
Ameliorating the Adverse Effects of Tomato mosaic tobamovirus Infecting Tomato Plants in Egypt by Boosting Immunity in Tomato Plants Using Zinc Oxide Nanoparticles

 ,
,  , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Source of the Virus Isolate
2.2. Improving Negative Effects of ToMV Using ZnO-NPs
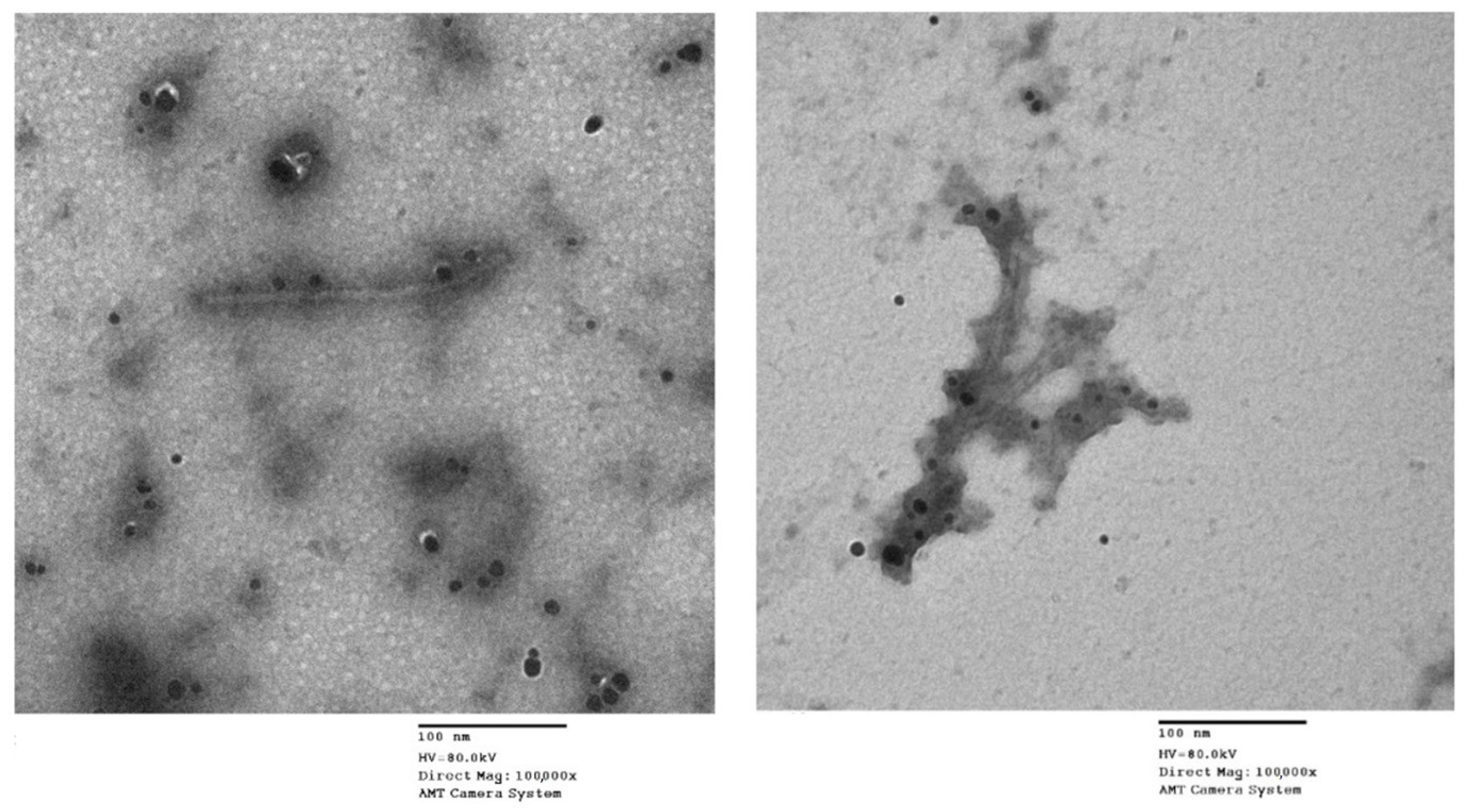
2.2.1. Direct Inactivation of ToMV by ZnO-NPs
2.2.2. Systemic Protection against ToMV in Tomato Plants Using ZnO-NPs
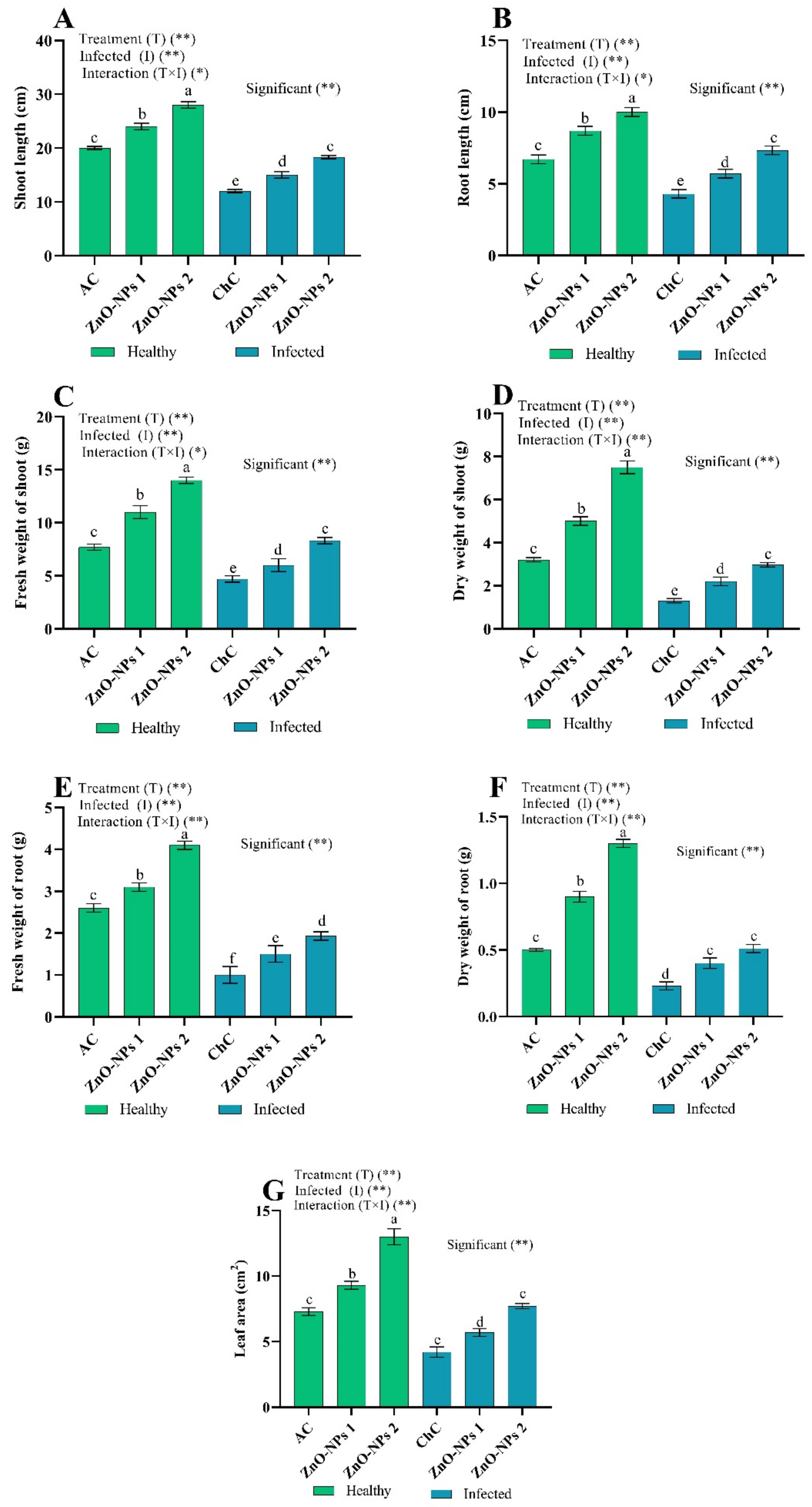
Growth Indices
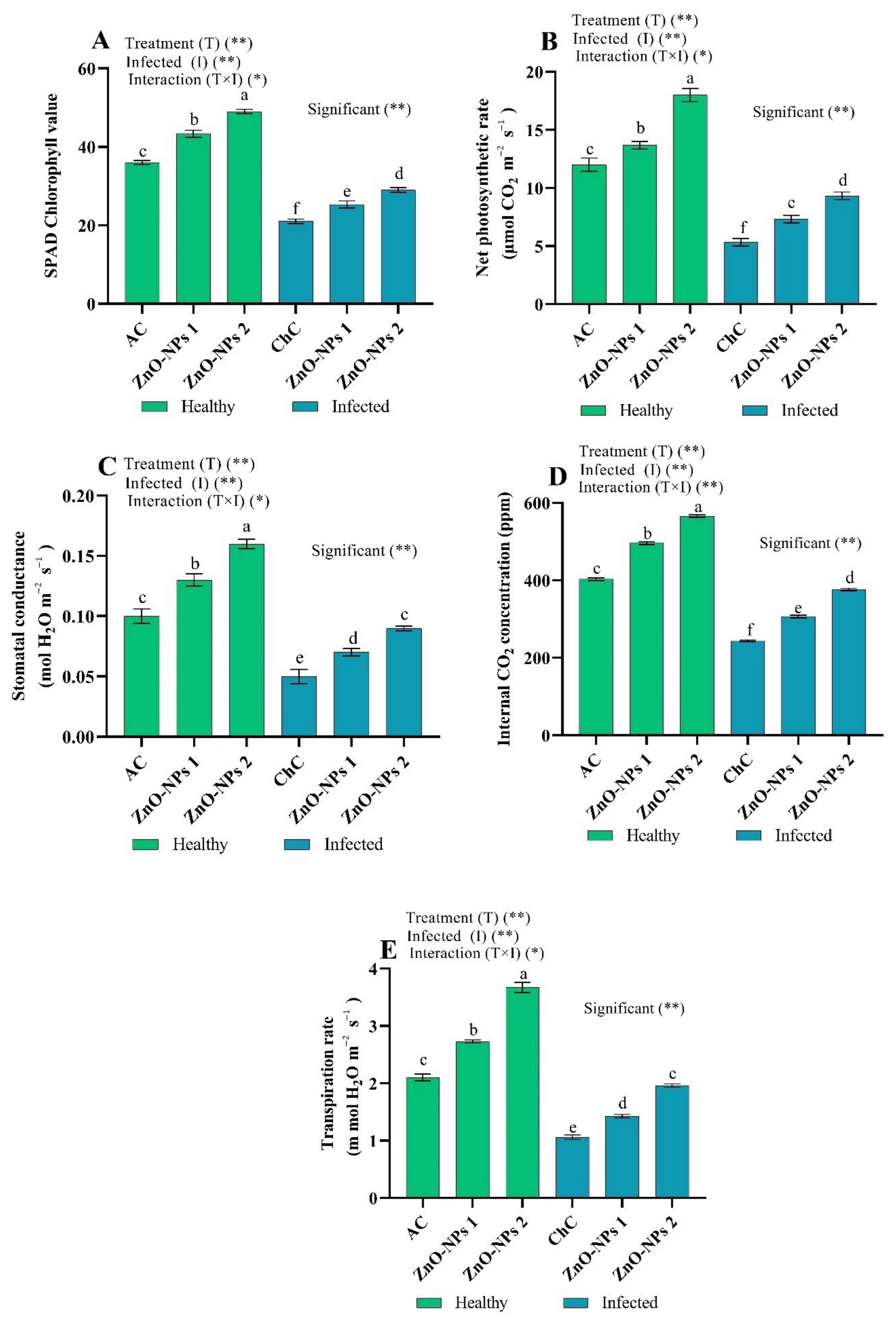
Chlorophyll Content and Photosynthetic Characteristics
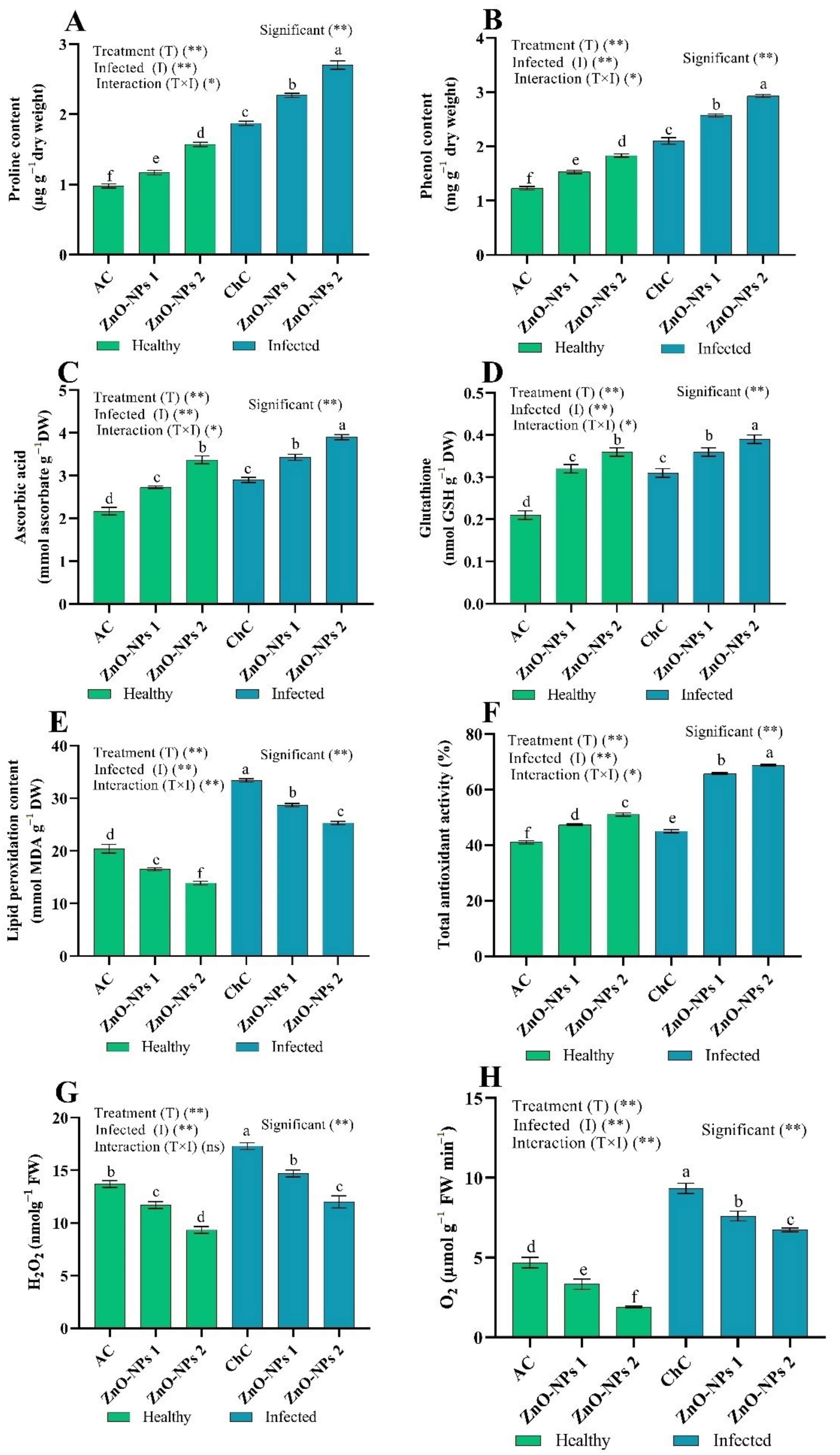
Oxidative Stress Markers
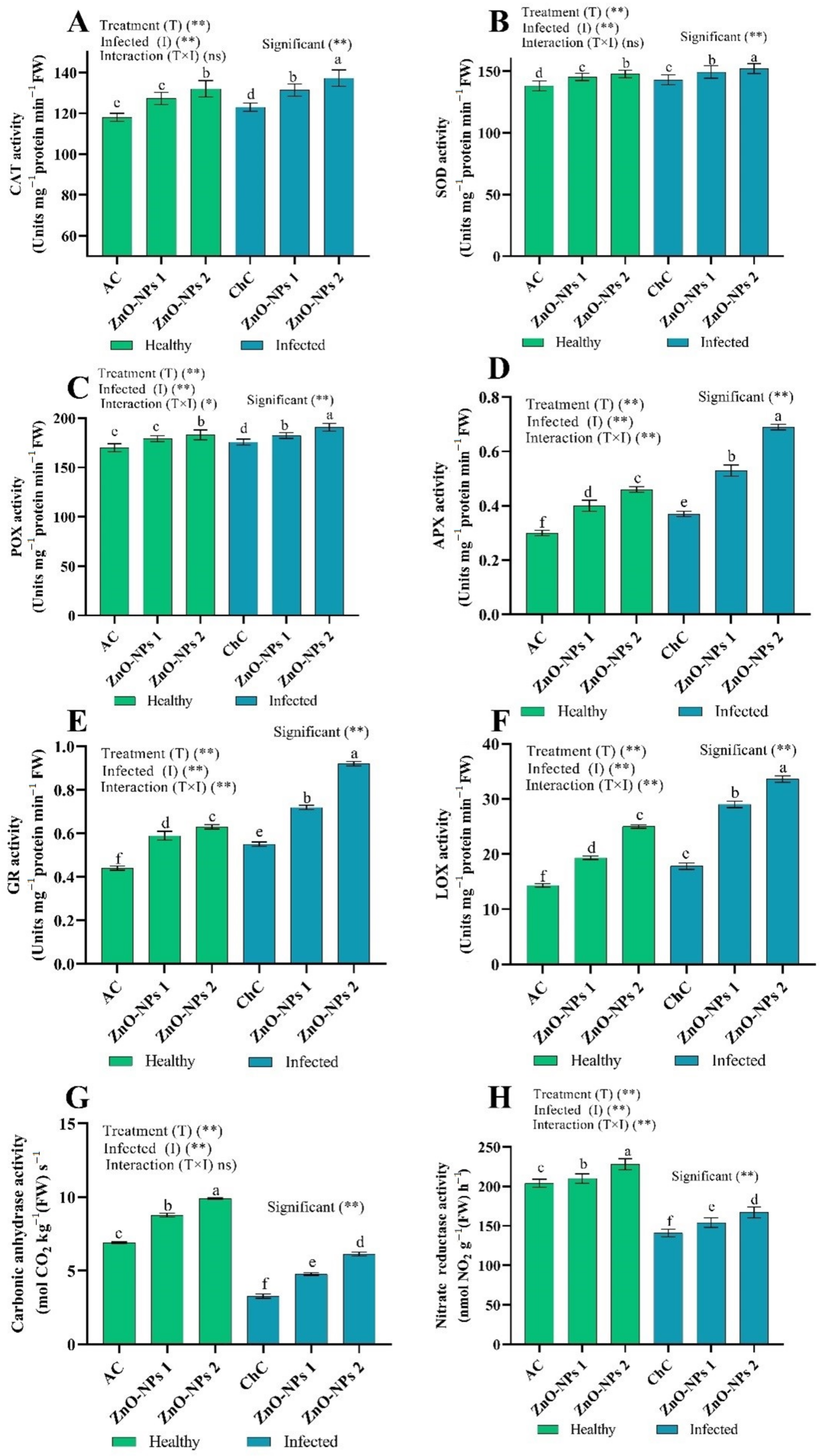
ROS Scavenging Enzymes
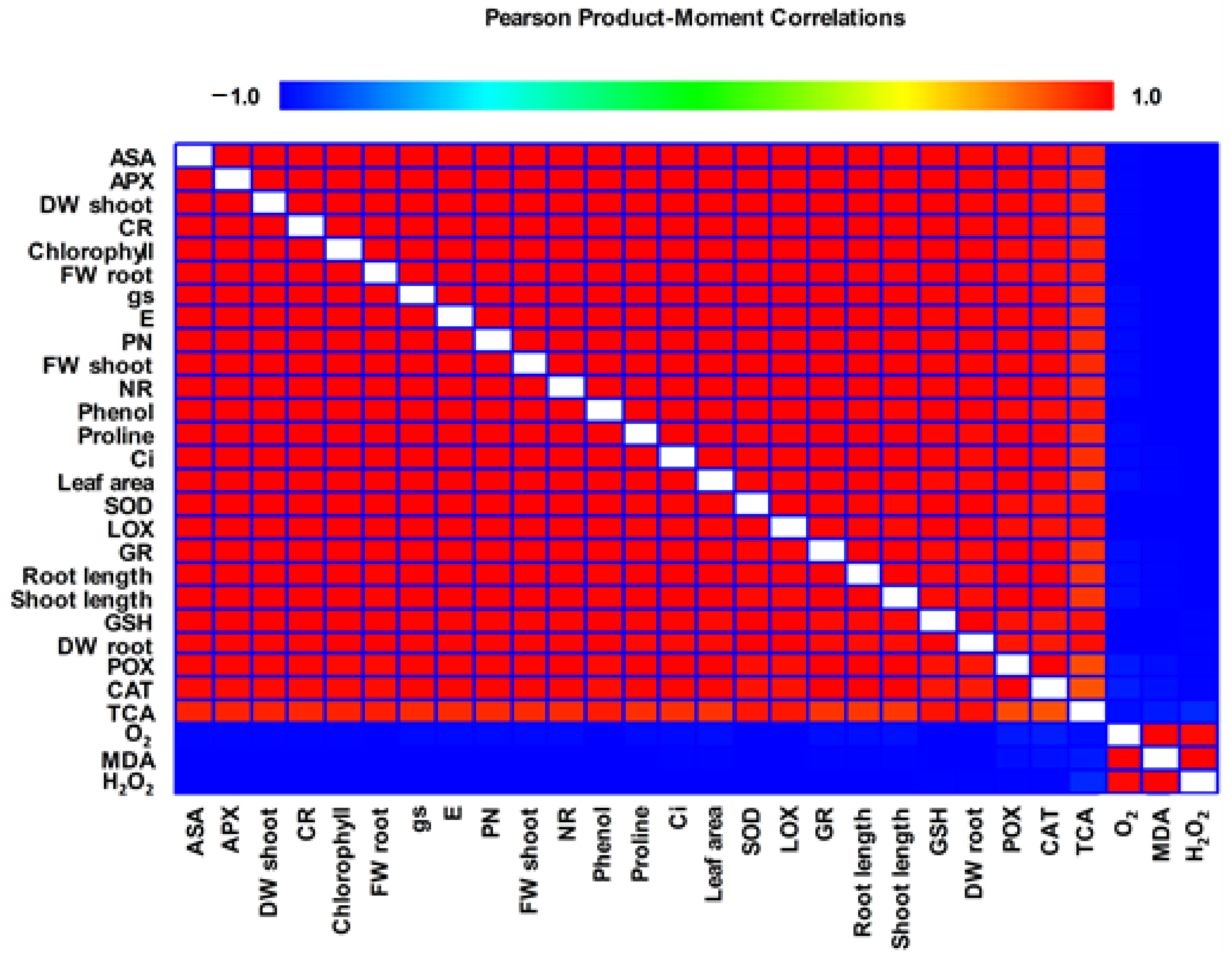
Correlation Analysis
3. Materials and Methods
3.1. Source of the Virus Isolate
3.2. Zinc Nanoparticles
3.3. Evaluation of Direct Antiviral Action
3.4. Systemic Protection against ToMV in Tomato Plants Using ZnO-NPs
3.4.1. Plant Materials
- In the first group, the plants were sprayed with water as untreated control (absolute control) (AC).
- In the second group, the plants were foliar sprayed with 50 mg/L ZnO-NPs (ZnO-NPs1).
- In the third group, the plants were foliar sprayed with 100 mg/L ZnO-NPs (ZnO-NPs2).
- In the fourth group, the plants were inoculated with Tomato mosaic virus (ToMV) as challenge control (ChC).
- In the fifth group, the plants were foliar sprayed with 50 mg/L ZnO-NPs and then inoculated after three days with ToMV (ZnO-NPs1 + V).
- In the sixth group, the plants were foliar sprayed with 100 mg/L ZnO-NPs and then inoculated after three days with ToMV (ZnO-NPs2 + V).
3.4.2. Growth Indices
3.4.3. Chlorophyll Content and Photosynthetic Characteristics
3.4.4. Oxidative Stress Markers
Proline and Phenols
Ascorbic Acid (ASA) and Glutathione (GSH) Content
Lipid Peroxidation and Total Antioxidant Activity
Quantification of H2O2 and O2
3.4.5. ROS Scavenging Enzymes
3.4.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Sofy, A.R.; Sofy, M.R.; Hmed, A.A.; El-Dougdoug, N.K. Potential Effect of Plant Growth-Promoting Rhizobacteria (PGPR) on Enhancing Protection against Viral Diseases. In Field Crops: Sustainable Management by PGPR. Sustainable Development and Bi-Odiversity; Maheshwari, D.K., Dheeman, S., Eds.; Springer: Cham, Switzerland, 2019; Volume 23, pp. 411–445. [Google Scholar]
- Megahed, A.A.; El-Dougdoug, K.A.; Othman, B.A.; Lashin, S.M.; Ibrahim, M.A.; Sofy, A.R. Induction of resistance in tomato plants against Tomato mosaic tobamovirus using beneficial microbial isolates. Pak. J. Biol. Sci. 2013, 16, 385–390. [Google Scholar] [CrossRef]
- Broadbent, L. The epidemiology of tomato mosaic. Ann. Appl. Biol. 1965, 56, 177–205. [Google Scholar] [CrossRef]
- Adams, M.J.; Heinze, C.; Jackson, A.O.; Kreuze, J.F.; Macfarlane, S.A.; Torrance, L. Tobamovirus. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier/Academic Press: London, UK, 2012; pp. 1153–1156. [Google Scholar]
- Cai, L.; Liu, C.; Fan, G.; Liu, C.; Sun, X. Preventing viral disease by ZnONPs through directly deactivating TMV and activating plant immunity in Nicotiana Benthamiana. Environ. Sci. Nano 2019, 6, 3653–3669. [Google Scholar] [CrossRef]
- Sofy, A.R.; Hmed, A.A.; Alnaggar, A.E.-A.M.; Dawoud, R.A.; Elshaarawy, R.F.M.; Sofy, M.R. Mitigating effects of Bean yellow mosaic virus infection in faba bean using new carboxymethyl chitosan-titania nanobiocomposites. Int. J. Biol. Macromol. 2020, 163, 1261–1275. [Google Scholar] [CrossRef] [PubMed]
- Sirelkhatim, A.; Mahmud, S.; Seeni, A.; Kaus, N.H.M.; Ann, L.C.; Bakhori, S.K.M.; Hasan, H.; Mohamad, D. Review on zinc oxide nanoparticles: Antibacterial activity and toxicity mechanism. Nano-Micro Lett. 2015, 7, 219–242. [Google Scholar] [CrossRef] [Green Version]
- Hoseinzadeh, E.; Makhdoumi, P.; Taha, P.; Hossini, H.; Stelling, J.; Amjad Kamal, M.; Ashraf, G.M. A review on nano-antimicrobials: Metal nanoparticles, methods and mechanisms. Curr. Drug Metab. 2017, 18, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.O.; McLean, J.E.; Britt, D.W.; Anderson, A.J. Antifungal activity of ZnO nanoparticles and their interactive effect with a biocontrol bacterium on growth antagonism of the plant pathogen Fusarium graminearum. Biometals 2013, 26, 913–924. [Google Scholar] [CrossRef]
- Sofy, A.R.; Hmed, A.A.; Abd El Haliem, N.F.; Zein, M.A.E.; Elshaarawy, R.F.M. Polyphosphonium-oligochitosans decorated with nanosilver as new prospective inhibitors for common human enteric viruses. Carbohydr. Polym. 2019, 226, 115261. [Google Scholar] [CrossRef]
- Elshaarawy, R.F.M.; Mustafa, F.H.A.; Sofy, A.R.; Hmed, A.A.; Janiak, C. A new synthetic antifouling coatings integrated novel aminothiazole-functionalized ionic liquids motifs with enhanced antibacterial performance. J. Environ. Chem. Eng. 2019, 7, 102800. [Google Scholar] [CrossRef]
- Abdelmoamen, A.; El-Dougdoug, N.; El-Borollosy, A.; El-Dougdoug, K. Serological and molecular characteristics of Tomato mosaic tobamovirus coat Protein. Arab Univ. J. Agric. Sci. 2018, 26, 1613–1626. [Google Scholar] [CrossRef] [Green Version]
- El-Ahdal, A.M.E.; Mayer, F.; El Afifi, S.I.; Allam, E.K. Characterization of two strains of Tobacco mosaic Virus by biological, biochemical, immunological and electron microscopical techniques. Z. Für Allg. Mikrobiol. 1984, 24, 507–514. [Google Scholar] [CrossRef]
- Singh, A.; Singh, N.B.; Afzal, S.; Singh, T.; Hussain, I. Zinc oxide nanoparticles: A review of their biological synthesis, antimicrobial activity, uptake, translocation and biotransformation in plants. J. Mater. Sci. 2018, 53, 185–201. [Google Scholar] [CrossRef]
- Jayaseelan, C.; Rahuman, A.A.; Kirthi, A.V.; Marimuthu, S.; Santhoshkumar, T.; Bagavan, A.; Gaurav, K.; Karthik, L.; Rao, K.V.B. Novel microbial route to synthesize ZnO nanoparticles using Aeromonas hydrophila and their activity against pathogenic bacteria and fungi. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2012, 90, 78–84. [Google Scholar] [CrossRef]
- Agarwal, H.; Venkat Kumar, S.; Rajeshkumar, S. A review on green synthesis of zinc oxide nanoparticles—An eco-friendly approach. Resour. Effic. Technol. 2017, 3, 406–413. [Google Scholar] [CrossRef]
- Raliya, R.; Nair, R.; Chavalmane, S.; Wang, W.; Biswas, P. Mechanistic evaluation of translocation and physiological impact of titanium dioxide and zinc oxide nanoparticles on the tomato (Solanum lycopersicum L.) plant. Metallomics 2015, 7, 1584–1594. [Google Scholar] [CrossRef]
- Khodakovskaya, M.; de Silva, K.; Biris, A.; Dervishi, E.; Villagarcia, H. Carbon Nanotubes Induce Growth Enhancement of Tobacco Cells. ACS Nano 2012, 6, 2128–2135. [Google Scholar] [CrossRef] [PubMed]
- Sofy, A.R.; Sofy, M.R.; Hmed, A.A.; Dawoud, R.A.; Refaey, E.E.; Mohamed, H.I.; El-Dougdoug, N.K. Molecular characterization of the Alfalfa mosaic virus infecting Solanum melongena in Egypt and the control of its deleterious effects with melatonin and salicylic acid. Plants 2021, 10, 459. [Google Scholar] [CrossRef]
- El-Dougdou, K.A.; Sofy, A.R.; Hameed, G.A.; Dawood, R.A. Intraspecific diversity of Cucumber mosaic cucumoviridae in Egypt. Int. J. Virol. 2014, 10, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Sofy, A.R.; Soliman, A.M.; Mousa, A.A.; El-Dougdou, K.A. Molecular characterization and bioinformatics analysis of viroid isolate associated with citrus gummy bark disease in Egypt. Int. J. Virol. 2012, 8, 133–150. [Google Scholar] [CrossRef]
- Sofy, A.R.; Mousa, A.A.; Soliman, A.M.; Dougdoug, K.A.E. The limiting of climatic factors and predicting of suitable habitat for citrus gummy bark disease occurrence using GIS. Int. J. Virol. 2012, 8, 165–177. [Google Scholar] [CrossRef] [Green Version]
- El-Dougdou, K.A.; Ghazal, S.A.; Mousa, A.A.; Fahmy, H.; Sofy, A.R. Differentiation among three Egyptian isolates of Citrus psorosis virus. Int. J. Virol. 2009, 5, 49–63. [Google Scholar] [CrossRef]
- Ghazal, S.A.; El-Dougdoug Kh, A.; Mousa, A.A.; Fahmy, H.; Sofy, A.R. Isolation and identification of Citrus psorosis virus Egyptian isolate (CPsV-EG). Commun. Agric. Appl. Biol. Sci. 2008, 73, 285–295. [Google Scholar] [PubMed]
- Faizan, M.; Faraz, A.; Hayat, S. Effective use of zinc oxide nanoparticles through root dipping on the performance of growth, quality, photosynthesis and antioxidant system in tomato. J. Plant. Biochem. Biotechnol. 2019, 1–15. [Google Scholar] [CrossRef]
- García-Gómez, C.; García, S.; Obrador, A.F.; González, D.; Babín, M.; Fernández, M.D. Effects of aged ZnO NPs and soil type on Zn availability, accumulation and toxicity to pea and beet in a greenhouse experiment. Ecotoxicol. Environ. Saf. 2018, 160, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Abdel Latef, A.A.H.; Abu Alhmad, M.F.; Abdelfattah, K.E. The possible roles of priming with ZnO nanoparticles in mitigation of salinity stress in lupine (Lupinus termis) plants. J. Plant. Growth Regul. 2017, 36, 60–70. [Google Scholar] [CrossRef]
- Cakmak, I. Tansley Review No. 111. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef]
- Sofy, M.R.; Elhindi, K.M.; Farouk, S.; Alotaibi, M.A. Zinc and paclobutrazol mediated regulation of growth, upregulating antioxidant aptitude and plant productivity of pea plants under salinity. Plants 2020, 9, 1197. [Google Scholar] [CrossRef]
- Akladious, S.A.; Mohamed, H.I. Physiological role of exogenous nitric oxide in improving performance, yield and some biochemical aspects of sunflower plant under zinc stress. Acta Biol. Hung. 2017, 68, 101–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faizan, M.; Faraz, A.; Yusuf, M.; Khan, S.T.; Hayat, S. Zinc oxide nanoparticle-mediated changes in photosynthetic efficiency and antioxidant system of tomato plants. Photosynthetica 2018, 56, 678–686. [Google Scholar] [CrossRef]
- Vitti, A.; Pellegrini, E.; Nali, C.; Lovelli, S.; Sofo, A.; Valerio, M.; Scopa, A.; Nuzzaci, M. Trichoderma harzianum T-22 induces systemic resistance in tomato infected by Cucumber mosaic virus. Front. Plant Sci. 2016, 7, 1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofy, A.R.; Dawoud, R.A.; Sofy, M.R.; Mohamed, H.I.; Hmed, A.A.; El-Dougdoug, N.K. Improving Regulation of Enzymatic and Non-Enzymatic Antioxidants and Stress-Related Gene Stimulation in Cucumber mosaic cucumovirus-Infected Cucumber Plants Treated with Glycine Betaine, Chitosan and Combination. Molecules 2020, 25, 2341. [Google Scholar] [CrossRef]
- Megahed, A.A.; El-Dougdoug, K.A.; Othman, B.A.; Lashin, S.M.; Ibrahim, M.A.; Sofy, A.R. A new Egyptian satellite strain of Cucumber mosaic cucumovirus. Int. J. Virol. 2012, 8, 240–257. [Google Scholar] [CrossRef] [Green Version]
- Rahoutei, J.; García-Luque, I.; Barón, M. Inhibition of photosynthesis by viral infection: Effect on PSII structure and function. Physiol. Plant. 2000, 110, 286–292. [Google Scholar] [CrossRef]
- Balachandran, S.; Hurry, V.M.; Kelley, S.E.; Osmond, C.B.; Robinson, S.A.; Rohozinski, J.; Seaton, G.G.R.; Sims, D.A. Concepts of plant biotic stress. Some insights into the stress physiology of virus-infected plants, from the perspective of photosynthesis. Physiol. Plant. 1997, 100, 203–213. [Google Scholar] [CrossRef]
- Fan, H.; Song, B.; Bhadury, P.; Jin, L.; Hu, D.; Yang, S. Antiviral Activity and Mechanism of Action of Novel Thiourea Containing Chiral Phosphonate on Tobacco Mosaic Virus. Int. J. Mol. Sci. 2011, 12, 4522–4535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govorov, A.O.; Carmeli, I. Hybrid Structures Composed of Photosynthetic System and Metal Nanoparticles: Plasmon Enhancement Effect. Nano Lett. 2007, 7, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Noji, T.; Kamidaki, C.; Kawakami, K.; Shen, J.; Kajino, T.; Fukushima, Y.; Sekitoh, T.; Itoh, S. Photosynthetic Oxygen Evolution in Mesoporous Silica Material: Adsorption of Photosystem II Reaction Center Complex into 23 nm Nanopores in SBA. Langmuir 2011, 27, 705–713. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Faisal, M.; Al Sahli, A.A. Nano-silicon dioxide mitigates the adverse effects of salt stress on Cucurbita pepo L. Environ. Toxicol. Chem. 2014, 33, 2429–2437. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, B.; Tao, G.; Zhang, Q.; Zhang, C. Effects of nano-silicon dioxide on photosynthetic fluorescence characteristics of Indocalamus barbatus McClure. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2012, 36, 59–63. [Google Scholar]
- Aly, A.A.; Mohamed, H.I.; Mansour, M.T.M.; Omar, M.R. Suppression of Powdery Mildew on Flax by Foliar Application of Essential Oils. J. Phytopathol. 2013, 161, 376–381. [Google Scholar] [CrossRef]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.; Mittler, R.; Rivero, R. Tolerance to Stress Combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant. Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- El-Beltagi, H.S.; Sofy, M.R.; Aldaej, M.I.; Mohamed, H.I. Silicon Alleviates Copper Toxicity in Flax Plants by Up-Regulating Antioxidant Defense and Secondary Metabolites and Decreasing Oxidative Damage. Sustainability 2020, 12, 4732. [Google Scholar] [CrossRef]
- Abu-Shahba, M.S.; Mansour, M.M.; Mohamed, H.I.; Sofy, M.R. Comparative Cultivation and Biochemical Analysis of Iceberg Lettuce Grown in Sand Soil and Hydroponics with or Without Microbubbles and Macrobubbles. J. Soil Sci. Plant Nutr. 2020, 1–15. [Google Scholar] [CrossRef]
- Sofy, A.R.; Mahfouze, S.A.; El-Enany, M.A. Isozyme markers for response of wild potato species to potato spindle tuber viroid Egyptian isolate. World Appl. Sci. J. 2013, 27, 1010–1022. [Google Scholar] [CrossRef]
- Abd El-Rahman, S.S.; Mazen, M.M.; Mohamed, H.I.; Mahmoud, N.M. Induction of defence related enzymes and phenolic compounds in lupin (Lupinus albus L.) and their effects on host resistance against Fusarium wilt. Eur. J. Plant Pathol. 2012, 134, 105–116. [Google Scholar] [CrossRef]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Helaly, M.; El-Metwally, M.; El-Hoseiny, H.; Omar, S.; El-Sheery, N. Effect of nanoparticles on biological contamination of’in vitro’cultures and organogenic regeneration of banana. Aust. J. Crop. Sci. 2014, 8, 612. [Google Scholar]
- Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing Adverse Effects of Pb on Maize Plants by Combined Treatment with Jasmonic, Salicylic Acids and Proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- Sofy, M.R.; Elhawat, N.; Tarek, A. Glycine betaine counters salinity stress by maintaining high K+/Na+ ratio and antioxidant defense via limiting Na+ uptake in common bean (Phaseolus vulgaris L.). Ecotoxicol. Environ. Saf. 2020, 200, 110732. [Google Scholar] [CrossRef]
- Kaushik, D.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar]
- Pérez-Labrada, F.; López-Vargas, E.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Responses of Tomato Plants under Saline Stress to Foliar Application of Copper Nanoparticles. Plants 2019, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Turkan, I. ROS and RNS: Key signalling molecules in plants. J. Exp. Bot. 2018, 69, 3313–3315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.; Rahman, A.; Suzuki, T.; Fujita, M. Polyamine and nitric oxide crosstalk: Antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense and methylglyoxal detoxification systems. Ecotoxicol. Environ. Saf. 2016, 126, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.H.M.; Mahmud, J.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Bhuyan, M.; Oku, H.; Fujita, M. Exogenous nitric oxide pretreatment protects Brassica napus L. seedlings from paraquat toxicity through the modulation of antioxidant defense and glyoxalase systems. Plant Physiol. Biochem. 2018, 126, 173–186. [Google Scholar] [CrossRef]
- Soares, C.; Carvalho, M.; Azevedo, R.; Fidalgo, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Lv, D.; Zhu, G.; Zhu, D.; Bian, Y.; Liang, X.; Cheng, Z.; Deng, X.; Yan, Y. Proteomic and phosphoproteomic analysis reveals the response and defense mechanism in leaves of diploid wheat T. monococcum under salt stress and recovery. J. Proteom. 2016, 143, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Mathioudakis, M.; Veiga, R.; Canto, T.; Medina, V.; Mossialos, D.; Makris, A.; Livieratos, I. Pepino mosaic virus triple gene block protein 1 (TGBp1) interacts with and increases tomato catalase 1 activity to enhance virus accumulation. Mol. Plant Pathol. 2013, 14, 589–601. [Google Scholar] [CrossRef]
- Derbalah, A.S.; Elsharkawy, M.M. A new strategy to control Cucumber mosaic virus using fabricated NiO-nanostructures. J. Biotechnol. 2019, 306, 134–141. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- Gooding, G.V., Jr.; Hebert, T.T. A simple technique for purification of Tobacco mosaic virus in large quantities. Phytopathology 1967, 57, 1285. [Google Scholar]
- Christie, S.R.; Purcifull, D.E.; Crawford, W.E.; Ahmed, N.A. Electron microscopy of negatively stained clarified viral concentrates obtained from small tissue samples with appendices on negative staining techniques. Bull. Agric. Exp. Stn. Univ. Fla. 1987, 872, 45. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Galicia, L.; Rosales, E.; Palacios, R. Laboratory Protocols 2008: Maize Nutrition Quality and Plant Tissue Analysis Laboratory; CIMMYT: El Batán, Mexico, 2009. [Google Scholar]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Griffith, O. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Patterson, B.D.; MacRae, E.A.; Ferguson, I.B. Estimation of hydrogen peroxide in plant extracts using titanium (IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef]
- Jabs, T.; Dietrich, R.A.; Dang, J.L. Initiation of runaway cell death in an Arabidopsis mutant by extracellular superoxide. Science 1996, 273, 1853–1856. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, X.; Lu, Y.; Wang, X. Effects of rare earth metal ions and their EDTA complexes on antioxidant enzymes of fish liver. Bull. Environ. Contam. Toxicol. 2000, 65, 357–365. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.L.; Jen, J.J.; Morr, C.V. Changes in Soluble and Bound Peroxidase—IAA Oxidase During Tomato Fruit Development. J. Food Sci. 1982, 47, 158–161. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [Green Version]
- Todd, J.; Paliyath, G.; Thompson, J. Characteristics of a Membrane-Associated Lipoxygenase in Tomato Fruit. Plant Physiol. 1990, 94, 1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworski, E. Nitrate reductase assay in intact plant tissues. Biochem. Biophys. Res. Commun. 1971, 43, 1274–1279. [Google Scholar] [CrossRef]
- Dwivedi, R.; Randhawa, N.S. Evaluation of a rapid test for the hidden hunger of zinc in plants. Plant Soil 1974, 40, 445–451. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons Inc.: Singapore, 1984. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sofy, A.R.; Sofy, M.R.; Hmed, A.A.; Dawoud, R.A.; Alnaggar, A.E.-A.M.; Soliman, A.M.; El-Dougdoug, N.K. Ameliorating the Adverse Effects of Tomato mosaic tobamovirus Infecting Tomato Plants in Egypt by Boosting Immunity in Tomato Plants Using Zinc Oxide Nanoparticles. Molecules 2021, 26, 1337. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051337
Sofy AR, Sofy MR, Hmed AA, Dawoud RA, Alnaggar AE-AM, Soliman AM, El-Dougdoug NK. Ameliorating the Adverse Effects of Tomato mosaic tobamovirus Infecting Tomato Plants in Egypt by Boosting Immunity in Tomato Plants Using Zinc Oxide Nanoparticles. Molecules. 2021; 26(5):1337. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051337
Chicago/Turabian StyleSofy, Ahmed R., Mahmoud R. Sofy, Ahmed A. Hmed, Rehab A. Dawoud, Abd El-Aleem M. Alnaggar, Ahmed M. Soliman, and Noha K. El-Dougdoug. 2021. "Ameliorating the Adverse Effects of Tomato mosaic tobamovirus Infecting Tomato Plants in Egypt by Boosting Immunity in Tomato Plants Using Zinc Oxide Nanoparticles" Molecules 26, no. 5: 1337. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051337