
A Synergic Potential of Antimicrobial Peptides against Pseudomonas syringae pv. actinidiae

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
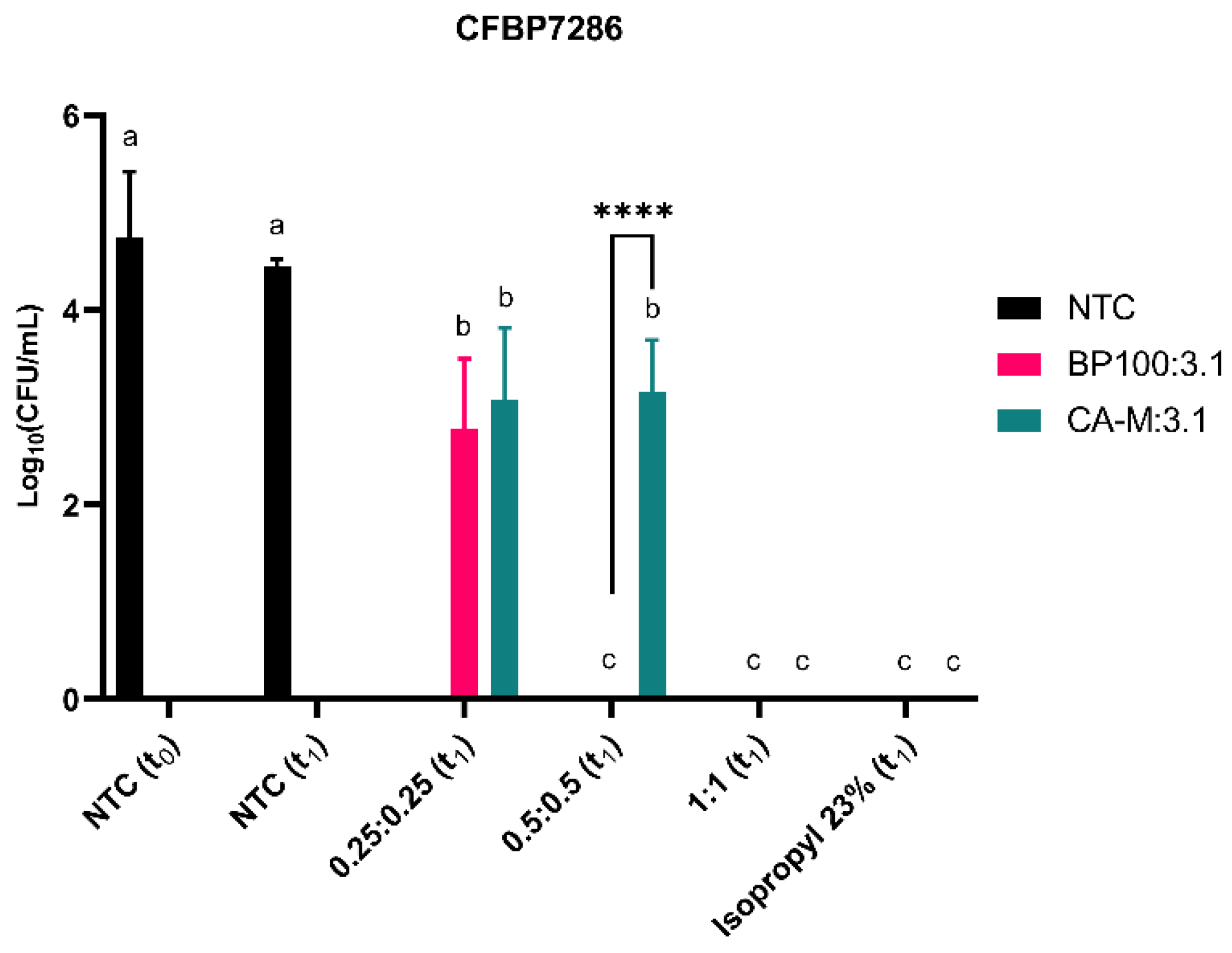
2. Results
2.1. Peptide Synthesis
2.2. Antibiogram Assays
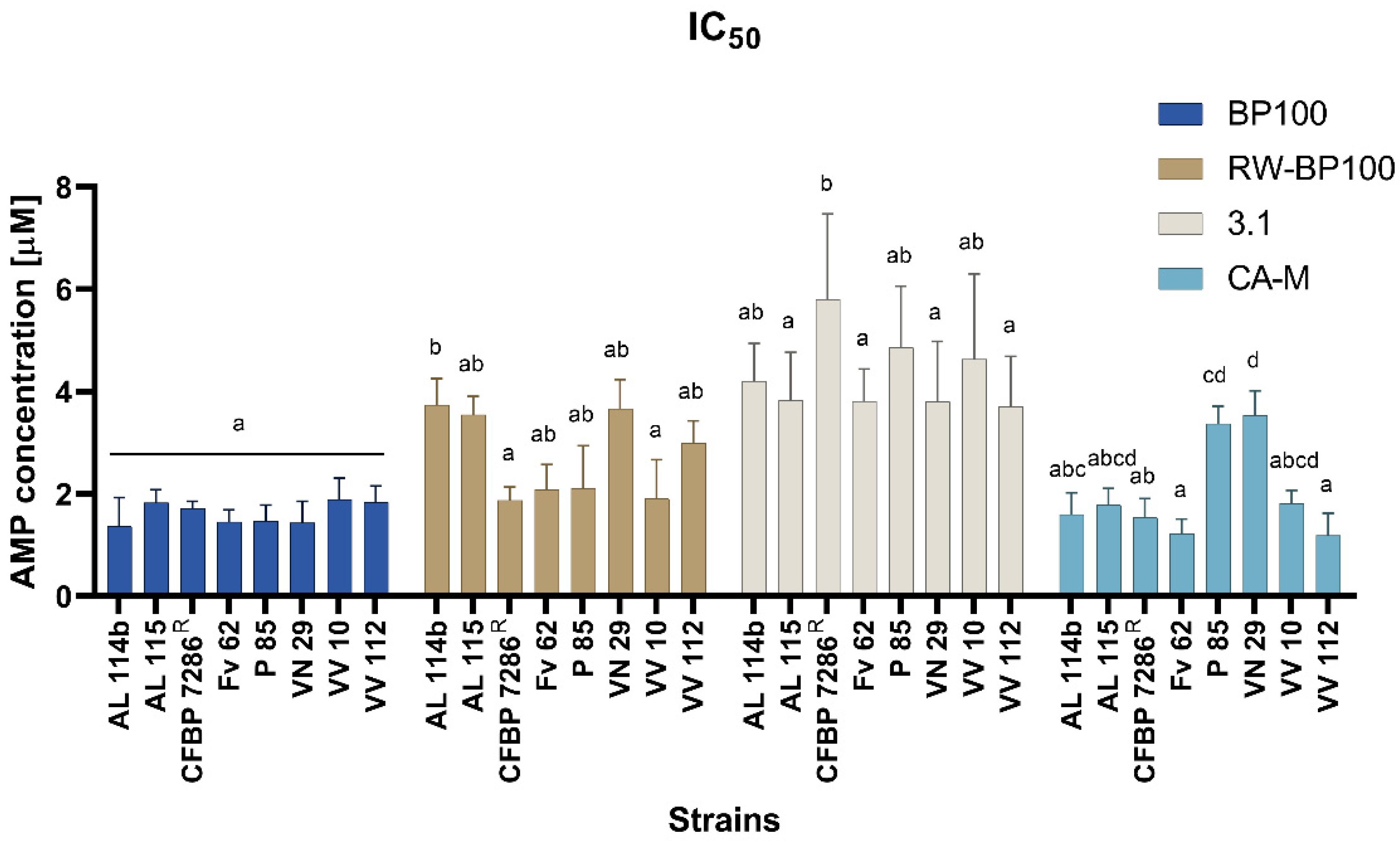
2.3. Minimal Inhibitory and Bactericidal Concentration and IC50
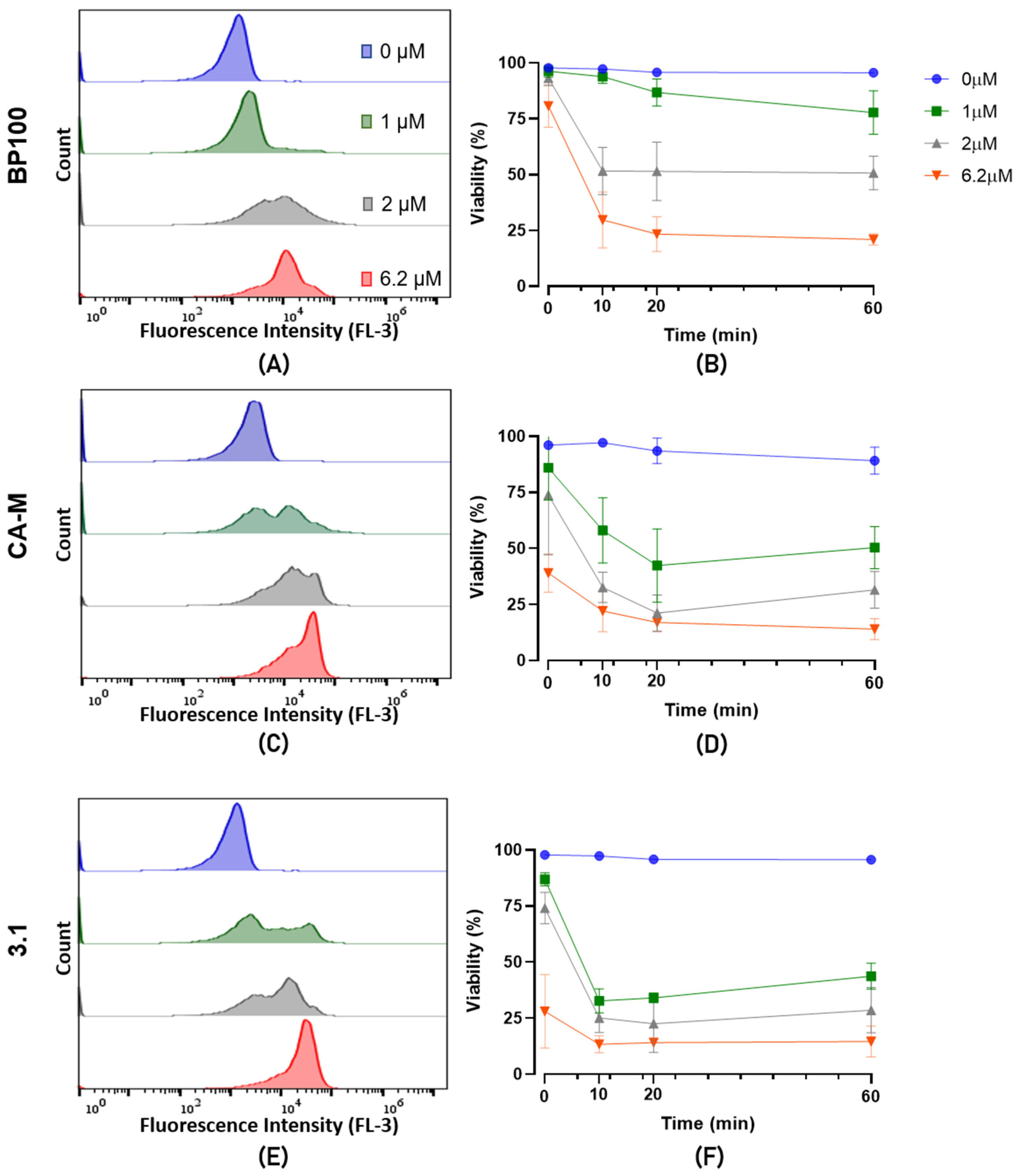
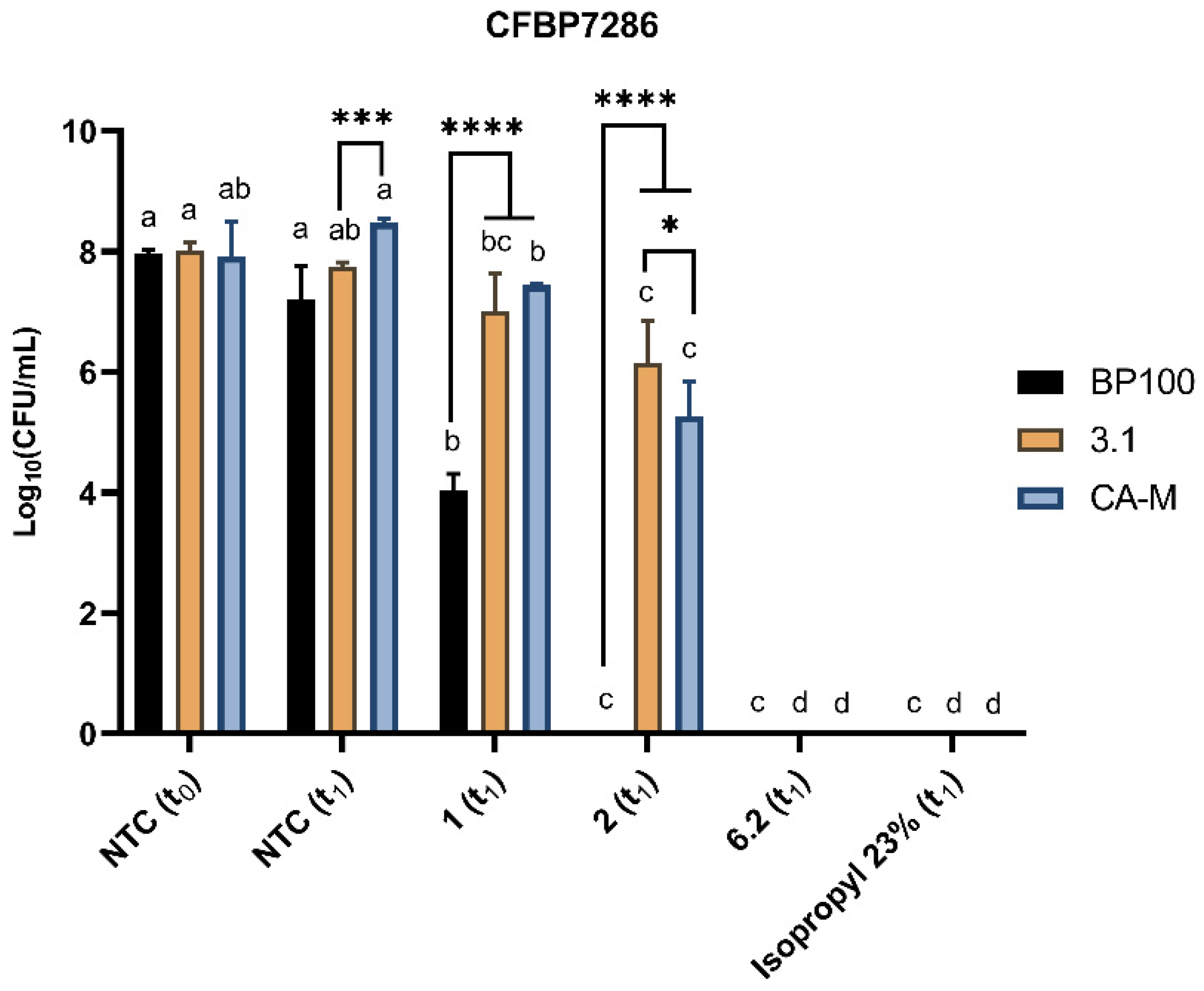
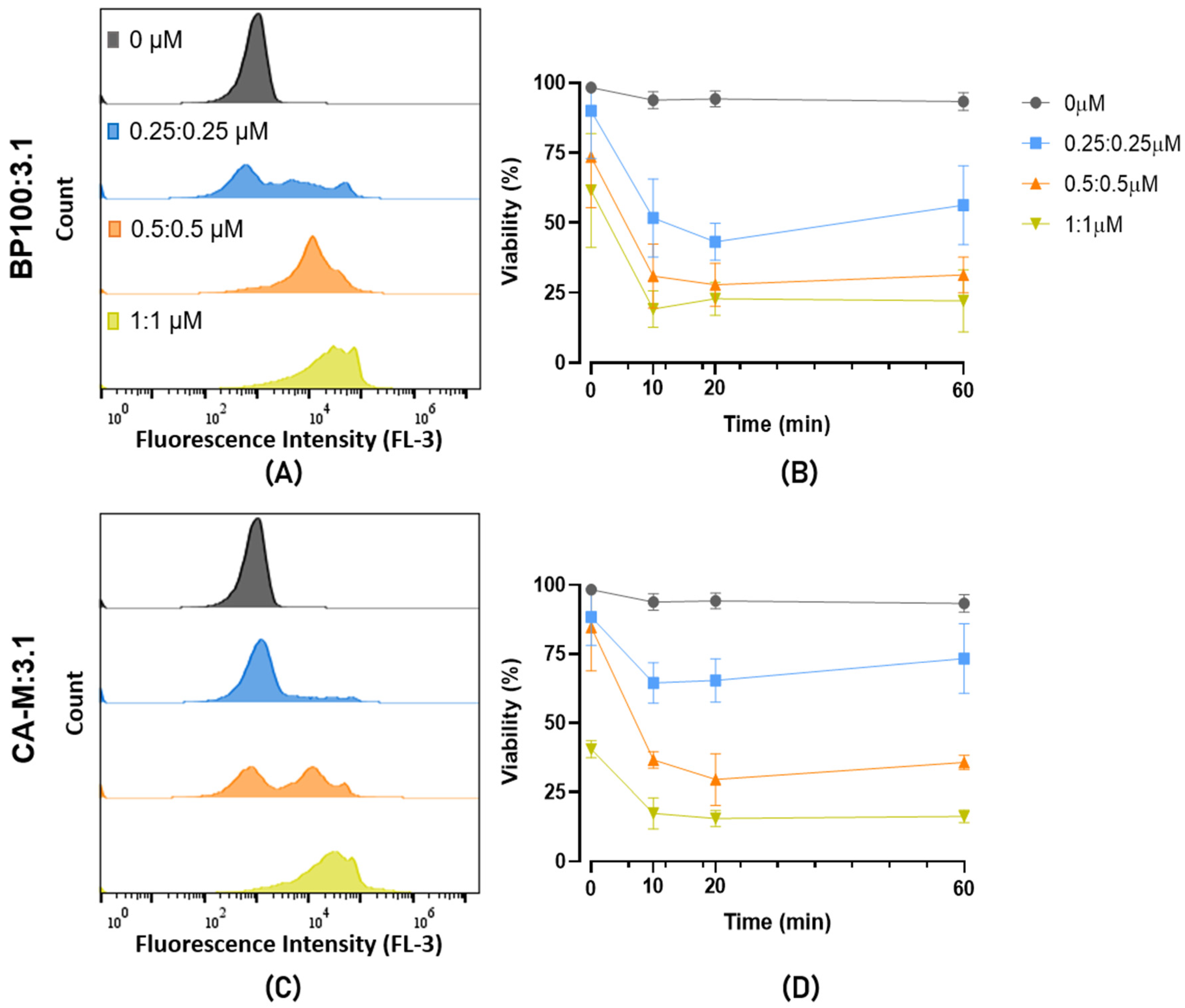
2.4. Evaluation of Bacterial Viability
2.5. Evaluation of AMPs Treatments on Tobacco Hypersensitivity Response
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Peptide Synthesis
4.3. Antibacterial Activity
4.3.1. Antibiogram Assay
4.3.2. Determination of Minimal Inhibitory Concentration and Minimal Bactericidal Concentration of AMPs
4.3.3. Flow Cytometry in the Evaluation of AMP Function and Viability
4.3.4. Assessment of Colony Forming Units (CFUs)
4.4. Hypersensitivity Response (HR) in Tobacco Leaves
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Donati, I.; Cellini, A.; Sangiorgio, D.; Vanneste, J.L.; Scortichini, M.; Balestra, G.M.; Spinelli, F. Pseudomonas syringae pv. actinidiae: Ecology, infection dynamics and disease epidemiology. Microb. Ecol. 2020, 80, 81–102. [Google Scholar] [CrossRef]
- Vanneste, J.L. The scientific, economic, and social impacts of the New Zealand outbreak of bacterial canker of kiwifruit (Pseudomonas syringae pv. actinidiae). Ann. Rev. Phytopathol. 2017, 55, 377–399. [Google Scholar] [CrossRef]
- Vanneste, J.L. Pseudomonas syringae pv. actinidiae (Psa): A threat to the New Zealand and global kiwifruit industry. N. Z. J. Crop Hortic. Sci. 2012, 40, 265–267. [Google Scholar] [CrossRef] [Green Version]
- McCann, H.C.; Rikkerink, E.H.; Bertels, F.; Fiers, M.; Lu, A.; Rees-George, J.; Andersen, M.T.; Gleave, A.P.; Haubold, B.; Wohlers, M.W.; et al. Genomic analysis of the kiwifruit pathogen Pseudomonas syringae pv. actinidiae provides insight into the origins of an emergent plant disease. PLoS Pathog. 2013, 9, e1003503. [Google Scholar] [CrossRef]
- Chapman, J.R.; Taylor, R.K.; Weir, B.S.; Romberg, M.K.; Vanneste, J.L.; Luck, J.; Alexander, B.J.R. Phylogenetic relationships among global populations of Pseudomonas syringae pv. actinidiae. Phytopathology 2012, 102, 1034–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujikawa, T.; Sawada, H. Genome analysis of the kiwifruit canker pathogen Pseudomonas syringae pv. actinidiae biovar 5. Sci. Rep. 2016, 6, 21399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, H.; Fujikawa, T. Genetic diversity of Pseudomonas syringae pv. actinidiae, pathogen of kiwifruit bacterial canker. Plant. Pathol. 2019, 68, 1235–1248. [Google Scholar] [CrossRef]
- Fujikawa, T.; Sawada, H. Genome analysis of Pseudomonas syringae pv. actinidiae biovar 6, which produces the phytotoxins, phaseolotoxin and coronatine. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.; Moura, L.; Abelleira, A.; Aguín, O.; Ares, A.; Mansilla, P. Characterization of Pseudomonas syringae pv. actinidiae biovar 3 on kiwifruit in north-west Portugal. J. Appl. Microbiol. 2018, 125, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Firrao, G.; Torelli, E.; Polano, C.; Ferrante, P.; Ferrini, F.; Martini, M.; Marcelletti, S.; Scortichini, M.; Ermacora, P. Genomic structural variations affecting virulence during clonal expansion of Pseudomonas syringae pv. actinidiae biovar 3 in Europe. Front. Microbiol. 2018, 9, 656. [Google Scholar] [CrossRef]
- Scortichini, M.; Marcelletti, S.; Ferrante, P.; Petriccione, M.; Firrao, G. Pseudomonas syringae pv. actinidiae: A re-emerging, multi-faceted, pandemic pathogen. Mol. Plant. Pathol. 2012, 13, 631–640. [Google Scholar] [CrossRef]
- Moura, L.; Garcia, E.; Aguín, O.; Ares, A.; Abelleira, A.; Mansilla, P. Identificação e caracterização de Pseudomonas syringae pv. actinidiae (Psa) na Região do Entre Douro e Minho (Portugal). Rev. Ciências Agrárias 2015, 38, 196–205. [Google Scholar]
- EPPO Global Database. Available online: https://gd.eppo.int/rppo/EU/categorization (accessed on 10 January 2021).
- Woodcock, S. A review of research and development undertaken on Psa. Kiwi Vine Health 2016, 5, 871–874. [Google Scholar]
- Cameron, A.; Sarojini, V. Pseudomonas syringae pv. actinidiae: Chemical control, resistance mechanisms and possible alternatives. Plant. Pathol. 2014, 63, 1–11. [Google Scholar] [CrossRef]
- Monchiero, M.; Gullino, M.L.; Pugliese, M.; Spadaro, D.; Garibaldi, A. Efficacy of different chemical and biological products in the control of Pseudomonas syringae pv. actinidiae on kiwifruit. Australas. Plant. Pathol. 2015, 44, 13–23. [Google Scholar] [CrossRef]
- Ministry for Primary Industries. A Strategy for New Zealand Food Safety 2019–2024; New Zealand Food Safety: Wellington, New Zealand, 2019.
- Taylor, P.; Reeder, R. Antibiotic use on crops in low and middle-income countries based on recommendations made by agricultural advisors. CABI Agricult. Biosci. 2020, 1, 1–14. [Google Scholar] [CrossRef]
- O’Niell, J. Antimicrobials in Agriculture and the Environment: Reducing Unnecessary Use and Waste: Review on Antimicrobial Resistance; European Union: Brussels, Belhium, 2015. [Google Scholar]
- Han, H.S.; Nam, H.Y.; Koh, Y.J.; Hur, J.S.; Jung, J.S. Molecular bases of high-level streptomycin resistance in Pseudomonas marginalis and Pseudomonas syringae pv. actinidiae. J. Microbiol. 2003, 41, 16–21. [Google Scholar]
- Jeyakumar, P.; Anderson, C.W.N.; Holmes, A.; Miller, S. Optimising copper sprays on kiwifruit: A review. In Proceedings of the 27th Annual Fertilizer & Lime Research Centre (FLRC) Workshop, Palmerston North, New Zealand, 18–20 February 2014. [Google Scholar]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Köhl, J.; Jones, J.B.; Aubertot, J.N. Thirteen decades of antimicrobial copper compounds applied in agriculture. A review. Agron. Sustain. Dev. 2018, 38, 28. [Google Scholar] [CrossRef] [Green Version]
- Tyson, J.L.; Dobson, S.J.; Manning, M.A. Effect of a protectant copper application on Psa infection of kiwifruit trap plants. N. Z. Plant Protect. 2017, 70, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ma, Y.B.; Hu, H.W.; Wang, J.T.; Liu, Y.R.; He, J.Z. Field-based evidence for consistent responses of bacterial communities to copper contamination in two contrasting agricultural soils. Front. Microbiol. 2015, 6, 31. [Google Scholar] [CrossRef]
- Colombi, E.; Straub, C.; Künzel, S.; Templeton, M.D.; McCann, H.C.; Rainey, P.B. Evolution of copper resistance in the kiwifruit pathogen Pseudomonas syringae pv. actinidiae through acquisition of integrative conjugative elements and plasmids. Environ. Microbiol. 2017, 19, 819–832. [Google Scholar] [CrossRef]
- Hu, H.W.; Wang, J.T.; Li, J.; Li, J.J.; Ma, Y.B.; Chen, D.; He, J.Z. Field-based evidence for copper contamination induced changes of antibiotic resistance in agricultural soils. Environ. Microbiol. 2016, 18, 3896–3909. [Google Scholar] [CrossRef] [PubMed]
- Ghods, S.; Sims, I.M.; Moradali, M.F.; Rehm, B.H. Bactericidal compounds controlling growth of the plant pathogen Pseudomonas syringae pv. actinidiae, which forms biofilms composed of a novel exopolysaccharide. Appl. Environ. Microbiol. 2015, 81, 4026–4036. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.; Zoysa, G.H.D.; Sarojini, V. Antimicrobial peptides against Pseudomonas syringae pv. actinidiae and Erwinia amylovora: Chemical synthesis, secondary structure, efficacy, and mechanistic investigations. Pept. Sci. 2014, 102, 88–96. [Google Scholar] [CrossRef]
- Oliveras, À.; Baró, A.; Montesinos, L.; Badosa, E.; Montesinos, E.; Feliu, L.; Planas, M. Antimicrobial activity of linear lipopeptides derived from BP100 towards plant pathogens. PLoS ONE 2018, 13, e0201571. [Google Scholar] [CrossRef] [PubMed]
- Camó, C.; Bonaterra, A.; Badosa, E.; Baró, A.; Montesinos, L.; Montesinos, E.; Planas, M.; Feliu, L. Antimicrobial peptide KSL-W and analogues: Promising agents to control plant diseases. Peptides 2019, 112, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Flores, O.; Retamales, J.; Núñez, M.; León, M.; Salinas, P.; Besoain, X.; Yañez, C.; Bastías, R. Characterization of bacteriophages against Pseudomonas syringae pv. actinidiae with potential use as natural antimicrobials in kiwifruit plants. Microorganisms 2020, 8, 974. [Google Scholar] [CrossRef] [PubMed]
- Vavala, E.; Passariello, C.; Pepi, F.; Colone, M.; Garzoli, S.; Ragno, R.; Pirolli, A.; Stringaro, A.; Angiolella, L. Antibacterial activity of essential oils mixture against Psa. Nat. Prod. Res. 2016, 30, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Pucci, N.; Orzali, L.; Modesti, V.; Lumia, V.; Brunetti, A.; Pilotti, M.; Loreti, S. Essential oils with inhibitory capacities on Pseudomonas syringae pv. actinidiae, the causal agent of kiwifruit bacterial canker. Asian J. Plant Pathol. 2018, 12, 16–26. [Google Scholar]
- Simonetti, G.; Pucci, N.; Brasili, E.; Valletta, A.; Sammarco, I.; Carnevale, E.; Pasqua, G.; Loreti, S. In vitro antimicrobial activity of plant extracts against Pseudomonas syringae pv. actinidiae causal agent of bacterial canker in kiwifruit. Plant Biosyst. Int. J. Deal. Aspects Plant Biol. 2020, 154, 100–106. [Google Scholar] [CrossRef]
- Wang, G.; Mishra, B. The importance of amino acid composition in natural AMPs: An evolutional, structural, and functional perspective. Front. Immunol. 2012, 3, 221. [Google Scholar]
- Aisenbrey, C.; Marquette, A.; Bechinger, B. The mechanisms of action of cationic antimicrobial peptides refined by novel concepts from biophysical investigations. Adv. Exp. Med. Biol. 2019, 1117, 33–64. [Google Scholar] [PubMed] [Green Version]
- Silva, F.A.C.; de Sousa Oliveira, M.; Souza, J.M.; Martins, P.G.S.; Pestana-Calsa, M.C.; Junior, T.C. Plant proteomics and peptidomics in host-pathogen interactions: The weapons used by each side. Curr. Prot. Pept. Sci. 2017, 18, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Mansour, S.C.; Hancock, R.E. Antimicrobial peptides: An introduction. Methods Mol. Biol. 2017, 1548, 3–22. [Google Scholar] [PubMed]
- Xu, L.; Shao, C.; Li, G.; Shan, A.; Chou, S.; Wang, J.; Ma, Q.; Dong, N. Conversion of broad-spectrum antimicrobial peptides into species-specific antimicrobials capable of precisely targeting pathogenic bacteria. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Li, J.; Koh, J.J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane active antimicrobial peptides: Translating mechanistic insights to design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.; Moghal, M.M.R.; Saha, S.K.; Yamazaki, M. The role of membrane tension in the action of antimicrobial peptides and cell-penetrating peptides in biomembranes. Biophys. Rev. 2019, 11, 431–448. [Google Scholar] [CrossRef] [PubMed]
- Hollmann, A.; Martinez, M.; Maturana, P.; Semorile, L.C.; Maffia, P.C. Antimicrobial peptides: Interaction with model and biological membranes and synergism with chemical antibiotics. Front. Chem. 2018, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Rios, A.C.; Moutinho, C.G.; Pinto, F.C.; Del Fiol, F.S.; Jozala, A.; Chaud, M.V.; Vila, M.M.; Teixeira, J.A.; Balcão, V.M. Alternatives to overcoming bacterial resistances: State-of-the-art. Microbiol. Res. 2016, 191, 51–80. [Google Scholar] [CrossRef]
- Ilyas, H.; Datta, A.; Bhunia, A. An approach towards structure based antimicrobial peptide design for use in development of transgenic plants: A strategy for plant disease management. Curr. Med. Chem. 2017, 24, 1350–1364. [Google Scholar] [CrossRef]
- Datta, A.; Ghosh, A.; Airoldi, C.; Sperandeo, P.; Mroue, K.H.; Jiménez-Barbero, J.; Kundu, P.; Ramamoorthy, A.; Bhunia, A. Antimicrobial peptides: Insights into membrane permeabilization, lipopolysaccharide fragmentation and application in plant disease control. Sci. Rep. 2015, 5, 11951. [Google Scholar] [CrossRef] [Green Version]
- Spohn, R.; Daruka, L.; Lázár, V.; Martins, A.; Vidovics, F.; Grézal, G.; Méhi, O.; Kintses, B.; Számel, M.; Jangir, P.K.; et al. Integrated evolutionary analysis reveals antimicrobial peptides with limited resistance. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Montesinos, E.; Badosa, E.; Cabrefiga, J.; Planas, M.; Feliu, L.; Bardají, E. Antimicrobial peptides for plant disease control. From discovery to application. In Small Wonders: Peptides for Disease Control; Rajasekaran, K., Cary, J.W., Jaynes, M., Montesinos, E., Eds.; American Chemical Society: Washington, DC, USA, 2012; Volume 1095, pp. 235–261. [Google Scholar]
- Cabrefiga, J.; Montesinos, E. Lysozyme enhances the bactericidal effect of BP100 peptide against Erwinia amylovora, the causal agent of fire blight of rosaceous plants. BMC Microbiol. 2017, 17, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inui Kishi, R.N.; Stach-Machado, D.; Singulani, J.D.L.; Dos Santos, C.T.; Fusco-Almeida, A.M.; Cilli, E.M.; Freitas-Astúa, J.; Picchi, S.C.; Machado, M.A. Evaluation of cytotoxicity features of antimicrobial peptides with potential to control bacterial diseases of citrus. PLoS ONE 2018, 13, e0203451. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, E. Antimicrobial peptides and plant disease control. FEMS Microbiol. Lett. 2007, 270, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baró, A.; Mora, I.; Montesinos, L.; Montesinos, E. Differential susceptibility of Xylella fastidiosa strains to synthetic bactericidal peptides. Phytopathology 2020, 110, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Velivelli, S.L.; Islam, K.T.; Hobson, E.; Shah, D.M. Modes of action of a bi-domain plant defensin MtDef5 against a bacterial pathogen Xanthomonas campestris. Front. Microb. 2018, 9, 934. [Google Scholar] [CrossRef]
- Rajasekaran, K.; Stromberg, K.D.; Cary, J.W.; Cleveland, T.E. Broad-spectrum antimicrobial activity in vitro of the synthetic peptide D4E1. J. Agric. Food Chem. 2001, 49, 2799–2803. [Google Scholar] [CrossRef] [PubMed]
- Topman, S.; Tamir-Ariel, D.; Bochnic-Tamir, H.; Stern Bauer, T.; Shafir, S.; Burdman, S.; Hayouka, Z. Random peptide mixtures as new crop protection agents. Microb. Biotech. 2018, 11, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.; Zhang, S.; Stover, E. Transgenic expression of antimicrobial peptide D2A21 confers resistance to diseases incited by Pseudomonas syringae pv. tabaci and Xanthomonas citri, but not Candidatus Liberibacter asiaticus. PLoS ONE 2017, 12, e0186810. [Google Scholar] [CrossRef]
- Zeitler, B.; Diaz, A.H.; Dangel, A.; Thellmann, M.; Meyer, H.; Sattler, M.; Lindermayr, C. De-novo design of antimicrobial peptides for plant protection. PLoS ONE 2013, 8, e71687. [Google Scholar] [CrossRef] [Green Version]
- Badosa, E.; Moiset, G.; Montesinos, L.; Talleda, M.; Bardají, E.; Feliu, L.; Planas, M.; Montesinos, E. Derivatives of the antimicrobial peptide BP100 for expression in plant systems. PLoS ONE 2013, 8, e85515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badosa, E.; Ferre, R.; Planas, M.; Feliu, L.; Besalú, E.; Cabrefiga, J.; Bardají, E.; Montesinos, E. A library of linear undecapeptides with bactericidal activity against phytopathogenic bacteria. Peptides 2007, 28, 2276–2285. [Google Scholar] [CrossRef]
- Nadal, A.; Montero, M.; Badosa, E.; Messeguer, J.; Montesinos, L.; Montesinos, E.; Pla, M. Constitutive expression of transgenes encoding derivatives of the synthetic antimicrobial peptide BP100: Impact on rice host plant fitness. BMC Plant. Biol. 2012, 12, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torcato, I.M.; Huang, Y.H.; Franquelim, H.G.; Gaspar, D.; Craik, D.J.; Castanho, M.A.; Henriques, S.T. Design and characterization of novel antimicrobial peptides, R-BP100 and RW-BP100, with activity against Gram-negative and Gram-positive bacteria. Biochim. Biophys. Acta 2013, 1828, 944–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oddo, A.; Thomsen, T.T.; Kjelstrup, S.; Gorey, C.; Franzyk, H.; Frimodt-Møller, N.; Løbner-Olesen, A.; Hansen, P.R. An amphipathic undecapeptide with all d-amino acids shows promising activity against colistin-resistant strains of Acinetobacter baumannii and a dual mode of action. Antimicrob. Agents Chemother. 2016, 60, 592–599. [Google Scholar] [CrossRef] [Green Version]
- De Lucca, A.J.; Bland, J.M.; Grimm, C.; Jacks, T.J.; Cary, J.W.; Jaynes, J.M.; Cleveland, T.E.; Walsh, T.J. Fungicidal properties, sterol binding, and proteolytic resistance of the synthetic peptide D4E1. Can. J. Microb. 1998, 44, 514–520. [Google Scholar] [CrossRef]
- Mentag, R.; Luckevich, M.; Morency, M.J.; Seguin, A. Bacterial disease resistance of transgenic hybrid poplar expressing the synthetic antimicrobial peptide D4E1. Tree Physiol. 2003, 23, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.J.; Won, H.S.; Choi, W.S.; Lee, B.J. De novo generation of antimicrobial LK peptides with a single tryptophan at the critical amphipathic interface. J. Pept. Sci. 2009, 15, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.S.M.; Bessa, L.J.; Fernandes, I.; Ferraz, R.; Mateus, N.; Gameiro, P.; Teixeira, C.; Gomes, P. Turning a collagenesis-inducing peptide into a potent antibacterial and antibiofilm agent against multidrug-resistant Gram-negative bacteria. Front. Microb. 2019, 10, 1915. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.M.; Maia, S.R.; Gomes, P.A.; Martins, M.C.L. Dhvar5 antimicrobial peptide (AMP) chemoselective covalent immobilization results on higher antiadherence effect than simple physical adsorption. Biomaterials 2015, 52, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Lupetti, A.; Van Dissel, J.T.; Brouwer, C.P.J.M.; Nibbering, P.H. Human antimicrobial peptides’ antifungal activity against Aspergillus fumigatus. Eur. J. Clin. Microb. Infect. Dis. 2008, 27, 1125–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Hertog, A.L.; Sang, H.W.W.F.; Kraayenhof, R.; Bolscher, J.G.; HOF, W.V.T.; Veerman, E.C.; Amerongen, A.V.N. Interactions of histatin 5 and histatin 5-derived peptides with liposome membranes: Surface effects, translocation and permeabilization. Biochem. J. 2004, 379, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Andreu, D.; Ubach, J.; Boman, A.; Wåhlin, B.; Wade, D.; Merrifield, R.B.; Boman, H.G. Shortened cecropin A-melittin hybrids. Significant size reduction retains potent antibiotic activity. FEBS Lett. 1992, 296, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Ciandrini, E.; Morroni, G.; Arzeni, D.; Kamysz, W.; Neubauer, D.; Kamysz, E.; Cirioni, O.; Brescini, L.; Baffone, W.; Campana, R. Antimicrobial activity of different antimicrobial peptides (AMPs) against clinical methicillin-resistant Staphylococcus aureus (MRSA). Curr. Top. Med. Chem. 2018, 18, 2116–2126. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.; Claro, B.; Silva, B.F.; Vale, N.; Gomes, P.; Gomes, M.S.; Funari, S.S.; Teixeira, J.; Uhríková, D.; Bastos, M. Unravelling a mechanism of action for a Cecropin A-Melittin hybrid antimicrobial peptide: The induced formation of multilamellar lipid stacks. Langmuir 2018, 34, 2158–2170. [Google Scholar] [CrossRef]
- Fields, F.R.; Manzo, G.; Hind, C.K.; Janardhanan, J.; Foik, I.P.; Carmo Silva, P.D.; Balsara, R.D.; Clifford, M.; Vu, H.M.; Ross, J.N.; et al. Synthetic antimicrobial peptide tuning permits membrane disruption and interpeptide synergy. ACS Pharmacol. Transl. Sci. 2020, 3, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Amso, Z.; Hayouka, Z. Antimicrobial random peptide cocktails: A new approach to fight pathogenic bacteria. Chem. Commun. 2019, 55, 2007–2014. [Google Scholar] [CrossRef]
- Rudilla, H.; Merlos, A.; Sans-Serramitjana, E.; Fuste, E.; Sierra, J.M.; Zalacain, A.; Vinuesa, T.; Vinas, M. New and old tools to evaluate new antimicrobial peptides. AIMS Microb. 2018, 4, 522. [Google Scholar] [CrossRef] [PubMed]
- Bessa, L.J.; Eaton, P.; Dematei, A.; Plácido, A.; Vale, N.; Gomes, P.; Delerue-Matos, C.; Sa Leite, J.R.; Gameiro, P. Synergistic and antibiofilm properties of ocellatin peptides against multidrug-resistant Pseudomonas aeruginosa. Future Microb. 2018, 13, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Goltermann, L.; Nielsen, P.E. PNA antisense targeting in bacteria: Determination of antibacterial activity (MIC) of PNA-peptide conjugates. Methods Mol. Biol. 2020, 2105, 231–239. [Google Scholar] [PubMed]
- Hong, J.; Guan, W.; Jin, G.; Zhao, H.; Jiang, X.; Dai, J. Mechanism of tachyplesin I injury to bacterial membranes and intracellular enzymes, determined by laser confocal scanning microscopy and flow cytometry. Microb. Res. 2015, 170, 69–77. [Google Scholar] [CrossRef] [PubMed]
- O’Brien-Simpson, N.M.; Pantarat, N.; Attard, T.J.; Walsh, K.A.; Reynolds, E.C. A rapid and quantitative flow cytometry method for the analysis of membrane disruptive antimicrobial activity. PLoS ONE 2016, 11, e0151694. [Google Scholar] [CrossRef] [Green Version]
- Behrendt, R.; White, P.; Offer, J. Advances in Fmoc solid-phase peptide synthesis. J. Pept. Sci. 2016, 22, 4–27. [Google Scholar] [CrossRef] [Green Version]
- Fernando, M.S.; Ligia, B.A.; Mayra, D.B.; Ignacio, I.F. A review of a promising therapeutic and agronomical alternative: Antimicrobial peptides from Capsicum sp. Afr. J. Biotech. 2011, 10, 19918–19928. [Google Scholar]
- López-García, B.; San Segundo, B.; Coca, M. Antimicrobial peptides as a promising alternative for plant disease protection. In Small Wonders: Peptides for Disease Control; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2012; Volume 1095, pp. 263–294. [Google Scholar]
- Vestaron. Available online: https://www.vestaron.com/ (accessed on 20 December 2020).
- Kumaresan, V.; Bhatt, P.; Ganesh, M.R.; Harikrishnan, R.; Arasu, M.; Al-Dhabi, N.A.; Pasupuleti, M.; Marimuthu, K.; Arockiaraj, J. A novel antimicrobial peptide derived from fish goose type lysozyme disrupts the membrane of Salmonella enterica. Mol. Immun. 2015, 68, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Raju, V.S.; Sarkar, P.; Pachaiappan, R.; Paray, B.A.; Al-Sadoon, M.K.; Arockiaraj, J. Defense involvement of piscidin from striped murrel Channa striatus and its peptides CsRG12 and CsLC11 involvement in an antimicrobial and antibiofilm activity. Fish. Shellfish Immun. 2020, 99, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, M.M.; Eftekhary, M.; Erfanimanesh, S.; Hashemi, A.; Omrani, V.F.; Farhadihosseinabadi, B.; Lasjerdi, Z.; Mossahebi-Mohammadi, M.; Chauhan, N.P.S.; Seifalian, A.M.; et al. Comparison of the antibacterial effects of a short cationic peptide and 1% silver bioactive glass against extensively drug-resistant bacteria, Pseudomonas aeruginosa and Acinetobacter baumannii, isolated from burn patients. Amino Acids 2018, 50, 1617–1628. [Google Scholar] [CrossRef]
- Duperthuy, M. Antimicrobial peptides: Virulence and resistance modulation in gram-negative bacteria. Microorganisms 2020, 8, 280. [Google Scholar] [CrossRef] [Green Version]
- Vasilchenko, A.S.; Rogozhin, E.A. Sub-inhibitory effects of antimicrobial peptides. Front. Microb. 2019, 10, 1160. [Google Scholar] [CrossRef] [Green Version]
- Chung, P.Y.; Khanum, R. Antimicrobial peptides as potential anti-biofilm agents against multidrug-resistant bacteria. J. Microb. Immun. Infect. 2017, 50, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Benincasa, M.; Barrière, Q.; Runti, G.; Pierre, O.; Bourge, M.; Scocchi, M.; Mergaert, P. Single cell flow cytometry assay for peptide uptake by bacteria. Bio-Protocol 2016, 6, e2038. [Google Scholar] [CrossRef] [Green Version]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Comparative mode of action of the antimicrobial peptide melimine and its derivative Mel4 against Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 7063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefani, E.; Loreti, S. PM 7/120 (1) Pseudomonas syringae pv. actinidiae. EPPO Bull. 2014, 44, 360–375. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | BP100 | RW-BP100 | CA-M | 3.1 | ||||
|---|---|---|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | |
| AL114b | 3.4 | 3.4 | 6.2 | 6.2 | 3.4 | 10 | 25 | 25 |
| AL115 | 3.4 | 3.4 | 6.2 | 6.2 | 3.4 | 6.2 | 10 | 10 |
| CFBP7286 R | 3.4 | 3.4 | 3.4 | 6.2 | 3.4 | 6.2 | 10 | 25 |
| Fv62 | 3.4 | 10 | 6.2 | 6.2 | 3.4 | 10 | 10 | 25 |
| P85 | 3.4 | 6.2 | 6.2 | 6.2 | 6.2 | 6.2 | 25 | 25 |
| VN29 | 3.4 | 6.2 | 6.2 | 6.2 | 6.2 | 6.2 | 10 | 25 |
| VV10 | 3.4 | 6.2 | 6.2 | 6.2 | 3.4 | 6.2 | 10 | 25 |
| VV112 | 3.4 | 10 | 6.2 | 6.2 | 3.4 | 6.2 | 10 | 25 |
| Bacterial Strains | Host Plant | Origin | Year of Isolation |
|---|---|---|---|
| Al114a | Actinidia deliciosa | Amares, Portugal | 2014 |
| AL114b | A. deliciosa | Amares, Portugal | 2014 |
| AL115 | A. deliciosa | Amares, Portugal | 2014 |
| AL116b | A. deliciosa | Amares, Portugal | 2014 |
| AL13 | A. deliciosa | Amares, Portugal | 2013 |
| Am63 | A. deliciosa | Amarante, Portugal | 2013 |
| Fv62 | A. deliciosa | Felgueiras, Portugal | 2013 |
| P18 | A. deliciosa | Vila de Prado, Portugal | 2013 |
| P84 | A. deliciosa | Vila de Prado, Portugal | 2013 |
| P85 | A. deliciosa | Vila de Prado, Portugal | 2013 |
| P93 | A. deliciosa | Vila de Prado, Portugal | 2013 |
| Pn16 | A. deliciosa | Penafiel, Portugal | 2013 |
| VC104b | A. deliciosa | Vila do Conde, Portugal | 2013 |
| VN23 | A. deliciosa | Valença, Portugal | 2016 |
| VN28 | A. deliciosa | Valença, Portugal | 2017 |
| VN29 | A. deliciosa | Valença, Portugal | 2017 |
| VV3 | A. deliciosa | Valença, Portugal | 2016 |
| VV10 | A. deliciosa | Valença, Portugal | 2017 |
| VV14 | A. deliciosa | Valença, Portugal | 2017 |
| VV15 | A. deliciosa | Valença, Portugal | 2017 |
| VV112 | A. deliciosa | Vila Verde, Portugal | 2014 |
| VV113 | A. deliciosa | Vila Verde, Portugal | 2014 |
| CFBP7286 R, a | A. chinensis | Latina, Italy | 2008 |
| Peptide | Sequence | Net Charge a,b | MW (Da) b |
|---|---|---|---|
| BP100 | KKLFKKILKYL-NH2 | 6 | 1419.9 |
| RW-BP100 | RRLFRRILRWL-NH2 | 6 | 1583.0 |
| CA-M | KWKLFKKIGAVLKVL-NH2 | 6 | 1769.2 |
| D4E1 | FKLRAKIKVRLRAKIKL-NH2 | 9 | 2079.4 |
| 3.1 | KKLLKWLLKLL-NH2 | 5 | 1393.9 |
| Dhvar-5 | LLLFLLKKRKKRKY-NH2 | 8 | 1845.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariz-Ponte, N.; Regalado, L.; Gimranov, E.; Tassi, N.; Moura, L.; Gomes, P.; Tavares, F.; Santos, C.; Teixeira, C. A Synergic Potential of Antimicrobial Peptides against Pseudomonas syringae pv. actinidiae. Molecules 2021, 26, 1461. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051461
Mariz-Ponte N, Regalado L, Gimranov E, Tassi N, Moura L, Gomes P, Tavares F, Santos C, Teixeira C. A Synergic Potential of Antimicrobial Peptides against Pseudomonas syringae pv. actinidiae. Molecules. 2021; 26(5):1461. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051461
Chicago/Turabian StyleMariz-Ponte, Nuno, Laura Regalado, Emil Gimranov, Natália Tassi, Luísa Moura, Paula Gomes, Fernando Tavares, Conceição Santos, and Cátia Teixeira. 2021. "A Synergic Potential of Antimicrobial Peptides against Pseudomonas syringae pv. actinidiae" Molecules 26, no. 5: 1461. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051461