
Spermidine Prevents Ethanol and Lipopolysaccharide-Induced Hepatic Injury in Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
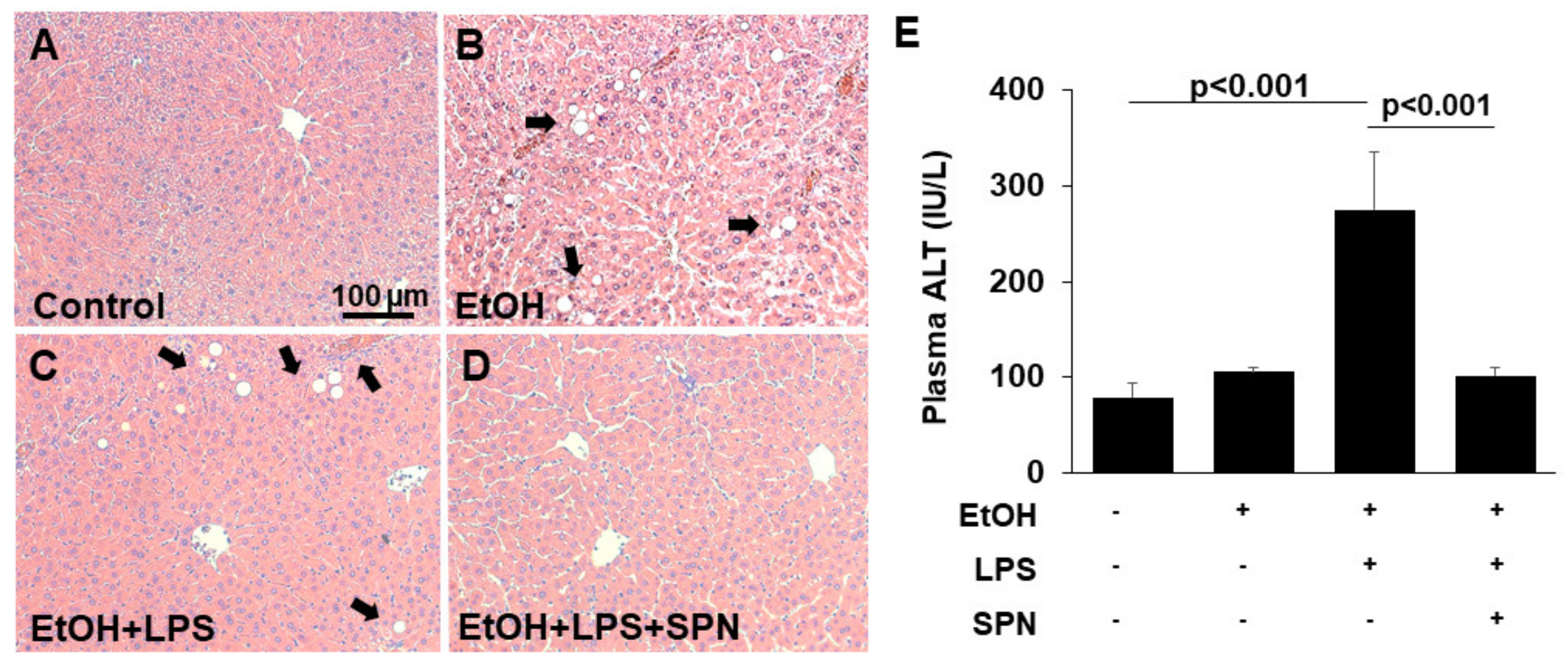
2.1. Spermidine Prevents Ethanol and LPS-induced Liver Injury
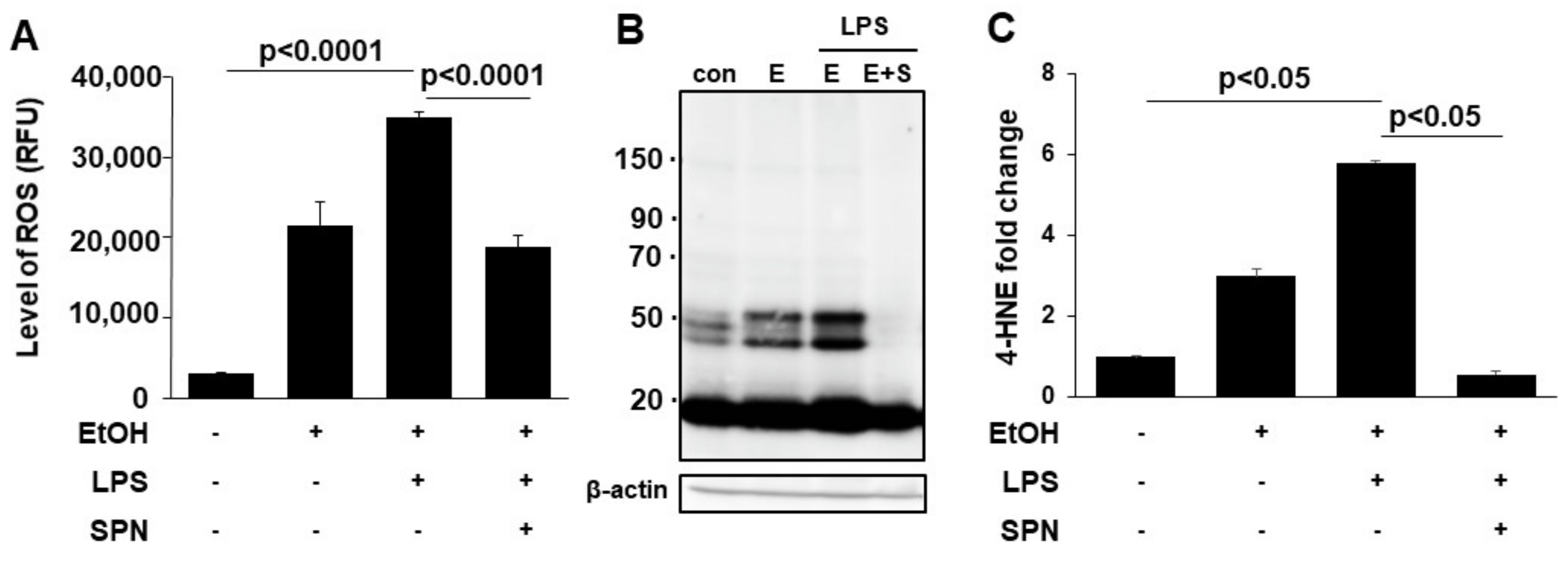
2.2. Spermidine Prevents Ethanol and LPS-induced Oxidative Stress and Lipid Peroxidation
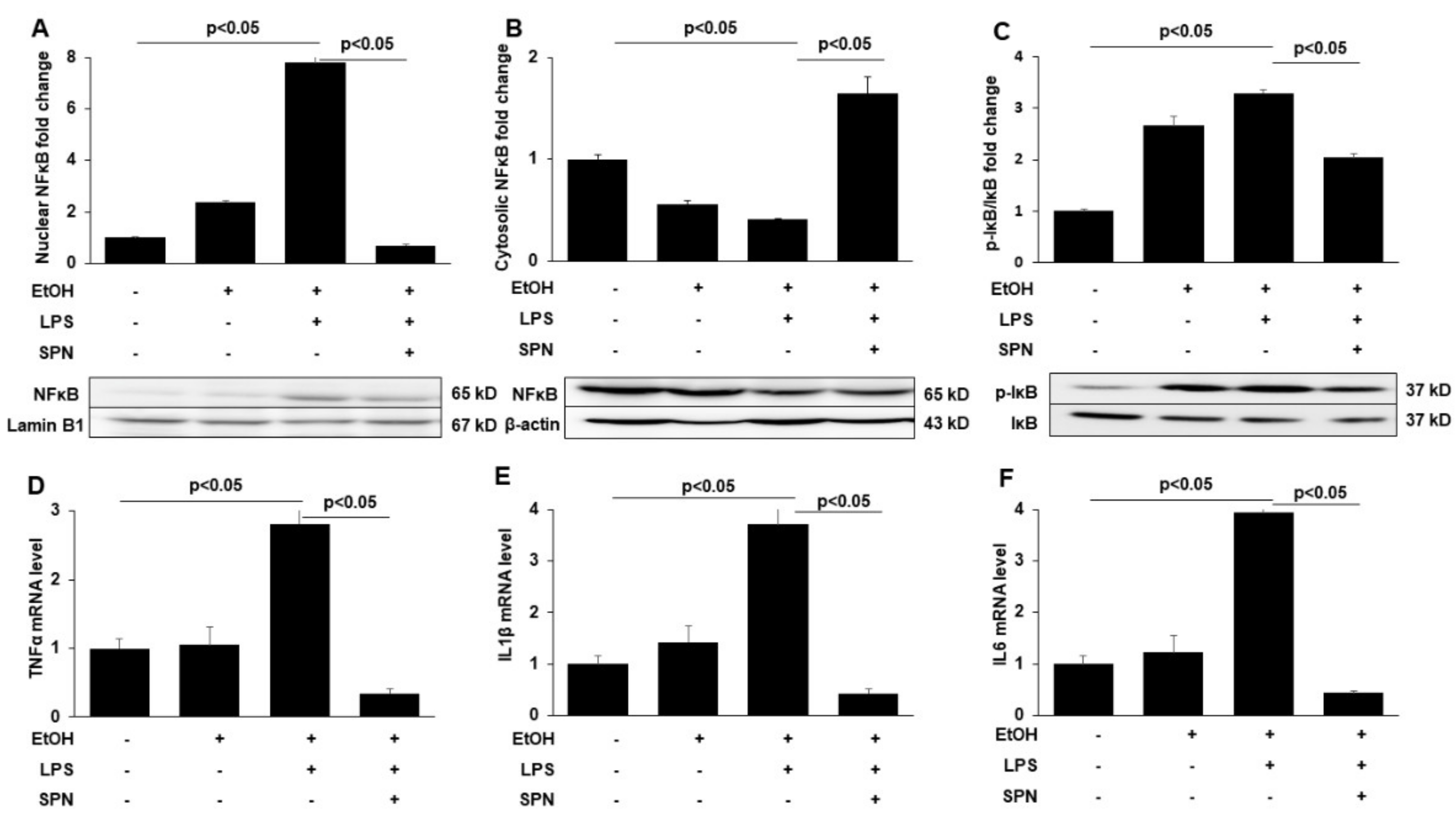
2.3. Spermidine Prevents Ethanol and LPS-induced Activation of Inflammatory Mediators
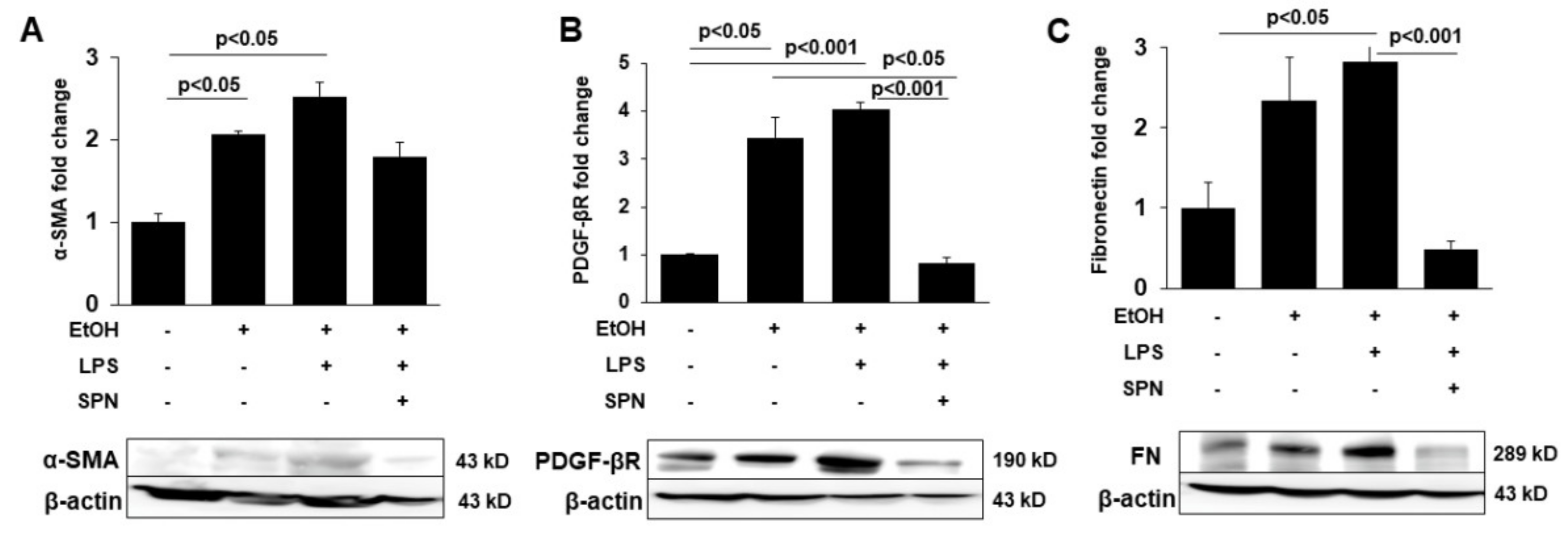
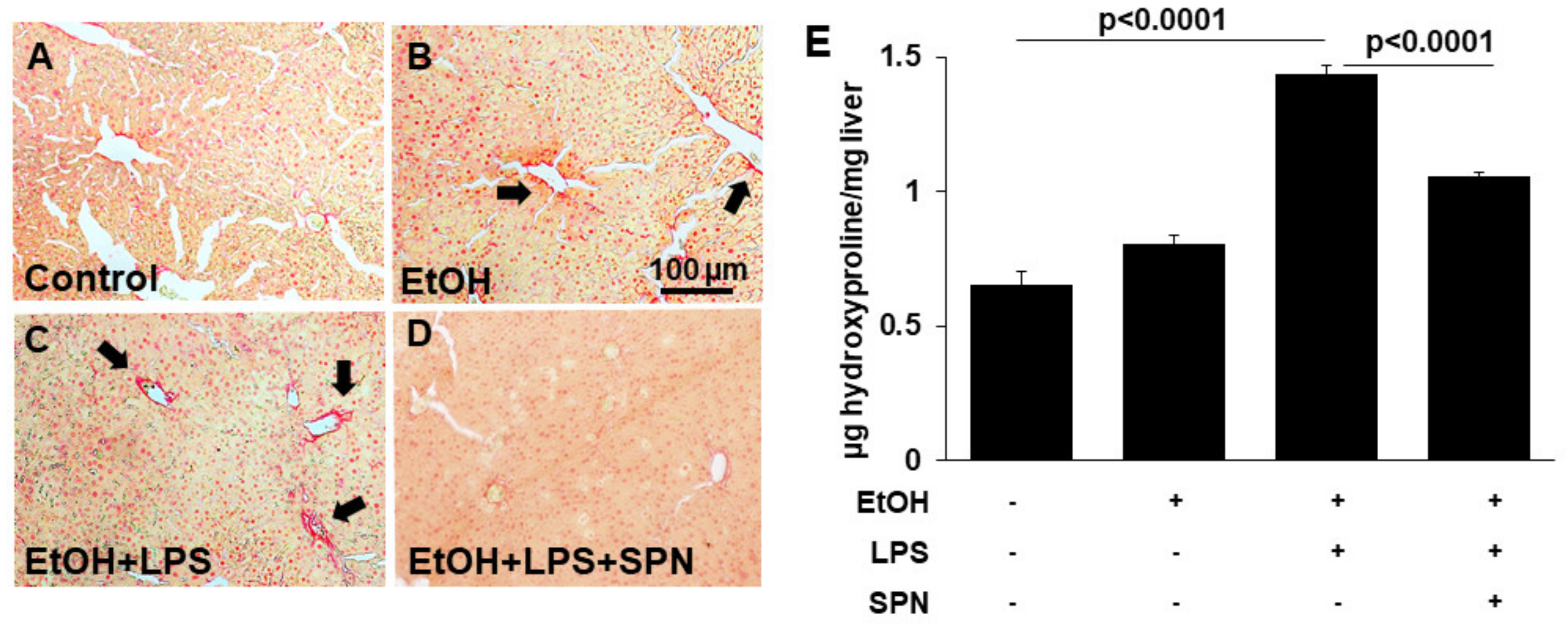
2.4. Spermidine Prevents Ethanol and LPS-induced Hepatic Fibrosis
3. Discussion
4. Materials and Methods
4.1. Mice and Diet
4.2. Plasma Liver Injury Markers
4.3. Hematoxylin and Eosin Staining
4.4. Reactive Oxygen Species and Hydroxyproline Determinations
4.5. Total, Nuclear and Cytosolic Protein Extraction
4.6. SDS-PAGE and Western Blot Analysis
4.7. RNA Extraction and Quantitative RT-PCR
4.8. Sirius Red Staining
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations:
| 4-HNE | 4-hydroxy-2-nonenal |
| DEN | diethylnitrosamine |
| EtOH | ethanol |
| IκB | inhibitor of nuclear factor kappa B |
| IL1β | interleukin 1 beta |
| IL6 | interleukin 6 |
| LPS | lipopolysaccharide |
| NFκB | nuclear factor kappa B |
| PDGF-βR | platelet-derived growth factor beta receptor |
| ROS | reactive oxygen species |
| αSMA | alpha smooth muscle actin |
| SPN | spermidine |
| TNFα | tumor necrosis factor alpha |
References
- Tuma, D.J.; Casey, C.A. Dangerous byproducts of alcohol breakdown—Focus on adducts. Alcohol. Res. Health 2003, 27, 285–290. [Google Scholar]
- Seitz, H.K.; Bataller, R.; Cortez-Pinto, H.; Gao, B.; Gual, A.; Lackner, C.; Mathurin, P.; Mueller, S.; Szabo, G.; Tsukamoto, H. Alcoholic liver disease. Nat. Rev. Dis. Primers 2018, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Rao, R. Endotoxemia and gut barrier dysfunction in alcoholic liver disease. Hepatology 2009, 50, 638–644. [Google Scholar] [CrossRef]
- Seki, E.; Schwabe, R.F. Hepatic inflammation and fibrosis: Functional links and key pathways. Hepatology 2015, 61, 1066–1079. [Google Scholar] [CrossRef] [PubMed]
- Bode, C.; Kugler, V.; Bode, J.C. Endotoxemia in patients with alcoholic and non-alcoholic cirrhosis and in subjects with no evidence of chronic liver disease following acute alcohol excess. J. Hepatol. 1987, 4, 8–14. [Google Scholar] [CrossRef]
- Nanji, A.A.; Khettry, U.; Sadrzadeh, S.M.; Yamanaka, T. Severity of liver injury in experimental alcoholic liver disease. Correlation with plasma endotoxin, prostaglandin E2, leukotriene B4, and thromboxane B2. Am. J. Pathol. 1993, 142, 367–373. [Google Scholar]
- Hernandez-Gea, V.; Friedman, S.L. Pathogenesis of liver fibrosis. Annu. Rev. Pathol. 2011, 6, 425–456. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef]
- Madeo, F.; Eisenberg, T.; Pietrocola, F.; Kroemer, G. Spermidine in health and disease. Science 2018, 359, eaan 2788. [Google Scholar] [CrossRef] [Green Version]
- Hoek, J. Endotoxin and alcoholic liver disease: Tolerance and susceptibility. Hepatology 1999, 29, 1602–1604. [Google Scholar] [CrossRef]
- Bertola, A.; Mathews, S.; Ki, S.H.; Wang, H.; Gao, B. Mouse model of chronic and binge ethanol feeding (the NIAAA model). Nat. Protoc. 2013, 8, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Sessa, A.; Perin, A. Ethanol and polyamine metabolism: Physiologic and pathologic implications: A review. Alcohol. Clin. Exp. Res. 1997, 21, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Atiya, A.M.; Poortvliet, E.; Strömberg, R.; Yngve, A. Polyamines in foods: Development of a food database. Food Nutr. Res. 2011, 55, 5572. [Google Scholar] [CrossRef] [Green Version]
- Milovic, V. Polyamines in the gut lumen: Bioavailability and biodistribution. Eur. J. Gastroenterol. Hepatol. 2001, 13, 1021–1025. [Google Scholar] [CrossRef]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Büttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L.; et al. Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Pietrocola, F. Caloric Restriction Mimetics Enhance Anticancer Immunosurveillance. Cancer Cell 2016, 30, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Okumura, S.; Teratani, T.; Fujimoto, Y.; Zhao, X.; Tsuruyama, T.; Masano, Y.; Kasahara, N.; Iida, T.; Yagi, S.; Uemura, T.; et al. Oral administration of polyamines ameliorates liver ischemia/reperfusion injury and promotes liver regeneration in rats. Liver Transpl. 2016, 22, 1231–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, F.; Li, W.; Zou, J.; Jiang, X.; Xu, G.; Huang, H.L.; Liu, L. Spermidine Prolongs Lifespan and Prevents Liver Fibrosis and Hepatocellular Carcinoma by Activating MAP1S-Mediated Autophagy. Cancer Res. 2017, 77, 2938–2951. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; de la Vega, M.R.; Dodson, M.; Yue, F.; Shi, B.; Fang, D.; Chapman, E.; Liu, L.; Zhang, D.D. Spermidine Confers Liver Protection by Enhancing NRF2 Signaling Through a MAP1S-Mediated Noncanonical Mechanism. Hepatology 2019, 70, 372–388. [Google Scholar] [CrossRef] [PubMed]
- Rigobello, M.P.; Toninello, A.; Siliprandi, D.; Bindoli, A. Effect of spermine on mitochondrial glutathione release. Biochem. Biophys. Res. Commun. 1993, 194, 1276–1281. [Google Scholar] [CrossRef] [PubMed]
- Elustondo, P.A.; Negoda, A.; Kane, C.L.; Kane, D.A.; Pavlov, E.V. Spermine selectively inhibits high-conductance, but not low-conductance calcium-induced permeability transition pore. Biochim. Biophys. Acta 2015, 1847, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ter Steege, J.C.; Forget, P.P.; Buurman, W.A. Oral spermine administration inhibits nitric oxide-mediated intestinal damage and levels of systemic inflammatory mediators in a mouse endotoxin model. Shock 1999, 11, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Lieber, C.S.; DeCarli, L.M. Liquid diet technique of ethanol administration: 1989 update. Alcohol Alcohol. 1989, 24, 197–211. [Google Scholar] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Gordillo, K.; Segovia, J.; Shibayama, M.; Vergara, P.; Moreno, M.G.; Muriel, P. Curcumin protects against acute liver damage in the rat by inhibiting NF-kappaB, proinflammatory cytokines production and oxidative stress. Biochim. Biophys. Acta 2007, 1770, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Garige, M.; Hirsch, K.; Lakshman, M.R. Liver Galbeta1,4GlcNAc alpha2,6-sialyltransferase is down-regulated in human alcoholics: Possible cause for the appearance of asialoconjugates. Metabolism 2007, 56, 1241–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhikari, R.; Shah, R.; Reyes-Gordillo, K.; Arellanes-Robledo, J.; Cheng, Y.; Ibrahim, J.; Tuma, P.L. Spermidine Prevents Ethanol and Lipopolysaccharide-Induced Hepatic Injury in Mice. Molecules 2021, 26, 1786. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26061786
Adhikari R, Shah R, Reyes-Gordillo K, Arellanes-Robledo J, Cheng Y, Ibrahim J, Tuma PL. Spermidine Prevents Ethanol and Lipopolysaccharide-Induced Hepatic Injury in Mice. Molecules. 2021; 26(6):1786. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26061786
Chicago/Turabian StyleAdhikari, Raghabendra, Ruchi Shah, Karina Reyes-Gordillo, Jaime Arellanes-Robledo, Ying Cheng, Joseph Ibrahim, and Pamela L. Tuma. 2021. "Spermidine Prevents Ethanol and Lipopolysaccharide-Induced Hepatic Injury in Mice" Molecules 26, no. 6: 1786. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26061786