
Suramin Targets the Conserved Ligand-Binding Pocket of Human Raf1 Kinase Inhibitory Protein

Abstract
:1. Introduction
2. Results and Discussion
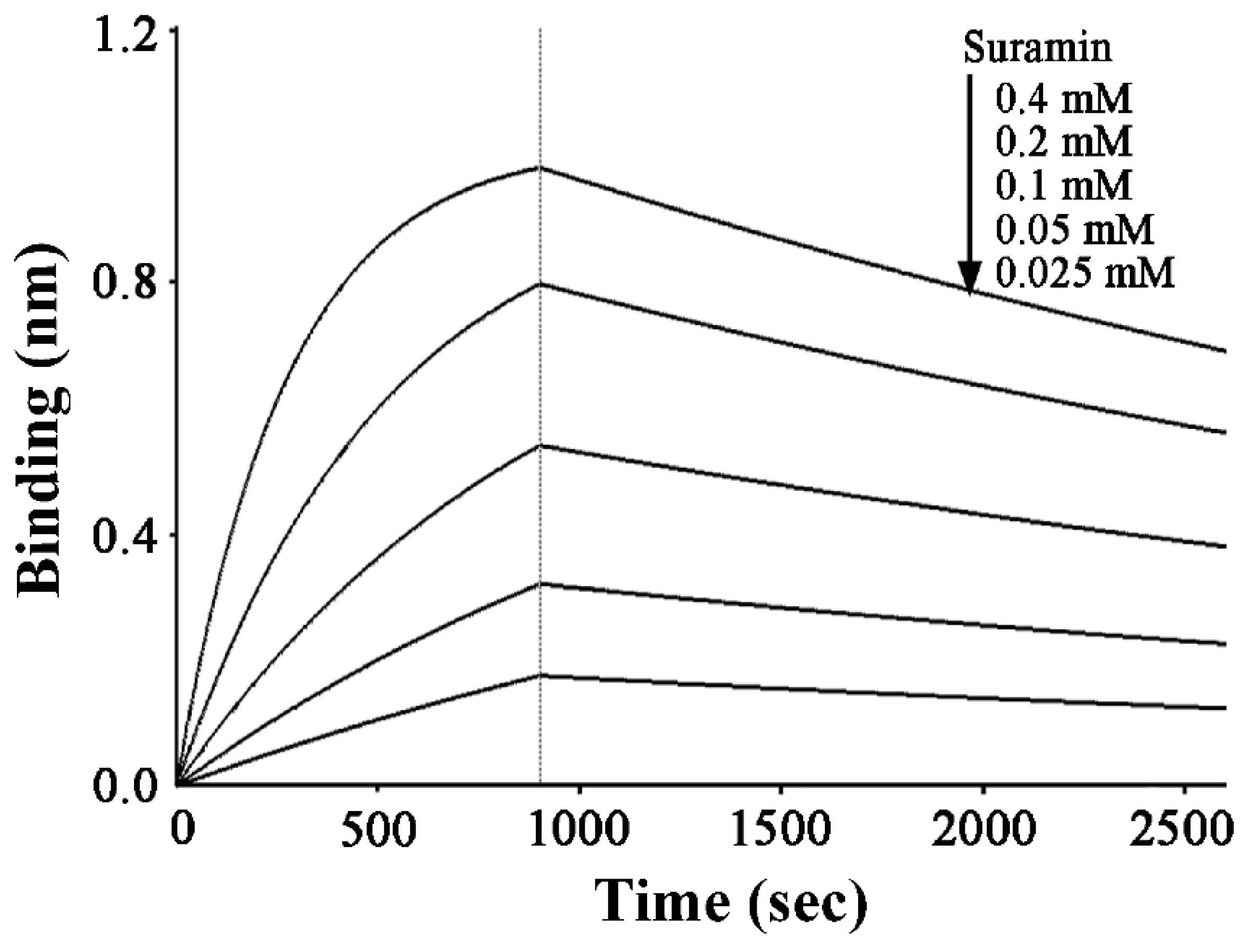
2.1. Suramin has a Micromolar Affinity for hRKIP
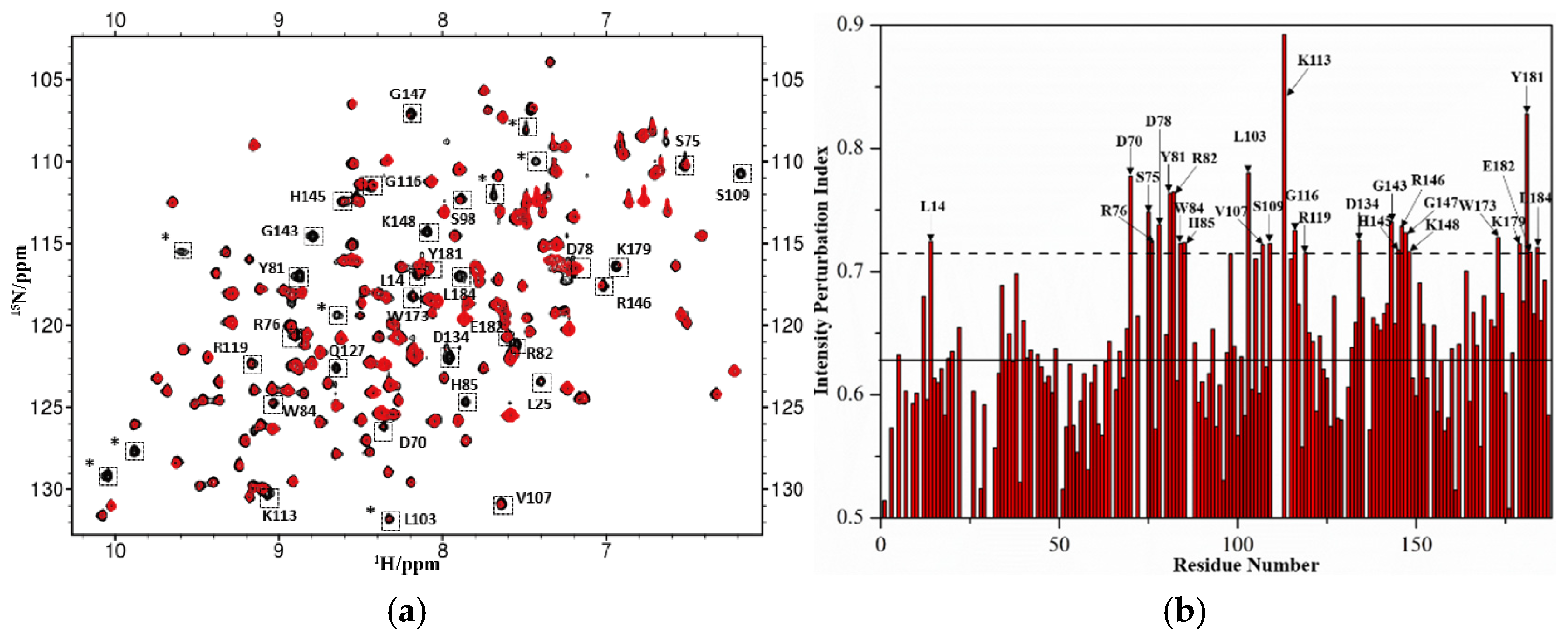
2.2. Identification of Suramin Binding Sites on hRKIP
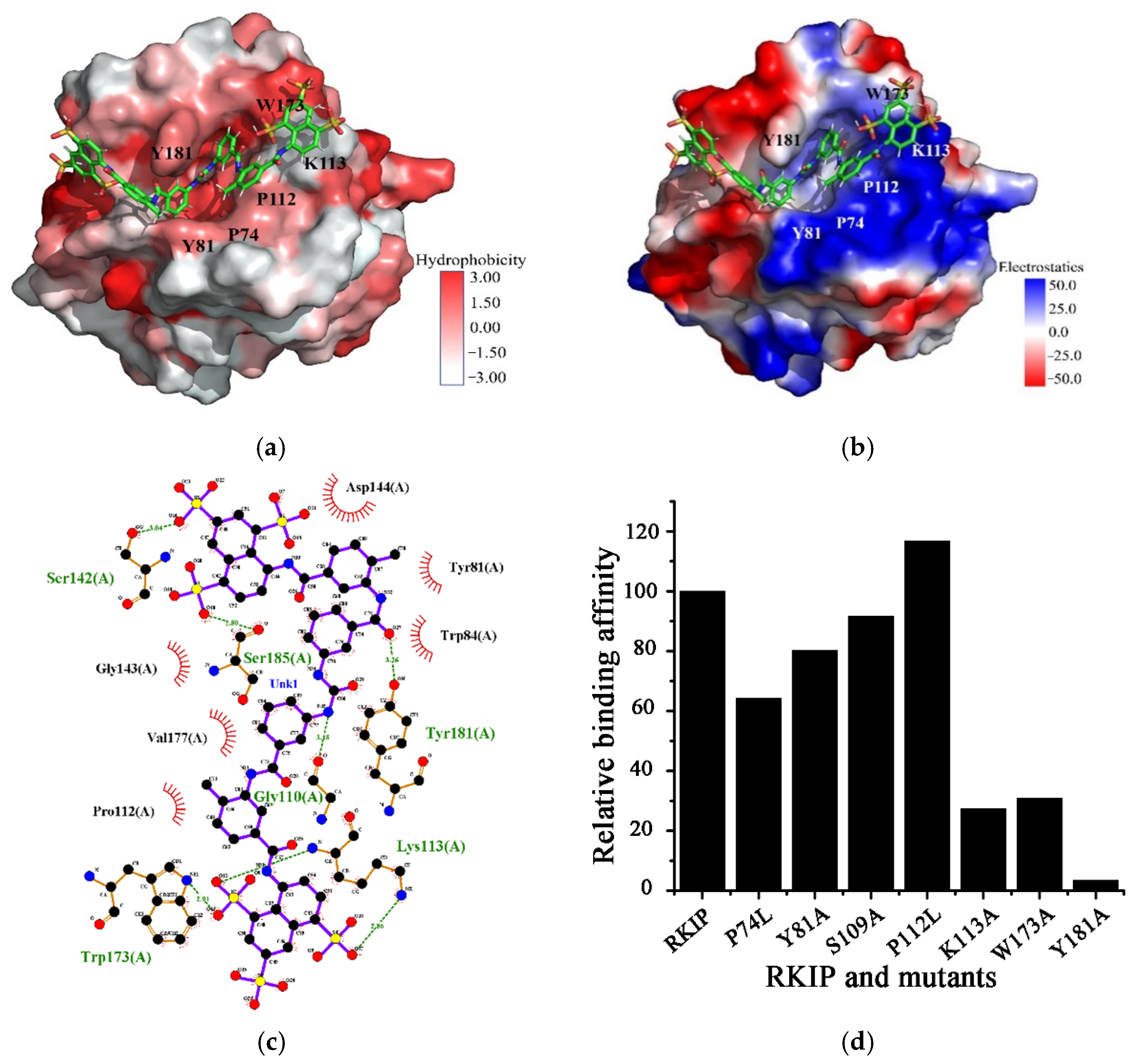
2.3. Structural Model of Suramin Binding to hRKIP
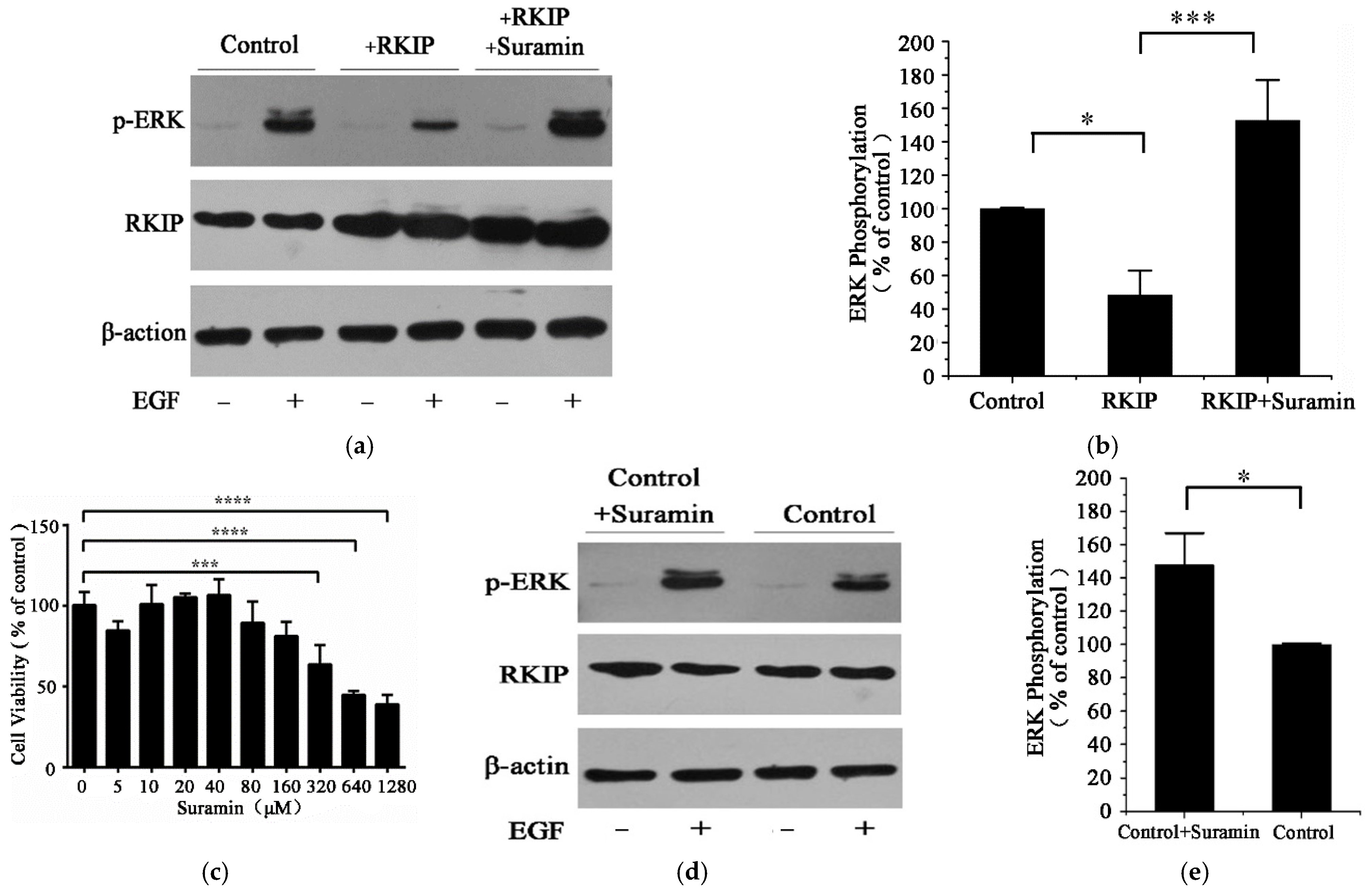
2.4. Suramin Enhances the Phosphorylation of ERK
3. Materials and Methods
3.1. Protein Expression and Purification
3.2. Biolayer Interferometry (BLI) Assays
3.3. Nuclear Magnetic Resonance (NMR) Spectroscopy
3.4. Molecular Docking
3.5. ERK Phosphorylation Analyses
3.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Hawking, F. Suramin: With special reference to onchocerciasis. Adv. Pharm. Chemother. 1978, 15, 289–322. [Google Scholar]
- Broder, S.; Yarchoan, R.; Collins, J.M.; Lane, H.C.; Markham, P.D.; Klecker, R.W.; Redfield, R.R.; Mitsuya, H.; Hoth, D.F.; Gelmann, E.; et al. Effects of suramin on HTLV-III/LAV infection presenting as Kaposi’s sarcoma or AIDS-related complex: Clinical pharmacology and suppression of virus replication in vivo. Lancet 1985, 2, 627–630. [Google Scholar] [CrossRef]
- Cheson, B.D.; Levine, A.M.; Mildvan, D.; Kaplan, L.D.; Wolfe, P.; Rios, A.; Groopman, J.E.; Gill, P.; Volberding, P.A.; Poiesz, B.J.; et al. Suramin Therapy in Aids and Related Disorders—Report of the United-States-Suramin-Working-Group. JAMA 1987, 258, 1347–1351. [Google Scholar] [CrossRef]
- Walther, M.M.; Figg, W.D.; Linehan, W.M. Intravesical suramin: A novel agent for the treatment of superficial transitional-cell carcinoma of the bladder. World J. Urol. 1996, 14, S8–S11. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.A.; Larocca, R.V.; Thomas, R.; Mcatee, N.; Myers, C.E. Suramin—An Anticancer Drug with a Unique Mechanism of Action. J. Clin. Oncol. 1989, 7, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Walz, T.M.; Abdiu, A.; Wingren, S.; Smeds, S.; Larsson, S.E.; Wasteson, A. Suramin Inhibits Growth of Human Osteosarcoma Xenografts in Nude-Mice. Cancer Res. 1991, 51, 3585–3589. [Google Scholar] [PubMed]
- Myers, C.; Cooper, M.; Stein, C.; Larocca, R.; Walther, M.M.; Weiss, G.; Choyke, P.; Dawson, N.; Steinberg, S.; Uhrich, M.M.; et al. Suramin—A Novel Growth-Factor Antagonist with Activity in Hormone-Refractory Metastatic Prostate-Cancer. J. Clin. Oncol. 1992, 10, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Foekens, J.A.; Sieuwerts, A.M.; Stuurman-Smeets, E.M.; Dorssers, L.C.; Berns, E.M.; Klijn, J.G. Pleiotropic actions of suramin on the proliferation of human breast-cancer cells in vitro. Int. J. Cancer 1992, 51, 439–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokshin, A.; Peng, X.J.; Campbell, P.G.; Barsouk, A.; Levitt, M.L. Mechanisms of growth stimulation by suramin in non-small-cell lung cancer cell lines. Cancer Chemoth. Pharm. 1999, 43, 341–347. [Google Scholar] [CrossRef]
- Morocz, I.A.; Lauber, B.; Schmitter, D.; Stahel, R.A. In vitro effect of suramin on lung tumour cells. Eur. J. Cancer 1993, 29A, 245–247. [Google Scholar] [CrossRef]
- Rubio, G.J.; Pinedo, H.M.; Virizuela, J.; van Ark-Otte, J.; Giaccone, G. Effects of suramin on human lung cancer cell lines. Eur. J. Cancer 1995, 31A, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Kaur, M.; Reed, E.; Sartor, O.; Dahut, W.; Figg, W.D. Suramin’s development: What did we learn? Investig. New Drugs 2002, 20, 209–219. [Google Scholar] [CrossRef]
- Stein, C.A. Suramin: A novel antineoplastic agent with multiple potential mechanisms of action. Cancer Res. 1993, 53, 2239–2248. [Google Scholar] [PubMed]
- Bojanowski, K.; Lelievre, S.; Markovits, J.; Couprie, J.; Jacquemin-Sablon, A.; Larsen, A.K. Suramin is an inhibitor of DNA topoisomerase II in vitro and in Chinese hamster fibrosarcoma cells. Proc. Natl. Acad. Sci. USA 1992, 89, 3025–3029. [Google Scholar] [CrossRef] [Green Version]
- Castro, E.; Mateo, J.; Tome, A.R.; Barbosa, R.M.; Miras-Portugal, M.T.; Rosario, L.M. Cell-specific purinergic receptors coupled to Ca2+ entry and Ca2+ release from internal stores in adrenal chromaffin cells. Differential sensitivity to UTP and suramin. J. Biol. Chem. 1995, 270, 5098–5106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horne, M.K., III; Stein, C.A.; LaRocca, R.V.; Myers, C.E. Circulating glycosaminoglycan anticoagulants associated with suramin treatment. Blood 1988, 71, 273–279. [Google Scholar] [PubMed]
- Jindal, H.K.; Anderson, C.W.; Davis, R.G.; Vishwanatha, J.K. Suramin affects DNA synthesis in HeLa cells by inhibition of DNA polymerases. Cancer Res. 1990, 50, 7754–7757. [Google Scholar] [PubMed]
- Pommier, G.J.; Garrouste, F.L.; el Atiq, F.; Roccabianca, M.; Marvaldi, J.L.; Remacle-Bonnet, M.M. Potential autocrine role of insulin-like growth factor II during suramin-induced differentiation of HT29-D4 human colonic adenocarcinoma cell line. Cancer Res. 1992, 52, 3182–3188. [Google Scholar]
- Schrell, U.M.; Gauer, S.; Kiesewetter, F.; Bickel, A.; Hren, J.; Adams, E.F.; Fahlbusch, R. Inhibition of proliferation of human cerebral meningioma cells by suramin: Effects on cell growth, cell cycle phases, extracellular growth factors, and PDGF-BB autocrine growth loop. J. Neurosurg. 1995, 82, 600–607. [Google Scholar] [CrossRef]
- Wiedemar, N.; Hauser, D.A.; Maser, P. 100 Years of Suramin. Antimicrob. Agents Chemother. 2020, 64, e01168-19. [Google Scholar] [CrossRef]
- Trapp, J.; Meier, R.; Hongwiset, D.; Kassack, M.U.; Sippl, W.; Jung, M. Structure-activity studies on suramin analogues as inhibitors of NAD(+)-dependent histone deacetylases (Sirtuins). Chemmedchem 2007, 2, 1419–1431. [Google Scholar] [CrossRef]
- Zhang, X.Z.; Shi, S.D.; Su, Y.; Yang, X.L.; He, S.N.; Yang, X.Y.; Wu, J.; Zhang, J.; Rao, F. Suramin and NF449 are IP5K inhibitors that disrupt inositol hexakisphosphate-mediated regulation of cullin-RING ligase and sensitize cancer cells to MLN4924/pevonedistat. J. Biol. Chem. 2020, 295, 10281–10292. [Google Scholar] [CrossRef]
- Guo, C.; Yi, C.; Peng, Y.; Wen, Y.; Lin, D. Solution structure and backbone dynamics of human Raf-1 kinase inhibitor protein. Biochem. Biophys. Res. Commun. 2013, 438, 129–132. [Google Scholar] [CrossRef]
- Granovsky, A.E.; Clark, M.C.; McElheny, D.; Heil, G.; Hong, J.; Liu, X.D.; Kim, Y.; Joachimiak, G.; Joachimiak, A.; Koide, S.; et al. Raf Kinase Inhibitory Protein Function Is Regulated via a Flexible Pocket and Novel Phosphorylation-Dependent Mechanism. Mol. Cell. Biol. 2009, 29, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Santos, E.; Crespo, P. The RAS-ERK pathway: A route for couples. Sci. Signal. 2018, 11, eaav0917. [Google Scholar] [CrossRef] [PubMed]
- Tidyman, W.E.; Rauen, K.A. The RASopathies: Developmental syndromes of Ras/MAPK pathway dysregulation. Curr. Opin. Genet. Dev. 2009, 19, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Auer, J.S.; Nagel, A.C.; Schulz, A.; Wahl, V.; Preiss, A. MAPK-dependent phosphorylation modulates the activity of Suppressor of Hairless in Drosophila. Cell Signal. 2015, 27, 115–124. [Google Scholar] [CrossRef]
- Cheng, X.; Wan, Y.; Xu, Y.; Zhou, Q.; Wang, Y.; Zhu, H. Melatonin alleviates myosin light chain kinase expression and activity via the mitogen-activated protein kinase pathway during atherosclerosis in rabbits. Mol. Med. Rep. 2015, 11, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Li, P.; Li, P.; Xu, Y.; Li, Y.; Tang, B. Isoquercitrin inhibits the progression of pancreatic cancer in vivo and in vitro by regulating opioid receptors and the mitogen-activated protein kinase signalling pathway. Oncol. Rep. 2015, 33, 840–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, D.; Zeng, X.X.; Liu, R.H.; Ye, H. Potential mechanism of mycelium polysaccharide from Pholiota dinghuensis Bi in regulating the proliferation and apoptosis of human breast cancer MCF-7 cells through p38/MAPK pathway. J. Funct. Foods 2015, 12, 375–388. [Google Scholar] [CrossRef]
- Kim, D.S.; Shin, M.R.; Kim, Y.S.; Bae, W.J.; Roh, D.H.; Hwang, Y.S.; Kim, E.C. Anti-inflammatory effects of glutamine on LPS-stimulated human dental pulp cells correlate with activation of MKP-1 and attenuation of the MAPK and NF-kappa B pathways. Int. Endod. J. 2015, 48, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Korashy, H.M.; Al-Suwayeh, H.A.; Maayah, Z.H.; Ansari, M.A.; Ahmad, S.F.; Bakheet, S.A. Mitogen-Activated Protein Kinases Pathways Mediate the Sunitinib-Induced Hypertrophy in Rat Cardiomyocyte H9c2 Cells. Cardiovasc. Toxicol. 2015, 15, 41–51. [Google Scholar] [CrossRef]
- Pastuhov, S.I.; Hisamoto, N.; Matsumoto, K. MAP kinase cascades regulating axon regeneration in C. elegans. P. Jpn. Acad. B-Phys. 2015, 91, 63–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.; Chang, T.; Sun, T.; Wu, Z.; Dai, Y.; Yao, H.; Lin, D. Anti-leprosy drug Clofazimine binds to human Raf1 kinase inhibitory protein and enhances ERK phosphorylation. Acta Biochim. Biophys. Sin. 2018, 50, 1062–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, C.; Peng, Y.; Guo, C.; Lin, D. 1H, 13C, 15N backbone and side-chain resonance assignments of the human Raf-1 kinase inhibitor protein. Biomol. Nmr. Assign. 2011, 5, 63–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Fu, C.; Shi, L.; Ruan, L.; Lin, D.; Guo, C. Structural basis for RKIP binding with its substrate Raf1 kinase. Biotechnol. Lett. 2014, 36, 1869–1874. [Google Scholar] [CrossRef]
- Schoentgen, F.; Jolles, P. From structure to function: Possible biological roles of a new widespread protein family binding hydrophobic ligands and displaying a nucleotide binding site. FEBS Lett. 1995, 369, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C. Molecular Recognition of Receptor-Sites Using a Genetic Algorithm with a Description of Desolvation. J. Mol. Biol. 1995, 245, 43–53. [Google Scholar] [CrossRef]
- Korb, O.; Stutzle, T.; Exner, T.E. Empirical scoring functions for advanced protein-ligand docking with PLANTS. J. Chem. Inf. Model. 2009, 49, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wu, Z.; Luo, M.; Lin, D.; Guo, C. Pranlukast, a novel binding ligand of human Raf1 kinase inhibitory protein. Biotechnol. Lett. 2016, 38, 1375–1380. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | KD (×10−5 M) | kon (M−1s−1) | koff (×10−4 s−1) | kobs (×10−4 s−1) | Full R2 (×10−1) |
|---|---|---|---|---|---|
| RKIP | 2.38 | 8.69 | 2.07 | 36.8 | 9.68 |
| P74L | 3.71 | 7.39 | 2.75 | 3.67 | 9.92 |
| Y81A | 2.97 | 5.31 | 1.58 | 2.91 | 9.94 |
| S109A | 2.60 | 6.93 | 1.80 | 29.5 | 9.90 |
| P112L | 2.04 | 8.14 | 1.66 | 3.70 | 9.57 |
| K113A | 8.72 | 3.38 | 2.95 | 3.80 | 8.15 |
| W173A | 7.73 | 3.12 | 2.41 | 5.53 | 9.76 |
| Y181A | 71.1 | 0.22 | 1.56 | 1.77 | 9.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, C.; Wu, Z.; Lin, W.; Xu, H.; Chang, T.; Dai, Y.; Lin, D. Suramin Targets the Conserved Ligand-Binding Pocket of Human Raf1 Kinase Inhibitory Protein. Molecules 2021, 26, 1151. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26041151
Guo C, Wu Z, Lin W, Xu H, Chang T, Dai Y, Lin D. Suramin Targets the Conserved Ligand-Binding Pocket of Human Raf1 Kinase Inhibitory Protein. Molecules. 2021; 26(4):1151. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26041151
Chicago/Turabian StyleGuo, Chenyun, Zhihua Wu, Weiliang Lin, Hao Xu, Ting Chang, Yazhuang Dai, and Donghai Lin. 2021. "Suramin Targets the Conserved Ligand-Binding Pocket of Human Raf1 Kinase Inhibitory Protein" Molecules 26, no. 4: 1151. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26041151