
Preparation of Amylose-Oligo[(R)-3-hydroxybutyrate] Inclusion Complex by Vine-Twining Polymerization


Graduate School of Science and Engineering, Kagoshima University, 1-21-40 Korimoto, Kagoshima 890-0065, Japan
*
Author to whom correspondence should be addressed.
Molecules 2021, 26(9), 2595; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092595
Submission received: 13 April 2021
/
Revised: 27 April 2021
/
Accepted: 28 April 2021
/
Published: 29 April 2021
(This article belongs to the Special Issue Polysaccharide-Based Materials II)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:In this study, we attempted to prepare an amylose-oligo[(R)-3-hydroxybutyrate] (ORHB) inclusion complex using a vine-twining polymerization approach. Our previous studies indicated that glucan phosphorylase (GP)-catalyzed enzymatic polymerization in the presence of appropriate hydrophobic guest polymers produces the corresponding amylose–polymer inclusion complexes, a process named vine-twining polymerization. When vine-twining polymerization was conducted in the presence of ORHB under general enzymatic polymerization conditions (45 °C), the enzymatically produced amylose did not undergo complexation with ORHB. However, using a maltotriose primer in the same polymerization system at 70 °C for 48 h to obtain water-soluble amylose, called single amylose, followed by cooling the system over 7 h to 45 °C, successfully induced the formation of the inclusion complex. Furthermore, enzymatic polymerization initiated from a longer primer under the same conditions induced the partial formation of the inclusion complex. The structures of the different products were analyzed by X-ray diffraction, 1H-NMR, and IR measurements. The mechanism of formation of the inclusion complexes discussed in the study is proposed based on the additional experimental results.
1. Introduction
Amylose is an abundant polysaccharide composed of α(1→4)-linked glucose (G) repeating units [1]. Besides its role in nature as an energy resource as a component of starch, amylose has been identified as a functional supramolecular material owing to its left-handed helical conformation with controlled regularity [2,3]. For example, as the inside of the helix is hydrophobic, amylose readily complexes within its cavity hydrophobic guest molecules of appropriate sizes and geometries, such as fatty acids, to form supramolecular inclusion complexes [4].
The complete separation of amylose from amylopectin, the other component in natural starch sources, is difficult. However, an enzymatic approach using α-glucan phosphorylase (GP) catalysis to form a regio- and stereo-controlled α(1→4)-glycosidic arrangement is a powerful tool to obtain pure amylose [5,6,7,8,9,10]. GP catalyzes the enzymatic polymerization of α-d-glucose 1-phosphate (G-1-P), the monomer, initiated from a maltooligosaccharide primer, resulting in the formation of α(1→4)-glucan (amylose) according to the following reversible reaction: [α(1→4)-G]n + G-1-P ⇄ [α(1→4)-G]n+1 + inorganic phosphate. GP-catalyzed enzymatic polymerization under ambient conditions, such as at 45 °C, yields an insoluble amylosic product in aqueous media through the formation of a well-known double-helical assembly [11,12]. We found that enzymatic polymerization using thermostable GP (isolated from the thermophilic bacterium Aquifex aeolicus VF5) at elevated temperatures, for example, at 70 °C, produced a water-soluble amylosic product with a single chain (called single amylose) in an equilibrium state between chain elongation and cleavage reactions (polymerization and phosphorolysis, respectively) [13].
Interestingly, when the GP-catalyzed enzymatic polymerization is conducted in the presence of appropriate hydrophobic polymers dispersed in aqueous buffer solvents, these polymers are gradually included in the helical cavity of the produced amylose chains during the propagation from the short primer toward longer α(1→4)-glucan chains, leading to the formation of amylose–polymer inclusion complexes [14,15,16,17,18]. As a controlled water-insoluble assembly is formed, these complexes gradually precipitate in the system. As the polymerization resembles the growth of plant vines that gradually twine around a support rod, the system was termed vine-twining polymerization. Both hydrophilic and strongly hydrophobic polymers are not suitable as guests for vine-twining polymerization as they undergo weak hydrophobic interactions with the amylosic cavity and aggregate in aqueous media, respectively [19,20]. Therefore, polyesters with moderate hydrophobicity, such as poly(ε-caprolactone) (PCL), poly(δ-valerolactone) (PVL), and poly(l-lactide), are ideal guests for vine-twining polymerization and have been shown to form the corresponding amylosic inclusion complexes [21,22,23].
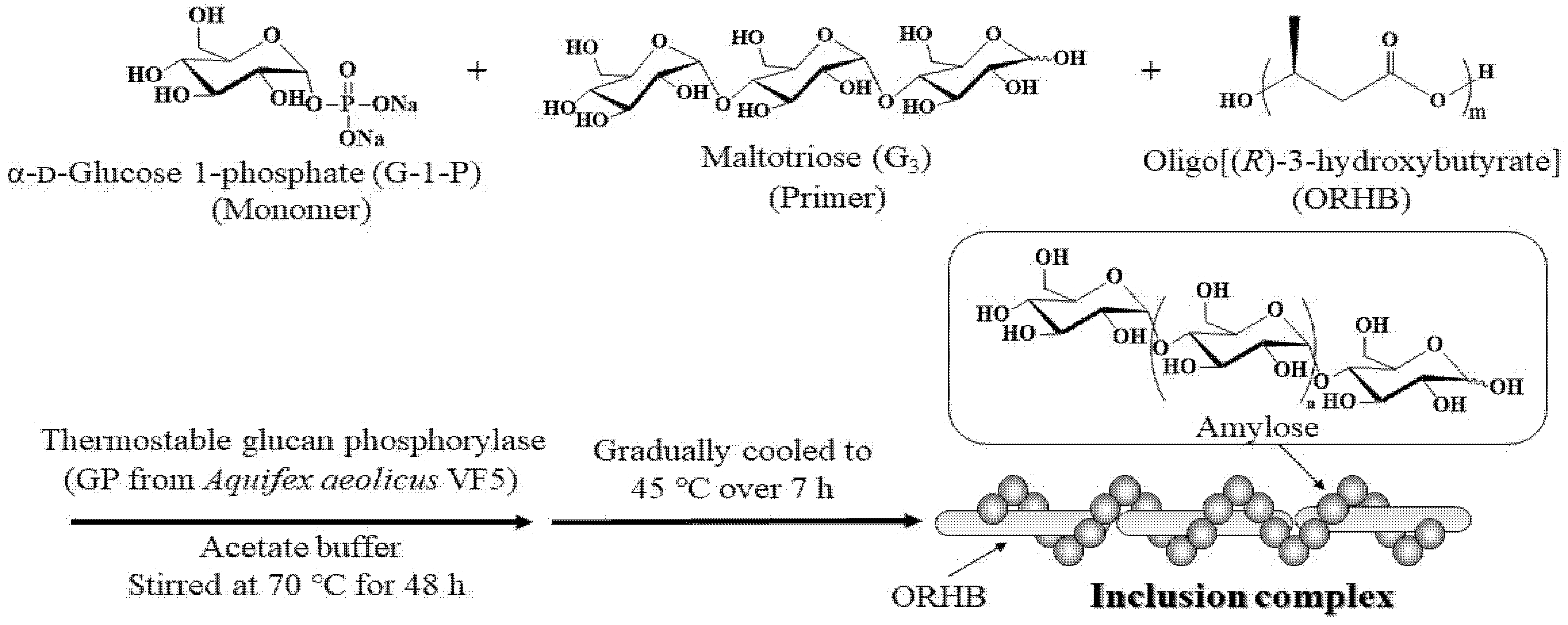
We attempted to use poly[(R)-3-hydroxybutyrate] (PRHB), a well-known polyester biosynthesized by microorganisms [24], as a new guest polyester with a similar structure to those discussed above for vine-twining polymerization. Owing to the high molecular weight (MW = ~500,000) and strong hydrophobicity of biosynthesized PRHB, our attempt at vine-twining polymerization under typical conditions at 45–50 °C mostly has not induced the inclusion of PRHB into the cavity of amylose. Therefore, in this study, we employed low-molecular-weight oligo[(R)-3-hydroxybutyrate] (ORHB) (<1000), prepared by partial alkaline hydrolysis of the parent PRHB, as the guest for the vine-twining polymerization. Thermostable GP-catalyzed enzymatic polymerization in the presence of ORHB was found to produce the amylose–ORHB inclusion complexes under controlled temperature conditions, as shown in Scheme 1. In addition, this study focuses on the extension of guest polymers, i.e., ORHB, in the vine-twining polymerization and, furthermore, the amylose–ORHB inclusion complex can be expected as a new bio-based and biocompatible material owing to biodegradability in both the components.
2. Results and Discussion
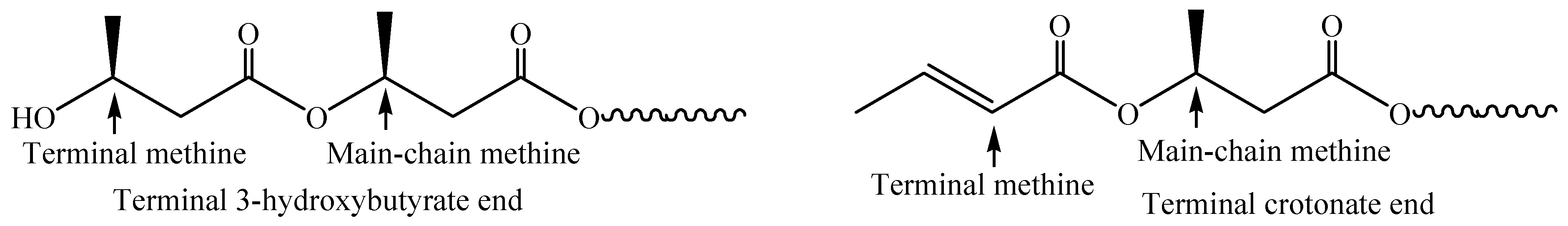
The low-molecular-weight ORHB was first prepared by partial alkaline hydrolysis of a commercially available PRHB (MW = ~500,000) in the presence of sodium hydroxide in chloroform at room temperature for 7 h. The presence of signals corresponding to both terminal 3-hydroxybutyrate and crotonate (derived by the dehydration of terminal 3-hydroxybuturate) in the 1H-NMR spectrum of the isolated product in CDCl3 supported the formation of the desired ORHB (Figure 1) [25]. Based on the ratio of the integrals of the terminal methine signals to the main-chain methine signals, the MW of the product was calculated to be, e.g., ~530.
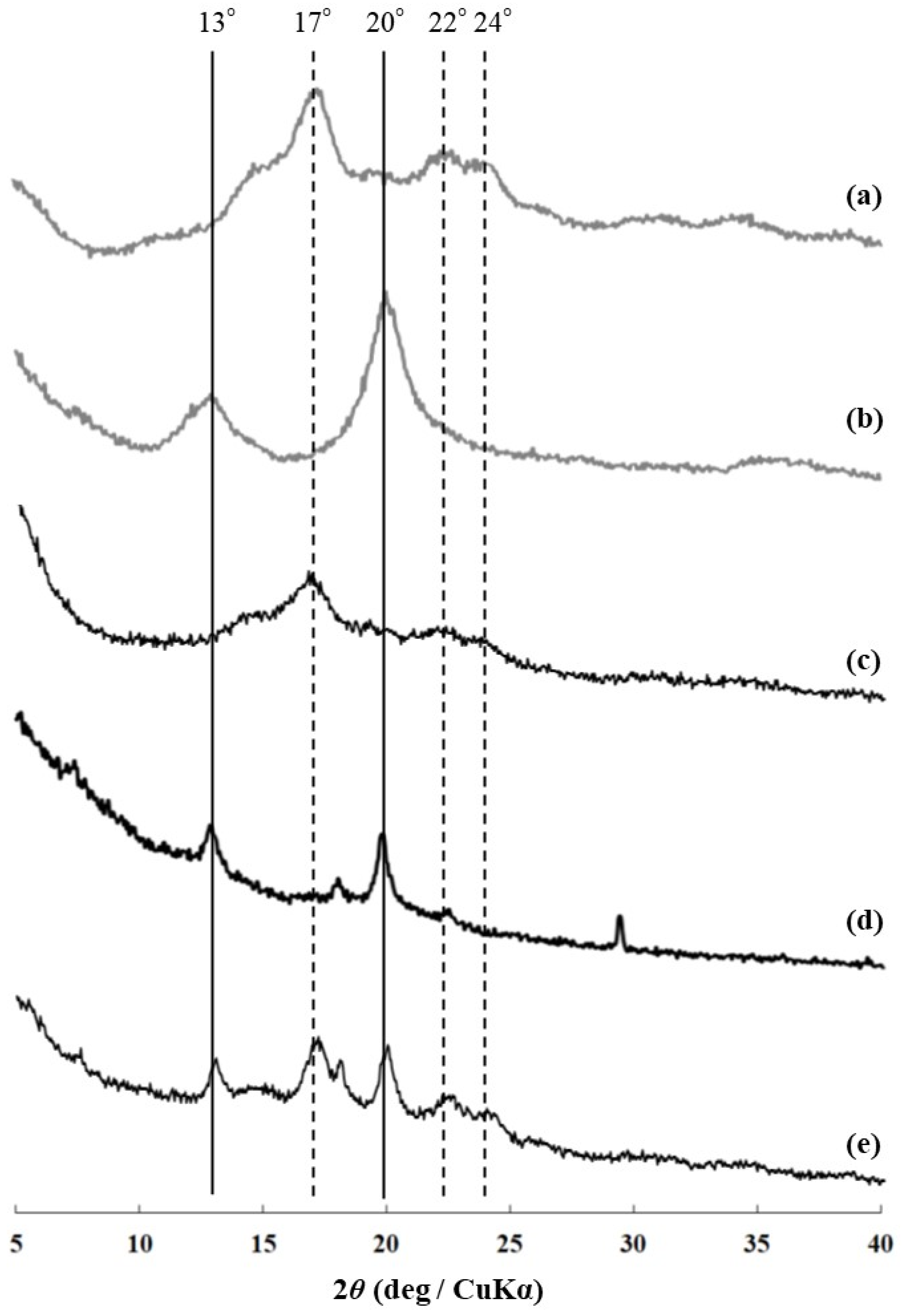
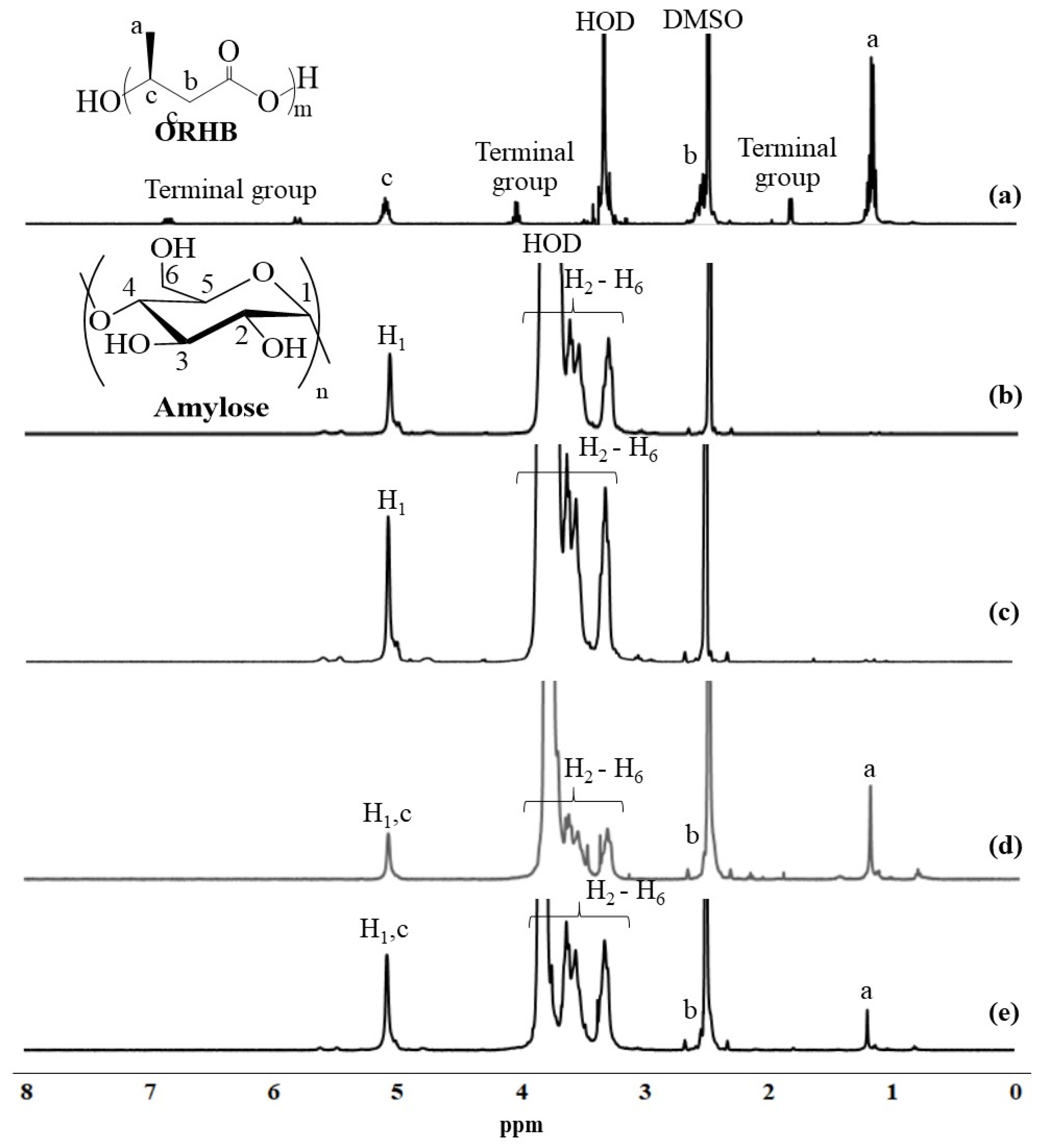
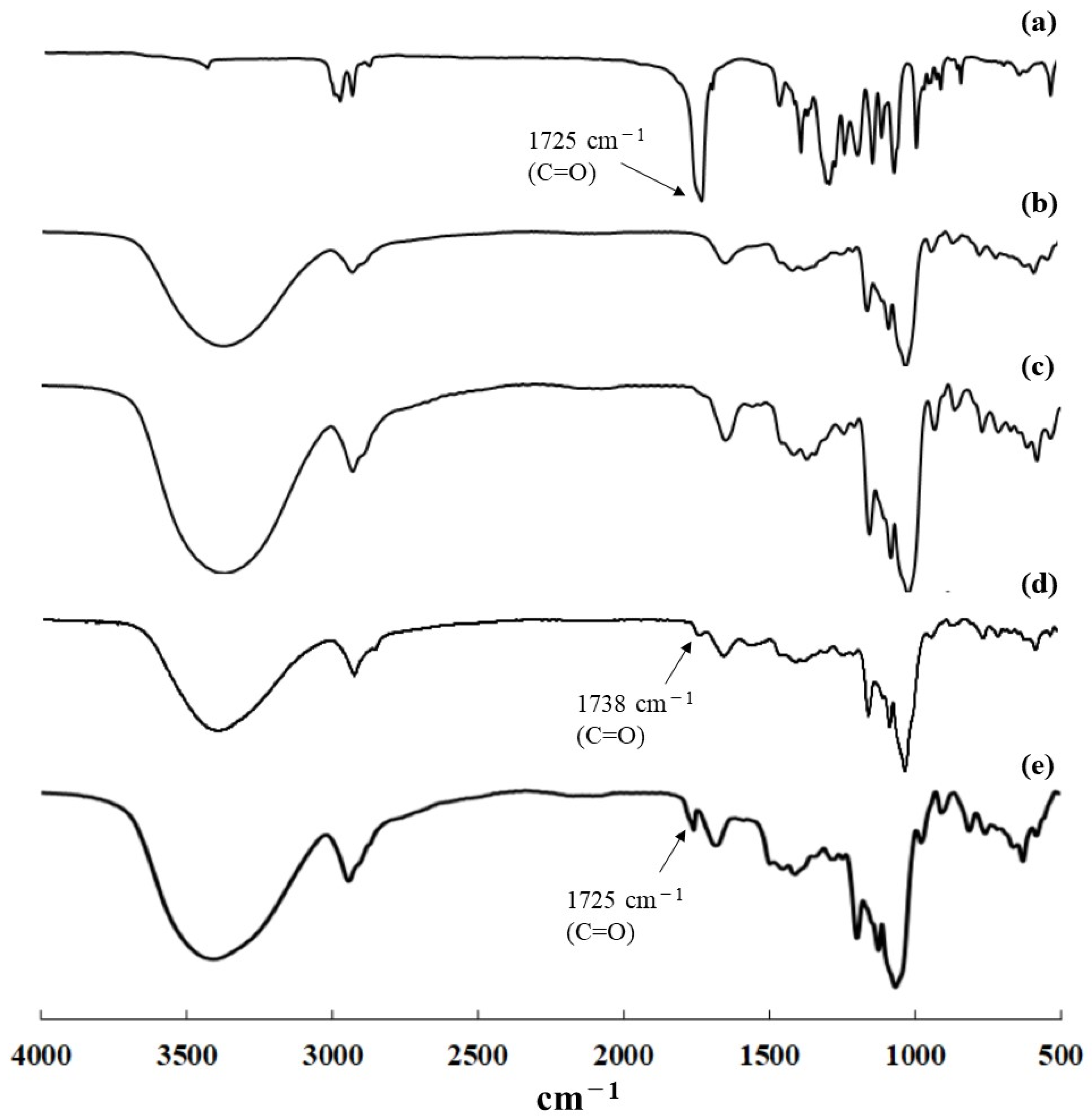
Vine-twining polymerization using ORHB was first performed by the thermostable GP (from Aquifex aeolicus VF5)-catalyzed enzymatic polymerization of G-1-P from a maltoheptaose primer in acetate buffer solvent under the typical conditions reported in our previous studies: ORHB was dispersed in the mixture at 45 °C for 24 h with vigorous stirring [21,22]. The precipitate was isolated by centrifugation, washed successively with water, acetone, and chloroform, and subsequently dried under reduced pressure to obtain the product. The powder X-ray diffraction (XRD) profile of the product was identical to that of the pure double-helical amylose (17°, 22°, and 24°), but completely different from that of the previously reported amylose–PCL inclusion complex (13° and 20°) [21,22] (Figure 2a–c). The 1H-NMR in DMSO-d6 + D2O and IR spectra of the product showed signals corresponding to pure amylose but did not exhibit characteristic signals derived from ORHB, such as a methyl signal and a carbonyl absorption peak, respectively (Figure 3a–c and Figure 4a–c). These results strongly suggest that under these conditions, the enzymatically produced amylose did not include ORHB, and instead favorably formed a double helix structure.
We then attempted to obtain the inclusion complex from ORHB by enzymatic production through the formation of water-soluble amylose without a double-helical assembly, called single amylose, at elevated temperatures (Scheme 1) [13]. The thermostable GP-catalyzed enzymatic polymerization of G-1-P from a maltotriose (G3) primer (80:1) was conducted in the presence of ORHB (MW = 530) dispersed in an acetate buffer solvent. The mixture was maintained at 70 °C for 48 h; we have already confirmed that such elevated reaction temperatures in the thermostable GP-catalyzed enzymatic polymerization do not affect the chemical structure of the produced amylose [13,23]. Subsequently, to permit the further progress of the polymerization and simultaneously accelerate the complexation and precipitation, the mixture was stirred and gradually cooled to a temperature of 45 °C over 7 h. The product was then isolated using the same procedure as described above and characterized by XRD, 1H-NMR, and IR measurements. The XRD profile of the isolated product showed diffraction peaks at 13° and 20°, which was identical to that of the 61 helix in the amylose–PCL inclusion complex [26], and completely different from that of the pure amylose (Figure 2a,b,d), suggesting the successful formation of the inclusion complex. The structure of the inclusion complex was also supported by the 1H-NMR and IR results. The 1H-NMR spectrum of the isolated product in DMSO-d6 + D2O showed signals corresponding to both amylose and ORHB (Figure 3d). Generally, one helical turn of amylose is composed of approximately six repeating G units when linear molecules, such as PCL, are included, as suggested by the corresponding XRD profiles (Figure 2b,d) that show a repeat distance of 0.800 nm for the amylose helix [27]. On the other hand, the unit length of ORHB was calculated to be 0.446 nm. Therefore, an average of 3.38 repeating G units in amylose corresponds to the length of one unit of ORHB. Based on these calculations, the ratio of the integrals of the H1 (anomeric) signals at δ 4.98–5.18 to the methyl signal a at δ 1.19 (a/H1) in the 1H NMR spectrum is theoretically 0.897. The a/H1 value in the 1H NMR spectrum of the product in Figure 3d was calculated to be 1.18, which is slightly higher than the theoretical value. This indicates that the amylose cavity quantitatively includes ORHB. The IR spectrum of the product exhibited a band corresponding to ester C=O absorption at 1738 cm−1 (Figure 4d). This band indicated the presence of ORHB, although it was detected at a different wavenumber from the carbonyl absorption of pure crystalline ORHB, at 1725 cm−1 (Figure 4a), similar to the absorption shift reported in a previous study on vine-twining polymerization using PVL [22]. The considerable difference in these peaks is because of the non-crystalline nature of ORHB in the product, indicating that the inclusion of amylose prevents the formation of a crystalline structure in the product. Dissociation of the inclusion complex was carried out by the treatment with DMSO at elevated temperatures according to our reported procedure [28]. The degree of polymerization (DP) and MW of the dissociated amylose were determined from the λmax value in the UV–Vis spectra of the violet solution of a complex with iodine to be 62.3 and 10,100, respectively [29,30]. The lengths of the amylose helix and the ORHB chain estimated from their DP values (62.3 and 3.95, respectively) were 8.3 and 1.76 nm, respectively, which suggests that one amylose chain includes, on average, four or five ORHB molecules. From the amylose/ORHB unit ratio of 1:0.28 in the product (calculated by the actual a/H1 value = 1.18), which corresponded to their weight ratio of 1:0.15, the yield of the inclusion complex was calculated to be ca. 10% based on the amounts of the G3 primer in the feed and the product (5.5 and 12.6 mg, respectively, Section 3.3). The relatively low yield of the amylosic inclusion complex owed to equilibrium nature of the GP-catalyzed enzymatic polymerization, as a similar value was reported in the vine-twining polymerization using PCL [21].
The following process is proposed for the formation of the amylose–ORHB inclusion complex during the enzymatic polymerization (Figure 5). At an elevated temperature (70 °C), the thermostable GP-catalyzed enzymatic polymerization from the G3 primer produces water-soluble single amylose with some chain elongation. To reach equilibrium during chain elongation, the single amylose weakly interacts with ORHB. The cooling of the reaction mixture to 45 °C over 7 h induces the weak assembly of complexes, shifting the equilibrium to further chain elongation. The elongated amylose chain then includes additional ORHB molecules to form a water-insoluble assembly from the regular inclusion complexes.
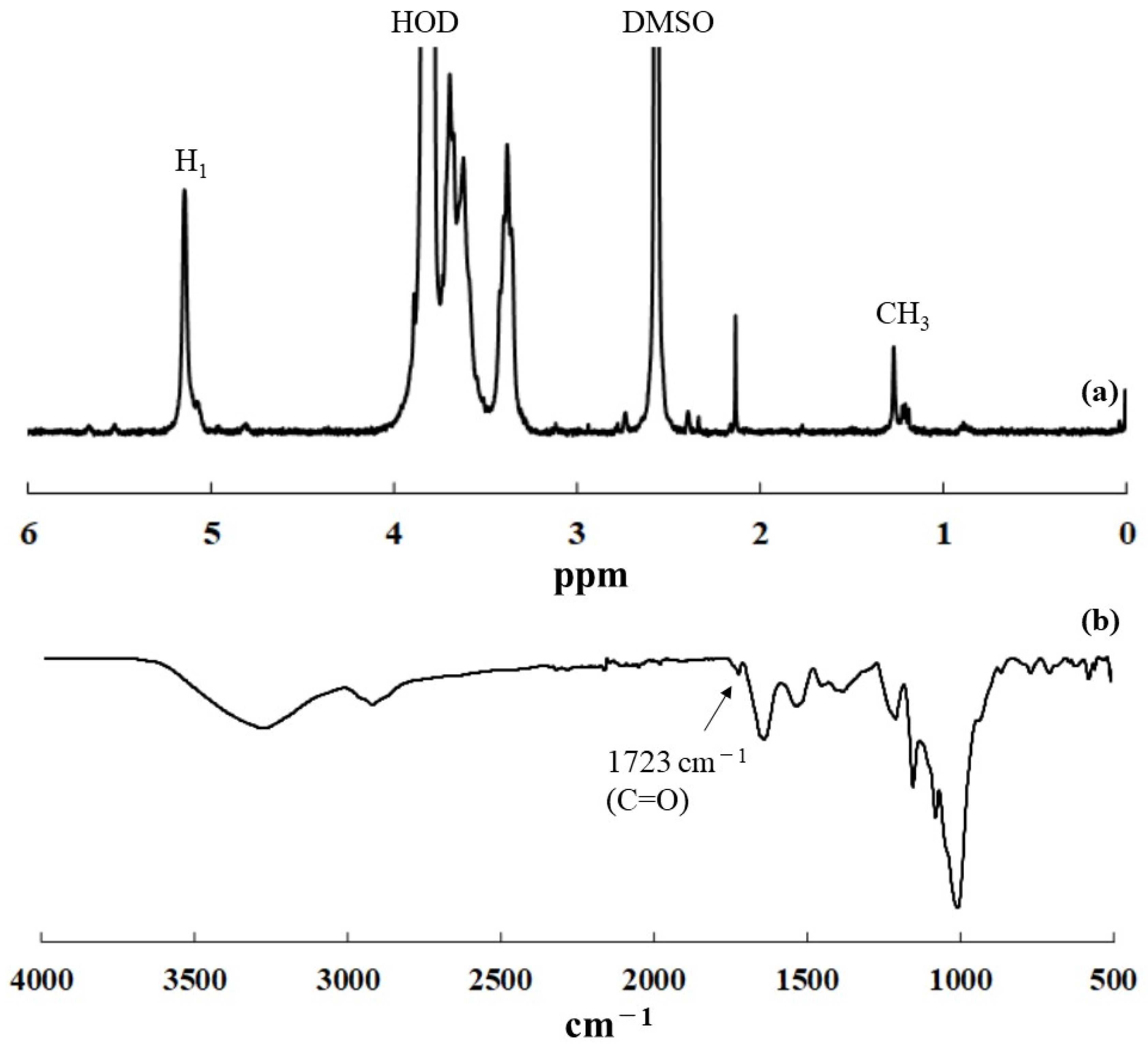
To support the above hypothesis, thermostable GP-catalyzed enzymatic polymerization was conducted in the presence of ORHB dispersed in acetate buffer solvent at 70 °C for 48 h to attain equilibrium with the production of single amylose. The resulting reaction mixture was immediately subjected to centrifugation without cooling to isolate a water-insoluble fraction. The resulting product was washed with water and chloroform. Despite thorough washing with appropriate solvents to remove the single amylose (water) and ORHB (chloroform), the 1H-NMR spectrum of the product in DMSO-d6 + D2O showed a methyl signal at δ 1.27 from ORHB and an H1 signal at δ 5.15 from amylose (Figure 6a). In addition, the IR spectrum of the product exhibited a carbonyl absorption at 1723 cm−1 (Figure 6b), indicating the presence of ORHB in the product. These analytical results indicated the occurrence of some interaction between the single amylose and ORHB, which may be a key material at the initial stage for the formation of the inclusion complex in the present system.
To confirm the necessity of the chain-elongation process from the shorter G3 to the longer single amylose at the early stage of complexation, thermostable GP-catalyzed enzymatic polymerization was performed in the presence of a water-soluble amylose (MW = 2800) as the primer instead of G3. The reaction was performed under similar conditions as above, at controlled temperatures (70 °C for 48 h and subsequent cooling to 45 °C over 7 h). The XRD profile of the product obtained using the same isolation procedure showed diffraction peaks that could be assigned to both the amylosic inclusion complex at 13° and 20° and the double-helical amylose at 17°, 22°, and 24° (Figure 2e). This result indicated that the enzymatically produced amylose from the water-soluble amylose primer partially formed an inclusion complex with ORHB. Although the 1H-NMR spectrum of the product in DMSO-d6 + D2O showed a methyl signal at δ 1.26 from ORHB (Figure 3e), its carbonyl absorption in the IR spectrum was detected at 1725 cm−1, which corresponded to the crystalline structure (Figure 4e). The IR data suggested that ORHB was partly included in the cavity of amylose, and the excluded ORHB chains probably formed a crystalline structure. However, the formation of the crystalline structure from the ORHB in the product was not confirmed by XRD measurements, because the diffraction peaks ascribed to the ORHB crystal completely overlapped with the peaks from the inclusion complex and the double helix of amylose (data not shown).
3. Materials and Methods
3.1. Materials
Thermostable GP from Aquifex aeolicus VF5 was supplied by Ezaki Glico Co., Ltd. (Osaka, Japan) [31,32]. An amylose sample was prepared by the thermostable GP-catalyzed enzymatic polymerization of Glc-1-P [9]. The amylose–PCL inclusion complex was prepared according to a procedure from the literature [21]. The biosynthesized PRHB with MW = ~500,000 and water-soluble amylose with MW = 2800 were purchased from Sigma-Aldrich (Darmstadt, Germany) and Tokyo Chemical Industry Co., Ltd. (Tokyo, Japan), respectively. Other reagents and solvents were available commercially and used without further purification.
3.2. Preparation of ORHB
A typical experimental procedure for the preparation of ORHB is as follows. A mixture of the commercially available PRHB (0.54 g) with sodium hydroxide (0.89 g) and chloroform (80 mL) was ultrasonicated at 50 °C for 1 h and then stirred at room temperature for 7 h. The reaction mixture was washed successively with 1 mol/L hydrochloric acid (5.0 mL) and water (80 mL). The chloroform layer was dried over anhydrous sodium sulfate, filtered, evaporated, and dried under reduced pressure to give ORHB (0.16 g). 1H-NMR (CDCl3): δ 1.22–1.27 (m, CH3 of main-chain), 1.85–1.88 (m, CH3 of terminal crotonate), 2.42–2.68 (m, CH2 of main-chain), 4.11–4.18 (m, CH of terminal 3-hydroxybutyrate), 5.18–5.36 (m, CH of main-chain), 5.76–5.87, 6.91–7.03 (m, =CH of terminal crotonate) (Figure 1). From the integrated ratio of the terminal methine signals at δ 4.11–4.18 and 5.76–5.87 to the main-chain methine signals at δ 5.18–5.36, the MW value was calculated to be ~530.
3.3. Preparation of Amylose–ORHB Inclusion Complex Using G3 as Primer
A mixture of G-1-P (0.204 g, 0.786 mmol), G3 (5.5 mg, 0.011 mmol), and ORHB (MW = 530, 0.159 g) in sodium acetate buffer (pH = 6.2, 0.2 mol/L, 5.0 mL) was ultrasonicated for 3 h to obtain a dispersion. After the addition of thermostable GP (14.4 U), the mixture was stirred at 70 °C for 48 h. The reaction mixture was then cooled to 45 °C over 7 h with stirring. The water-insoluble fraction was isolated by centrifugation, which was washed with water (25 mL), acetone (5.0 mL), and chloroform (30 mL), and dried under reduced pressure to give the inclusion complex (12.6 mg). 1H-NMR (DMSO-d6 + D2O) δ 1.19 (s, CH3 of ORHB), 2.53 (br, CH2 of ORHB, overlapping with DMSO), 3.31–3.92 (m, H2-H6 of amylose, overlapping with HOD), 4.98–5.18 (br, H1 of amylose, CH of ORHB).
3.4. Interaction between Single Amylose and ORHB in Thermostable Phosphorylase-Catalyzed Enzymatic Polymerization Using G3 as Primer
A mixture of G-1-P (0.207 g, 0.796 mmol), G3 (5.6 mg, 0.011 mmol), and ORHB (MW = 550, 0.106 g) in sodium acetate buffer (pH = 6.2, 0.2 mol/L, 4.0 mL) was ultrasonicated for 3 h to obtain a dispersion. After the addition of thermostable GP (14.4 U), the mixture was stirred at 70 °C for 48 h. The water-insoluble fraction was isolated by centrifugation, which was washed with water (30 mL) and chloroform (30 mL), and dried under reduced pressure to give the product (2.8 mg).
3.5. Preparation of Amylose–ORHB Inclusion Complex Using Water-Soluble Amylose as Primer
A mixture of G-1-P (0.129 g, 0.497 mmol), water-soluble amylose (MW = 2800, 13.9 mg, 5.0 μmol), and ORHB (MW = 530, 0.108 g) in sodium acetate buffer (pH = 6.2, 0.2 mol/L, 5.0 mL) was ultrasonicated for 3 h to obtain a dispersion. After the addition of thermostable GP (7.2 U), the mixture was stirred at 70 °C for 48 h. The reaction mixture was then cooled to 45 °C over 7 h with stirring. The water-insoluble fraction was isolated by centrifugation, which was washed with water (25 mL), acetone (5.0 mL), and chloroform (30 mL), and dried under reduced pressure to give the inclusion complex (23.4 mg). 1H-NMR (DMSO-d6 + D2O) δ 1.26 (s, CH3 of ORHB), 2.64 (br, CH2 of ORHB, overlapping with DMSO), 3.37–4.18 (m, H2-H6 of amylose, overlapping with HOD), 4.99–5.29 (br, H1 of amylose, CH of ORHB).
3.6. Measurements
Powder XRD measurements were conducted using a Rigaku Geigerflex RADIIB diffractometer (PANalytical B.V., EA Almelo, the Netherlands) with Ni-filtered CuKα radiation (λ = 0.15418 nm). 1H-NMR spectra were recorded on JEOL ECA 600 and ECX 400 spectrometers (JEOL, Akishima, Tokyo, Japan). IR spectra were recorded on a PerkinElmer Spectrum Two spectrometer (PerkinElmer Japan Co., Ltd., Yokohama, Japan).
4. Conclusions
When the thermostable GP-catalyzed enzymatic polymerization of G-1-P from the G3 primer was carried out in the presence of ORHB at 70 °C for 48 h and the mixture was subsequently cooled to 45 °C over 7 h, the amylose–ORHB inclusion complex was produced by vine-twining polymerization. On the other hand, the same system using the water-soluble amylose primer induced partial formation of the inclusion complex. The interaction between the single amylose obtained by the thermostable GP-catalyzed enzymatic polymerization at an elevated temperature and ORHB was speculated to be key for the formation of the inclusion complex in this study. As the two components in the inclusion complex are biodegradable polymers, we are going to investigate its biodegradability and biocompatibility. In the future, other biosynthesized polyesters, a family of PRHB, will be employed as guest polymers in the present vine-twining polymerization system to produce the corresponding inclusion complexes with different structures.
Author Contributions
J.-i.K. and K.Y. contributed to the methodology and writing of the paper. Y.W. performed the experiments and analysis. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We acknowledge the supply of thermostable α-glucan phosphorylase from Ezaki Glico Co. Ltd., Osaka, Japan.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Not applicable.
References
- Schuerch, C. Polysaccharides. In Encyclopedia of Polymer Science and Engineering, 2nd ed.; Mark, H.F., Bilkales, N., Overberger, C.G., Eds.; John Wiley & Sons: New York, NY, USA, 1986; Volume 13, pp. 87–162. [Google Scholar]
- Rostamabadi, H.; Falsafi, S.R.; Jafari, S.M. Nano-helices of amylose for encapsulation of food ingredients. In Biopolymer Nanostructures for Food Encapsulation Purposes; Elsevier: Amsterdam, The Netherlands, 2019; pp. 463–491. [Google Scholar]
- Seung, D. Amylose in starch: Towards an understanding of biosynthesis, structure and function. New Phytol. 2020, 228, 1490–1504. [Google Scholar] [CrossRef] [PubMed]
- Putseys, J.A.; Lamberts, L.; Delcour, J.A. Amylose-inclusion complexes: Formation, identity and physico-chemical properties. J. Cereal Sci. 2010, 51, 238–247. [Google Scholar] [CrossRef]
- Ziegast, G.; Pfannemuller, B. Linear and star-shaped hybrid polymers 4. Phosphorolytic syntheses with di-functional, oligo-functional and multifunctional primers. Carbohydr. Res. 1987, 160, 185–204. [Google Scholar] [CrossRef]
- Fujii, K.; Takata, H.; Yanase, M.; Terada, Y.; Ohdan, K.; Takaha, T.; Okada, S.; Kuriki, T. Bioengineering and application of novel glucose polymers. Biocatal. Biotransform. 2003, 21, 167–172. [Google Scholar] [CrossRef]
- Yanase, M.; Takaha, T.; Kuriki, T. α-Glucan phosphorylase and its use in carbohydrate engineering. J. Sci. Food Agric. 2006, 86, 1631–1635. [Google Scholar] [CrossRef]
- Shoda, S.; Uyama, H.; Kadokawa, J.; Kimura, S.; Kobayashi, S. Enzymes as green catalysts for precision macromolecular synthesis. Chem. Rev. 2016, 116, 2307–2413. [Google Scholar] [CrossRef] [PubMed]
- Kadokawa, J. Enzymatic synthesis of functional amylosic materials and amylose analog polysaccharides. Methods Enzymol. 2019, 627, 189–213. [Google Scholar]
- Awad, F.N. Glycoside phosphorylases for carbohydrate synthesis: An insight into the diversity and potentiality. Biocatal. Agric. Biotechnol. 2021, 31, 101886. [Google Scholar] [CrossRef]
- Imberty, A.; Chanzy, H.; Perez, S.; Buleon, A.; Tran, V. The double-helical nature of the crystalline part of A-starch. J. Mol. Biol. 1988, 201, 365–378. [Google Scholar] [CrossRef]
- Imberty, A.; Perez, S. A revisit to the three-dimensional structure of B-type starch. Biopolymers 1988, 27, 1205–1221. [Google Scholar] [CrossRef]
- Kadokawa, J.; Orio, S.; Yamamoto, K. Formation of microparticles from amylose-grafted poly(γ-glutamic acid) networks obtained by thermostable phosphorylase-catalyzed enzymatic polymerization. RSC Adv. 2019, 9, 16176–16182. [Google Scholar] [CrossRef] [Green Version]
- Kadokawa, J. Preparation and applications of amylose supramolecules by means of phosphorylase-catalyzed enzymatic polymerization. Polymers 2012, 4, 116–133. [Google Scholar] [CrossRef]
- Kadokawa, J. Architecture of amylose supramolecules in form of inclusion complexes by phosphorylase-catalyzed enzymatic polymerization. Biomolecules 2013, 3, 369–385. [Google Scholar] [CrossRef]
- Kadokawa, J. Precision polysaccharide synthesis catalyzed by enzymes. Chem. Rev. 2011, 111, 4308–4345. [Google Scholar] [CrossRef] [PubMed]
- Orio, S.; Yamamoto, K.; Kadokawa, J. Preparation and material application of amylose-polymer inclusion complexes by enzymatic polymerization approach. Polymers 2017, 9, 729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadokawa, J. Synthesis of amylosic supramolecular materials by glucan phosphorylase-catalyzed enzymatic polymerization according to the vine-twining approach. Synlett 2020, 31, 648–656. [Google Scholar] [CrossRef]
- Kadokawa, J.; Kaneko, Y.; Nagase, S.; Takahashi, T.; Tagaya, H. Vine-twining polymerization: Amylose twines around polyethers to form amylose–Polyether inclusion complexes. Chem. Eur. J. 2002, 8, 3321–3326. [Google Scholar] [CrossRef]
- Kaneko, Y.; Beppu, K.; Kadokawa, J. Preparation of amylose/polycarbonate inclusion complexes by means of vine-twining polymerization. Macromol. Chem. Phys. 2008, 209, 1037–1042. [Google Scholar] [CrossRef]
- Kadokawa, J.; Kaneko, Y.; Nakaya, A.; Tagaya, H. Formation of an amylose-polyester inclusion complex by means of phosphorylase-catalyzed enzymatic polymerization of α-D-glucose 1-phosphate monomer in the presence of poly(ε-caprolactone). Macromolecules 2001, 34, 6536–6538. [Google Scholar] [CrossRef]
- Kadokawa, J.; Nakaya, A.; Kaneko, Y.; Tagaya, H. Preparation of inclusion complexes between amylose and ester-containing polymers by means of vine-twining polymerization. Macromol. Chem. Phys. 2003, 204, 1451–1457. [Google Scholar] [CrossRef]
- Kaneko, Y.; Ueno, K.; Yui, T.; Nakahara, K.; Kadokawa, J. Amylose’s recognition of chirality in polylactides on formation of inclusion complexes in vine-twining polymerization. Macromol. Biosci. 2011, 11, 1407–1415. [Google Scholar] [CrossRef]
- Meereboer, K.W.; Misra, M.; Mohanty, A.K. Review of recent advances in the biodegradability of polyhydroxyalkanoate (PHA) bioplastics and their composites. Green Chem. 2020, 22, 5519–5558. [Google Scholar] [CrossRef]
- Kučera, F.; Petruš, J.; Jančář, J. The structure-hydrolysis relationship of poly(3-hydroxybutyrate). Polym. Test. 2019, 80, 106095. [Google Scholar] [CrossRef]
- Yamashita, Y. Single crystals of amylose V complexes. J. Polym. Sci. Part A 1965, 3, 3251–3260. [Google Scholar] [CrossRef] [PubMed]
- Obiro, W.C.; Ray, S.S.; Emmambux, M.N. V-Amylose structural characteristics, methods of preparation, significance, and potential applications. Food Rev. Int. 2012, 28, 412–438. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Tsutsui, A.; Tanaka, K.; Yamamoto, K.; Kadokawa, J. Evaluation of stability of amylose inclusion complexes depending on guest polymers and their application to supramolecular polymeric materials. Biomolecules 2017, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, W.; Greenwood, C.T.; Khan, K.M. The interaction of linear, amylose oligomers with iodine. Carbohydr. Res. 1971, 17, 25–33. [Google Scholar] [CrossRef]
- Kobayashi, K.; Kamiya, S.; Enomoto, N. Amylose-carrying styrene macromonomer and its homo- and copolymers: Synthesis via enzyme-catalyzed polymerization and complex formation with iodine. Macromolecules 1996, 29, 8670–8676. [Google Scholar] [CrossRef]
- Bhuiyan, S.H.; Rus’d, A.A.; Kitaoka, M.; Hayashi, K. Characterization of a hyperthermostable glycogen phosphorylase from Aquifex aeolicus expressed in Escherichia coli. J. Mol. Catal. B Enzym. 2003, 22, 173–180. [Google Scholar] [CrossRef]
- Yanase, M.; Takata, H.; Fujii, K.; Takaha, T.; Kuriki, T. Cumulative effect of amino acid replacements results in enhanced thermostability of potato type L α-glucan phosphorylase. Appl. Environ. Microbiol. 2005, 71, 5433–5439. [Google Scholar] [CrossRef] [Green Version]
Scheme 1.
Preparation of amylose–ORHB inclusion complex by vine-twining polymerization under conditions at controlled temperatures.
Scheme 1.
Preparation of amylose–ORHB inclusion complex by vine-twining polymerization under conditions at controlled temperatures.

Figure 1.
Terminal structures of ORHB.

Figure 2.
XRD profiles of (a) amylose, (b) amylose–PCL inclusion complex, (c) product obtained at 45 °C, (d) product obtained using G3 primer at elevated temperature, and (e) product obtained using water-soluble amylose primer at elevated temperature.
Figure 2.
XRD profiles of (a) amylose, (b) amylose–PCL inclusion complex, (c) product obtained at 45 °C, (d) product obtained using G3 primer at elevated temperature, and (e) product obtained using water-soluble amylose primer at elevated temperature.

Figure 3.
1H-NMR spectra of (a) ORHB in DMSO-d6, (b) amylose, (c) product obtained at 45 °C, (d) product obtained using G3 primer at elevated temperature, and (e) product obtained using water-soluble amylose primer at elevated temperature in DMSO-d6 + D2O.
Figure 3.
1H-NMR spectra of (a) ORHB in DMSO-d6, (b) amylose, (c) product obtained at 45 °C, (d) product obtained using G3 primer at elevated temperature, and (e) product obtained using water-soluble amylose primer at elevated temperature in DMSO-d6 + D2O.

Figure 4.
IR spectra of (a) ORHB, (b) amylose, (c) product obtained at 45 °C, (d) product obtained using G3 primer at elevated temperature, and (e) product obtained using water-soluble amylose primer at elevated temperature.
Figure 4.
IR spectra of (a) ORHB, (b) amylose, (c) product obtained at 45 °C, (d) product obtained using G3 primer at elevated temperature, and (e) product obtained using water-soluble amylose primer at elevated temperature.

Figure 5.
Plausible process for formation of amylose–ORHB inclusion complexes in thermostable GP-catalyzed enzymatic polymerization under conditions at controlled temperatures.
Figure 5.
Plausible process for formation of amylose–ORHB inclusion complexes in thermostable GP-catalyzed enzymatic polymerization under conditions at controlled temperatures.

Figure 6.
(a) 1H-NMR spectrum in DMSO-d6 + D2O and (b) IR spectrum of the interaction product between single amylose and ORHB.
Figure 6.
(a) 1H-NMR spectrum in DMSO-d6 + D2O and (b) IR spectrum of the interaction product between single amylose and ORHB.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kadokawa, J.-i.; Wada, Y.; Yamamoto, K. Preparation of Amylose-Oligo[(R)-3-hydroxybutyrate] Inclusion Complex by Vine-Twining Polymerization. Molecules 2021, 26, 2595. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092595
AMA Style
Kadokawa J-i, Wada Y, Yamamoto K. Preparation of Amylose-Oligo[(R)-3-hydroxybutyrate] Inclusion Complex by Vine-Twining Polymerization. Molecules. 2021; 26(9):2595. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092595
Chicago/Turabian StyleKadokawa, Jun-ichi, Yuki Wada, and Kazuya Yamamoto. 2021. "Preparation of Amylose-Oligo[(R)-3-hydroxybutyrate] Inclusion Complex by Vine-Twining Polymerization" Molecules 26, no. 9: 2595. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092595