
Enhancement of β-Glucan Biological Activity Using a Modified Acid-Base Extraction Method from Saccharomyces cerevisiae

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Extraction of β-Glucan
2.1.1. Acid-Base Extraction Method
2.1.2. Water Extraction Method
2.2. Characterization of β-Glucan
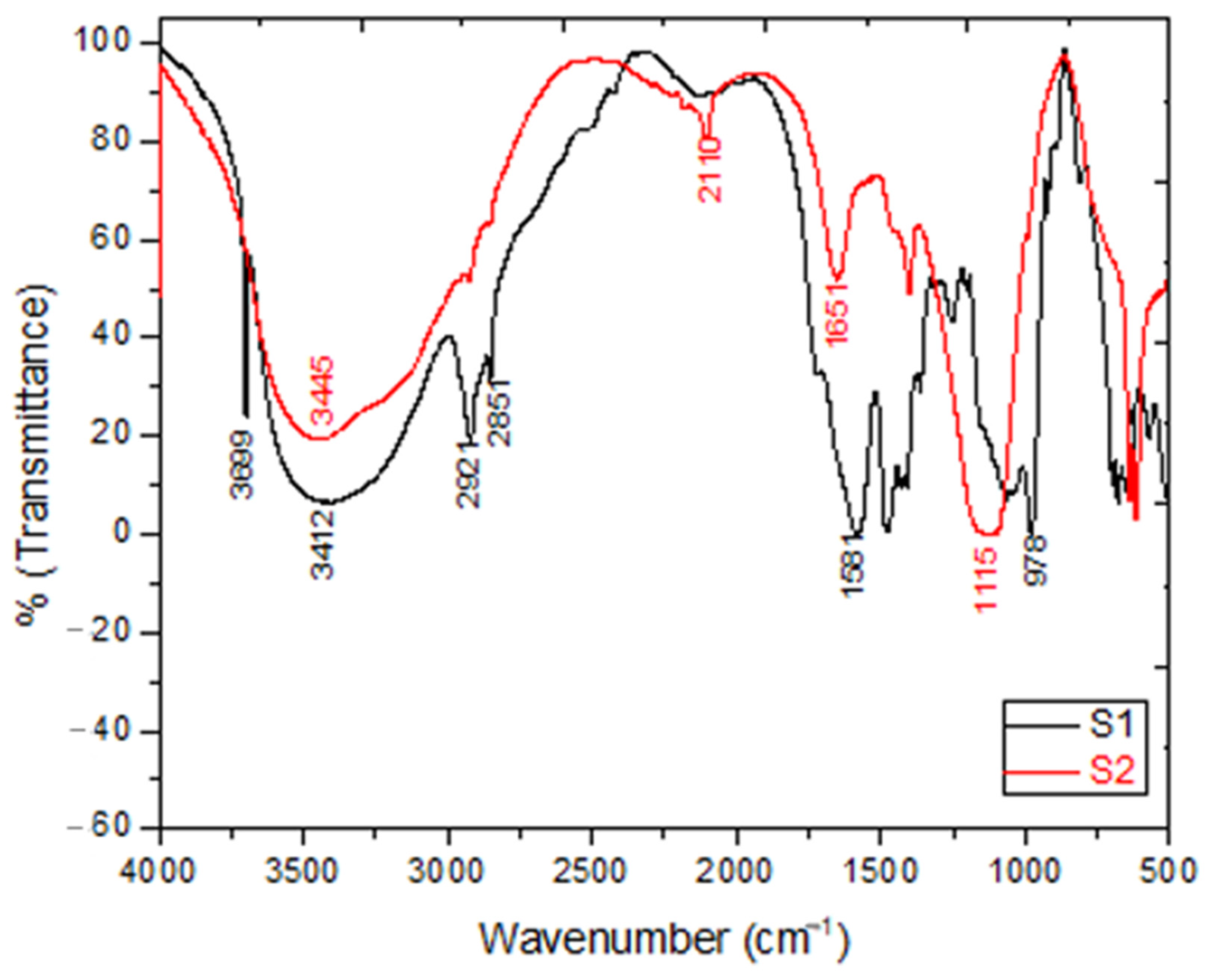
2.2.1. FTIR Spectroscopic Analysis
2.2.2. HPLC Analysis
2.2.3. Optical Properties
2.3. Detoxification Ability
2.4. Possible Mechanism for Interaction β-Glucan with AFs
2.5. Antimicrobial Activity of β-Glucan
2.6. β-Glucan as an Immune-Modulatory Molecule
2.7. Anticancer Activity of β-Glucan
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. β-Glucan Extraction Methods
4.3. β-Glucan Characterization
4.4. Aflatoxins Removal
4.5. Antimicrobial Activity
4.6. Immunomodulatory Effects
4.7. Anticancer Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Volman, J.J.; Ramakers, J.D.; Plat, J. Dietary Modulation of Immune Function by β-Glucans. Physiol. Behav. 2008, 94, 276–284. [Google Scholar] [CrossRef]
- Upadhyay, T.K.; Fatima, N.; Sharma, D.; Saravanakumar, V.; Sharma, R. Preparation and Characterization of Beta-Glucan Particles Containing a Payload of Nanoembedded Rifabutin for Enhanced Targeted Delivery to Macrophages. Excli J. 2017, 16, 210. [Google Scholar]
- Chamidah, A.; Hardoko; Prihanto, A.A. Antibacterial Activities of β-Glucan (Laminaran) against Gram-Negative and Gram-Positive Bacteria. In AIP Conference Proceedings; AIP Publishing LLC, 2017; Volume 1844, p. 20011. Available online: https://aip.scitation.org/doi/abs/10.1063/1.4983422# (accessed on 23 March 2021).
- Marasca, E.; Boulos, S.; Nyström, L. Bile Acid-Retention by Native and Modified Oat and Barley β-Glucan. Carbohydr. Polym. 2020, 236, 116034. [Google Scholar] [CrossRef] [PubMed]
- Muta, T. Molecular Basis for Invertebrate Innate Immune Recognition of (1→3)-β-D-Glucan as a Pathogen-Associated Molecular Pattern. Curr. Pharm. Des. 2006, 12, 4155–4161. [Google Scholar] [CrossRef]
- Gantner, B.N.; Simmons, R.M.; Canavera, S.J.; Akira, S.; Underhill, D.M. Collaborative Induction of Inflammatory Responses by Dectin-1 and Toll-like Receptor 2. J. Exp. Med. 2003, 197, 1107–1117. [Google Scholar] [CrossRef] [Green Version]
- Chan, G.C.-F.; Chan, W.K.; Sze, D.M.-Y. The Effects of β-Glucan on Human Immune and Cancer Cells. J. Hematol. Oncol. 2009, 2, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, C.; Cai, Y.; Gunn, L.; Ding, C.; Li, B.; Kloecker, G.; Qian, K.; Vasilakos, J.; Saijo, S.; Iwakura, Y. Differential Pathways Regulating Innate and Adaptive Antitumor Immune Responses by Particulate and Soluble Yeast-Derived β-Glucans. Blood J. Am. Soc. Hematol. 2011, 117, 6825–6836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yan, L.; Kim, J.A. Modulating Mammary Tumor Growth, Metastasis and Immunosuppression by SiRNA-Induced MIF Reduction in Tumor Microenvironment. Cancer Gene Ther. 2015, 22, 463–474. [Google Scholar] [CrossRef]
- Pelley, R.P.; Strickland, F.M. Plants, Polysaccharides, and the Treatment and Prevention of Neoplasia. Crit. Rev. Oncog. 2000, 11. [Google Scholar] [CrossRef]
- Camilli, G.; Tabouret, G.; Quintin, J. The Complexity of Fungal β-Glucan in Health and Disease: Effects on the Mononuclear Phagocyte System. Front. Immunol. 2018, 9, 673. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Kim, J.A.; Huang, A.Y.-C. Optimizing Tumor Microenvironment for Cancer Immunotherapy: β-Glucan-Based Nanoparticles. Front. Immunol. 2018, 9, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Würsch, P.; Pi-Sunyer, F.X. The Role of Viscous Soluble Fiber in the Metabolic Control of Diabetes: A Review with Special Emphasis on Cereals Rich in β-Glucan. Diabetes Care 1997, 20, 1774–1780. [Google Scholar] [CrossRef] [PubMed]
- Browder, W.; Williams, D.; Lucore, P.; Pretus, H.; Jones, E.; McNamee, R. Effect of Enhanced Macrophage Function on Early Wound Healing. Surgery 1988, 104, 224–230. [Google Scholar] [PubMed]
- Vetvicka, V.; Vetvickova, J. Β-Glucan Attenuates Chronic Fatique Syndrome in Murine Model. J. Nat. Sci. 2015, 1, e112. [Google Scholar]
- Braaten, J.T.; Wood, P.J.; Scott, F.W.; Wolynetz, M.S.; Lowe, M.K.; Bradley-White, P.; Collins, M.W. Oat Beta-Glucan Reduces Blood Cholesterol Concentration in Hypercholesterolemic Subjects. Eur. J. Clin. Nutr. 1994, 48, 465–474. [Google Scholar]
- Sima, P.; Vannucci, L.; Vetvicka, V. Glucans and Cancer: Historical Perspective. Cancer Transl. Med. 2015, 1, 209. [Google Scholar]
- Kushner, B.H.; Cheung, I.Y.; Modak, S.; Kramer, K.; Ragupathi, G.; Cheung, N.-K. V Phase I Trial of a Bivalent Gangliosides Vaccine in Combination with β-Glucan for High-Risk Neuroblastoma in Second or Later Remission. Clin. Cancer Res. 2014, 20, 1375–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, J.; Svozil, V.; Král, V.; Dobiášová, L.R.; Stiborová, I.; Vetvicka, V. Clinical Trials of Yeast-Derived β-(1, 3) Glucan in Children: Effects on Innate Immunity. Ann. Transl. Med. 2014, 2, 15. [Google Scholar] [PubMed]
- Vetvicka, V.; Richter, J.; Svozil, V.; Dobiášová, L.R.; Král, V. Placebo-Driven Clinical Trials of Yeast-Derived β-(1-3) Glucan in Children with Chronic Respiratory Problems. Ann. Transl. Med. 2013, 1, 26. [Google Scholar] [PubMed]
- LEGRAS, J.; Merdinoglu, D.; CORNUET, J.; Karst, F. Bread, Beer and Wine: Saccharomyces Cerevisiae Diversity Reflects Human History. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M.; Mol, P.; Hellingwerf, K.; Brul, S. Dynamics of Cell Wall Structure in Saccharomyces Cerevisiae. FEMS Microbiol. Rev. 2002, 26, 239–256. [Google Scholar] [CrossRef]
- Sánchez-Madrigal, M.Á.; Neder-Suárez, D.; Quintero-Ramos, A.; Ruiz-Gutiérrez, M.G.; Meléndez-Pizarro, C.O.; Piñón-Castillo, H.A.; Galicia-García, T.; Ramírez-Wong, B. Physicochemical Properties of Frozen Tortillas from Nixtamalized Maize Flours Enriched with β-Glucans. Food Sci. Technol. 2015, 35, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Stier, H.; Ebbeskotte, V.; Gruenwald, J. Immune-Modulatory Effects of Dietary Yeast Beta-1, 3/1, 6-D-Glucan. Nutr. J. 2014, 13, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, A.; Anjum, F.M.; Zahoor, T.; Nawaz, H.; Dilshad, S.M.R. Beta Glucan: A Valuable Functional Ingredient in Foods. Crit. Rev. Food Sci. Nutr. 2012, 52, 201–212. [Google Scholar] [CrossRef]
- Hromádková, Z.; Ebringerová, A.; Sasinková, V.; Šandula, J.; Hříbalová, V.; Omelková, J. Influence of the Drying Method on the Physical Properties and Immunomodulatory Activity of the Particulate (1→3)-β-D-Glucan from Saccharomyces Cerevisiae. Carbohydr. Polym. 2003, 51, 9–15. [Google Scholar] [CrossRef]
- Allan, S.E.; Crome, S.Q.; Crellin, N.K.; Passerini, L.; Steiner, T.S.; Bacchetta, R.; Roncarolo, M.G.; Levings, M.K. Activation-Induced FOXP3 in Human T Effector Cells Does Not Suppress Proliferation or Cytokine Production. Int. Immunol. 2007, 19, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenger, M.D.; DePhillips, P.; Bracewell, D.G. A Microscale Yeast Cell Disruption Technique for Integrated Process Development Strategies. Biotechnol. Prog. 2008, 24, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, P. Revisiting the Cellulosimicrobium Cellulans Yeast-Lytic β-1, 3-Glucanases Toolbox: A Review. Microb. Cell Fact. 2006, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pengkumsri, N.; Sivamaruthi, B.S.; Sirilun, S.; Peerajan, S.; Kesika, P.; Chaiyasut, K.; Chaiyasut, C. Extraction of β-Glucan from Saccharomyces Cerevisiae: Comparison of Different Extraction Methods and in Vivo Assessment of Immunomodulatory Effect in Mice. Food Sci. Technol. 2017, 37, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Babayan, T.L.; Bezrukov, M.G. Autolysis in Yeasts. Acta Biotechnol. 1985, 5, 129–136. [Google Scholar] [CrossRef]
- Babayan, T.L.; Bezrukov, M.G.; Latov, V.K.; Belikov, V.M.; Belavtseva, E.M.; Titova, E.F. Induced Autolysis of Saccharomyces Cerevisiae: Morphological Effects, Rheological Effects, and Dynamics of Accumulation of Extracellular Hydrolysis Products. Curr. Microbiol. 1981, 5, 163–168. [Google Scholar] [CrossRef]
- Charpentier, C.; Freyssinet, M. The Mechanism of Yeast Autolysis in Wine. Yeast (Chichester) 1989, 5, 181–186. [Google Scholar]
- Zohri, A.N.A.; Moubasher, H.; Abdel-Hay, H.M.; Orban, M.A.I. Biotechnological β-Glucan Production from Returned Bakers Yeast and Yeast Remaining after Ethanol Fermentation. Egypt. Sugar J. 2019, 4, 5. [Google Scholar]
- Bzducha-Wróbel, A.; Błażejak, S.; Kawarska, A.; Stasiak-Różańska, L.; Gientka, I.; Majewska, E. Evaluation of the Efficiency of Different Disruption Methods on Yeast Cell Wall Preparation for β-Glucan Isolation. Molecules 2014, 19, 20941–20961. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zeng, X.-A.; Sun, D.-W.; Han, Z. Disruption and Protein Release by Ultrasonication of Yeast Cells. Innov. Food Sci. Emerg. Technol. 2013, 18, 132–137. [Google Scholar] [CrossRef]
- Pattanayak, S.; Chakraborty, S.; Biswas, S.; Chattopadhyay, D.; Chakraborty, M. Degradation of Methyl Parathion, a Common Pesticide and Fluorescence Quenching of Rhodamine B, a Carcinogen Using β-d Glucan Stabilized Gold Nanoparticles. J. Saudi Chem. Soc. 2018, 22, 937–948. [Google Scholar] [CrossRef]
- Reddy, K.R.N.; Salleh, B.; Saad, B.; Abbas, H.K.; Abel, C.A.; Shier, W.T. Erratum: An Overview of Mycotoxin Contamination in Foods and Its Implications for Human Health (Toxin Reviews (2010) 29 (326)). Toxin Rev. 2010, 29. [Google Scholar] [CrossRef]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human Aflatoxicosis in Developing Countries: A Review of Toxicology, Exposure, Potential Health Consequences, and Interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Santillán, E.; Morales-González, J.A.; Vargas-Mendoza, N.; Reyes-Ramírez, P.; Cruz-Jaime, S.; Sumaya-Martínez, T.; Pérez-Pastén, R.; Madrigal-Bujaidar, E. Antigenotoxic Studies of Different Substances to Reduce the DNA Damage Induced by Aflatoxin B1 and Ochratoxin A. Toxins 2010, 2, 738–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleveland, T.E.; Dowd, P.F.; Desjardins, A.E.; Bhatnagar, D.; Cotty, P.J. United States Department of Agriculture—Agricultural Research Service Research on Pre-harvest Prevention of Mycotoxins and Mycotoxigenic Fungi in US Crops. Pest Manag. Sci. Former. Pestic. Sci. 2003, 59, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Anttila, A.; Bhat, R.V.; Bond, J.A.; Borghoff, S.J.; Bosch, F.X.; Carlson, G.P.; Castegnaro, M.; Cruzan, G.; Gelderblom, W.C.A.; Hass, U. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans: Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene. IARC Monogr. Eval. Carcinog. Risks Hum. 2002, 82. Available online: https://0-ucdavis-pure-elsevier-com.brum.beds.ac.uk/en/publications/iarc-monographs-on-the-evaluation-of-carcinogenic-risks-to-humans (accessed on 1 April 2021).
- Baptista, A.S.; Horii, J.; Calori-Domingues, M.A.; Da Glória, E.M.; Salgado, J.M.; Vizioli, M.R. The Capacity of Manno-Oligosaccharides, Thermolysed Yeast and Active Yeast to Attenuate Aflatoxicosis. World J. Microbiol. Biotechnol. 2004, 20, 475–481. [Google Scholar] [CrossRef]
- Bianco, G.; Russo, R.; Marzocco, S.; Velotto, S.; Autore, G.; Severino, L. Modulation of Macrophage Activity by Aflatoxins B1 and B2 and Their Metabolites Aflatoxins M1 and M2. Toxicon 2012, 59, 644–650. [Google Scholar] [CrossRef]
- Roda, E.; Coccini, T.; Acerbi, D.; Castoldi, A.F.; Manzo, L. Comparative in Vitro and Ex-Vivo Myelotoxicity of Aflatoxins B1 and M1 on Haematopoietic Progenitors (BFU-E, CFU-E, and CFU-GM): Species-Related Susceptibility. Toxicol. In Vitro 2010, 24, 217–223. [Google Scholar] [CrossRef]
- Shetty, P.H.; Jespersen, L. Saccharomyces Cerevisiae and Lactic Acid Bacteria as Potential Mycotoxin Decontaminating Agents. Trends Food Sci. Technol. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic Acid Bacteria—Potential for Control of Mould Growth and Mycotoxins: A Review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Piotrowska, M. Adsorption of Ochratoxin A by Saccharomyces Cerevisiae Living and Non-Living Cells. Acta Aliment. 2012, 41, 1–7. [Google Scholar] [CrossRef]
- Piotrowska, M.; Nowak, A.; Czyzowska, A. Removal of Ochratoxin A by Wine Saccharomyces Cerevisiae Strains. Eur. Food Res. Technol. 2013, 236, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Petruzzi, L.; Bevilacqua, A.; Baiano, A.; Beneduce, L.; Corbo, M.R.; Sinigaglia, M. In Vitro Removal of Ochratoxin A by Two Strains of S Accharomyces Cerevisiae and Their Performances under Fermentative and Stressing Conditions. J. Appl. Microbiol. 2014, 116, 60–70. [Google Scholar] [CrossRef]
- Zhao, L.; Jin, H.; Lan, J.; Zhang, R.; Ren, H.; Zhang, X.; Yu, G. Detoxification of Zearalenone by Three Strains of Lactobacillus Plantarum from Fermented Food in Vitro. Food Control 2015, 54, 158–164. [Google Scholar] [CrossRef]
- Raju, M.; Devegowda, G. Influence of Esterified-Glucomannan on Performance and Organ Morphology, Serum Biochemistry and Haematology in Broilers Exposed to Individual and Combined Mycotoxicosis (Aflatoxin, Ochratoxin and T-2 Toxin). Br. Poult. Sci. 2000, 41, 640–650. [Google Scholar] [CrossRef]
- Yiannikouris, A.; FRANCois, J.; Poughon, L.; Dussap, C.-G.; Bertin, G.; Jeminet, G.; Jouany, J.-P. Adsorption of Zearalenone by β-D-Glucans in the Saccharomyces Cerevisiae Cell Wall. J. Food Prot. 2004, 67, 1195–1200. [Google Scholar] [CrossRef]
- Yiannikouris, A.; André, G.; Poughon, L.; François, J.; Dussap, C.-G.; Jeminet, G.; Bertin, G.; Jouany, J.-P. Chemical and Conformational Study of the Interactions Involved in Mycotoxin Complexation with β-D-Glucans. Biomacromolecules 2006, 7, 1147–1155. [Google Scholar] [CrossRef]
- Pereyra, C.M.; Cavaglieri, L.R.; Chiacchiera, S.M.; Dalcero, A. The Corn Influence on the Adsorption Levels of Aflatoxin B 1 and Zearalenone by Yeast Cell Wall. J. Appl. Microbiol. 2013, 114, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Bujaidar, E.; Morales-González, J.A.; Sánchez-Gutiérrez, M.; Izquierdo-Vega, J.A.; Reyes-Arellano, A.; Álvarez-González, I.; Pérez-Pasten, R.; Madrigal-Santillán, E. Prevention of Aflatoxin B1-Induced Dna Breaks by β-D-Glucan. Toxins 2015, 7, 2145–2158. [Google Scholar] [CrossRef] [Green Version]
- Aazami, M.H.; Nasri, M.H.F.; Mojtahedi, M.; Mohammadi, S.R. In Vitro Aflatoxin B1 Binding by the Cell Wall and (1→3)-β-d-Glucan of Baker’s Yeast. J. Food Prot. 2018, 81, 670–676. [Google Scholar] [CrossRef]
- Akhapkina, I.G.; Antropova, A.B.; Akhmatov, E.A.; Zheltikova, T.M. Effects of the Linear Fragments of Beta-(1→3)-Glucans on Cytokine Production in Vitro. Biochemistry 2018, 83, 1002–1006. [Google Scholar] [CrossRef] [PubMed]
- Ginovyan, M.; Keryan, A.; Bazukyan, I.; Ghazaryan, P.; Trchounian, A. The Large Scale Antibacterial, Antifungal and Anti-Phage Efficiency of Petamcin-A: New Multicomponent Preparation for Skin Diseases Treatment. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irshad, A.; Sarwar, N.; Sadia, H.; Malik, K.; Javed, I.; Irshad, A.; Afzal, M.; Abbas, M.; Rizvi, H. Comprehensive Facts on Dynamic Antimicrobial Properties of Polysaccharides and Biomolecules-Silver Nanoparticle Conjugate. Int. J. Biol. Macromol. 2020, 145, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Gao, Y.; Ding, Z.; Zhao, Y.; Yang, Y.; Sun, Q.; Yang, X.; Ge, W.; Xu, X.; Cheng, R. Soluble Beta-Glucan Salecan Improves Vaginal Infection of Candida Albicans in Mice. Int. J. Biol. Macromol. 2020, 148, 1053–1060. [Google Scholar] [CrossRef]
- Sudjaswadi, R. Increasing of the Bacteriostatic Effects of HCl Tetracycline–Polyethylene Glycol 6000–Tween 80 (PT). Indones. J. Pharm. 2006, 17, 98–103. [Google Scholar]
- Javmen, A.; NemeikaitĖ-Čėnienė, A.; Bratchikov, M.; Grigiškis, S.; Grigas, F.; Jonauskienė, I.; Zabulytė, D.; Mauricas, M. β-Glucan from Saccharomyces Cerevisiae Induces IFN-γ Production in Vivo in BALB/c Mice. In Vivo 2015, 29, 359–363. [Google Scholar] [PubMed]
- Liang, J.; Melican, D.; Cafro, L.; Palace, G.; Fisette, L.; Armstrong, R.; Patchen, M.L. Enhanced Clearance of a Multiple Antibiotic Resistant Staphylococcus Aureus in Rats Treated with PGG-Glucan Is Associated with Increased Leukocyte Counts and Increased Neutrophil Oxidative Burst Activity. Int. J. Immunopharmacol. 1998, 20, 595–614. [Google Scholar] [CrossRef]
- Soltys, J.; Quinn, M.T. Modulation of Endotoxin-and Enterotoxin-Induced Cytokine Release by in Vivo Treatment with β-(1, 6)-Branched β-(1, 3)-Glucan. Infect. Immun. 1999, 67, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.L.; Browder, I.W.; Di Luzio, N.R. Immunotherapeutic Modification of Escherichia Coli-Induced Experimental Peritonitis and Bacteremia by Glucan. Surgery 1983, 93, 448–454. [Google Scholar]
- Markova, N.; Kussovski, V.; Drandarska, I.; Nikolaeva, S.; Georgieva, N.; Radoucheva, T. Protective Activity of Lentinan in Experimental Tuberculosis. Int. Immunopharmacol. 2003, 3, 1557–1562. [Google Scholar] [CrossRef]
- Korotchenko, E.; Schießl, V.; Scheiblhofer, S.; Schubert, M.; Dall, E.; Joubert, I.A.; Strandt, H.; Neuper, T.; Sarajlic, M.; Bauer, R. Laser-facilitated Epicutaneous Immunotherapy with Hypoallergenic Beta-glucan Neoglycoconjugates Suppresses Lung Inflammation and Avoids Local Side Effects in a Mouse Model of Allergic Asthma. Allergy 2021, 76, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.Ø.; Lagos, L.; Lei, P.; Reveco-Urzua, F.E.; Morales-Lange, B.; Hansen, L.D.; Schiavone, M.; Mydland, L.T.; Arntzen, M.Ø.; Mercado, L. Down-Stream Processing of Baker’s Yeast (Saccharomyces Cerevisiae)—Effect on Nutrient Digestibility and Immune Response in Atlantic Salmon (Salmo Salar). Aquaculture 2021, 530, 735707. [Google Scholar] [CrossRef]
- Ina, K.; Kataoka, T.; Ando, T. The Use of Lentinan for Treating Gastric Cancer. Anti-Cancer Agents Med. Chem. (Formerly Curr. Med. Chem. Agents) 2013, 13, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Demir, G.; Klein, H.O.; Mandel-Molinas, N.; Tuzuner, N. Beta Glucan Induces Proliferation and Activation of Monocytes in Peripheral Blood of Patients with Advanced Breast Cancer. Int. Immunopharmacol. 2007, 7, 113–116. [Google Scholar] [CrossRef]
- Shomori, K.; Yamamoto, M.; Arifuku, I.; Teramachi, K.; Ito, H. Antitumor Effects of a Water-Soluble Extract from Maitake (Grifola Frondosa) on Human Gastric Cancer Cell Lines. Oncol. Rep. 2009, 22, 615–620. [Google Scholar] [CrossRef] [Green Version]
- Vetvicka, V.; Vetvickova, J. Glucans and Cancer: Comparison of Commercially Available β-Glucans—Part IV. Anticancer Res. 2018, 38, 1327–1333. [Google Scholar]
- Sambrani, R.; Abdolalizadeh, J.; Kohan, L.; Jafari, B. Recent Advances in the Application of Probiotic Yeasts, Particularly Saccharomyces, as an Adjuvant Therapy in the Management of Cancer with Focus on Colorectal Cancer. Mol. Biol. Rep. 2021, 48, 915–960. [Google Scholar]
- Vetvicka, V.; Vetvickova, J. Anti-Infectious and Anti-Tumor Activities of β-Glucans. Anticancer Res. 2020, 40, 3139–3145. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Hong, S.-Y.; Kim, S.-K.; Cheong, C.; Park, H.-J.; Chun, H.-K.; Jang, K.-H.; Yoon, B.-D.; Kim, C.-H.; Kang, S.A. β-Glucan Enhanced Apoptosis in Human Colon Cancer Cells SNU-C4. Nutr. Res. Pract. 2009, 3, 180–184. [Google Scholar] [CrossRef]
- Piotrowska, M.; Masek, A. Saccharomyces Cerevisiae Cell Wall Components as Tools for Ochratoxin A Decontamination. Toxins 2015, 7, 1151–1162. [Google Scholar] [CrossRef] [Green Version]
- Freimund, S.; Sauter, M.; Käppeli, O.; Dutler, H. A New Non-Degrading Isolation Process for 1, 3-β-D-Glucan of High Purity from Baker’s Yeast Saccharomyces Cerevisiae. Carbohydr. Polym. 2003, 54, 159–171. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Wang, Q.; Cui, S.W.; Liu, H.-Z. A New Isolation Method of β-D-Glucans from Spent Yeast Saccharomyces Cerevisiae. Food Hydrocoll. 2008, 22, 239–247. [Google Scholar] [CrossRef]
- Kesika, P.; Prasanth, M.I.; Balamurugan, K. Modulation of Caenorhabditis Elegans Immune Response and Modification of Shigella Endotoxin upon Interaction. J. Basic Microbiol. 2015, 55, 432–450. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Prasanth, M.I.; Balamurugan, K. Alterations in Caenorhabditis Elegans and Cronobacter Sakazakii Lipopolysaccharide during Interaction. Arch. Microbiol. 2015, 197, 327–337. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Zaki, S.A.; Abd-Elrahman, M.I.; Abu-Sehly, A.A.; Shaalan, N.M.; Hafiz, M.M. Thermal Annealing of SnS Thin Film Induced Mixed Tin Sulfide Oxides-Sn2S3 for Gas Sensing: Optical and Electrical Properties. Mater. Sci. Semicond. Process. 2018, 75, 214–220. [Google Scholar] [CrossRef]
- Zohri, A.N.; Aboul-Nasr, M.B.; Adam, M.; Mustafa, M.A.; Amer, E.M. Impact of Enzymes and Toxins Potentiality of Four Aspergillus Species to Cause Aspergillosis. Biol. Med. 2017, 9, 2. [Google Scholar]
- Farhan, M.A.; Moharram, A.M.; Salah, T.; Shaaban, O.M. Types of Yeasts That Cause Vulvovaginal Candidiasis in Chronic Users of Corticosteroids. Med. Mycol. 2019, 57, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Compounds | Test Strains/ZOI (mm) and MIC (mg/mL) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. aureus | MRSA | S. pneumoniae | E. coli | K. pneumoniae | P. aeruginosa | A. flavus | A. niger | C. albicans | ||||||||||
| ZOI | MIC | ZOI | MIC | ZOI | MIC | ZOI | MIC | ZOI | MIC | ZOI | MIC | ZOI | MIC | ZOI | MIC | ZOI | MIC | |
| S1 | 11.9 ± 0.3 | 0.39 | 11 ± 0.3 | 0.39 | 5.1 ± 0.08 | 3.13 | 2.2 ± 0.1 | 6.25 | 10.2 ± 0.1 | 0.78 | 13.2 ± 0.1 | 0.19 | 2.6 ± 0.1 | 6.25 | 6.1 ± 0.07 | 3.12 | 7.7 ± 0.1 | 3.12 |
| S2 | 9.2 ± 0.1 | 0.78 | 8.2 ± 0.2 | 1.56 | 1.6 ± 0.1 | 6.25 | R | R | 5.7 ± 0.2 | 3.125 | 11.1 ± 0.1 | 0.39 | R | R | R | R | 6.9 ± 0.4 | 3.12 |
| PC | 12 ± 0.3 | 0.78 | 11.3 ± 0.4 | 0.78 | 8.6 ± 0.1 | 0.31 | 16.9 ± 0.8 | 0.15 | 9.5 ± 0.3 | 0.31 | 17.2 ± 0.2 | 0.15 | 2.7 ± 0.1 | 0.78 | 5.1 ± 0.2 | 0.39 | 8.0 ± 0.3 | 0.39 |
| DMSO | R | R | R | R | R | R | R | R | R | 1.2 ± 0.05 | R | 4.2 ± 0.1 | R | 2.0 ± 0.04 | R | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoud Amer, E.; Saber, S.H.; Abo Markeb, A.; Elkhawaga, A.A.; Mekhemer, I.M.A.; Zohri, A.-N.A.; Abujamel, T.S.; Harakeh, S.; Abd-Allah, E.A. Enhancement of β-Glucan Biological Activity Using a Modified Acid-Base Extraction Method from Saccharomyces cerevisiae. Molecules 2021, 26, 2113. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082113
Mahmoud Amer E, Saber SH, Abo Markeb A, Elkhawaga AA, Mekhemer IMA, Zohri A-NA, Abujamel TS, Harakeh S, Abd-Allah EA. Enhancement of β-Glucan Biological Activity Using a Modified Acid-Base Extraction Method from Saccharomyces cerevisiae. Molecules. 2021; 26(8):2113. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082113
Chicago/Turabian StyleMahmoud Amer, Enas, Saber H. Saber, Ahmad Abo Markeb, Amal A. Elkhawaga, Islam M. A. Mekhemer, Abdel-Naser A. Zohri, Turki S. Abujamel, Steve Harakeh, and Elham A. Abd-Allah. 2021. "Enhancement of β-Glucan Biological Activity Using a Modified Acid-Base Extraction Method from Saccharomyces cerevisiae" Molecules 26, no. 8: 2113. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082113