’S DISEASE | In vivo | Lavandula angustifolia ssp. angustifolia Mill. (Lamiaceae) Lavandula hybrida Rev. (Lamiaceae) | [112] |
|
|
|
| 61. | ALZHEIMER’S DISEASE | In vivo | Ocimum basilicum | [113] |
|
|
|
| 62. | Ocimum sanctum L. Ocimum basilicum L. | [114] |
|
|
| ||
| 63. | ALZHEIMER’S DISEASE | In vivo | Pimpinella peregrina | [115] |
|
|
|
| 64. | Pinus halepensis | [116] |
|
|
| ||
| 65. | ALZHEIMER’S DISEASE | In vivo | Kushui rose (Rosa setate × Rosa rugosa) | [117] |
|
|
|
| 66. | ALZHEIMER’S DISEASE | In vivo | Rosmarinus officinalis | [118] |
|
|
|
| 67. | Salvia miltiorrhiza Bge | [119] |
|
|
| ||
| 68. | ALZHEIMER’S DISEASE | In vivo | Schisandra chinensis | [120] |
|
|
|
| 69. | SuHeXiang Wan (SHXW) | [121] |
|
|
| ||
| 70. | ALZHEIMER’S DISEASE | In vivo | Tetraclinis articulata | [122] |
|
|
|
| 71. | Zataria multiflora Boiss. | [123] |
|
|
| ||
| 72. | ALZHEIMER’S DISEASE | In vivo | Z. multiflora Boiss. | [124] |
|
|
|
| 73. | Origanum vulgare Thymus vulgaris | [125] |
|
|
| ||
| 74. | ALZHEIMER’S DISEASE | In vitro and In vivo | Acori graminei | [126] |
|
|
|
| 75. | ALZHEIMER’S DISEASE | In vitro and In vivo | Foeniculi vulgare aetheroleum | [127] |
|
|
|
| 76. | Lavandula angustifolia Mill. | [128] |
|
|
| ||
| 77. | ALZHEIMER’S DISEASE | In vitro and In vivo | Lavandula angustifolia Mill. | [129] |
|
|
|
| 78. | Lavandula luisieri | [130] |
|
|
| ||
| 79. | ALZHEIMER’S DISEASE | In vitro and In vivo | Mentha longifolia | [131] |
|
|
|
| 80. | Salviae aetheroleum | [8] |
|
|
| ||
| 81. | SHXW | [132] |
|
|
| ||
| 82. | ALZHEIMER’S DISEASE | In vitro and In vivo | Zelkova serrata | [133] |
|
|
|
| 83. | ALZHEIMER’S DISEASE | In vitro and In vivo | Zingiber officinale | [134] |
|
|
|
| 84. | Pistacia khinjuk Allium sativum | [135] |
|
|
| ||
| 85. | ALZHEIMER’S DISEASE | In vitro and Ex vivo | Salvia sp | [136] |
|
|
|
| 86. | Syzygium aromaticum Xylopia aethiopica | [137] |
|
|
| ||
| 87. | PARKINSON’S DISEASE | In vitro | Cinnamomum verum Cinnamomum cassia | [138] |
|
|
|
| 88. | Eryngium sp. | [139] |
|
|
| ||
| 89. | PARKINSON’S DISEASE | In vitro | Myrtus communis Ferula gummosa Eucalyptus globulus Satureja hortensis Rosmarinus officinalis Mentha spicata Mentha piperita Cuminum cyminum Artemisia dracunculus Citrus sinensis Citrus limonum Thymus vulgaris Lavandula officinalis Mentha pulegium Foeniculum vulgare | [140] |
|
|
|
| 90. | Cuminum cyminum | [141] |
|
|
| ||
| 91. | PARKINSON’S DISEASE | In vivo | Acorus tatarinowii Schott (Shi Chang Pu) | [142] |
|
|
|
| 92. | Acorus tatarinowii Schott (Shi Chang Pu) | [143] |
|
|
| ||
| 93. | PARKINSON’S DISEASE | In vivo | Eplingiella fruticosa | [144] |
|
|
|
| 94. | Foeniculum vulgare | [145] |
|
|
| ||
| 95. | PARKINSON’S DISEASE | In vivo | Pulicaria undulata | [146] |
|
|
|
| 96. | Zingiber officinale Syzygium aromaticum | [28] |
|
|
| ||
| 97. | PARKINSON’S DISEASE | In vitro and In vivo | SHXW | [147] |
|
|
|
| 98. | PARKINSON’S DISEASE | In vivo and ex vivo | Rosa damascena Mill. Lavandula angustifolia Mill, | [148] |
|
|
|
| 99. | PARKINSON’S DISEASE | In vivo and ex vivo | Rosa Damascena Mill. | [149] |
|
|
|
| 100. | OTHER NEURODEGENERATIVE DISEASE | In vitro | Lavandula pedunculata subsp lusitanica (Chaytor) Franco | [150] |
|
|
|
| 101. | Pelargonium graveolens | [151] |
|
|
| ||
| 102. | OTHER NEURODEGENERATIVE DISEASE | In vitro | Myrcianthes myrsinoides (Kunth) Grifo Myrcia mollis (Kunth) DC. (Myrtaceae) | [152] |
|
|
|
| 103. | OTHER NEURODEGENERATIVE DISEASE | In vivo | Anthriscus nemorosa | [153] |
|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd Rashed, A.; Abd Rahman, A.Z.; Rathi, D.N.G. Essential Oils as a Potential Neuroprotective Remedy for Age-Related Neurodegenerative Diseases: A Review. Molecules 2021, 26, 1107. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26041107
Abd Rashed A, Abd Rahman AZ, Rathi DNG. Essential Oils as a Potential Neuroprotective Remedy for Age-Related Neurodegenerative Diseases: A Review. Molecules. 2021; 26(4):1107. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26041107
Chicago/Turabian StyleAbd Rashed, Aswir, Ahmad Zuhairi Abd Rahman, and Devi Nair Gunasegavan Rathi. 2021. "Essential Oils as a Potential Neuroprotective Remedy for Age-Related Neurodegenerative Diseases: A Review" Molecules 26, no. 4: 1107. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26041107