
Protective Effects of Swertiamarin against Methylglyoxal-Induced Epithelial-Mesenchymal Transition by Improving Oxidative Stress in Rat Kidney Epithelial (NRK-52E) Cells



Abstract
:1. Introduction
2. Results
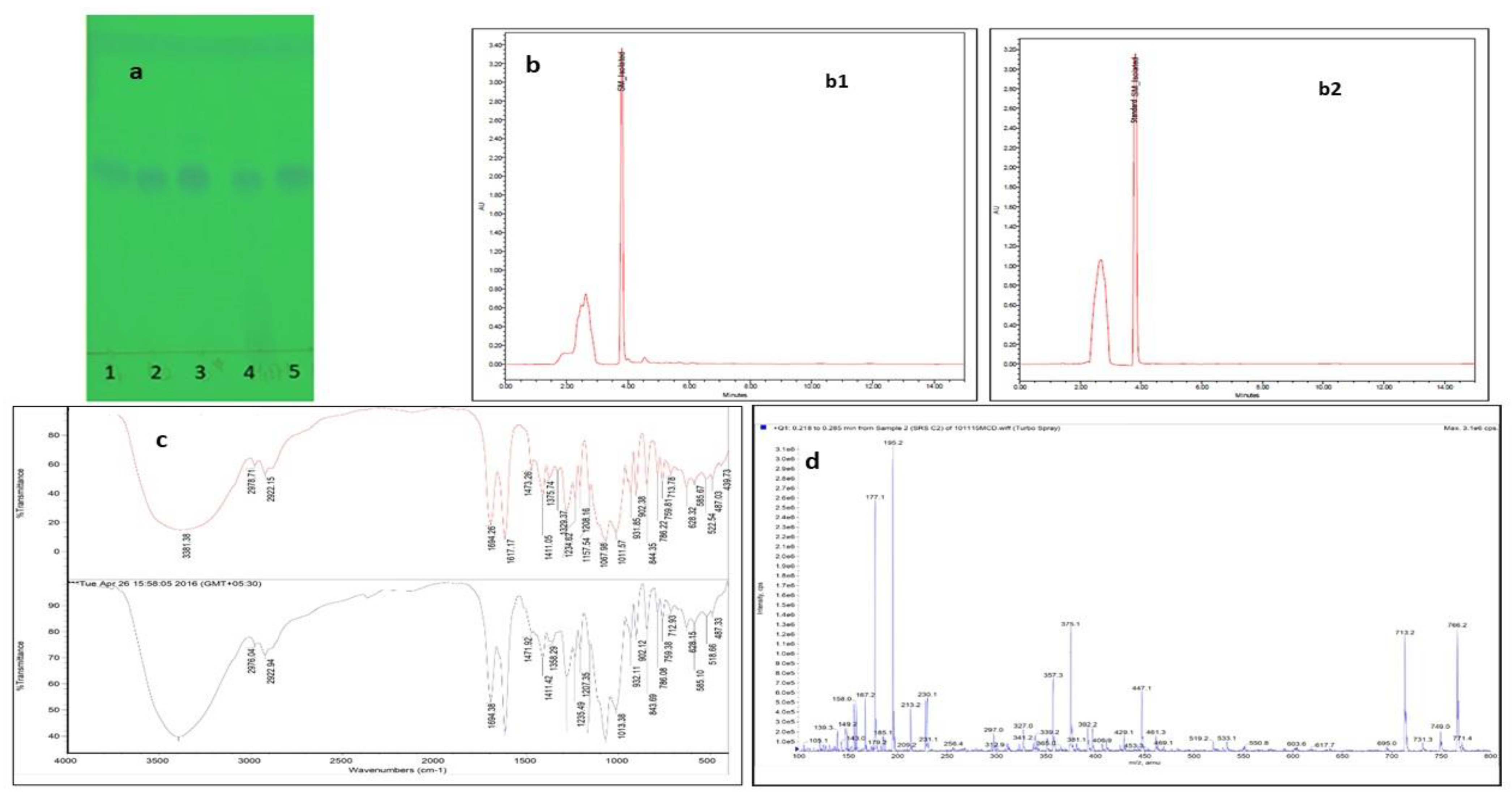
2.1. Isolation of SM from E. littorale and Characterization Using High-Performance Liquid Chromatography (HPLC), Fourier-Transform Infrared Spectroscopy (FTIR) and Liquid Chromatography-Mass Spectra(LC-MS)
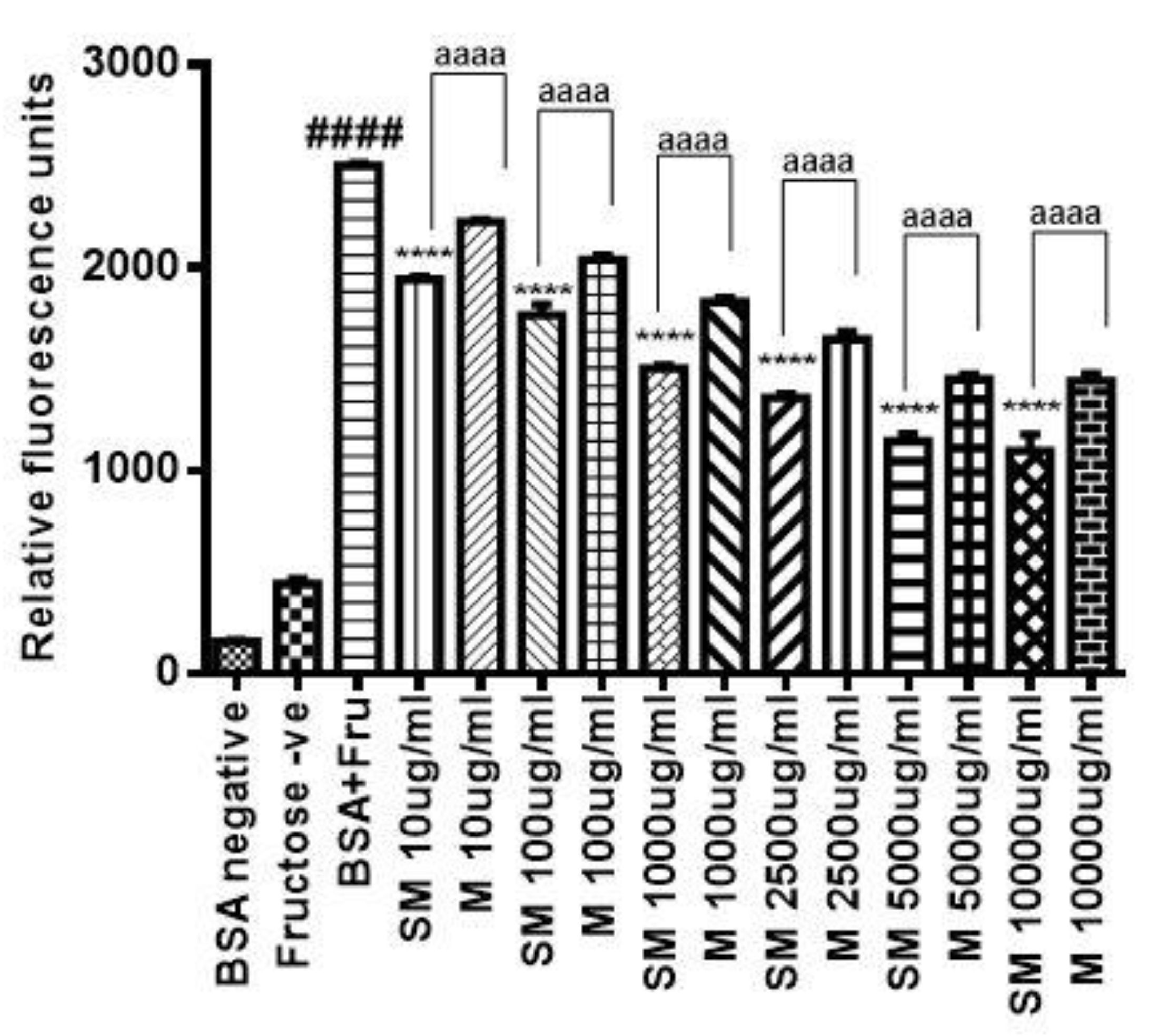
2.2. SM Inhibits the Formation of AGEs
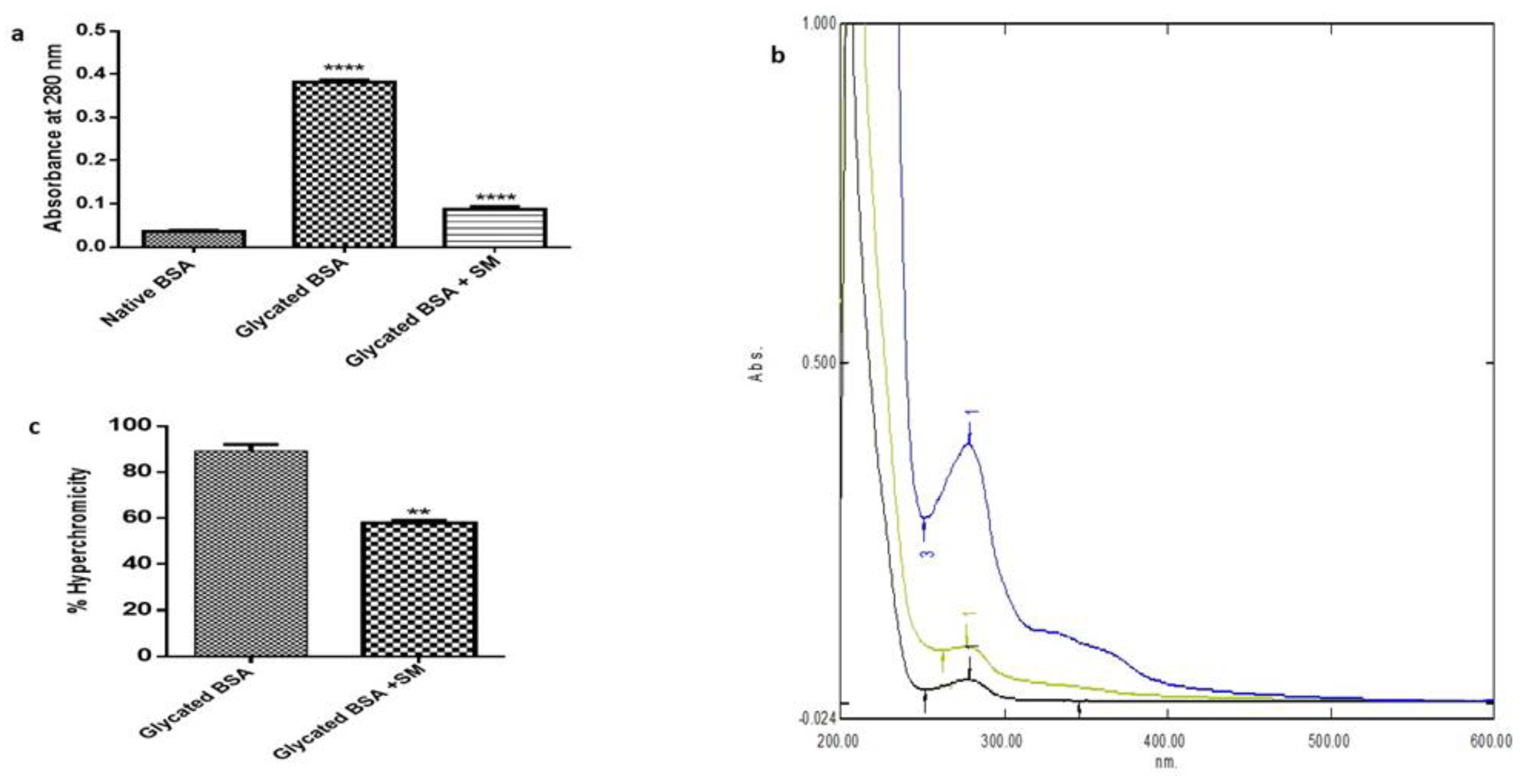
2.3. SM Reduces Fructose Mediated Hyperchromicity
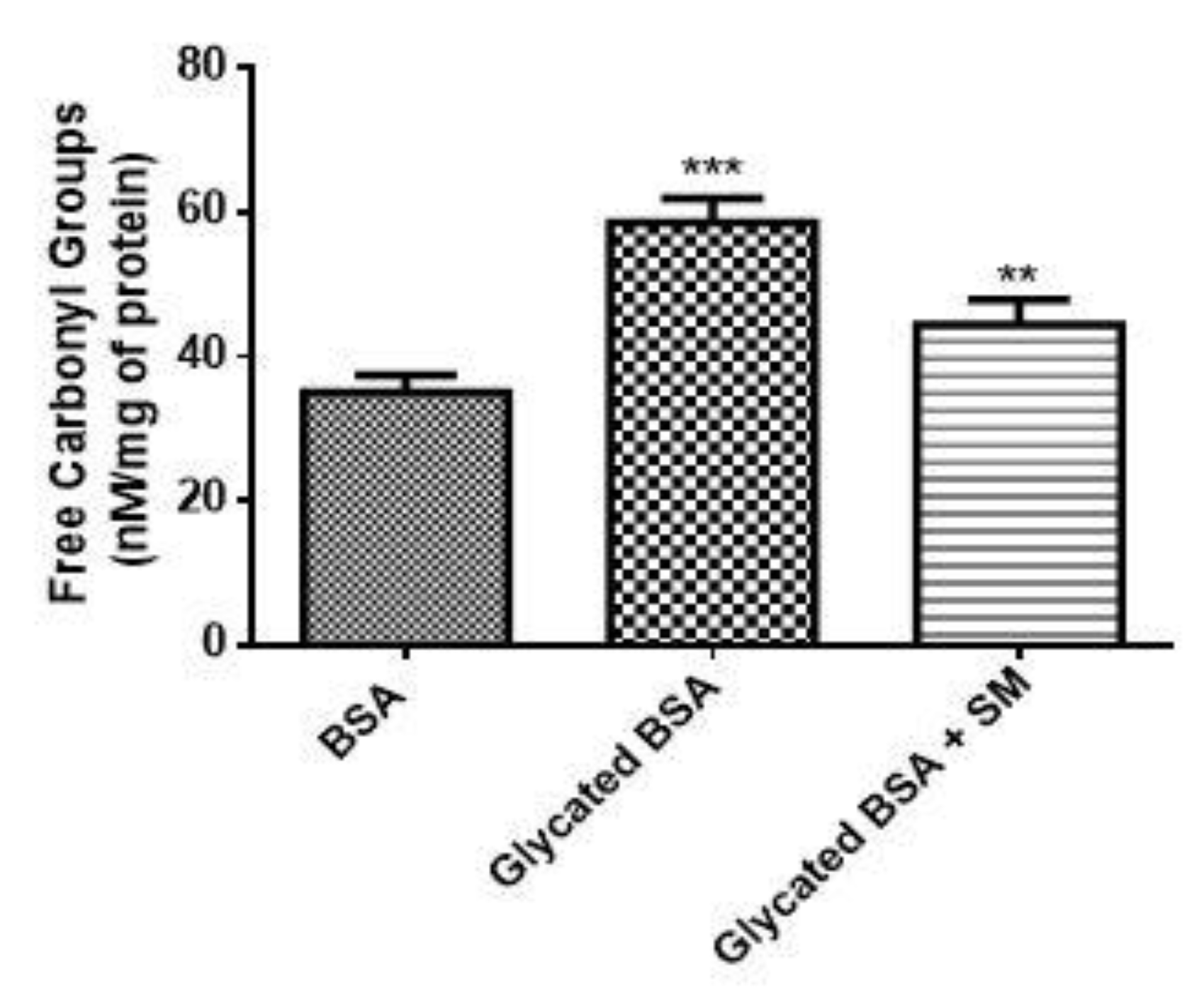
2.4. SM Reduces Carbonyl Content in the Glycated BSA
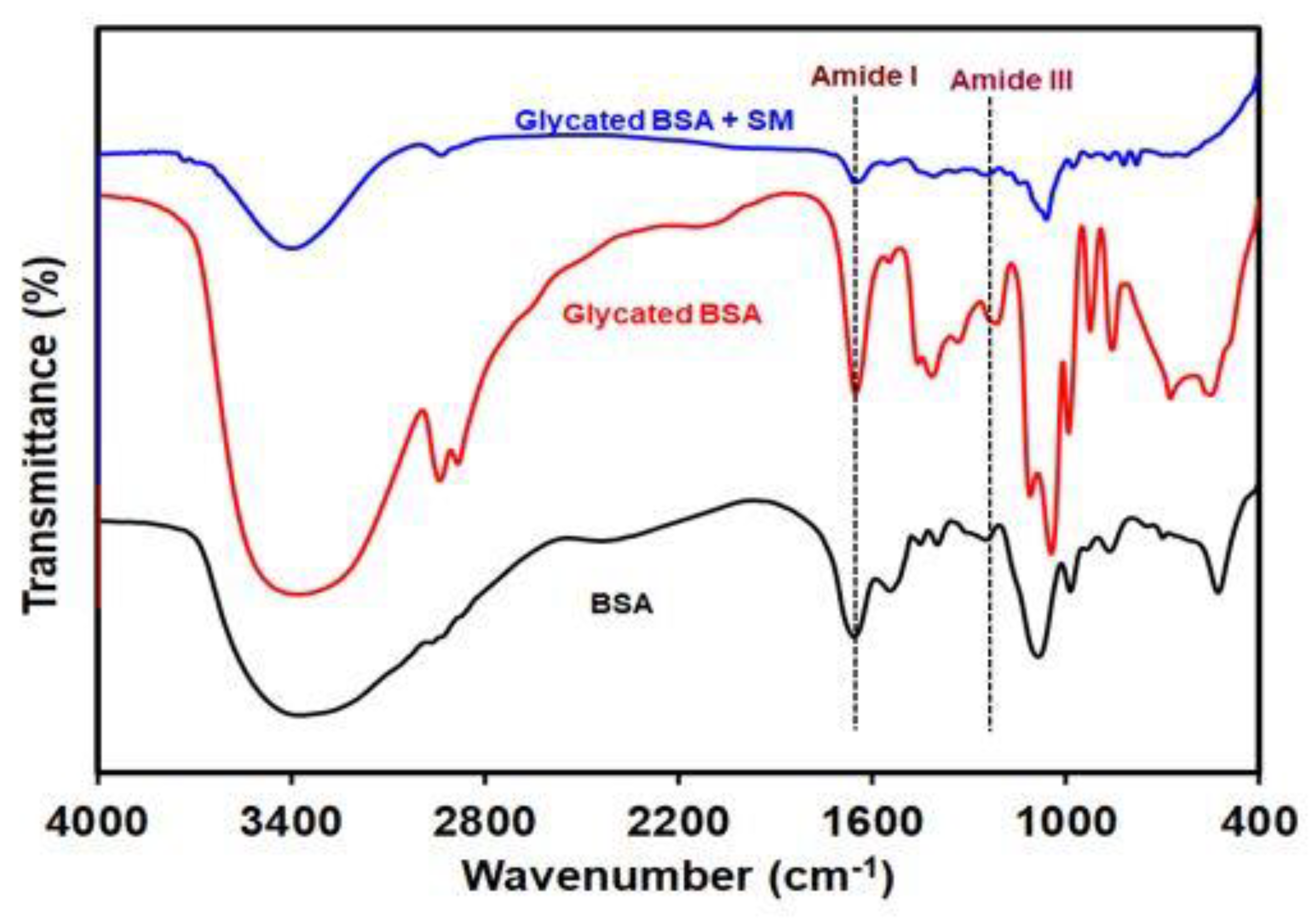
2.5. SM Prevents Fructose Mediated Side-Chain Modifications in BSA
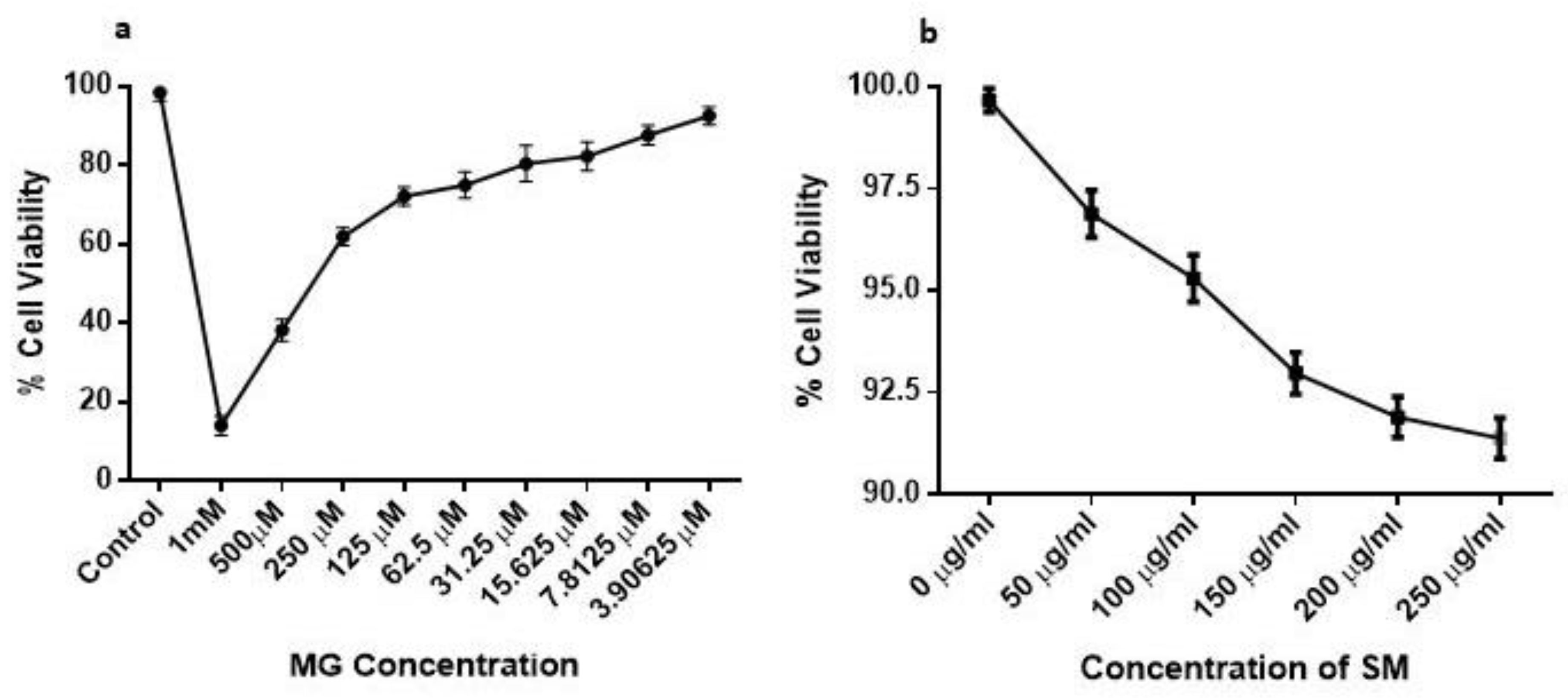
2.6. Effect of MG and SM on the Viability of NRK-52E Cells
2.7. SM Improves the Morphology of the NRK-52E Cells and Ameliorates against MG-Induced Damage
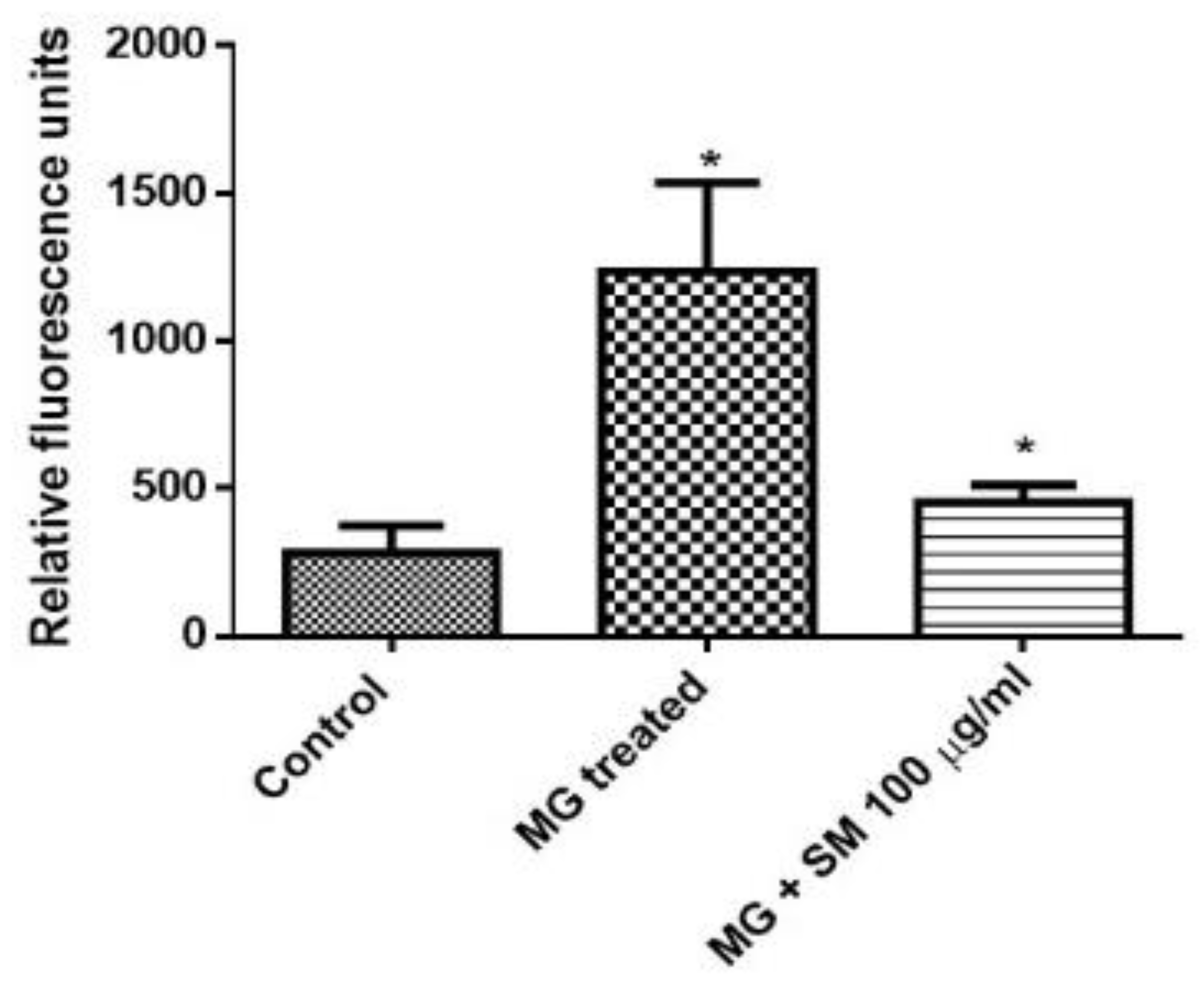
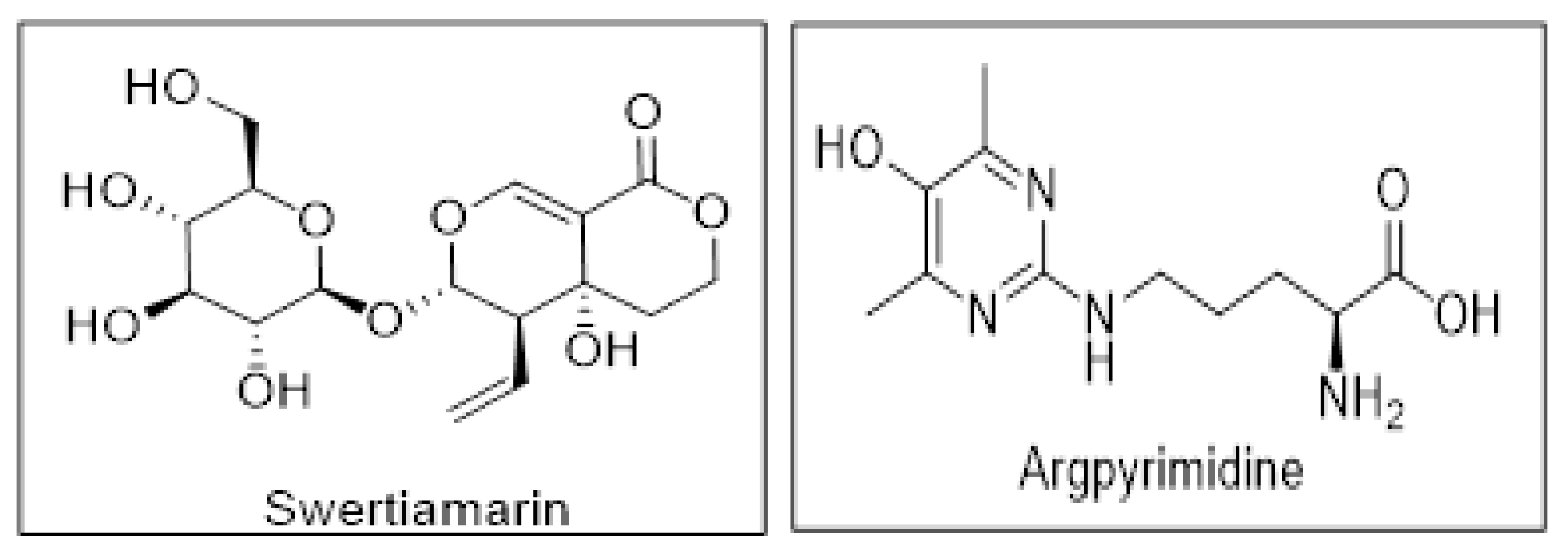
2.8. SM Alleviates the Formation of Argpyrimidine Like AGEs in NRK-52E Cells
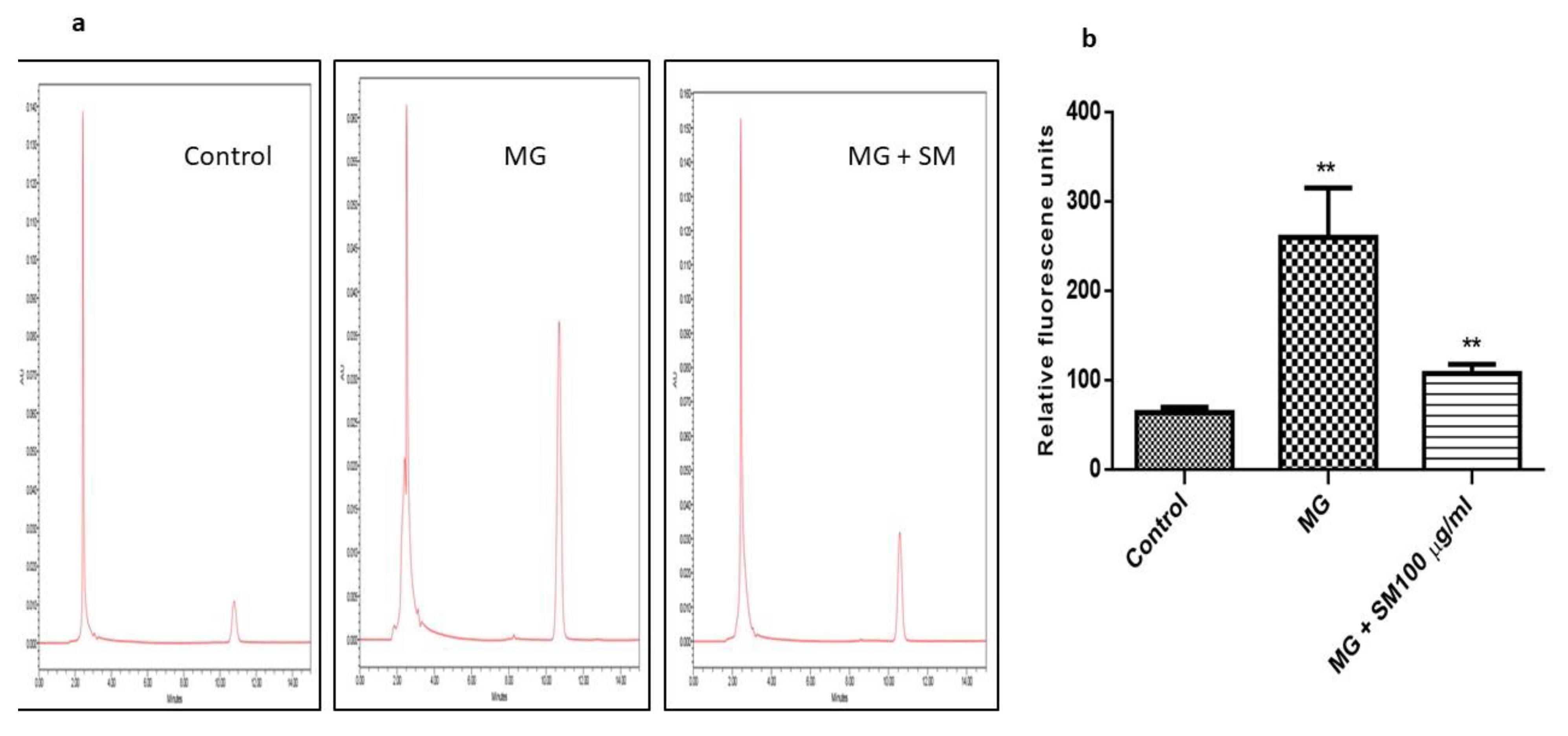
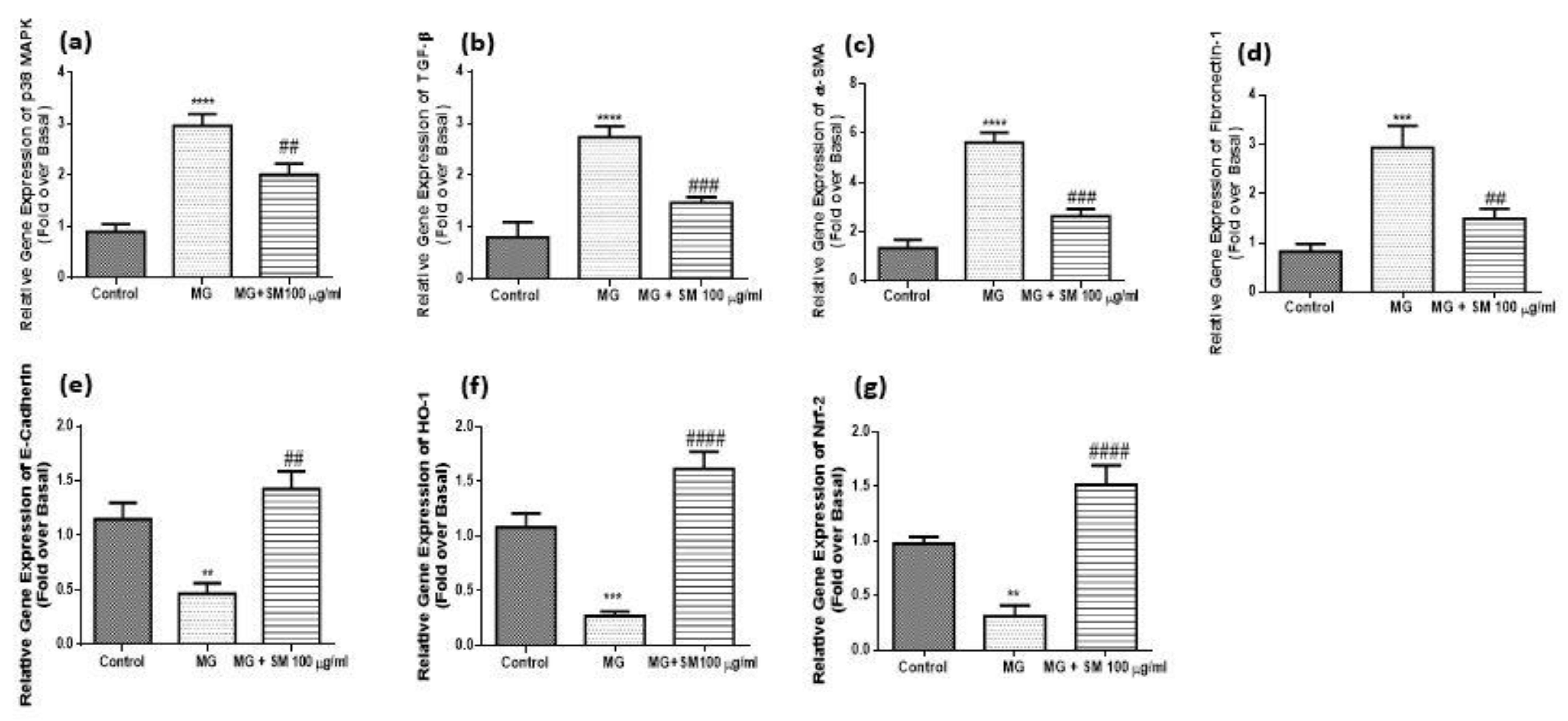
2.9. SM Alleviates the Formation of MG—Induced ROS and Mitigates the Oxidative Stress in NRK-52E Cells
2.10. SM Alleviates MG—Induced ER Stress in NRK-52E Cells
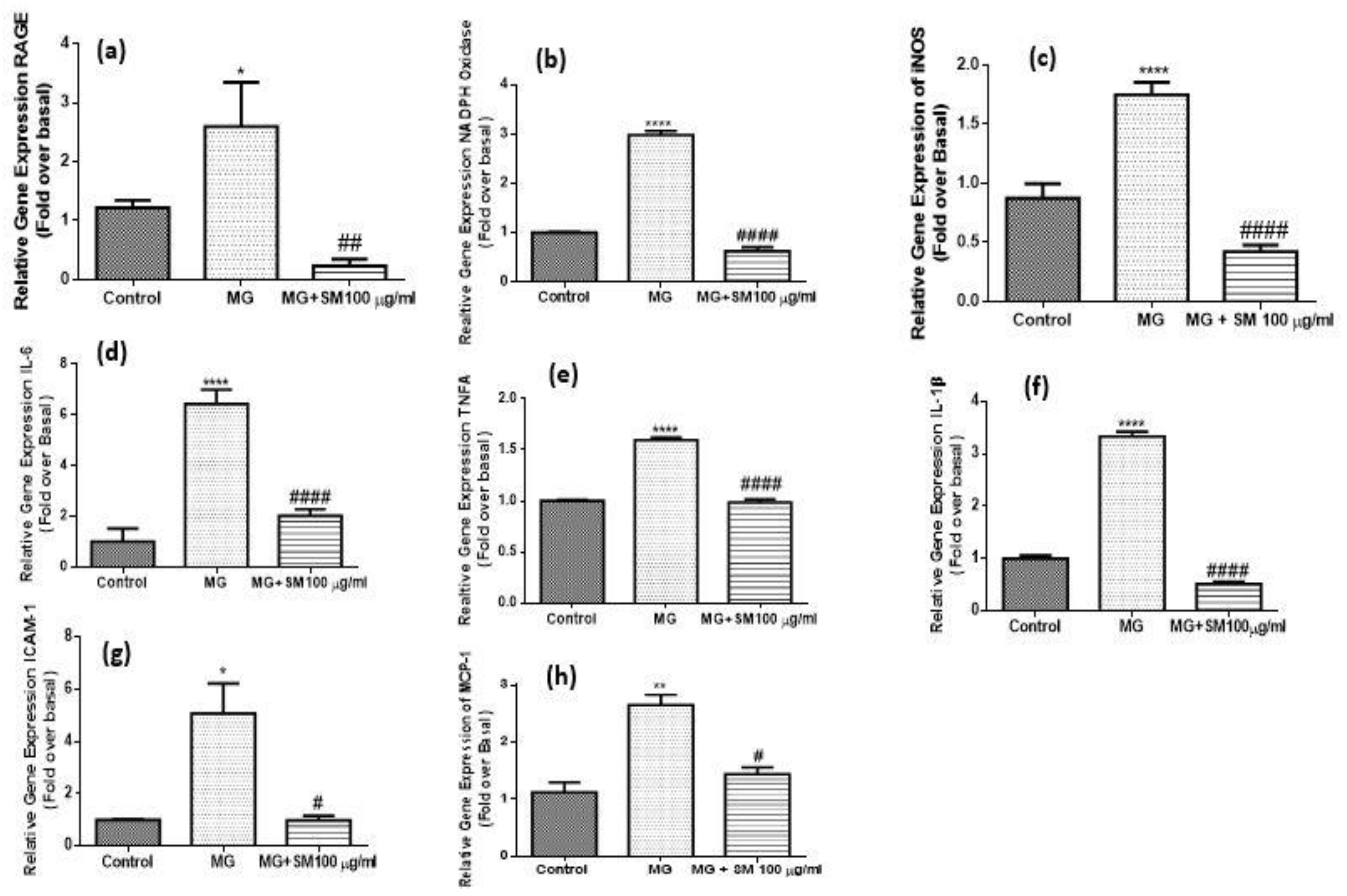
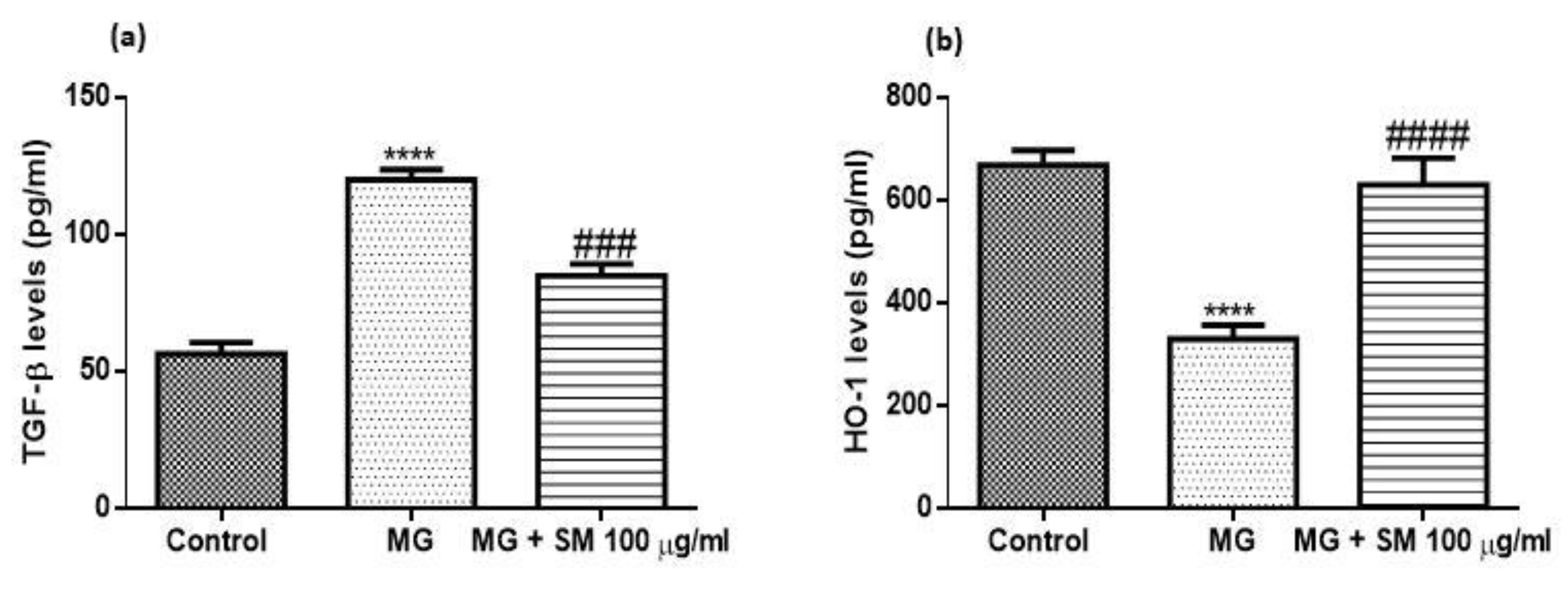
2.11. Treatment with SM Improves the Inflammatory Response in MG Treated NRK-52E Cells
2.12. SM Improves MG-Induced Epithelial Mesenchymal Transition (EMT) in NRK-52E Cells
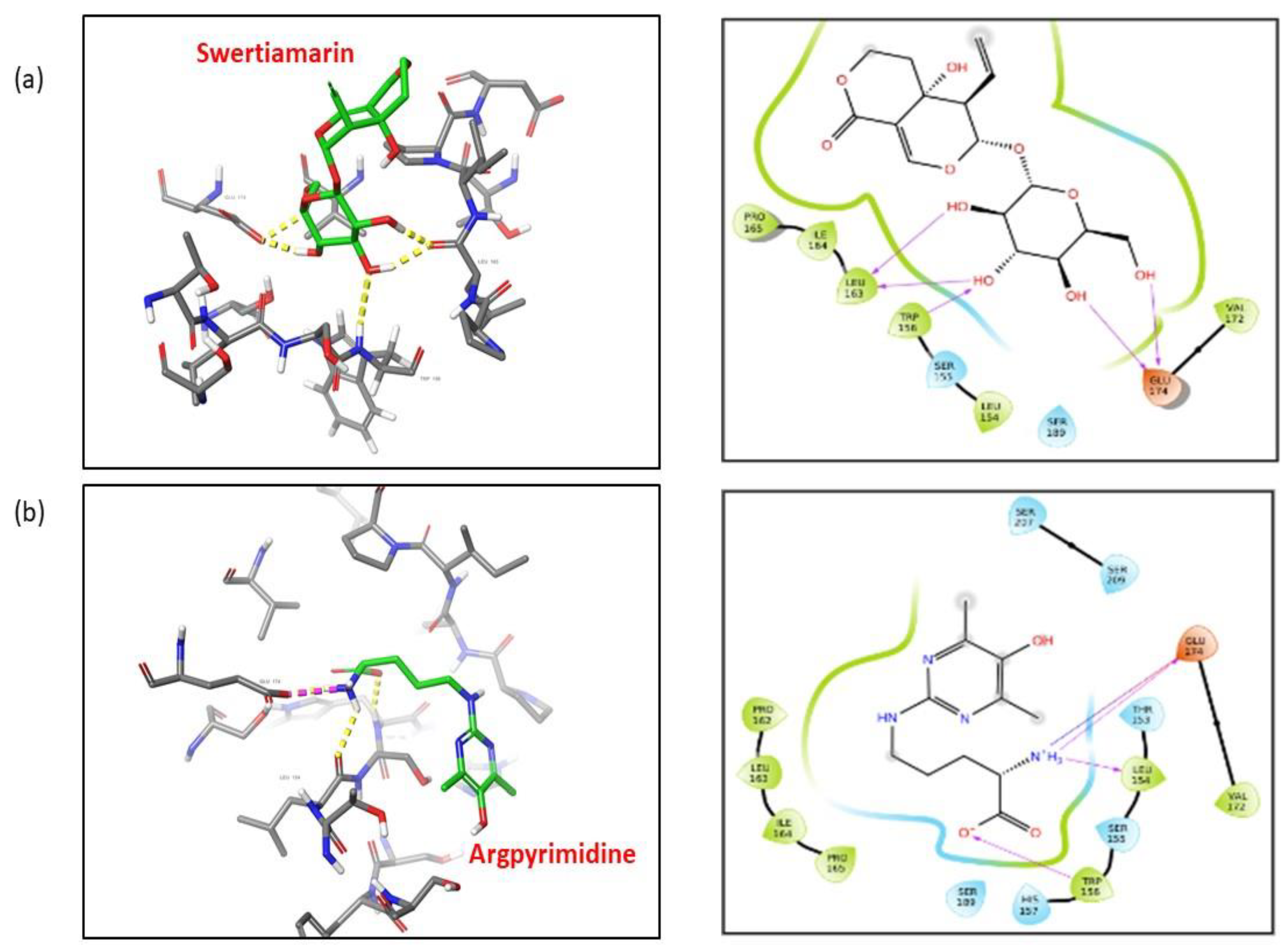
2.13. SM Shows Better Affinity and Binding with RAGE as Compared to Argpyrimidine
3. Discussion
4. Materials and Methods
4.1. Isolation of SM Using Silica Chromatography
4.2. Characterization of Isolated SM for Its Purity
4.2.1. Characterization Using High Performance Liquid Chromatography (HPLC)
4.2.2. Characterization Using Fourier-Transform Infrared Spectroscopy (FTIR) and Mass Spectrometer (MS)
4.3. Analysis of Antiglycative Potential of SM Using In Vitro Glycation Assay
4.4. Analysis of Protein Modifications Due to Glycation by UV Absorbance Spectroscopy
4.5. Estimation of Protein Carbonyl Content
4.6. Fourier Transform Infrared Spectroscopy of the Protein Samples
4.7. Culturing of NRK-52 E Cells
Cell Viability and Dose Determination of MG and SM Using MTT Assay
4.8. MG Stimulation and Different Treatment Groups
4.9. Estimation of Argpyrimidine
4.10. Estimation of Reactive Oxygen Species (ROS)
4.11. Estimation of MDA as a Measure of Lipid Peroxidation by HPLC
4.12. Isolation of RNA, cDNA Synthesis and Analysis of Gene Expressions of Various Markers by qRT-PCR
4.13. Estimation of Protein Levels of TGF-β and HO-1 by ELISA
4.14. Molecular Docking Studies
4.14.1. Homology Modelling
4.14.2. Protein Preparation Protocol
4.14.3. Receptor Grid Generation
4.14.4. Glide Molecular Docking and Analysis of Docking
4.14.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Writing Team for the Diabetes Control and Complications Trial/Epidemiology of Diabetes Interventions and Complications Research Group Sustained Effect of Intensive Treatment of Type 1 Diabetes Mellitus on Development and Progression of Diabetic Nephropathy. JAMA 2003, 290, 2159–2167. [CrossRef] [Green Version]
- Fioretto, P.; Steffes, M.W.; Sutherland, D.E.; Goetz, F.C.; Mauer, M. Reversal of Lesions of Diabetic Nephropathy after Pancreas Transplantation. N. Engl. J. Med. 1998, 339, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.; Steffes, M.W.; E Sutherland, D.; E Connett, J.; Rao, K.V.; Mauer, S.M. Effect of glycemic control on early diabetic renal lesions. A 5-year randomized controlled clinical trial of insulin-dependent diabetic kidney transplant recipients. JAMA 1994, 272, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Brownlee, M. Hyperglycemia-Induced Reactive Oxygen Species Increase Expression of the Receptor for Advanced Glycation End Products (RAGE) and RAGE Ligands. Diabetes 2009, 59, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neglia, C.; Cohen, H.J.; Garber, A.R.; Ellis, P.D.; Thorpe, S.R.; Baynes, J.W. 13C NMR investigation of nonenzymatic glucosylation of protein. Model studies using RNase A. J. Biol. Chem. 1983, 258, 14279–14283. [Google Scholar] [CrossRef]
- Lapolla, A.; Traldi, P.; Fedele, D. Importance of measuring products of non-enzymatic glycation of proteins. Clin. Biochem. 2005, 38, 103–115. [Google Scholar] [CrossRef]
- Kilhovd, B.; Giardino, I.; Torjesen, P.; Birkeland, K.; Berg, T.; Thornalley, P.; Brownlee, M.; Hanssen, K. Increased serum levels of the specific AGE-compound methylglyoxal-derived hydroimidazolone in patients with type 2 diabetes. Metabolism 2003, 52, 163–167. [Google Scholar] [CrossRef]
- Cooper, M.E. Interaction of metabolic and haemodynamic factors in mediating experimental diabetic nephropathy. Diabetologia 2001, 44, 1957–1972. [Google Scholar] [CrossRef]
- Pasupulati, A.K.; Chitra, P.S.; Reddy, G.B. Advanced glycation end products mediated cellular and molecular events in the pathology of diabetic nephropathy. Biomol. Concepts 2016, 7, 293–309. [Google Scholar] [CrossRef] [Green Version]
- Ceriello, A.; Bortolotti, N.; Falleti, E.; Taboga, C.; Tonutti, L.; Crescentini, A.; Motz, E.; Lizzio, S.; Russo, A.; Bartoli, E. Total Radical-Trapping Antioxidant Parameter in NIDDM Patients. Diabetes Care 1997, 20, 194–197. [Google Scholar] [CrossRef]
- Kang, J.H. Modification and inactivation of human Cu,Zn-superoxide dismutase by methylglyoxal. Mol. Cells 2003, 15, 194–199. [Google Scholar]
- Najjar, F.M.; Taghavi, F.; Ghadari, R.; Sheibani, N.; Moosavi-Movahedi, A.A. Destructive effect of non-enzymatic glycation on catalase and remediation via curcumin. Arch. Biochem. Biophys. 2017, 630, 81–90. [Google Scholar] [CrossRef]
- Choudhary, M.I.; Maher, S.; Begum, A.; Abbaskhan, A.; Ali, S.; Khan, A.; Rehman, S.-U.-; Rahman, A.-U. Characterization and Antiglycation Activity of Phenolic Constituents from Viscum album (European Mistletoe). Chem. Pharm. Bull. 2010, 58, 980–982. [Google Scholar] [CrossRef] [Green Version]
- Meng, G.; Zhu, H.; Yang, S.; Wu, F.; Zheng, H.; Chen, E.; Xu, J. Attenuating effects of Ganoderma lucidum polysaccharides on myocardial collagen cross-linking relates to advanced glycation end product and antioxidant enzymes in high-fat-diet and streptozotocin-induced diabetic rats. Carbohydr. Polym. 2011, 84, 180–185. [Google Scholar] [CrossRef]
- Sun, Z.; Peng, X.; Liu, J.; Fan, K.W.; Wang, M.; Chen, F. Inhibitory effects of microalgal extracts on the formation of advanced glycation endproducts (AGEs). Food Chem. 2010, 120, 261–267. [Google Scholar] [CrossRef]
- Yazdanparast, R.; Ardestani, A.; Jamshidi, S. Experimental diabetes treated with Achillea santolina: Effect on pancreatic oxidative parameters. J. Ethnopharmacol. 2007, 112, 13–18. [Google Scholar] [CrossRef]
- Murali, B.; Upadhyaya, U.; Goyal, R. Effect of chronic treatment with Enicostemma littorale in non-insulin-dependent diabetic (NIDDM) rats. J. Ethnopharmacol. 2002, 81, 199–204. [Google Scholar] [CrossRef]
- Yamahara, J.; Kobayashi, M.; Matsuda, H.; Aoki, S. Anticholinergic action of Swertia japonica and an active constituent. J. Ethnopharmacol. 1991, 33, 31–35. [Google Scholar] [CrossRef]
- Vaidya, H.; Rajani, M.; Sudarsanam, V.; Padh, H.; Goyal, R. Antihyperlipidaemic activity of swertiamarin, a secoiridoid glycoside in poloxamer-407-induced hyperlipidaemic rats. J. Nat. Med. 2009, 63, 437–442. [Google Scholar] [CrossRef]
- Vaidya, H.; Prajapati, A.; Rajani, M.; Sudarsanam, V.; Padh, H.; Goyal, R.K. Beneficial Effects of Swertiamarin on Dyslipidaemia in Streptozotocin-induced Type 2 Diabetic Rats. Phytotherapy Res. 2012, 26, 1259–1261. [Google Scholar] [CrossRef] [PubMed]
- Jaishree, V.; Badami, S. Antioxidant and hepatoprotective effect of swertiamarin from Enicostemma axillare against d-galactosamine induced acute liver damage in rats. J. Ethnopharmacol. 2010, 130, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Jaishree, V.; Narsimha, S. Swertiamarin and quercetin combination ameliorates hyperglycemia, hyperlipidemia and oxidative stress in streptozotocin-induced type 2 diabetes mellitus in wistar rats. Biomed. Pharmacother. 2020, 130, 110561. [Google Scholar] [CrossRef]
- Nakagawa, T.; Yokozawa, T.; Terasawa, K.; Shu, S.; Juneja, L.R. Protective Activity of Green Tea against Free Radical- and Glucose-Mediated Protein Damage. J. Agric. Food Chem. 2002, 50, 2418–2422. [Google Scholar] [CrossRef]
- Vishwakarma, S.; Rajani, M.; Bagul, M.; Goyal, R. A Rapid Method for the Isolation of Swertiamarin from Enicostemma littorale. Pharm. Biol. 2004, 42, 400–403. [Google Scholar] [CrossRef] [Green Version]
- Magora, H.B.; Rahman, M.; Gray, A.I.; Cole, M.D. Swertiamarin from Enicostemma axillare subsp. axillare (Gentianaceae). Biochem. Syst. Ecol. 2003, 31, 553–555. [Google Scholar] [CrossRef]
- Rana, V.S. Separation and Identification of Swertiamarin from Enicostema axillare Lam. Raynal by Centrifugal Partition Chromatography and Nuclear Magnetic Resonance-Mass Spectrometry. J. Pharm Sci. Emerg. Drugs 2014, 1, 2. [Google Scholar]
- Rana, V.S.; Dhanani, T.; Kumar, S. Improved and Rapid HPLC-PDA Method for Identification and Quantification of Swertiamarin in the Aerial Parts of Enicostemma Axillare. Malaysian J. Pharma Sci. 2012, 10, 1–10. [Google Scholar]
- Kumar, S.; Jairaj, V. An Effective Method for Isolation of Pure Swertiamarin from Enicostemma littorale Blume. Indo Glob. J. Pharm. Sci. 2018, 8, 01–08. [Google Scholar] [CrossRef]
- Wu, T.; Li, J.; Li, Y.; Song, H. Antioxidant and Hepatoprotective Effect of Swertiamarin on Carbon Tetrachloride-Induced Hepatotoxicity via the Nrf2/HO-1 Pathway. Cell. Physiol. Biochem. 2017, 41, 2242–2254. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxidative Med. Cell. Longev. 2016, 2016, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Fukami, K.; Yamagishi, S.-I.; Ueda, S.; Okuda, S. Role of AGEs in Diabetic Nephropathy. Curr. Pharm. Des. 2008, 14, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N. Advanced glycation endproducts—role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar] [CrossRef]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced Glycation End Products and Diabetic Complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunceford, N.; Gugliucci, A. Ilex paraguariensis extracts inhibit AGE formation more efficiently than green tea. Fitoter. 2005, 76, 419–427. [Google Scholar] [CrossRef]
- Lapolla, A.; Flamini, R.; Vedova, A.D.; Senesi, A.; Reitano, R.; Fedele, D.; Basso, E.; Seraglia, R.; Traldi, P. Glyoxal and Methylglyoxal Levels in Diabetic Patients: Quantitative Determination by a New GC/MS Method. Clin. Chem. Lab. Med. 2003, 41, 1166–1173. [Google Scholar] [CrossRef]
- Stinghen, A.E.; Massy, Z.A.; Vlassara, H.; Striker, G.E.; Boullier, A. Uremic Toxicity of Advanced Glycation End Products in CKD. J. Am. Soc. Nephrol. 2015, 27, 354–370. [Google Scholar] [CrossRef] [Green Version]
- Henning, C.; Liehr, K.; Girndt, M.; Ulrich, C.; Glomb, M.A. Extending the Spectrum of α-Dicarbonyl Compounds In Vivo. J. Biol. Chem. 2014, 289, 28676–28688. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yagiz, Y.; Buran, T.J.; Nunes, C.D.N.; Gu, L. Phytochemicals from berries and grapes inhibited the formation of advanced glycation end-products by scavenging reactive carbonyls. Food Res. Int. 2011, 44, 2666–2673. [Google Scholar] [CrossRef]
- Justino, A.B.; Franco, R.R.; Silva, H.C.G.; Saraiva, A.L.; Sousa, R.M.F.; Espindola, F.S. B procyanidins of Annona crassiflora fruit peel inhibited glycation, lipid peroxidation and protein-bound carbonyls, with protective effects on glycated catalase. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Ardestani, A.; Yazdanparast, R. Cyperus rotundus suppresses AGE formation and protein oxidation in a model of fructose-mediated protein glycoxidation. Int. J. Biol. Macromol. 2007, 41, 572–578. [Google Scholar] [CrossRef]
- Ahmad, S.; Shahab, U.; Baig, M.H.; Khan, M.S.; Khan, M.S.; Srivastava, A.K.; Saeed, M. Moinuddin Inhibitory Effect of Metformin and Pyridoxamine in the Formation of Early, Intermediate and Advanced Glycation End-Products. PLoS ONE 2013, 8, e72128. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Matsui, T.; Takeuchi, M.; Yamagishi, S. Metformin Inhibits Advanced Glycation End Products (AGEs)-induced Renal Tubular Cell Injury by Suppressing Reactive Oxygen Species Generation via Reducing Receptor for AGEs (RAGE) Expression. Horm. Metab. Res. 2012, 44, 891–895. [Google Scholar] [CrossRef]
- Meeprom, A.; Sompong, W.; Chan, C.B.; Adisakwattana, S. Isoferulic Acid, a New Anti-Glycation Agent, Inhibits Fructose- and Glucose-Mediated Protein Glycation In Vitro. Molecules 2013, 18, 6439–6454. [Google Scholar] [CrossRef] [Green Version]
- Jairajpuri, D.S.; Fatima, S.; Saleemuddin, M. Immunoglobulin glycation with fructose: A comparative study. Clin. Chim. Acta 2007, 378, 86–92. [Google Scholar] [CrossRef]
- Allarakha, S.; Ahmad, P.; Ishtikhar, M.; Zaheer, M.S.; Siddiqi, S.S.; Moinuddin; Ali, A. Fructosylation generates neo-epitopes on human serum albumin. IUBMB Life 2015, 67, 338–347. [Google Scholar] [CrossRef]
- Liu, J.; Xing, X.; Jing, H. Differentiation of glycated residue numbers on heat-induced structural changes of bovine serum albumin. J. Sci. Food Agric. 2018, 98, 2168–2175. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Harding, J.J. Glycation-induced inactivation and loss of antigenicity of catalase and superoxide dismutase. Biochem. J. 1997, 328, 599–605. [Google Scholar] [CrossRef] [Green Version]
- Ha, H.; Hwang, I.-A.; Park, J.H.; Lee, H.B. Role of reactive oxygen species in the pathogenesis of diabetic nephropathy. Diabetes Res. Clin. Pract. 2008, 82, S42–S45. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-F.; Miyata, S.; Hirota, Y.; Higo, S.; Miyazaki, H.; Fukunaga, M.; Hamada, Y.; Ueyama, S.; Muramoto, O.; Uriuhara, A.; et al. Methylglyoxal induces apoptosis through activation of p38 mitogen-activated protein kinase in rat mesangial cells. Kidney Int. 2003, 63, 947–957. [Google Scholar] [CrossRef] [Green Version]
- Oya, T.; Hattori, N.; Mizuno, Y.; Miyata, S.; Maeda, S.; Osawa, T.; Uchida, K. Methylglyoxal Modification of Protein. J. Biol. Chem. 1999, 274, 18492–18502. [Google Scholar] [CrossRef] [Green Version]
- Wilker, S.C.; Chellan, P.; Arnold, B.M.; Nagaraj, R.H. Chromatographic Quantification of Argpyrimidine, a Methylglyoxal-Derived Product in Tissue Proteins: Comparison with Pentosidine. Anal. Biochem. 2001, 290, 353–358. [Google Scholar] [CrossRef]
- Padival, A.K.; Crabb, J.W.; Nagaraj, R.H. Methylglyoxal modifies heat shock protein 27 in glomerular mesangial cells. FEBS Lett. 2003, 551, 113–118. [Google Scholar] [CrossRef]
- Niki, E. Lipid peroxidation products as oxidative stress biomarkers. BioFactors 2008, 34, 171–180. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 1–31. [Google Scholar] [CrossRef]
- Sompong, W.; Cheng, H.; Adisakwattana, S. Ferulic acid prevents methylglyoxal-induced protein glycation, DNA damage, and apoptosis in pancreatic β-cells. J. Physiol. Biochem. 2016, 73, 121–131. [Google Scholar] [CrossRef]
- Reiniger, N.; Lau, K.; McCalla, D.; Eby, B.; Cheng, B.; Lu, Y.; Qu, W.; Quadri, N.; Ananthakrishnan, R.; Furmansky, M.; et al. Deletion of the Receptor for Advanced Glycation End Products Reduces Glomerulosclerosis and Preserves Renal Function in the Diabetic OVE26 Mouse. Diabetes 2010, 59, 2043–2054. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.D.; Zheng, M.; Zhao, L.L.; Qian, Y.; Yao, X.M.; Ren, A.; Li, S.M.; Jing, C.Y. Intensive insulin therapy decreases urinary MCP-1 and ICAM-1 excretions in incipient diabetic nephropathy. Eur. J. Clin. Investig. 2009, 39, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.-I.; Matsui, T. Advanced Glycation end Products, Oxidative Stress and Diabetic Nephropathy. Oxidative Med. Cell. Longev. 2010, 3, 101–108. [Google Scholar] [CrossRef]
- Menon, M.C.; Ross, M.J. Epithelial-to-mesenchymal transition of tubular epithelial cells in renal fibrosis: A new twist on an old tale. Kidney Int. 2016, 89, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liang, D.; Guo, L.; Liang, W.; Jiang, Y.; Li, H.; Zhao, Y.; Lu, S.; Chi, Z.-H. Curcumin protects renal tubular epithelial cells from high glucose-induced epithelial-to-mesenchymal transition through Nrf2-mediated upregulation of heme oxygenase-1. Mol. Med. Rep. 2012, 12, 1347–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, W.C.; Twigg, S.M.; Forbes, J.M.; Pete, J.; Tikellis, C.; Thallas-Bonke, V.; Thomas, M.C.; Cooper, M.E.; Kantharidis, P. Connective Tissue Growth Factor Plays an Important Role in Advanced Glycation End Product–Induced Tubular Epithelial-to-Mesenchymal Transition: Implications for Diabetic Renal Disease. J. Am. Soc. Nephrol. 2006, 17, 2484–2494. [Google Scholar] [CrossRef] [Green Version]
- Lv, Z.-M.; Wang, Q.; Wan, Q.; Lin, J.-G.; Hu, M.-S.; Liu, Y.-X.; Wang, R. The Role of the p38 MAPK Signaling Pathway in High Glucose-Induced Epithelial-Mesenchymal Transition of Cultured Human Renal Tubular Epithelial Cells. PLoS ONE 2011, 6, e22806. [Google Scholar] [CrossRef]
- Lee, Y.J.; Han, H.J. Troglitazone ameliorates high glucose-induced EMT and dysfunction of SGLTs through PI3K/Akt, GSK-3β, Snail1, and β-catenin in renal proximal tubule cells. Am. J. Physiol. Physiol. 2010, 298, F1263–F1275. [Google Scholar] [CrossRef] [Green Version]
- Hung, T.-J.; Chen, W.-M.; Liu, S.-F.; Liao, T.-N.; Lee, T.-C.; Chuang, L.-Y.; Guh, J.-Y.; Hung, C.-Y.; Hung, Y.-J.; Chen, P.-Y.; et al. 20-Hydroxyecdysone attenuates TGF-β1-induced renal cellular fibrosis in proximal tubule cells. J. Diabetes Complicat. 2012, 26, 463–469. [Google Scholar] [CrossRef]
- Parwani, K.; Mandal, P. Role of advanced glycation end products and insulin resistance in diabetic nephropathy. Arch. Physiol. Biochem. 2020, 1–13. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, W.; Zhang, M.; Ma, Z.; Qiu, X.; Shen, M.; Yin, X. ROS induces epithelial-mesenchymal transition via the TGF-β1/PI3K/Akt/mTOR pathway in diabetic nephropathy. Exp. Ther. Med. 2018, 17, 835–846. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Huang, Z.; Lin, Y.; Zhang, Z.; Fang, D.; Zhang, D.D. The Protective Role of Nrf2 in Streptozotocin-Induced Diabetic Nephropathy. Diabetes 2010, 59, 850–860. [Google Scholar] [CrossRef] [Green Version]
- Kie, J.H.; Kapturczak, M.H.; Traylor, A.; Agarwal, A.; Hill-Kapturczak, N. Heme Oxygenase-1 Deficiency Promotes Epithelial-Mesenchymal Transition and Renal Fibrosis. J. Am. Soc. Nephrol. 2008, 19, 1681–1691. [Google Scholar] [CrossRef] [Green Version]
- Fatchiyah, F.; Hardiyanti, F.; Widodo, N. Selective Inhibition on RAGE-binding AGEs Required by Bioactive Peptide Alpha-S2 Case in Protein from Goat Ethawah Breed Milk: Study of Biological Modeling. Acta Inform. Medica 2015, 23, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Kshirsagar, P.R.; Pai, S.R.; Nimbalkar, M.S.; Gaikwad, N.B. RP-HPLC analysis of seco-iridoid glycoside swertiamarin from differentSwertiaspecies. Nat. Prod. Res. 2016, 30, 865–868. [Google Scholar] [CrossRef] [PubMed]
- McPherson, J.D.; Shilton, B.H.; Walton, D.J. Role of fructose in glycation and cross-linking of proteins. Biochemistry 1988, 27, 1901–1907. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Assay Guidance Manual; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004; pp. 296–302. [Google Scholar]
- Karbowski, M.; Kurono, C.; Wozniak, M.; Ostrowski, M.; Teranishi, M.; Nishizawa, Y.; Usukura, J.; Soji, T.; Wakabayashi, T. Free radical–induced megamitochondria formation and apoptosis. Free. Radic. Biol. Med. 1999, 26, 396–409. [Google Scholar] [CrossRef]
- Tukozkan, N.; Erdamar, H.; Seven, I. Measurement of total malondialdehyde in plasma and tissues by high-performance liquid chromatography and thiobarbituric acid assay. Firat Tip Dergisi. 2006, 11, 88–92. [Google Scholar]
- Kema, V.H.; Khan, I.; Jamal, R.; Vishwakarma, S.K.; Reddy, C.L.; Parwani, K.; Patel, F.; Patel, D.; Khan, A.A.; Mandal, P. Protective Effects of Diallyl Sulfide Against Ethanol-Induced Injury in Rat Adipose Tissue and Primary Human Adipocytes. Alcohol. Clin. Exp. Res. 2017, 41, 1078–1092. [Google Scholar] [CrossRef]
- Tsoporis, J.; Izhar, S.; Leong-Poi, H.; Desjardins, J.-F.; Huttunen, H.; Parker, T. S100B Interaction with the Receptor for Advanced Glycation End Products (RAGE). Circ. Res. 2010, 106, 93–101. [Google Scholar] [CrossRef]
- Apweiler, R. UniProt: The Universal Protein knowledgebase. Nucleic Acids Res. 2004, 32, 115–119. [Google Scholar] [CrossRef]
- Xu, D.; Young, J.H.; Krahn, J.M.; Song, D.; Corbett, K.D.; Chazin, W.J.; Pedersen, L.C.; Esko, J.D. Stable RAGE-Heparan Sulfate Complexes Are Essential for Signal Transduction. ACS Chem. Biol. 2013, 8, 1611–1620. [Google Scholar] [CrossRef] [Green Version]
- Schrödinger Release, Version 12.7.156; Maestro Schrödinger LLC: New York, NY, USA, 2021.
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Greenwood, J.R.; Calkins, D.; Sullivan, A.P.; Shelley, J.C. Towards the comprehensive, rapid, and accurate prediction of the favorable tautomeric states of drug-like molecules in aqueous solution. J. Comput. Mol. Des. 2010, 24, 591–604. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.P.; Pincus, D.L.; Rapp, C.S.; Day, T.J.F.; Honig, B.; Shaw, D.E.; Friesner, R.A. A hierarchical approach to all-atom protein loop prediction. Proteins: Struct. Funct. Bioinform. 2004, 55, 351–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Bands and Wavenumber (cm−1) | |
|---|---|---|
| Amide I | Amide III | |
| Native BSA | 1655 | 1245 |
| Glycated BSA | 1651 | 1236 |
| Glycated BSA + SM | 1654 | 1242 |
| Sr. No. | Interaction | Point Interaction | Donor Atom | Acceptor Atom | Type of Interaction | Bond Distance (Å) | Energy Binding (G Score) (kcal/mol) |
|---|---|---|---|---|---|---|---|
| 1 | RAGE: Swertiamarin | Trp 156–Swertiamarin | Trp 156: H | Swertiamarin: O | Hydrogen Bond | 2.62 | −4.7 |
| Leu 163–Swertiamarin | Swertiamarin: H | Leu 163: O | Hydrogen Bond | 1.90 | |||
| Leu 163–Swertiamarin | Swertiamarin: H | Leu 163: O | Hydrogen Bond | 1.90 | |||
| Glu 174–Swertiamarin | Swertiamarin: H | Glu 174: O | Hydrogen Bond | 2.23 | |||
| Glu 174–Swertiamarin | Swertiamarin: H | Glu 174: O | Hydrogen Bond | 1.75 | |||
| 2 | RAGE: Argpyrimidine | Leu 154–Argpyrimidine | Argpyrimidine: H | Leu 154: O | Hydrogen Bond | 2.21 | −2.75 |
| Trp 156–Argpyrimidine | Trp 156: H | Argpyrimidine: O | Hydrogen Bond | 2.38 | |||
| Glu 174–Argpyrimidine | Argpyrimidine: H | Glu 174: O | Hydrogen Bond | 1.82 | |||
| Glu 174–Argpyrimidine | Argpyrimidine: N | Glu 174: O | Electrostatic Interaction | 2.82 |
| Sr. No. | Name of the Gene | Sense Primer Sequence | Antisense Primer Sequence |
|---|---|---|---|
| 1. | 18s | 5′ACGGAAGGGCACCACCAGGA 3′ | 5′CACCACCACCCACGGAATCG 3′ |
| 2. | RAGE | 5′ GGTACTGGTTCTTGCTCT 3′ | 5′ATTCTAGCTTCTGGGTTG 3′ |
| 3. | TNF-α | 5′ CAAGGAGGAGAAGTTCCCAA 3′ | 5′CTCTGCTTGGTGGTTTGCTA 3′ |
| 4. | ICAM-1 | 5′CCCCACCTACATACATTCCTAC 3′ | 5′ACATTTTCTCCCAGGCATTC 3′ |
| 5. | NADPH oxidase | 5′ GGCATCCCTTTACTCTGACCT 3′ | 5′ TGCTGCTCGAATATGAATGG 3′ |
| 6. | IL-6 | 5′ GCCCTTCAGGAACAGCTATGA 3′ | 5′ TGTCAACAACATCAGTCCCAAGA 3′ |
| 7. | IL-1β | 5′ CCCTGCAGCTGGAGAGTGTGG 3′ | 5′ TGTGCTCTGCTTGAGAGGTGCT 3′ |
| 8. | TGF-β | 5′ TGCTTCAGCTCCACAGAGAA 3′ | 5′ TGTGTTGGTTGTAGAGGGCA 3′ |
| 9. | iNOS | 5′ TCACTGGGACAGCACAGAAT 3′ | 5′ TGTGTCTGCAGATGTGCTGA 3′ |
| 10. | Fibronectin | 5′ CATGGCTTTAGGCGAACCA 3′ | 5′ CATCTACATTCGGCAGGTATGG 3′ |
| 11. | α-SMA | 5′ GACCCTGAAGTATCCGATAGAACA 3′ | 5′CACGCGAAGCTCGTTATAGAAG 3′ |
| 12. | E-cadherin | 5′ TGATGATGCCCCCAACACTC 3′ | 5′ CCAAGCCCTTGGCTGTTTTC 3′ |
| 13. | p38 MAPK | 5′ CGAAATGACCGGCTACGTGG 3′ | 5′ CACTTCATCGTAGGTCAGGC 3′ |
| 14. | Grp78 | 5′ GAAACTGCCGAGGCGTAT 3′ | 5′ ATGTTCTTCTCTCCCTCTCTCTTT 3′ |
| 15. | CHOP | 5′ GAAAGCAGAAACCGGTCCAAT 3′ | 5′ GGATGAGATATAGGTGCCCCC 3′ |
| 16. | MCP-1 | 5′ CCTCCACCACTATGCAGGTCTC 3′ | 5′ GCACGTGGATGCTACAGGC 3′ |
| 17. | Nrf-2 | 5′ CAGAGTTTCTTCGCCAGAGG 3′ | 5′ TGAGTGTGAGGACCCATCG 3′ |
| 18. | HO-1 | 5′ CAAATCCCACCTTGAACACA 3′ | 5′ CGACTGACTAATGGCAGCAG 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parwani, K.; Patel, F.; Patel, D.; Mandal, P. Protective Effects of Swertiamarin against Methylglyoxal-Induced Epithelial-Mesenchymal Transition by Improving Oxidative Stress in Rat Kidney Epithelial (NRK-52E) Cells. Molecules 2021, 26, 2748. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092748
Parwani K, Patel F, Patel D, Mandal P. Protective Effects of Swertiamarin against Methylglyoxal-Induced Epithelial-Mesenchymal Transition by Improving Oxidative Stress in Rat Kidney Epithelial (NRK-52E) Cells. Molecules. 2021; 26(9):2748. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092748
Chicago/Turabian StyleParwani, Kirti, Farhin Patel, Dhara Patel, and Palash Mandal. 2021. "Protective Effects of Swertiamarin against Methylglyoxal-Induced Epithelial-Mesenchymal Transition by Improving Oxidative Stress in Rat Kidney Epithelial (NRK-52E) Cells" Molecules 26, no. 9: 2748. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092748