
Bioguided Fractionation of Local Plants against Matrix Metalloproteinase9 and Its Cytotoxicity against Breast Cancer Cell Models: In Silico and In Vitro Study (Part II)

 ,
,  ,
, 

 ,
,
Abstract
:1. Introduction
2. Results
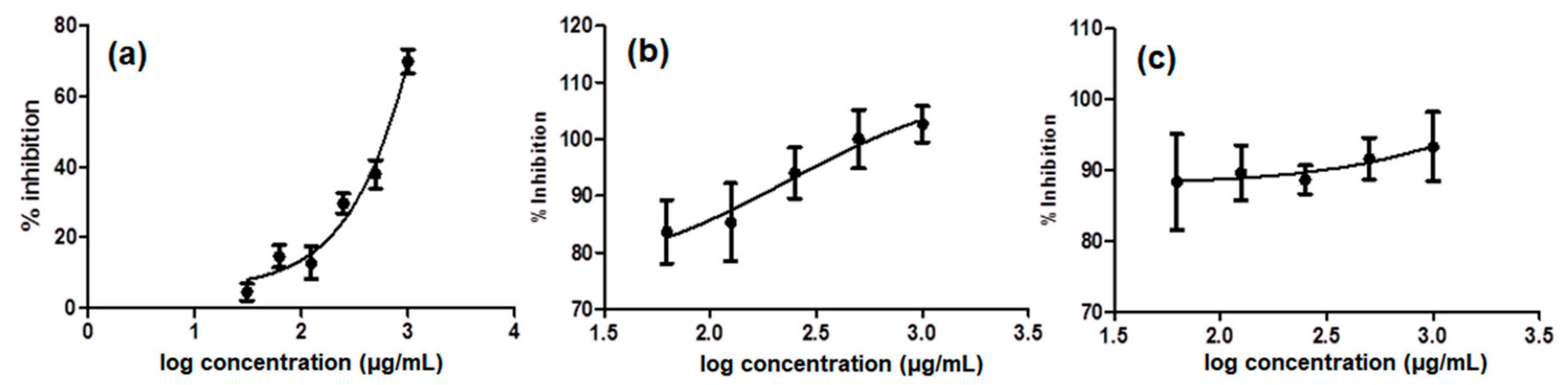
2.1. MMP9 In Vitro Assay
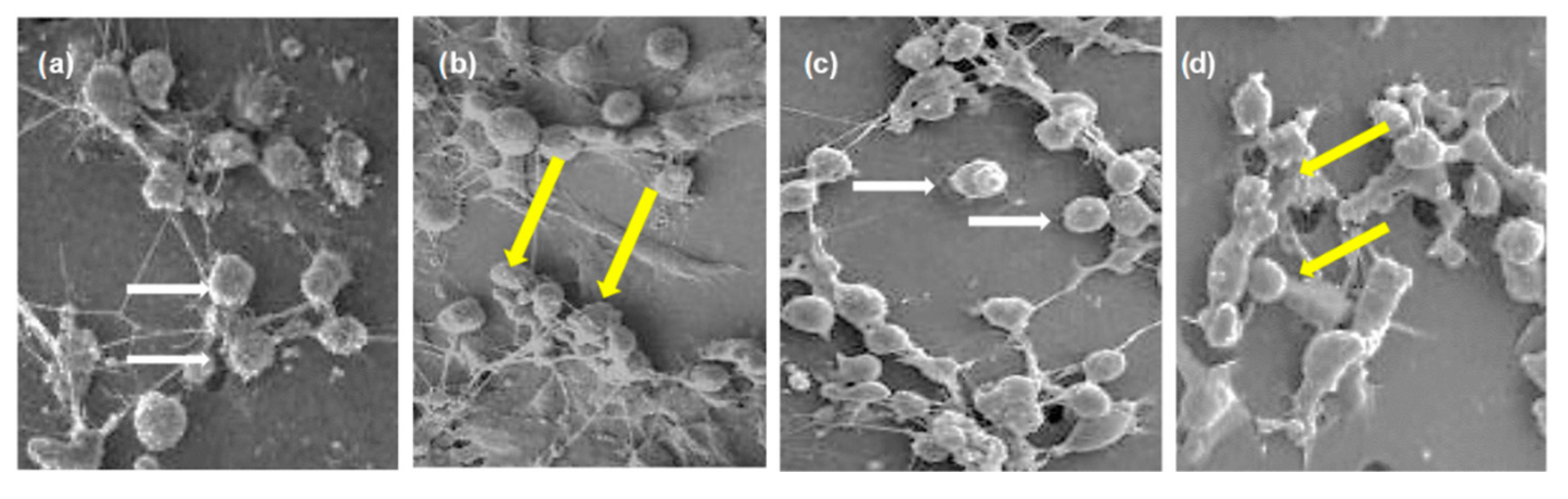
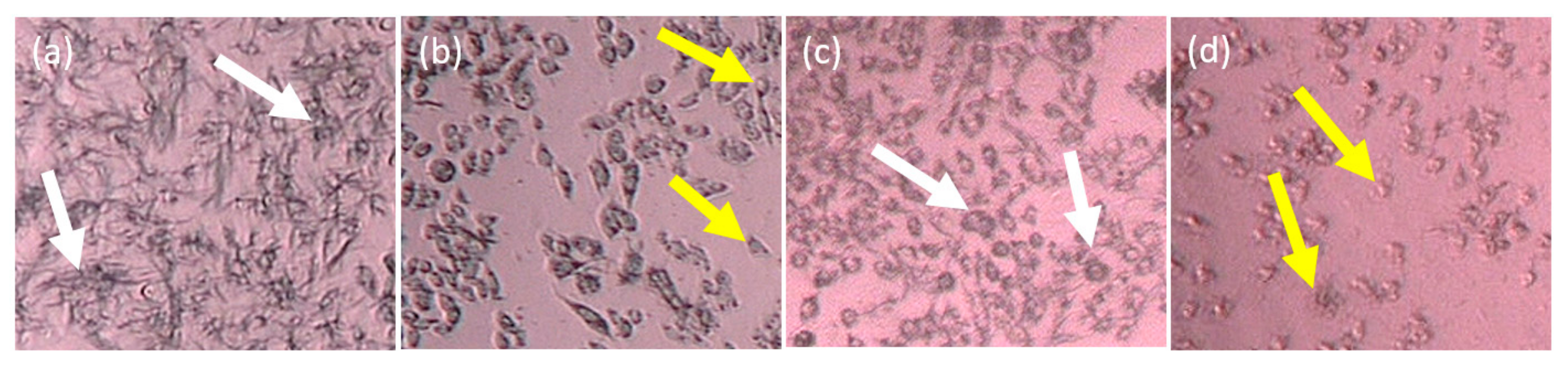
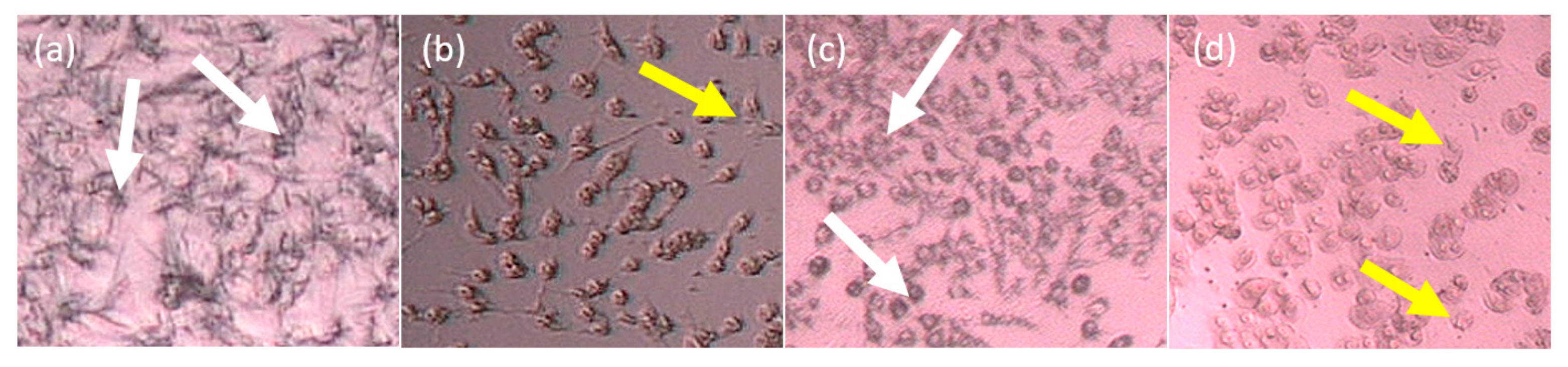
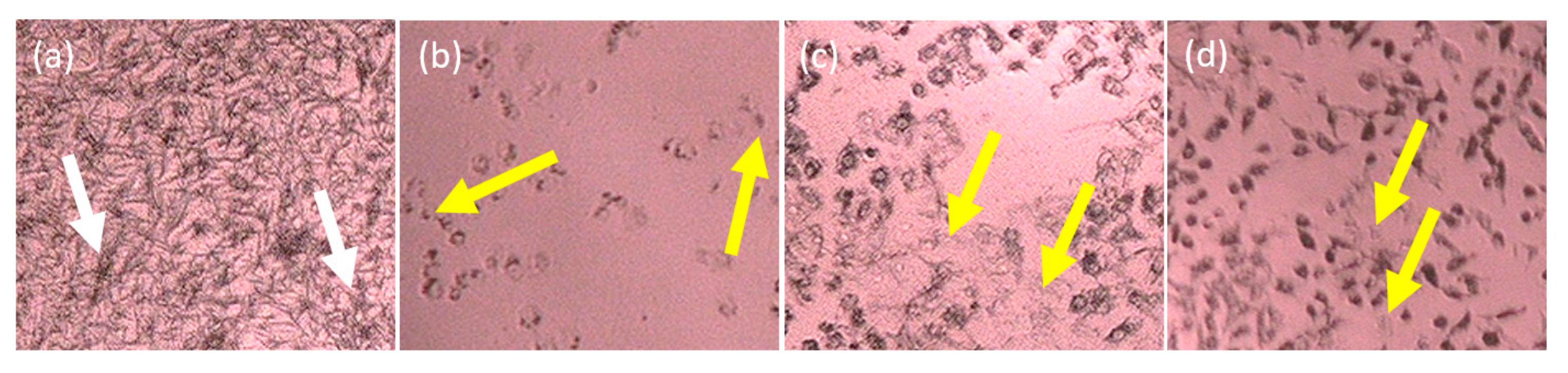
2.2. Cytotoxicity In Vitro Assay of the Partitions
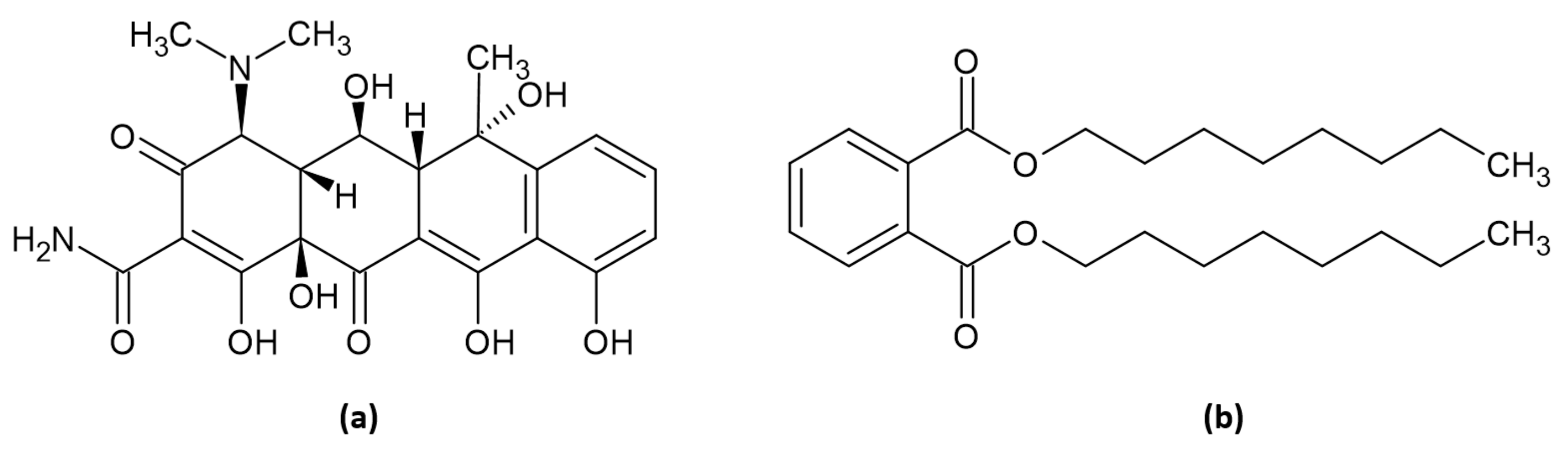
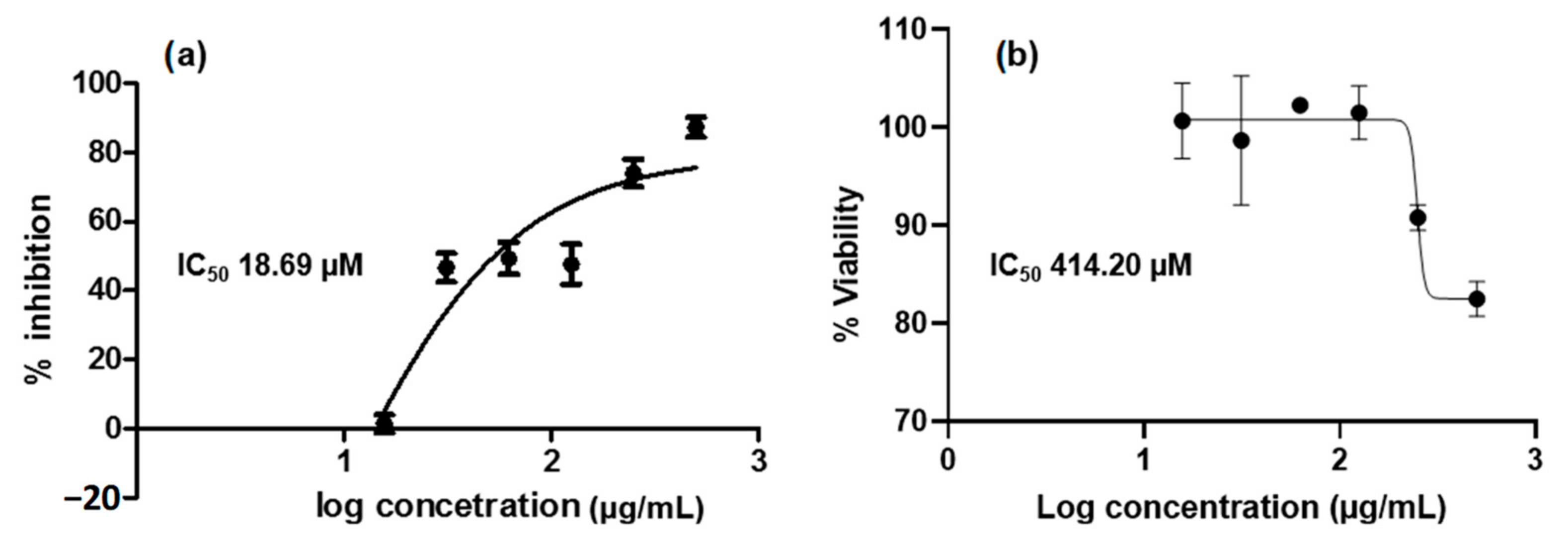
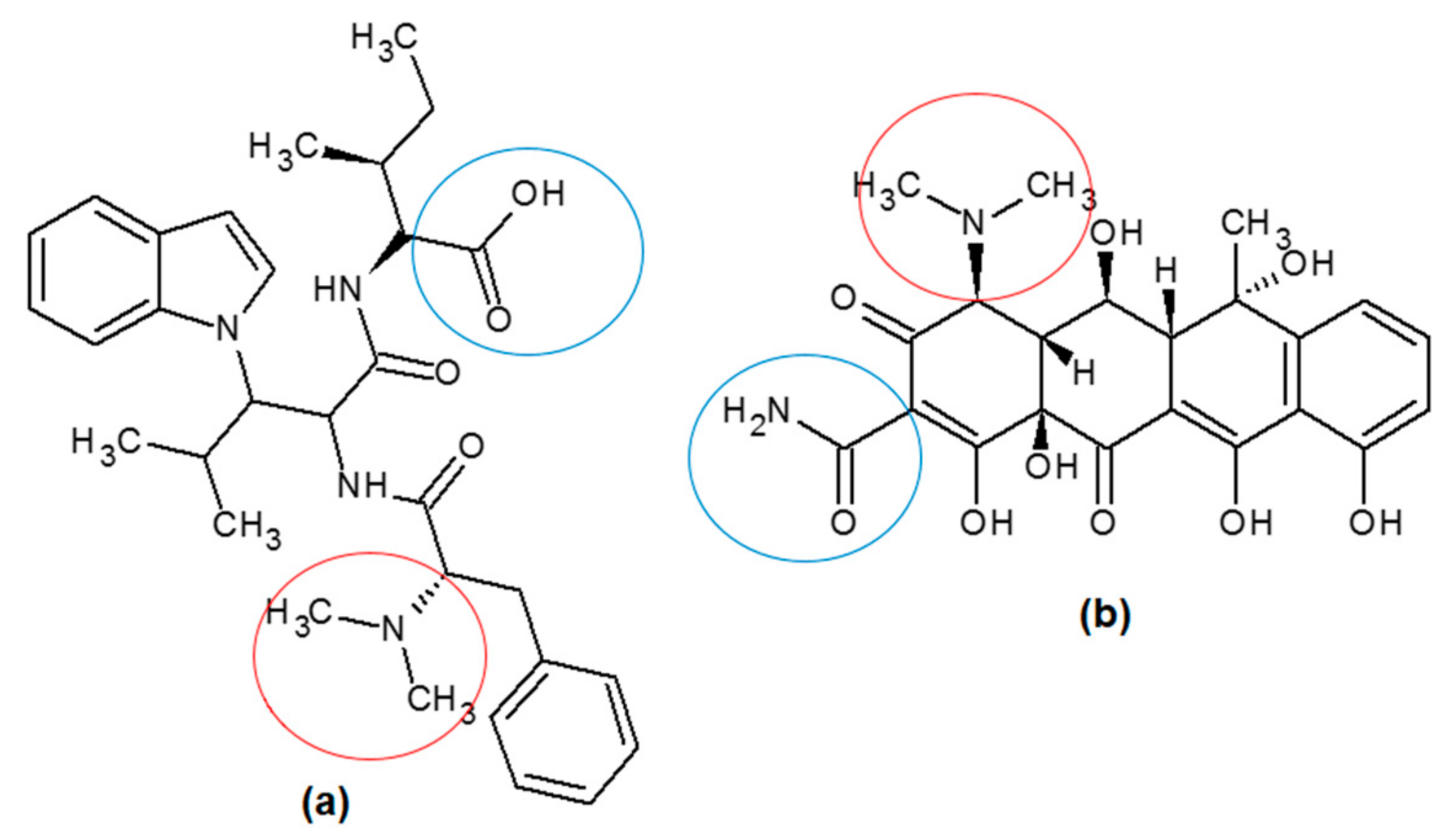
2.3. Isolation and In Vitro Assay
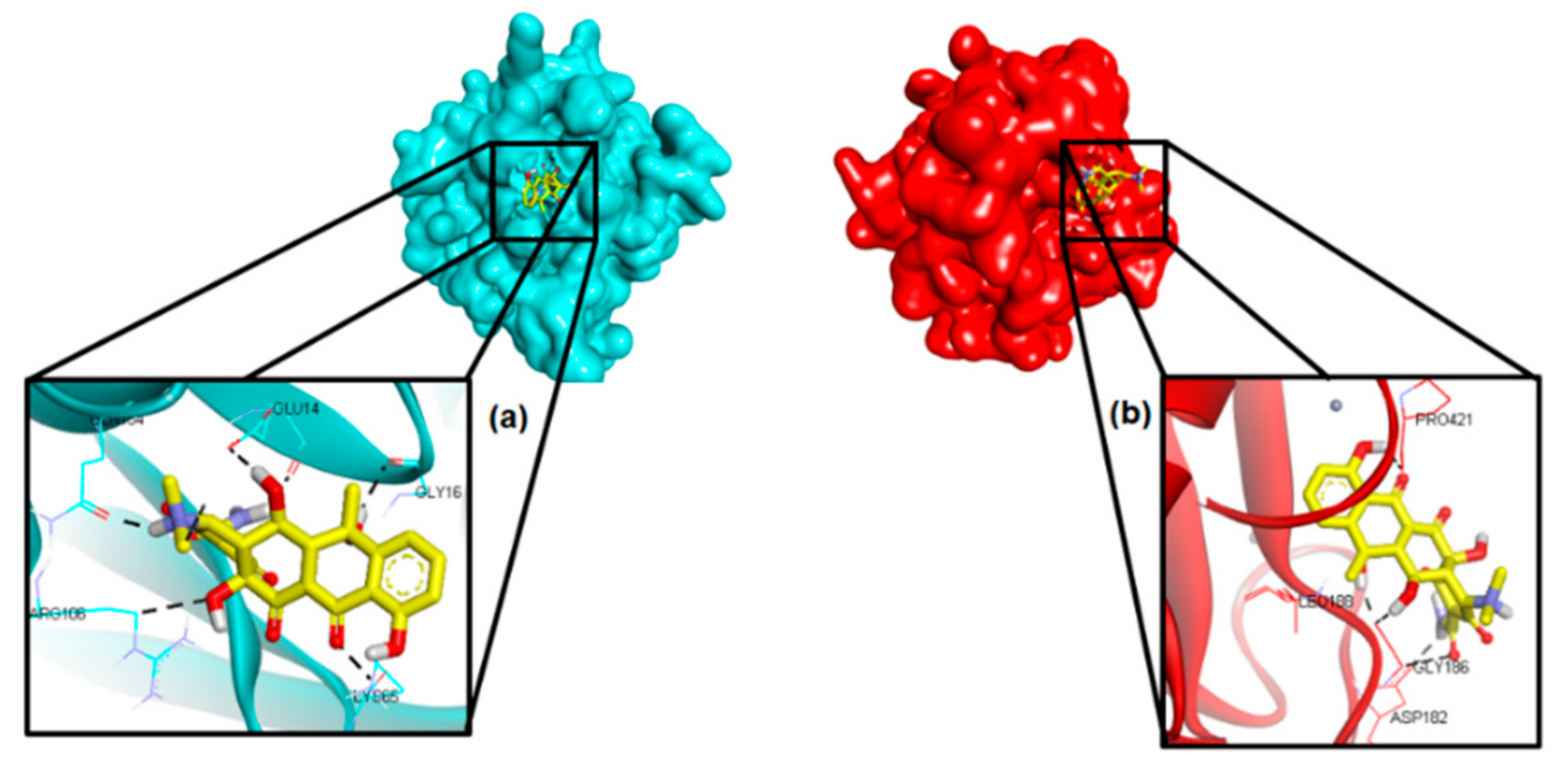
2.4. Molecular Docking
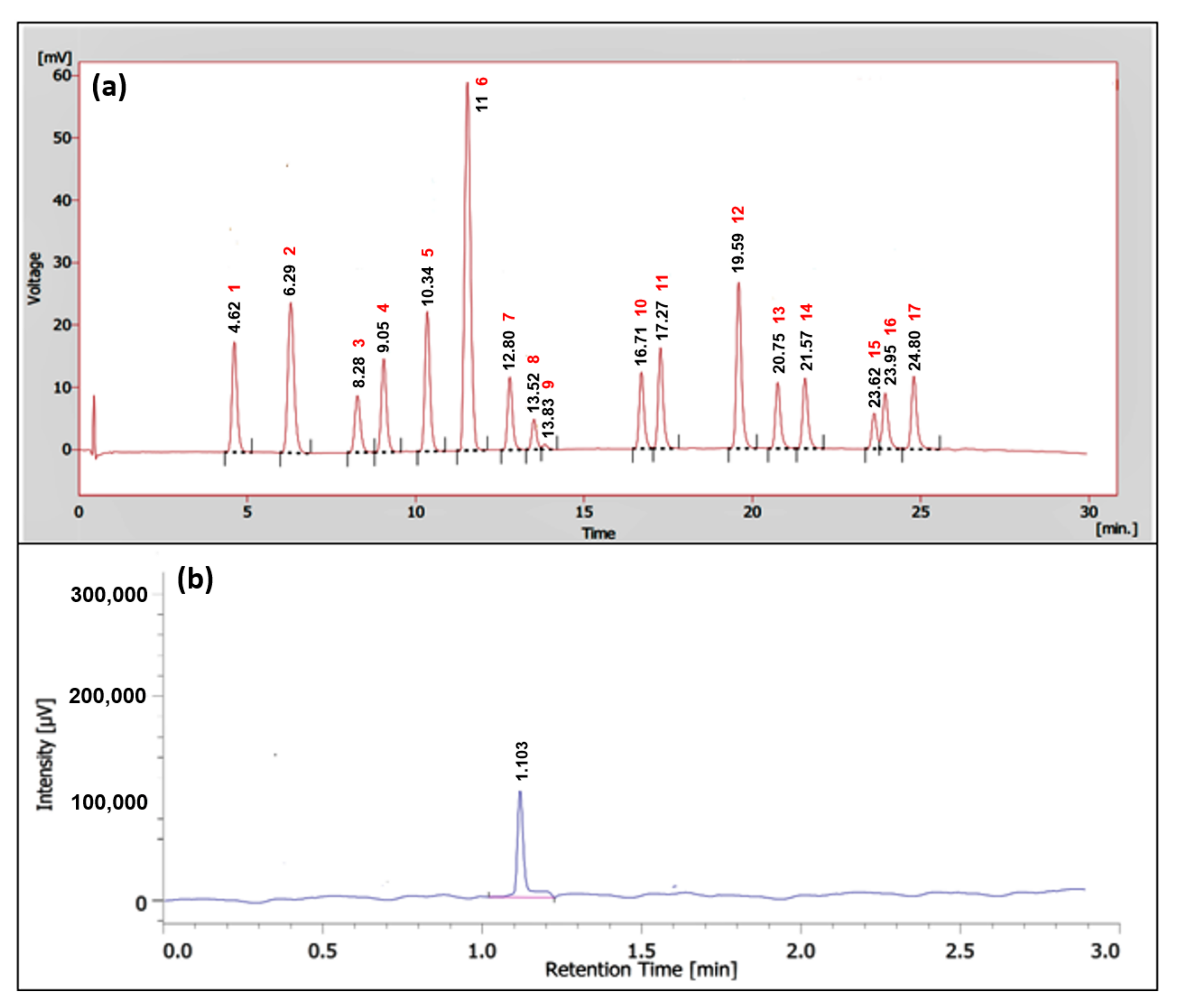
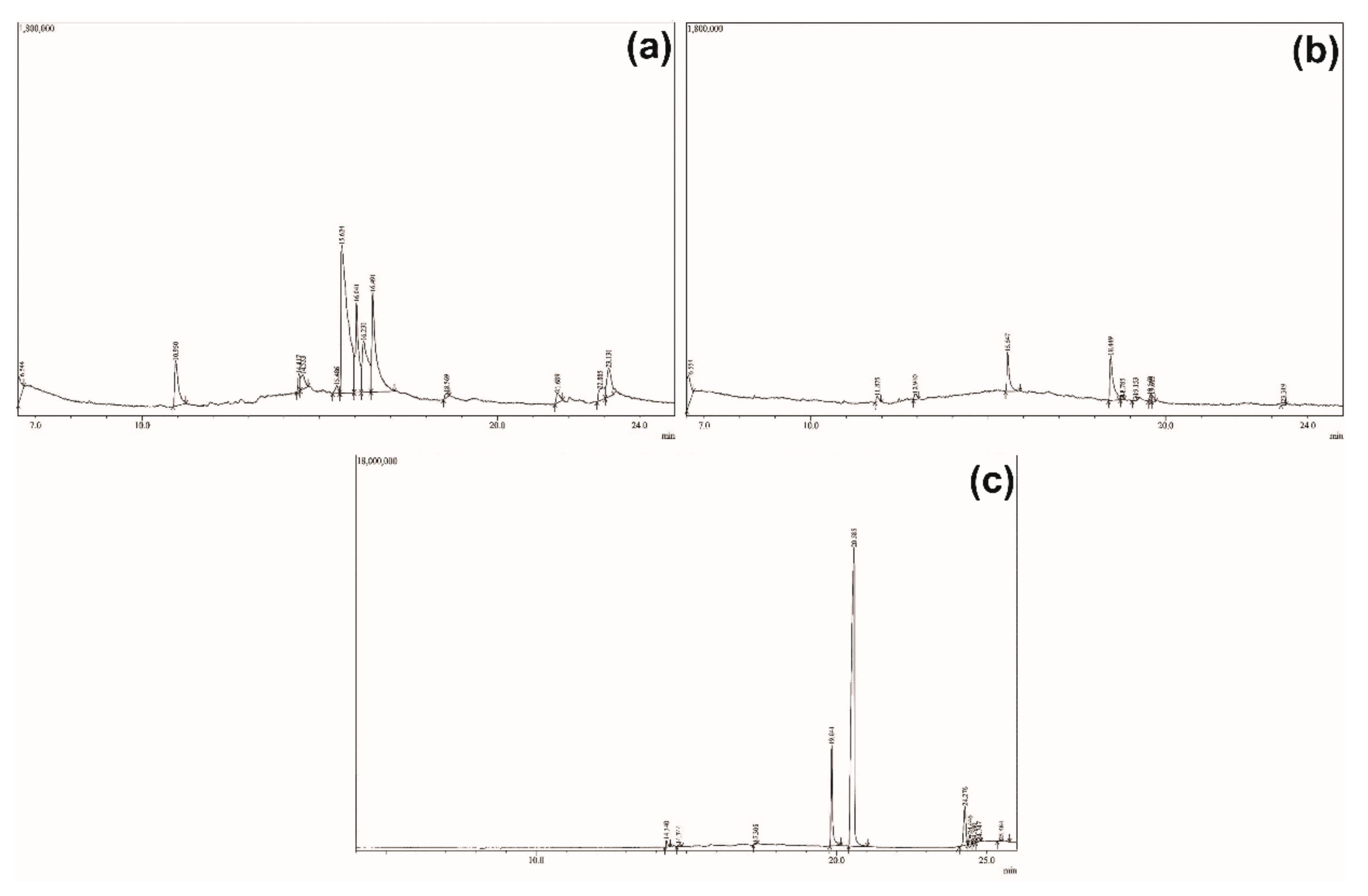
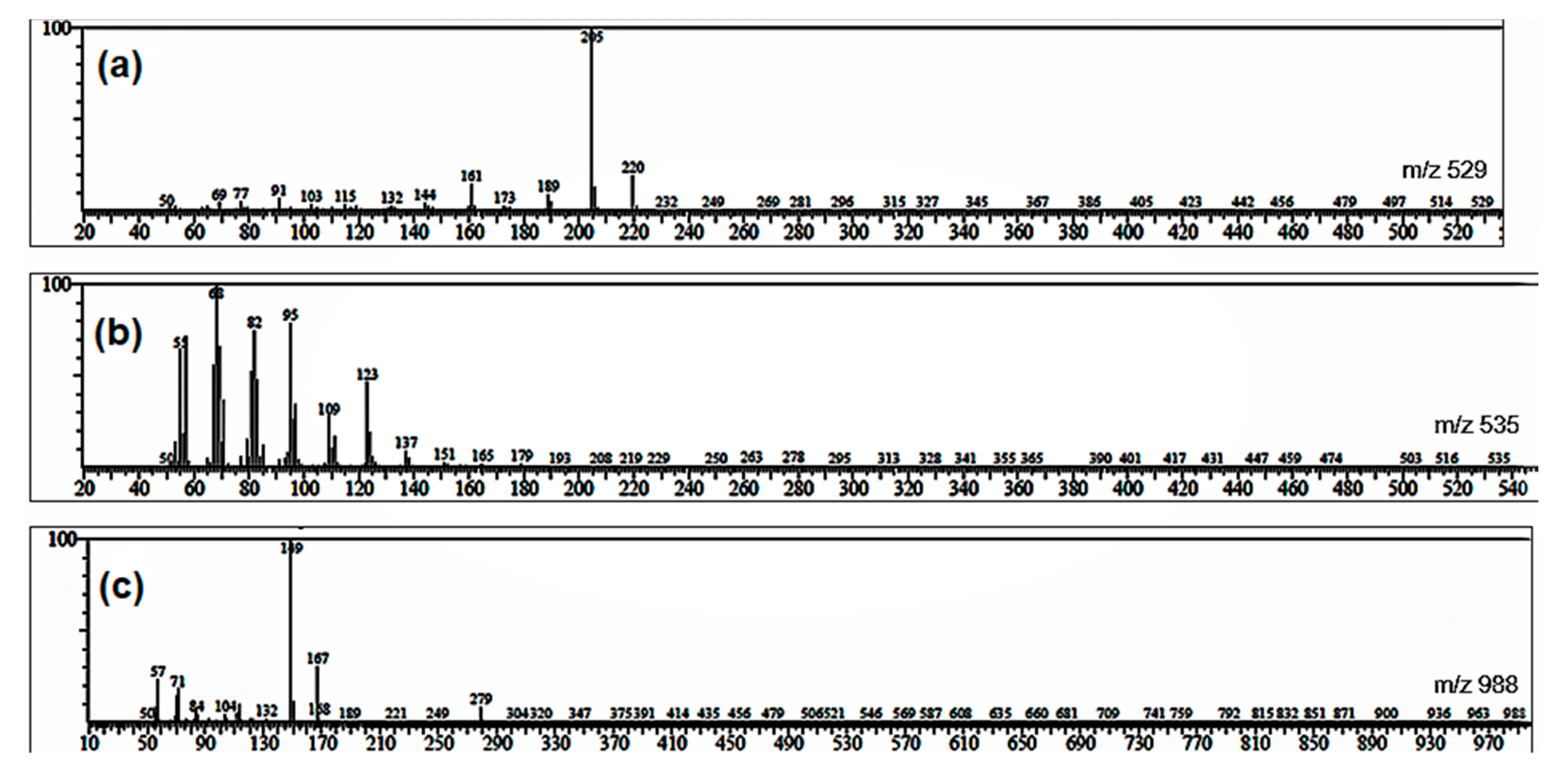
2.5. GC-MS Profiles of the Partitions
3. Discussion
4. Materials and Methods
4.1. Software and Hardware
4.2. Chemicals
4.3. In Vitro MMP9 Inhibition Assay
4.4. In Vitro Cytotoxicity Assay
4.5. Isolation
4.6. Molecular Docking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Ferrario, A.; Stephens, P.; Guan, X.; Ross-Degnan, D.; Wagner, A. Sales of anti-cancer medicines; China, Indonesia, Kazakhstan, Malaysia, Philippines and Thailand. Bull. World Health Organ. 2020, 98, 467. [Google Scholar] [CrossRef]
- Anwar, S.L.; Avanti, W.S.; Nugroho, A.C.; Choridah, L.; Dwianingsih, E.K.; Harahap, W.A.; Aryandono, T.; Wulaningsih, W. Risk factors of distant metastasis after surgery among different breast cancer subtypes: A hospital-based study in Indonesia. World J. Surg. Oncol. 2020, 18, 117. [Google Scholar] [CrossRef]
- Hariono, M.; Nuwarda, R.F.; Yusuf, M.; Rollando, R.; Jenie, R.I.; Al-Najjar, B.; Julianus, J.; Putra, K.C.; Nugroho, E.S.; Wisnumurti, Y.K.; et al. Arylamide as potential selective inhibitor for matrix metalloproteinase 9 (MMP9): Design, synthesis, biological evaluation, and molecular modeling. J. Chem. Inf. Model. 2020, 60, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Chen, S.; Huang, L.; Shao, Z.M. Predictive and prognostic value of Matrix metalloproteinase (MMP)-9 in neoadjuvant chemotherapy for triple-negative breast cancer patients. BMC Cancer 2018, 18, 909. [Google Scholar] [CrossRef] [PubMed]
- Adhipandito, C.F.; Ludji, D.P.K.S.; Aprilianto, E.; Jenie, R.I.; Al-Najjar, B.; Hariono, M. Matrix metalloproteinase9 as the protein target in anti-breast cancer drug discovery: An approach by targeting hemopexin domain. Future J. Pharm. Sci. 2019, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Krzeski, P.; Buckland-Wright, C.; Bálint, G.; Cline, G.A.; Stoner, K.; Lyon, R.; Beary, J.; Aronstein, W.S.; Spector, T.D. Development of musculoskeletal toxicity without clear benefit after administration of PG-116800, a matrix metalloproteinase inhibitor, to patients with knee osteoarthritis: A randomized, 12-month, double-blind, placebo-controlled study. Arthritis Res. Ther. 2007, 9, R109. [Google Scholar] [CrossRef] [Green Version]
- Hariono, M.; Rollando, R.; Karamoy, J.; Hariyono, P.; Atmono, M.; Djohan, M.; Wiwy, W.; Nuwarda, R.; Kurniawan, C.; Salin, N.; et al. Bioguided fractionation of local plants against matrix metalloproteinase9 and its cytotoxicity against breast cancer cell models: In silico and in vitro study. Molecules 2020, 25, 4691. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J. Animal models for studying prevention and treatment of breast cancer. In Animal Models for the Study of Human Disease, 1st ed.; Conn, P.M., Ed.; Elsevier Inc.: Cambridge, MA, USA, 2013; pp. 997–1018. [Google Scholar]
- Lv, Y.; Zhao, X.; Zhu, L.; Li, S.; Xiao, Q.; He, W.; Yin, L. Targeting intracellular MMPs efficiently inhibits tumor metastasis and angiogenesis. Theranostics 2018, 8, 2830. [Google Scholar] [CrossRef] [PubMed]
- Aka, J.A.; Lin, S.X. Comparison of functional proteomic analyses of human breast cancer cell lines T47D and MCF7. PLoS ONE 2012, 7, e31532. [Google Scholar] [CrossRef]
- van Kuppeveld, F.J.; Johansson, K.E.; Galama, J.M.; Kissing, J.; Bölske, G.; van der Logt, J.T.; Melchers, W.J. Detection of mycoplasma contamination in cell cultures by a mycoplasma group-specific PCR. Appl. Environ. Microbiol. 1994, 60, 149–152. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.; Tamez, P.; Pezzuto, J.; Soejarto, D. Evaluation of plants used for antimalarial treatment by the Massai of Kenya. J. Ethnopharmacol. 2005, 101, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, J.; Cadilha, B.L.; Markota, A.; Voigt, C.; Huang, Z.; Lin, P.P.; Wang, D.D.; Dai, J.; Kranz, G.; et al. Epithelial-type systemic breast carcinoma cells with a restricted mesenchymal transition are a major source of metastasis. Sci. Adv. 2019, 2, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenny, P.A.; Lee, G.Y.; Myers, C.A.; Neve, R.M.; Semeiks, J.R.; Spellman, P.T.; Lorenz, K.; Lee, E.H.; Barcellos-Hoff, M.H.; Petersen, O.W. The morphologies of breast cancer cell lines in three-dimensional assays correlate with their profiles of gene expression. Mol. Oncol. 2007, 1, 84–96. [Google Scholar] [CrossRef]
- Cooper, G.M. The Cell, a Molecular Approach, 2nd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2000. [Google Scholar]
- Goncalves, E.A.; Ventura, C.A.; Yano, T.; Macedo, M.L.R.; Genari, S.C. Morphological and growth alterations in Vero cells transformed by cisplatin. Cell Biol. Int. 2006, 30, 485–494. [Google Scholar] [CrossRef]
- Kotta, J.C.; Lestari, A.B.S.; Candrasari, D.S.; Hariono, M. Medicinal effect, in silico bioactivity prediction, and pharmaceutical formulation of Ageratum conyzoides L.: A review. Scientifica 2020, 2020, 6420909. [Google Scholar] [CrossRef]
- Dufour, A.; Sampson, N.S.; Li, J.; Kuscu, C.; Rizzo, R.C.; DeLeon, J.L.; Zhi, J.; Jaber, N.; Liu, E.; Zucker, S.; et al. Small-molecule anticancer compounds selectively target the hemopexin domain of matrix metalloproteinase-9. Cancer Res. 2011, 71, 4977–4988. [Google Scholar] [CrossRef] [Green Version]
- Alford, V.M.; Kamath, A.; Ren, X.; Kumar, K.; Gan, Q.; Awwa, M.; Tong, M.; Seeliger, M.A.; Cao, J.; Ojima, I.; et al. Targeting the hemopexin-like domain of latent matrix metalloproteinase-9 (proMMP-9) with a small molecule inhibitor prevents the formation of focal adhesion junctions. ACS Chem. Biol. 2017, 12, 2788–2803. [Google Scholar] [CrossRef] [PubMed]
- Versiani, M.A.; Ikram, A.; Khalid, S.; Faizi, S.; Tahiri, I.A. Ixoroid: A new triterpenoid from the flowers of Ixora coccinea. Nat. Prod. Commun. 2012, 7, 1934578X1200700706. [Google Scholar] [CrossRef] [Green Version]
- Netzel-Arnett, S.; Mallya, S.K.; Nagase, H.; Birkedal-Hansen, H.; Van Wart, H.E. Continuously recording fluorescent assays optimized for five human matrix metalloproteinases. Anal. Biochem. 1991, 195, 86–92. [Google Scholar] [CrossRef]
- Lee, C.L.; Liao, Y.C.; Hwang, T.L.; Wu, C.C.; Chang, F.R.; Wu, Y.C. Ixorapeptide I and ixorapeptide II, bioactive peptides isolated from Ixora coccinea. Bioorg. Med. Chem. Lett. 2010, 20, 7354–7357. [Google Scholar] [CrossRef]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast cancer cell line classification and its relevance with breast tumor subtyping. J. Cancer 2017, 8, 3131. [Google Scholar] [CrossRef] [Green Version]
- Mardianingrum, R.; Yusuf, M.; Hariono, M.; Mohd Gazzali, A.; Muchtaridi, M. α-Mangostin and its derivatives against estrogen receptor alpha. J. Biomol. Struct. Dyn. 2020, 1–14. [Google Scholar] [CrossRef]
- Bao, L.; Haque, A.; Jackson, K.; Hazari, S.; Moroz, K.; Jetly, R.; Dash, S. Increased expression of P-glycoprotein is associated with doxorubicin chemoresistance in the metastatic 4T1 breast cancer model. Am. J. Pathol. 2011, 178, 838–852. [Google Scholar] [CrossRef] [Green Version]
- Sztalmachova, M.; Gumulec, J.; Raudenska, M.; Polanska, H.; Holubova, M.; Balvan, J.; Hudcova, K.; Knopfova, L.; Kizek, R.; Adam, V.; et al. Molecular response of 4T1-induced mouse mammary tumours and healthy tissues to zinc treatment. Int. J. Oncol. 2015, 46, 1810–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickens, L.B.; Tang, Y. Oxytetracycline biosynthesis. J. Biol. Chem. 2010, 285, 27509–27515. [Google Scholar] [CrossRef] [Green Version]
- Helfrich, E.J.; Vogel, C.M.; Ueoka, R.; Schäfer, M.; Ryffel, F.; Müller, D.B.; Probst, S.; Kreuzer, M.; Piel, J.; Vorholt, J.A. Bipartite interactions, antibiotic production and biosynthetic potential of the Arabidopsis leaf microbiome. Nat. Microbiol. 2018, 3, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Samartzis, E.P.; Fink, D.; Stucki, M.; Imesch, P. Doxycycline reduces MMP-2 activity and inhibits invasion of 12Z epithelial endometriotic cells as well as MMP-2 and-9 activity in primary endometriotic stromal cells in vitro. Reprod. Biol. Endocrinol. 2019, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Benko, C.; Gill, S.E.; Dufour, A. The pharmacological TAILS of matrix metalloproteinases and their inhibitors. Pharmaceuticals 2021, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Arnoczky, S.P.; Lavagnino, M.; Gardner, K.L.; Tian, T.; Vaupel, Z.M.; Stick, J.A. In vitro effects of oxytetracycline on matrix metalloproteinase-1 mRNA expression and on collagen gel contraction by cultured myofibroblasts obtained from the accessory ligament of foals. Am. J. Vet. Res. 2004, 65, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Fugler, L.A.; Eades, S.C.; Moore, R.M.; Koch, C.E.; Keowen, M.L. Plasma matrix metalloproteinase activity in horses after intravenous infusion of lipopolysaccharide and treatment with matrix metalloproteinase inhibitors. Am. J. Vet. Res. 2013, 74, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Anusha, P.; Thangaviji, V.; Velmurugan, S.; Michaelbabu, M.; Citarasu, T. Protection of ornamental gold fish Carassius auratus against Aeromonas hydrophila by treating Ixora coccinea active principles. Fish Shellfish Immunol. 2014, 36, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.N. Bioactive natural derivatives of phthalate ester. Crit. Rev. Biotechnol. 2020, 40, 913–929. [Google Scholar] [CrossRef]
- Kini, R.M.; Evans, H.J. A novel approach to the design of potent bioactive peptides by incorporation of proline brackets: Antiplatelet effects of Arg-Gly-Asp peptides. FEBS Lett. 1995, 375, 15–17. [Google Scholar] [CrossRef] [Green Version]
- Rengasamy, K.R.; Khan, H.; Ahmad, I.; Lobine, D.; Mahomoodally, F.; Suroowan, S.; Hassan, S.T.; Xu, S.; Patel, S.; Daglia, M. Bioactive peptides and proteins as alternative antiplatelet drugs. Med. Res. Rev. 2019, 39, 2153–2171. [Google Scholar] [CrossRef]
- Cha, H.; Kopetzki, E.; Huber, R.; Lanzendörfer, M.; Brandstetter, H. Structural basis of the adaptive molecular recognition by MMP9. J. Mol. Biol. 2002, 320, 1065–1079. [Google Scholar] [CrossRef]
- Rowsell, S.; Hawtin, P.; Minshull, C.A.; Jepson, H.; Brockbank, S.M.V.; Barratt, D.G.; Slater, A.M.; McPheat, W.L.; Waterson, D.; Henney, A.M.; et al. Crystal structure of human MMP9 in complex with a reverse hydroxamate inhibitor. J. Mol. Biol. 2002, 319, 173–181. [Google Scholar] [CrossRef]
- Franchi, M.; Piperigkou, Z.; Karamanos, K.A.; Franchi, L.; Masola, V. Extracellular matrix-mediated breast cancer cells morphological alterations, invasiveness, and microvesicles/exosomes release. Cells 2020, 9, 2031. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Hevener, K.E.; Zhao, W.; Ball, D.M.; Babaoglu, K.; Qi, J.; White, S.W.; Lee, R.E. Validation of molecular docking programs for virtual screening against dihydropteroate synthase. J. Chem. Inf. Model. 2009, 49, 444–460. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | IC50 (µg/mL) | |||||
|---|---|---|---|---|---|---|
| MDA-MB-231 (R2) | 4T1 (R2) | T47D (R2) | MCF7 (R2) | Vero (R2) | ||
| 1 | AC n-hexane | 2.05 (0.98) | 265 (0.78) | 109.70 (0.98) | 2.11 (0.97) | 217.2 (0.99) |
| 2 | IC n-hexane | NA | 225.5(0.73) | 1320 (0.86) | NA | 972.2 (0.85) |
| 3 | IC ethylacetate | 1.92 (0.99) | 57.5 (0.85) | 371.5 (0.95) | 2.01 (0.98) | 429.5 (0.99) |
| 4 | doxorubicin | NA | 388.4 (0.52) | 5.13 (0.91) | NA | 69.58 (0.87) |
| Samples | SI | ||||
|---|---|---|---|---|---|
| MDA-MB-231 | 4T1 | T47D | MCF7 | ||
| 1 | AC n-hexane | 105.95 | 0.82 | 1.97 | 102.93 |
| 2 | IC n-hexane | NA | 4.31 | 0.74 | NA |
| 3 | IC ethylacetate | 223.69 | 7.47 | 1.16 | 213.68 |
| 4 | doxorubicin | NA | 0.18 | 13.56 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hariono, M.; Rollando, R.; Yoga, I.; Harjono, A.; Suryodanindro, A.; Yanuar, M.; Gonzaga, T.; Parabang, Z.; Hariyono, P.; Febriansah, R.; et al. Bioguided Fractionation of Local Plants against Matrix Metalloproteinase9 and Its Cytotoxicity against Breast Cancer Cell Models: In Silico and In Vitro Study (Part II). Molecules 2021, 26, 1464. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051464
Hariono M, Rollando R, Yoga I, Harjono A, Suryodanindro A, Yanuar M, Gonzaga T, Parabang Z, Hariyono P, Febriansah R, et al. Bioguided Fractionation of Local Plants against Matrix Metalloproteinase9 and Its Cytotoxicity against Breast Cancer Cell Models: In Silico and In Vitro Study (Part II). Molecules. 2021; 26(5):1464. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051464
Chicago/Turabian StyleHariono, Maywan, Rollando Rollando, I Yoga, Abraham Harjono, Alfonsus Suryodanindro, Michael Yanuar, Thomas Gonzaga, Zet Parabang, Pandu Hariyono, Rifki Febriansah, and et al. 2021. "Bioguided Fractionation of Local Plants against Matrix Metalloproteinase9 and Its Cytotoxicity against Breast Cancer Cell Models: In Silico and In Vitro Study (Part II)" Molecules 26, no. 5: 1464. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051464