
Thermodynamic Evaluation of the Interactions between Anticancer Pt(II) Complexes and Model Proteins

 ,
,  , , , ,
, , , ,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Stability over Time of Metal Complex in Solution
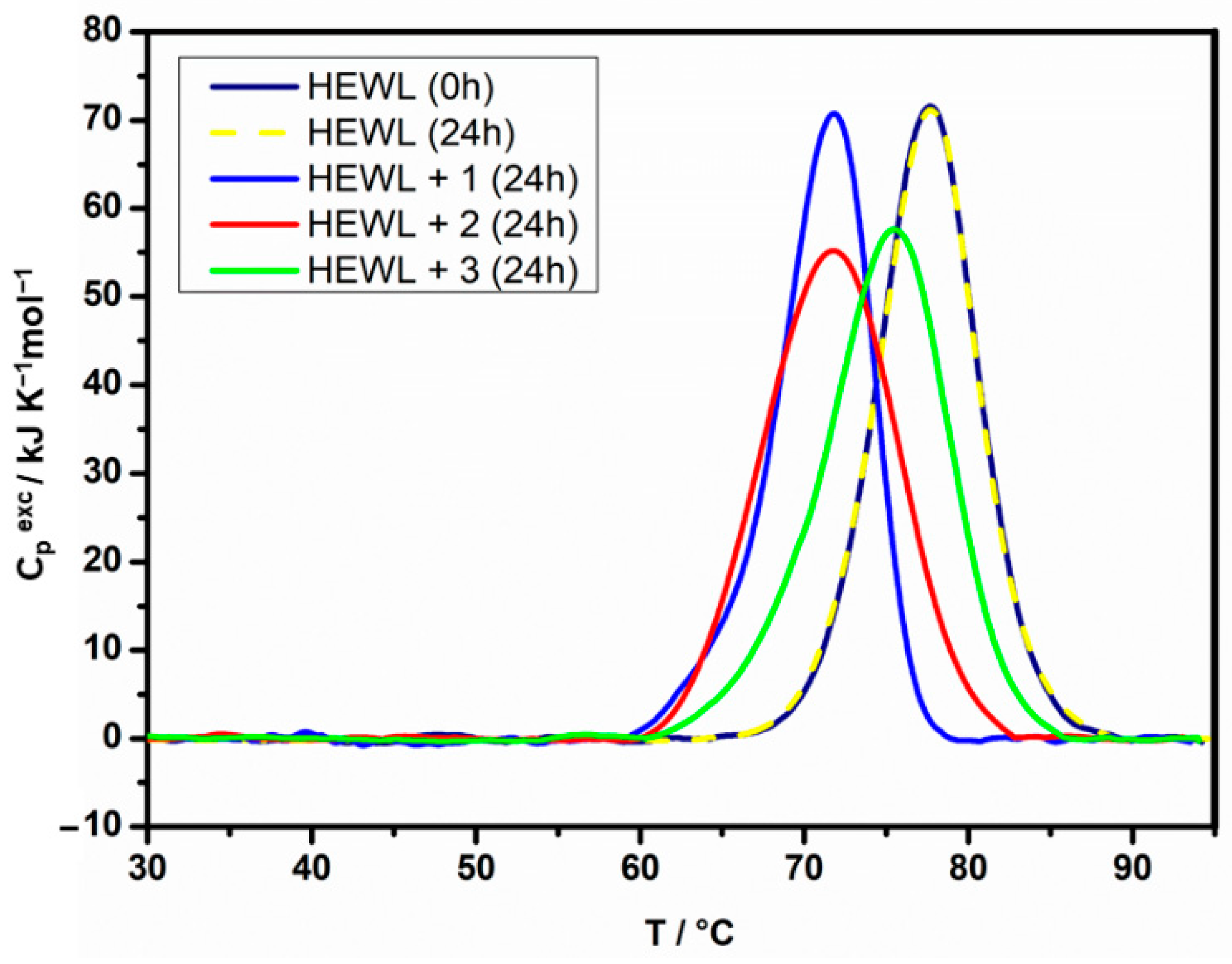
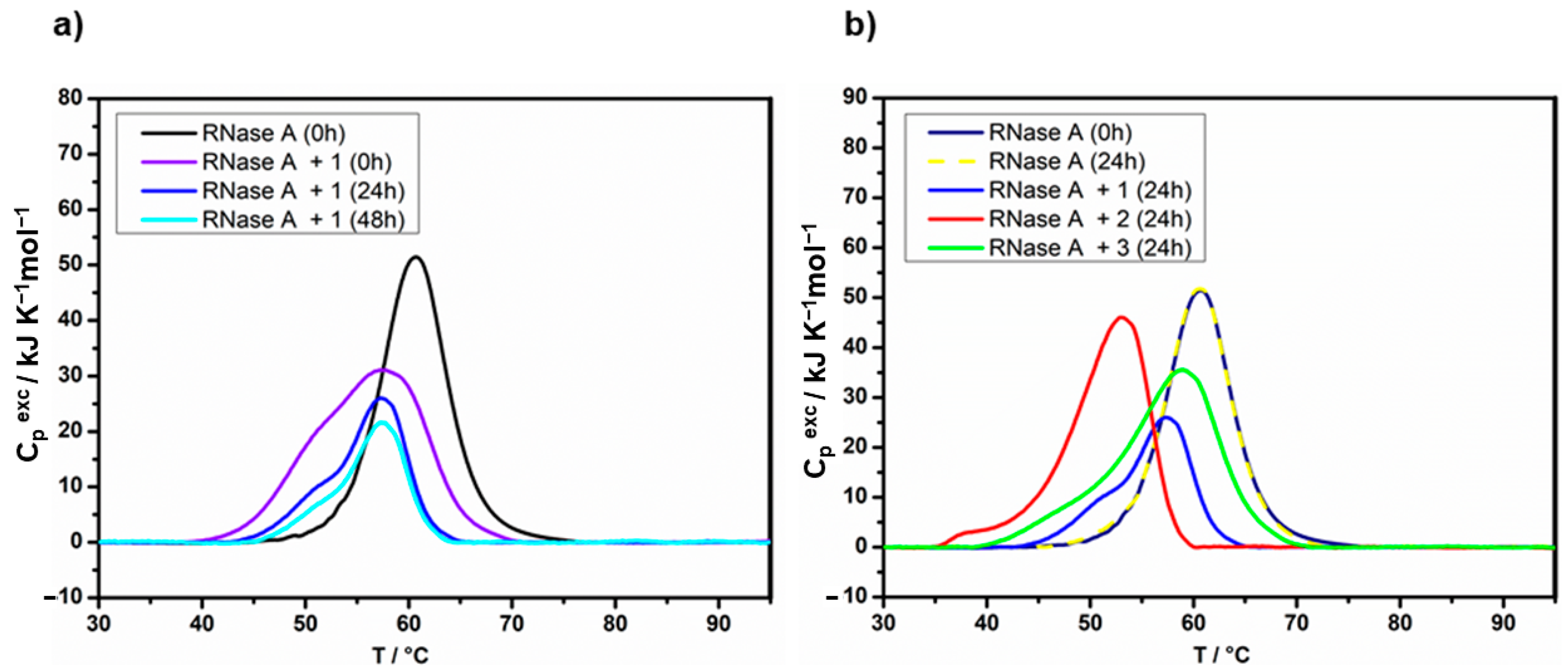
2.2. Protein Binding by Differential Scanning Calorimetry
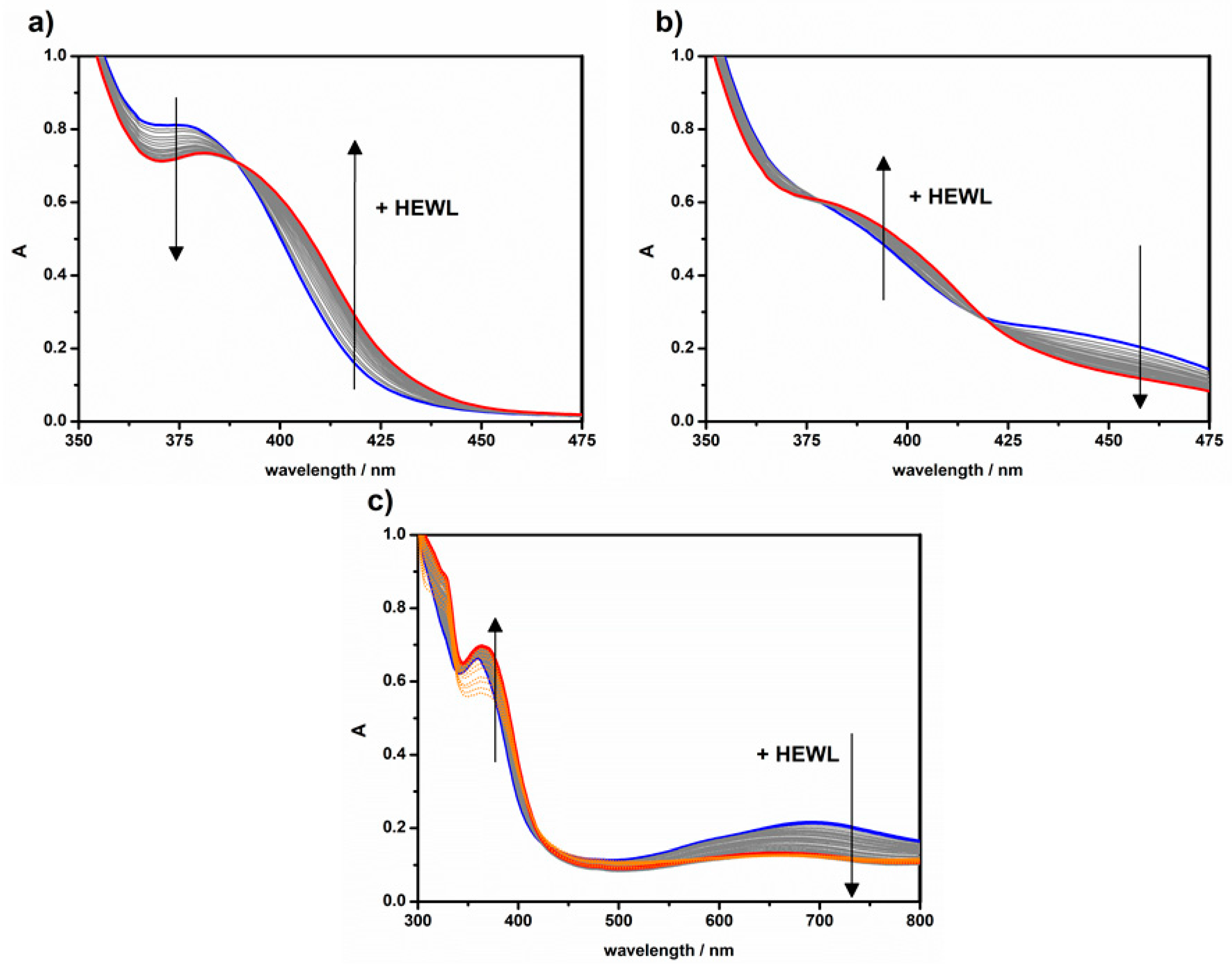
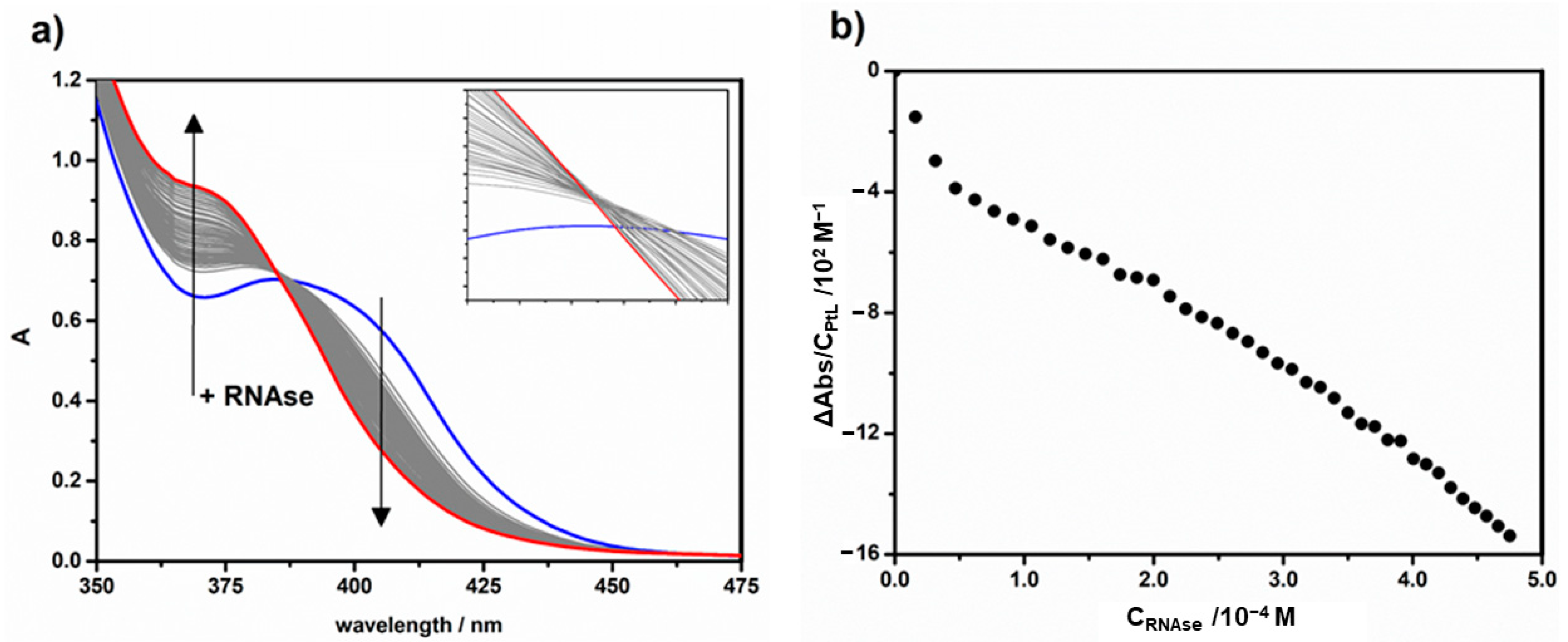
2.3. Protein Binding by UV-Vis Titrations
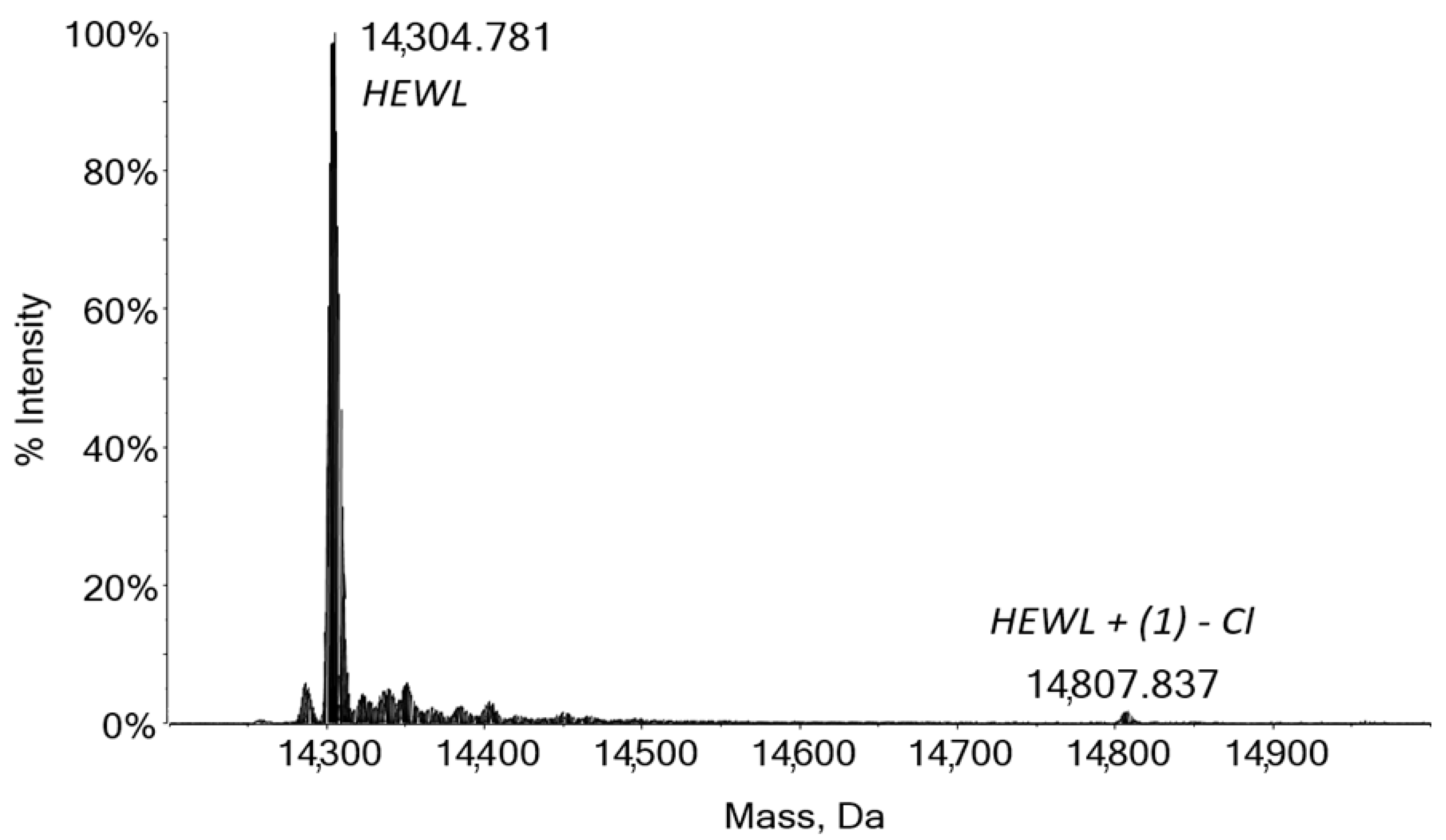
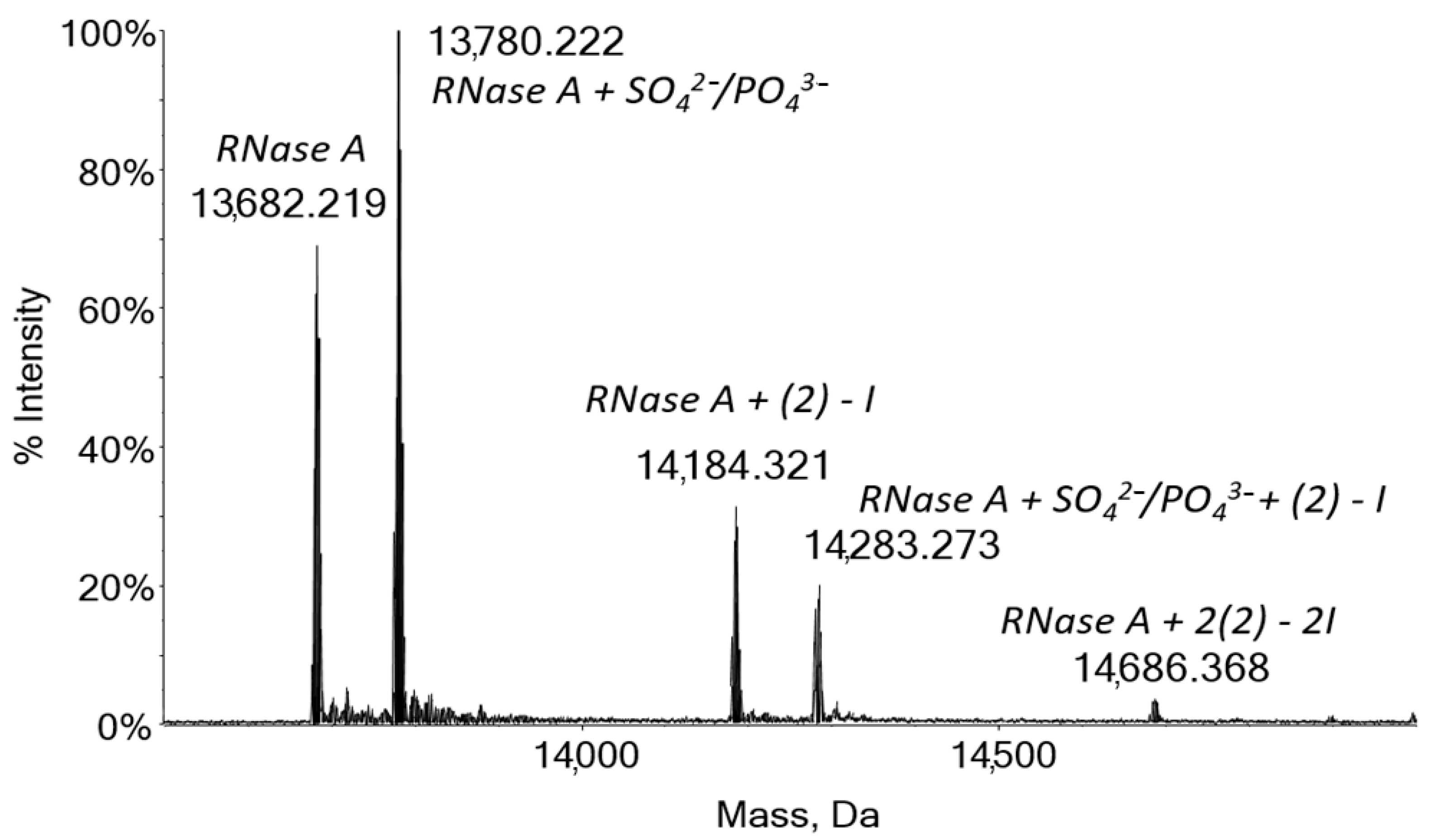
2.4. Protein Binding by Mass Spectrometry
3. Materials and Methods
3.1. Materials
3.2. Differential Scanning Calorimetry
3.3. UV-Vis Spectrophotometry
3.4. Mass Spectrometry
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Muggia, F.M.; Bonetti, A.; Hoeschele, J.D.; Rozencweig, M.; Howell, S.B. Platinum Antitumor Complexes: 50 Years Since Barnett Rosenberg’s Discovery. J. Clin. Oncol. 2015, 33, 4219–4226. [Google Scholar] [CrossRef] [PubMed]
- Alderden, R.A.; Hall, M.D.; Hambley, T.W. The Discovery and Development of Cisplatin. J. Chem. Educ. 2006, 83, 728–734. [Google Scholar] [CrossRef]
- Aldossary, S.A. Review on Pharmacology of Cisplatin: Clinical Use, Toxicity and Mechanism of Resistance of Cisplatin. Biomed. Pharmacol. J. 2019, 12, 07–15. [Google Scholar] [CrossRef]
- Waissbluth, S.; Daniel, S.J. Cisplatin-induced ototoxicity: Transporters playing a role in cisplatin toxicity. Hear. Res. 2013, 299, 37–45. [Google Scholar] [CrossRef]
- Gately, D.P.; Howell, S.B. Cellular accumulation of the anticancer agent cisplatin: A review. Br. J. Cancer 1993, 67, 1171–1176. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Lippard, S.J. Cellular processing of platinum anticancer drugs. Nat. Rev. Drug Discov. 2005, 4, 307–320. [Google Scholar] [CrossRef]
- Raymond, E.; Faivre, S.; Chaney, S.; Woynarowski, J.; Cvitkovic, E. Cellular and molecular pharmacology of oxaliplatin. Mol. Cancer Ther. 2002, 1, 227–235. [Google Scholar] [PubMed]
- Fuertes, M.A.; Castilla, J.; Alonso, C.; Pérez, J.M. Cisplatin Biochemical Mechanism of Action: From Cytotoxicity to Induction of Cell Death Through Interconnections Between Apoptotic and Necrotic Pathways. Curr. Med. Chem. 2003, 10, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Casini, A.; Hartinger, C.; Gabbiani, C.; Mini, E.; Dyson, P.J.; Keppler, B.K.; Messori, L. Gold(III) compounds as anticancer agents: Relevance of gold–protein interactions for their mechanism of action. J. Inorg. Biochem. 2008, 102, 564–575. [Google Scholar] [CrossRef]
- Reedijk, J. Improved understanding in platinium antitumour chemistry. Chem. Commun. 1996, 801–806. [Google Scholar] [CrossRef]
- O’Sullivan, D.A.; Lepkowski, W. Chemical Science. Chem. Eng. News Arch. 1990, 68, 42–61. [Google Scholar] [CrossRef]
- Messori, L.; Merlino, A. Cisplatin binding to proteins: A structural perspective. Co-ord. Chem. Rev. 2016, 315, 67–89. [Google Scholar] [CrossRef]
- Sooriyaarachchi, M.; Narendran, A.; Gailer, J. Comparative hydrolysis and plasma protein binding of cis-platin and carboplatin in human plasma in vitro. Metallomics 2011, 3, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Tsang, C.-N.; Sun, H. Identification and characterization of metallodrug binding proteins by (metallo)proteomics. Metallomics 2009, 1, 25–31. [Google Scholar] [CrossRef]
- Loreto, D.; Merlino, A. Interaction of Platinum-based Drugs with Proteins: An Overview of Representative Crystallographic Studies. Curr. Top. Med. Chem. 2021, 21, 6–27. [Google Scholar] [CrossRef]
- Matera-Witkiewicz, A.; Brasuń, J.; Świątek-Kozłowska, J.; Pratesi, A.; Ginanneschi, M.; Messori, L. Short-chain oligopeptides with copper(II) binding properties: The impact of specific structural modifications on the copper(II) coordination abilities. J. Inorg. Biochem. 2009, 103, 678–688. [Google Scholar] [CrossRef] [PubMed]
- Tolbatov, I.; Cirri, D.; Marchetti, L.; Marrone, A.; Coletti, C.; Re, N.; La Mendola, D.; Messori, L.; Marzo, T.; Gabbiani, C.; et al. Mechanistic Insights Into the Anticancer Properties of the Auranofin Analog Au(PEt3)I: A Theoretical and Experimental Study. Front. Chem. 2020, 8, 812. [Google Scholar] [CrossRef] [PubMed]
- Massai, L.; Zoppi, C.; Cirri, D.; Pratesi, A.; Messori, L. Reactions of Medicinal Gold(III) Compounds With Proteins and Peptides Explored by Electrospray Ionization Mass Spectrometry and Complementary Biophysical Methods. Front. Chem. 2020, 8, 581648. [Google Scholar] [CrossRef]
- Ferraro, G.; Cirri, D.; Marzo, T.; Pratesi, A.; Messori, L.; Merlino, A. The first step of arsenoplatin-1 aggregation in solution unveiled by solving the crystal structure of its protein adduct. Dalton Trans. 2021, 50, 68–71. [Google Scholar] [CrossRef]
- Marzo, T.; De Pascali, S.A.; Gabbiani, C.; Fanizzi, F.P.; Messori, L.; Pratesi, A. ESI–MS studies of the reactions of novel platinum(II) complexes containing O,O′-chelated acetylacetonate and sulfur ligands with selected model proteins. BioMetals 2017, 30, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Canil, G.; Braccini, S.; Marzo, T.; Marchetti, L.; Pratesi, A.; Biver, T.; Funaioli, T.; Chiellini, F.; Hoeschele, J.D.; Gabbiani, C.; et al. Photocytotoxic Pt(iv) complexes as prospective anticancer agents. Dalton Trans. 2019, 48, 10933–10944. [Google Scholar] [CrossRef] [PubMed]
- Canil, G.; Gurruchaga-Pereda, J.; Marchetti, L.; Pratesi, A.; Funaioli, T.; Salassa, L.; Gabbiani, C. Flavin-Catalyzed Light-Activatable Pt(IV) Compound as Potential Photochemothera-peutic. Inorg. Chem. (Unpublished work).
- Gabano, E.; Perin, E.; Fielden, C.; Platts, J.A.; Gallina, A.; Rangone, B.; Ravera, M. How to obtain Pt( iv ) complexes suitable for conjugation to nanovectors from the oxidation of [PtCl(terpyridine)] +. Dalton Trans. 2017, 46, 10246–10254. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, C.; Duce, C.; Russo, D.; Tiné, M.R.; Wurm, F.R. PPEylation of proteins: Synthesis, activity, and stability of myoglobin-polyphosphoester conjugates. Eur. Polym. J. 2018, 108, 357–363. [Google Scholar] [CrossRef]
- Pelosi, C.; Duce, C.; Tinè, M.R.; Wurm, F.R. Effect of polymer hydrophilicity and molar mass on the properties of the protein in protein-polymer conjugates: The case of PPE-ylated myoglobin. Biomacromolecules 2021. [Google Scholar] [CrossRef]
- Cooper, A.; Nutley, M.A.; Wadood, A. Differential Scanning Microcalorimetry. In Protein-Ligand Interactions: Hydrodynamics and calorimetry; Harding, S.E., Chowdhry, B.Z., Eds.; Oxford University Press: Oxford, UK, 2000; ISBN 9780199637461. [Google Scholar]
- Shrake, A.; Ross, P.D. Ligand-induced biphasic protein denaturation. J. Biol. Chem. 1990, 265, 5055–5059. [Google Scholar] [CrossRef]
- Macii, F.; Biver, T. Spectrofluorimetric analysis of the binding of a target molecule to serum albumin: Tricky aspects and tips. J. Inorg. Biochem. 2021, 216, 111305. [Google Scholar] [CrossRef]
- Musa, K.A.; Ridzwan, N.F.W.; Mohamad, S.B.; Tayyab, S. Exploring the combination characteristics of lumefantrine, an antimalarial drug and human serum albumin through spectroscopic and molecular docking studies. J. Biomol. Struct. Dyn. 2021, 39, 691–702. [Google Scholar] [CrossRef]
- Pelosi, C.; Saitta, F.; Wurm, F.R.; Fessas, D.; Tinè, M.R.; Duce, C. Thermodynamic stability of myoglobin-poly(ethylene glycol) bioconjugates: A calorimetric study. Thermochim. Acta 2019, 671, 26–31. [Google Scholar] [CrossRef]
- Barbiroli, A.; Bonomi, F.; Ferranti, P.; Fessas, D.; Nasi, A.; Rasmussen, P.; Iametti, S. Bound Fatty Acids Modulate the Sensitivity of Bovine β-Lactoglobulin to Chemical and Physical Denaturation. J. Agric. Food Chem. 2011, 59, 5729–5737. [Google Scholar] [CrossRef] [PubMed]
- Ausili, A.; Pennacchio, A.; Staiano, M.; Dattelbaum, J.; Fessas, D.; Schiraldi, A.; D’Auria, S. Amino acid transport in thermophiles: Characterization of an arginine-binding protein from Thermotoga maritima. 3. Conformational dynamics and stability. J. Photochem. Photobiol. B Biol. 2013, 118, 66–73. [Google Scholar] [CrossRef]
- Tsitsanou, K.E.; Drakou, C.E.; Thireou, T.; Gruber, A.V.; Kythreoti, G.; Azem, A.; Fessas, D.; Eliopoulos, E.; Iatrou, K.; Zographos, S.E. Crystal and Solution Studies of the “Plus-C” Odorant-binding Protein 48 from Anopheles gambiae. J. Biol. Chem. 2013, 288, 33427–33438. [Google Scholar] [CrossRef] [Green Version]
- Caldinelli, L.; Iametti, S.; Barbiroli, A.; Fessas, D.; Bonomi, F.; Piubelli, L.; Molla, G.; Pollegioni, L. Relevance of the flavin binding to the stability and folding of engineered cholesterol oxidase containing noncovalently bound FAD. Protein Sci. 2008, 17, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Biancardi, A.; Biver, T.; Burgalassi, A.; Mattonai, M.; Secco, F.; Venturini, M. Mechanistic aspects of thioflavin-T self-aggregation and DNA binding: Evidence for dimer attack on DNA grooves. Phys. Chem. Chem. Phys. 2014, 16, 20061–20072. [Google Scholar] [CrossRef]
- Zoppi, C.; Messori, L.; Pratesi, A. ESI MS studies highlight the selective interaction of Auranofin with protein free thiols. Dalton Trans. 2020, 49, 5906–5913. [Google Scholar] [CrossRef]
- Pratesi, A.; Cirri, D.; Ciofi, L.; Messori, L. Reactions of Auranofin and Its Pseudohalide Derivatives with Serum Albumin Investigated through ESI-Q-TOF MS. Inorg. Chem. 2018, 57, 10507–10510. [Google Scholar] [CrossRef] [PubMed]
- Zoppi, C.; Nocentini, A.; Supuran, C.T.; Pratesi, A.; Messori, L. Native mass spectrometry of human carbonic anhydrase I and its inhibitor complexes. JBIC J. Biol. Inorg. Chem. 2020, 25, 979–993. [Google Scholar] [CrossRef]
- Heck, A.J.R.; Heuvel, R.H.H.V.D. Investigation of intact protein complexes by mass spectrometry. Mass Spectrom. Rev. 2004, 23, 368–389. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Kellie, J.F.; Hottenstein, C.S.; Szapacs, M.E. Native high-resolution mass spectrometry analysis of noncovalent protein complexes up to 450 kDa. Bioanalysis 2020, 12, 1353–1362. [Google Scholar] [CrossRef] [PubMed]
- Corinti, D.; Gregori, B.; Guidoni, L.; Scuderi, D.; McMahon, T.B.; Chiavarino, B.; Fornarini, S.; Crestoni, M.E. Complexation of halide ions to tyrosine: Role of non-covalent interactions evidenced by IRMPD spectroscopy. Phys. Chem. Chem. Phys. 2018, 20, 4429–4441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.K.; Katta, V.; Beavis, R.C.; Chait, B.T. Origin and removal of adducts (molecular mass = 98 u) attached to peptide and protein ions in electrospray ionization mass spectra. J. Am. Soc. Mass Spectrom. 1990, 1, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Casini, A.; Mastrobuoni, G.; Temperini, C.; Gabbiani, C.; Francese, S.; Moneti, G.; Supuran, C.T.; Scozzafava, A.; Messori, L. ESI mass spectrometry and X-ray diffraction studies of adducts between anticancer platinum drugs and hen egg white lysozyme. Chem. Commun. 2006, 156–158. [Google Scholar] [CrossRef] [PubMed]
- Messori, L.; Marzo, T.; Gabbiani, C.; Valdes, A.A.; Quiroga, A.G.; Merlino, A. Peculiar Features in the Crystal Structure of the Adduct Formed between cis-PtI2(NH3)2 and Hen Egg White Lysozyme. Inorg. Chem. 2013, 52, 13827–13829. [Google Scholar] [CrossRef]
- Gabbiani, C.; Massai, L.; Scaletti, F.; Michelucci, E.; Maiore, L.; Cinellu, M.A.; Messori, L. Protein metalation by metal-based drugs: Reactions of cytotoxic gold compounds with cytochrome c and lysozyme. JBIC J. Biol. Inorg. Chem. 2012, 17, 1293–1302. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, D.J.; Echavarren, A.M.; De Arellano, M.C.R. Divergent Behavior of Palladium(II) and Platinum(II) in the Metalation of 1,3-Di(2-pyridyl)benzene. Organometallics 1999, 18, 3337–3341. [Google Scholar] [CrossRef]
- Barone, G.; Del Vecchio, P.G.G.; Fessas, D.; Giancola, C.; Graziano, G. Theseus: A new software package for the handling and analysis of thermal denaturation data of biological macromolecules. J. Therm. Anal. Calorim. 1992, 38, 2779–2790. [Google Scholar] [CrossRef]
- Saitta, F.; Signorelli, M.; Bramanti, E.; Pizzimenti, S.; Pelosi, C.; Duce, C.; Fessas, D.; Bonaduce, I.; Tinè, M.R. DSC on ovalbumin-hematite “tempera” paints: The role of water and pigment on protein stability. Thermochim. Acta 2020, 694, 178780. [Google Scholar] [CrossRef]
- Press, W.H.; Flannery, B.P.; Teukolski, S.A.; Weterling, W.T. Numerical Recipes: The Art of Scientific Computing; Cambridge University Press: Cambridge, UK, 1989; ISBN 9780521880688. [Google Scholar]
- Barone, G.; Del Vecchio, P.; Fessas, D.; Giancola, C.; Graziano, G.; Pucci, P.; Riccio, A.; Ruoppolo, M. Thermal denaturation of ribonuclease T1 a DSC study. J. Therm. Anal. Calorim. 1992, 38, 2791–2802. [Google Scholar] [CrossRef]
- Arakawa, T.; Kita, Y.; Timasheff, S.N. Protein precipitation and denaturation by dimethyl sulfoxide. Biophys. Chem. 2007, 131, 62–70. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample 1 | Incub. (h) | Tmax (°C) 2 | ΔdH (kJ mol−1) 3 | Sample 4 | Incub. (h) | Tmax (°C) 2 | ΔdH (kJ mol−1) 3 |
|---|---|---|---|---|---|---|---|
| HEWL | 0 | 76.3 | 550 | RNase A | 0 | 59.4 | 440 |
| HEWL + 1 | 24 | 71.8 | 515 | RNase A + 1 | 24 | 56.4 | 220 * |
| 48 | 71.8 | 510 | |||||
| HEWL + 2 | 24 | 71.8 | 550 | RNase A + 2 | 24 | 51.8 | 420 * |
| 48 | 71.8 | 550 | |||||
| HEWL + 3 | 24 | 75.5 | 550 | RNase A + 3 | 24 | 57.6 | 425 * |
| 48 | 75.5 | 550 |
| Titration | K (M−1) 1 | K′ (M−1) 2 | n 2 |
|---|---|---|---|
| HEWL + 1 | (1.7 ± 0.2) × 104 | (1.5 ± 0.8) × 104 | 0.91 ± 0.05 |
| HEWL + 2 | (1.2 ± 0.4) × 104 | (1.2 ± 0.5) × 104 | 1.00 ± 0.04 |
| HEWL + 3 | (4 ± 2) × 102 | - | ≅ 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pelosi, C.; Saitta, F.; Zerino, C.; Canil, G.; Biver, T.; Pratesi, A.; Duce, C.; Fessas, D.; Gabbiani, C.; Tiné, M.R. Thermodynamic Evaluation of the Interactions between Anticancer Pt(II) Complexes and Model Proteins. Molecules 2021, 26, 2376. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082376
Pelosi C, Saitta F, Zerino C, Canil G, Biver T, Pratesi A, Duce C, Fessas D, Gabbiani C, Tiné MR. Thermodynamic Evaluation of the Interactions between Anticancer Pt(II) Complexes and Model Proteins. Molecules. 2021; 26(8):2376. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082376
Chicago/Turabian StylePelosi, Chiara, Francesca Saitta, Caterina Zerino, Giovanni Canil, Tarita Biver, Alessandro Pratesi, Celia Duce, Dimitrios Fessas, Chiara Gabbiani, and Maria Rosaria Tiné. 2021. "Thermodynamic Evaluation of the Interactions between Anticancer Pt(II) Complexes and Model Proteins" Molecules 26, no. 8: 2376. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082376