
Structural Refolding and Thermal Stability of Myoglobin in the Presence of Mixture of Crowders: Importance of Various Interactions for Protein Stabilization in Crowded Conditions

 ,
,  ,
,  , ,
, , 
Abstract
:
1. Introduction
2. Results
2.1. Effect of Mixture of Crowders (PEG 400 + EG) on the Structure and Stability of Mb Using Various Spectroscopic Techniques
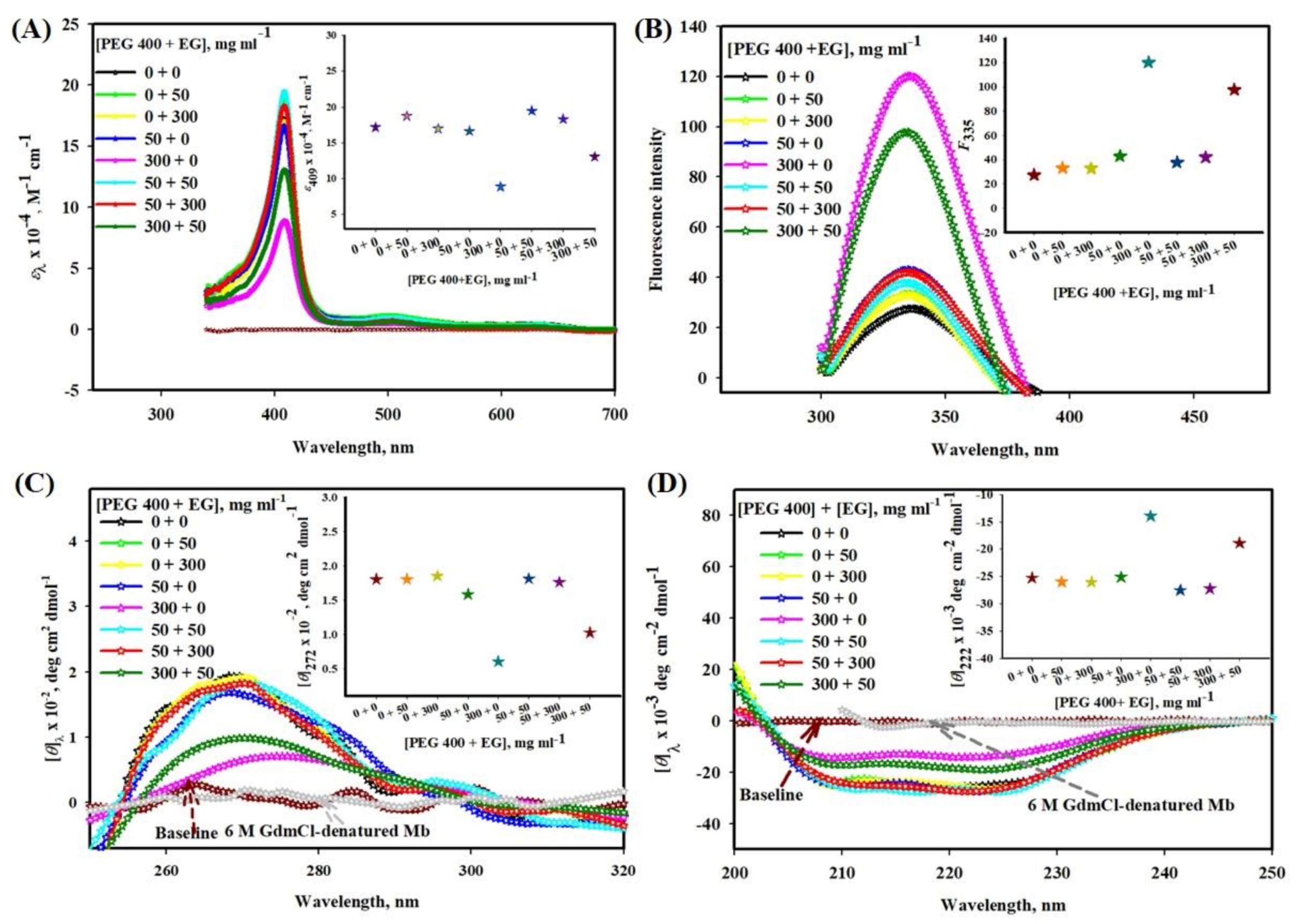
2.1.1. Influence of PEG 400-EG Mixtures on Absorption Spectra of Mb
2.1.2. Influence of PEG 400-EG Mixtures on Mb fluorescence
2.1.3. Conformational Studies of Mb in the Presence of PEG 400-EG Mixture and 6 M Guanidinium Chloride (GdmCl) Using Near- and Far-UV CD Spectra
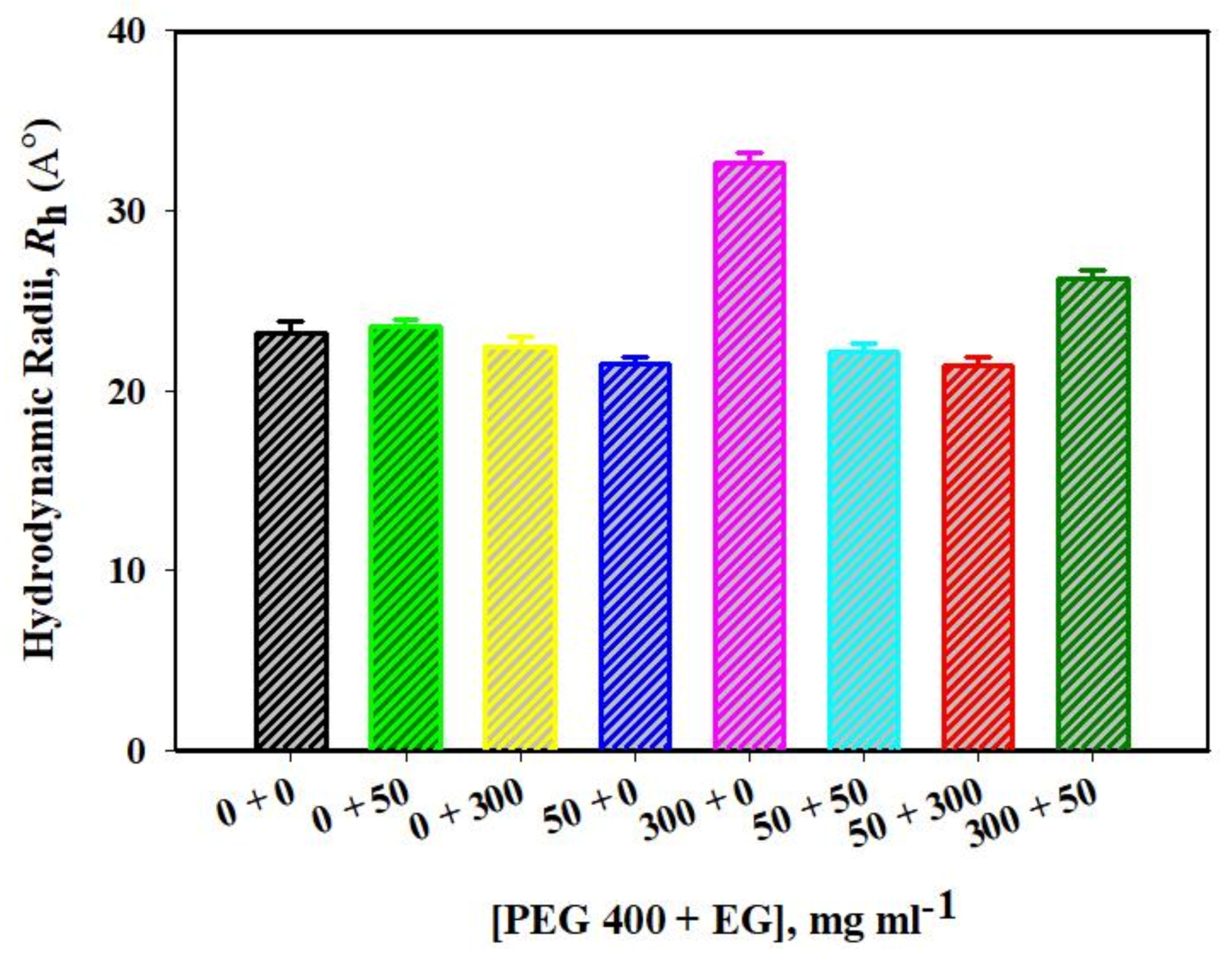
2.1.4. DLS Measurements of Mb in the Absence and Presence of EG, PEG 400 and Their Mixtures
2.2. Binding Studies of Mixture of Crowder Molecules (PEG + EG) with the Protein (Mb)
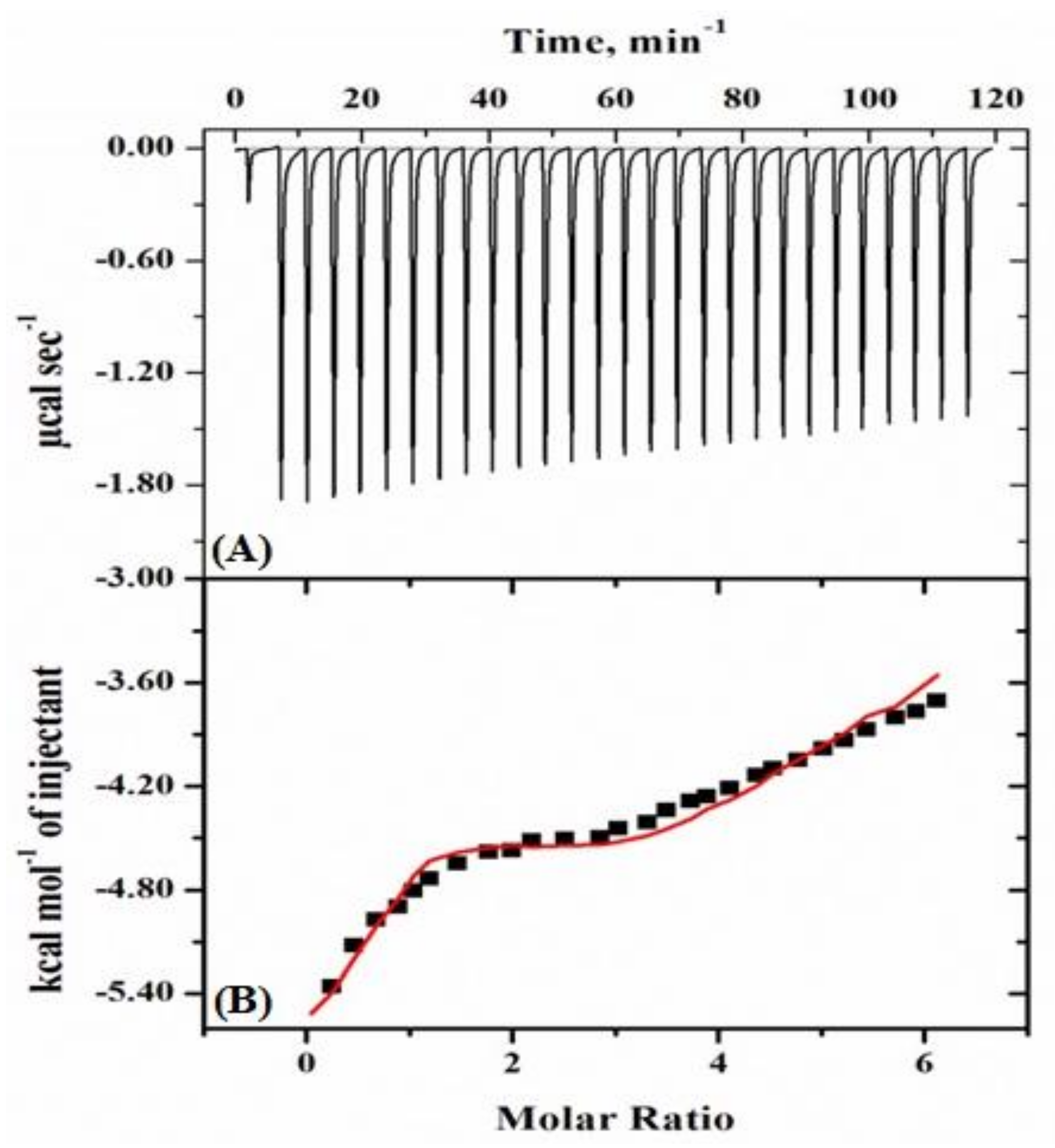
2.2.1. Isothermal Titration Calorimetry (ITC) Studies
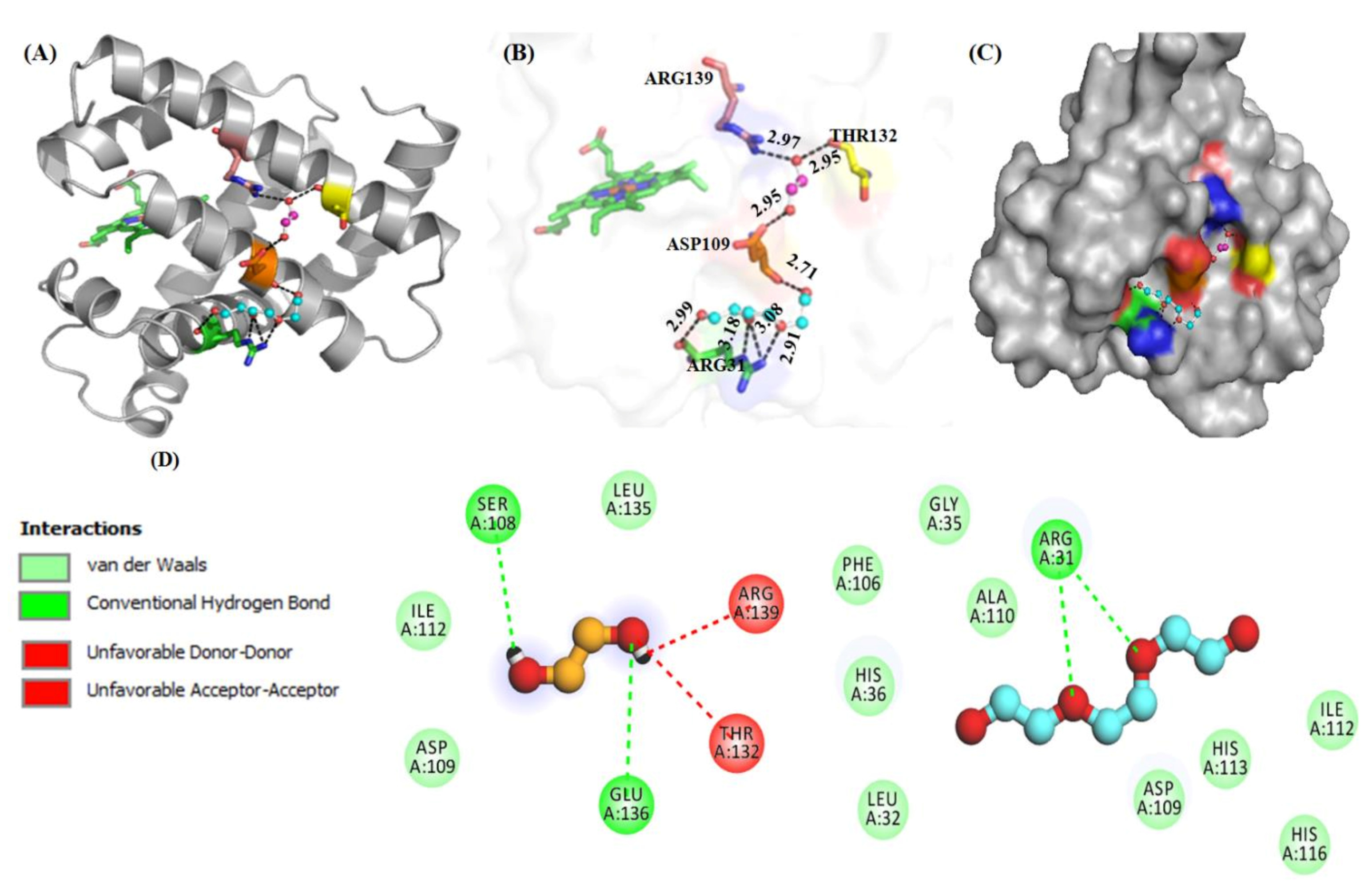
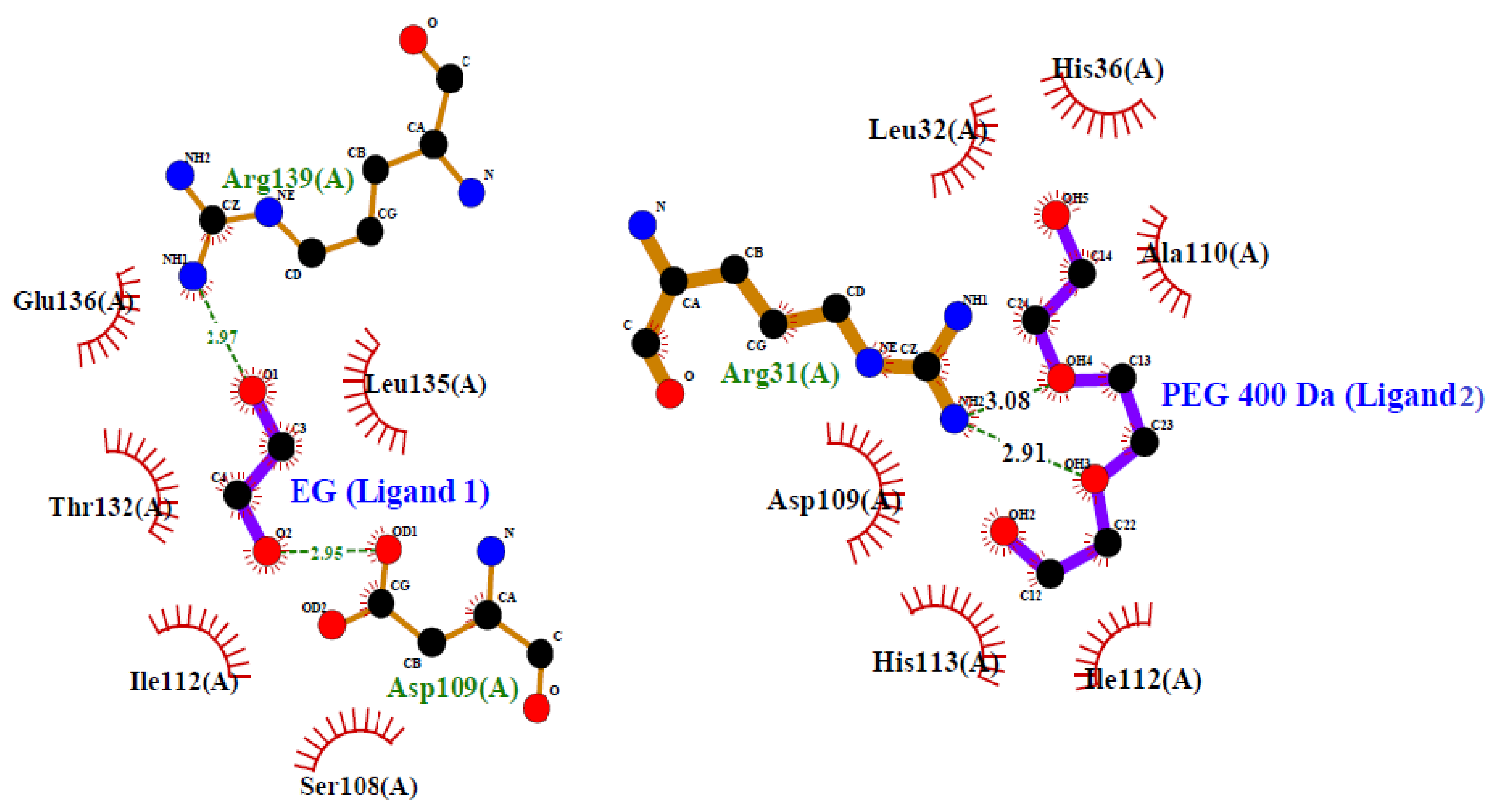
2.2.2. Molecular Docking Studies
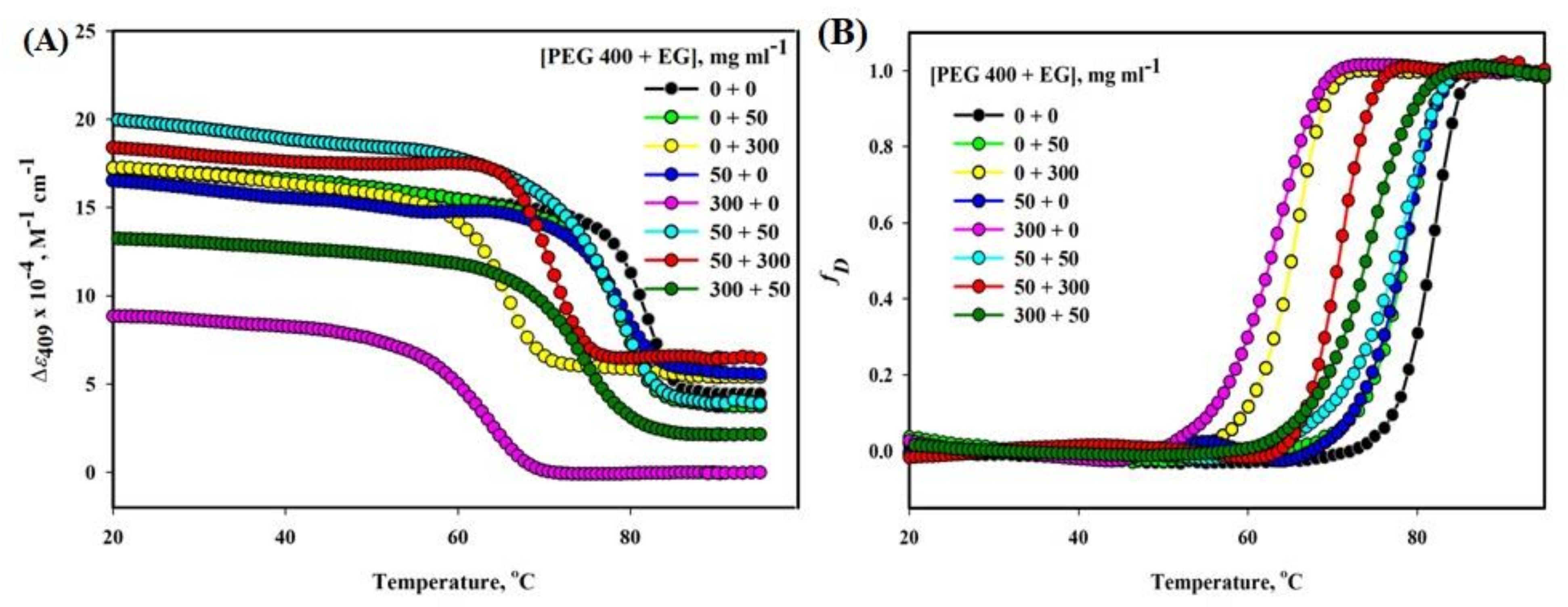
2.3. Heme-Mb Interaction Thermal Stability in the Presence of Various Concentration Mixtures of PEG 400 and EG
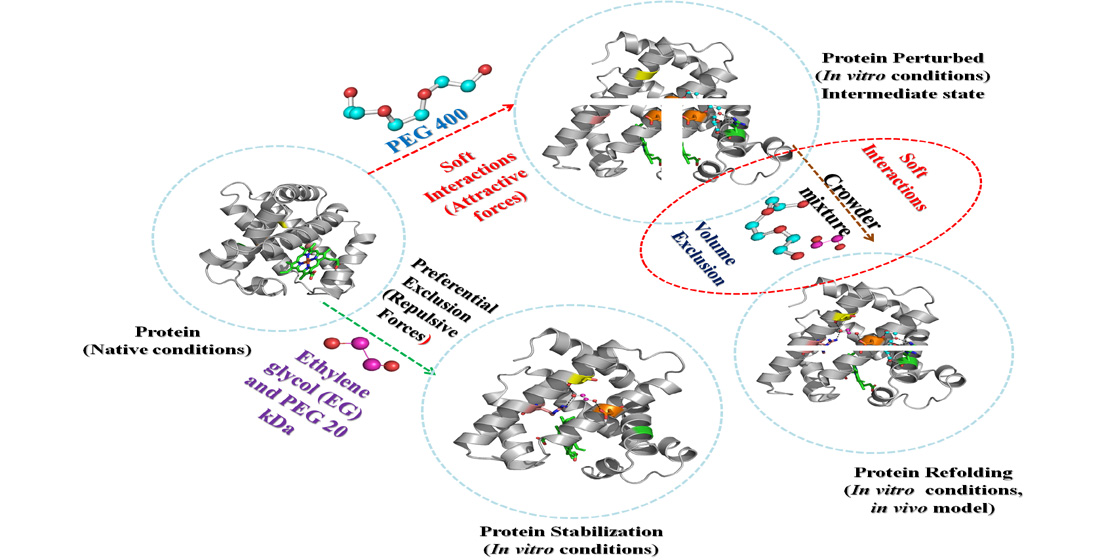
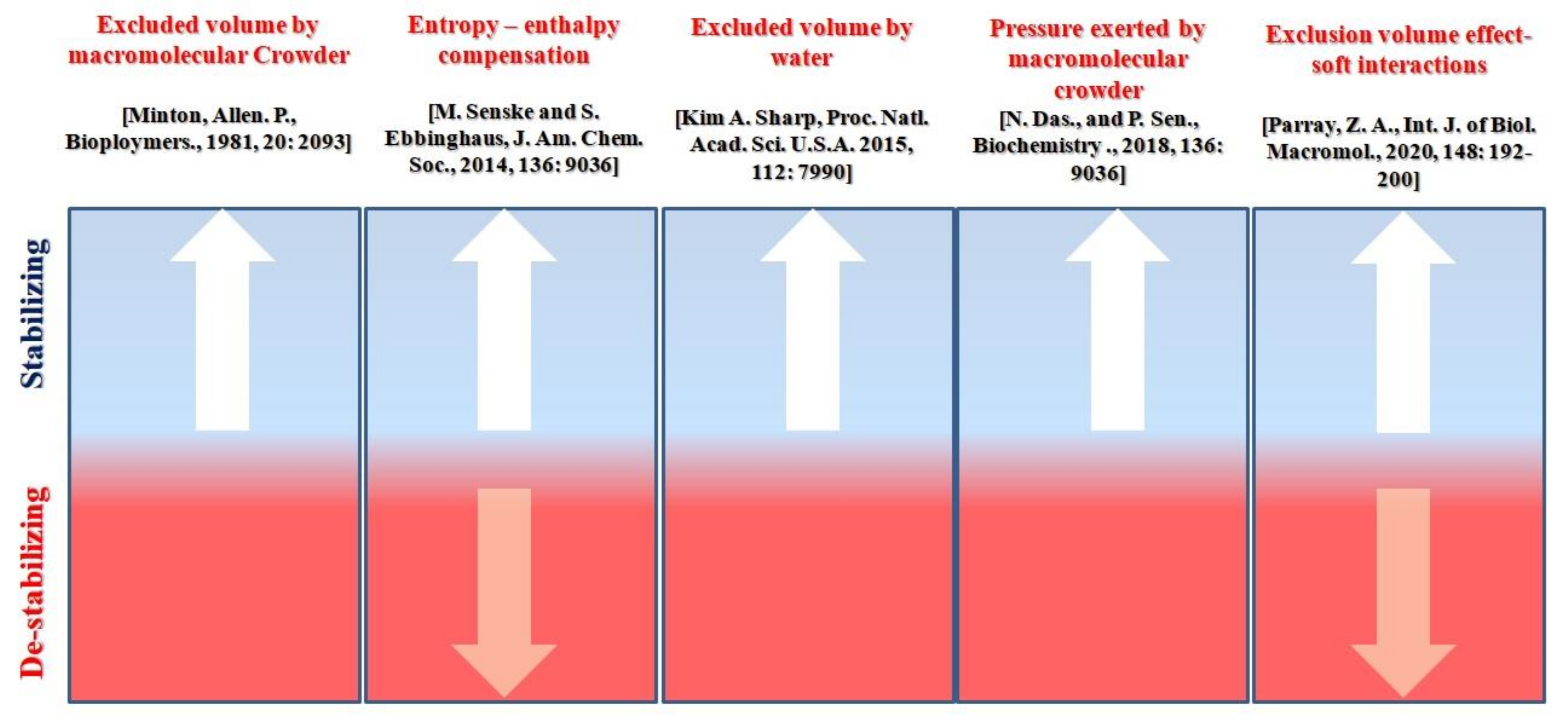
3. Discussion
4. Methodology
4.1. Materials
4.2. Methods
Preparation of Solutions of Protein and Reagents
4.3. Spectroscopic Techniques
4.3.1. Absorption Spectroscopy
4.3.2. Fluorescence Spectroscopy
4.3.3. Circular Dichroism (CD) Spectroscopy
4.3.4. Size Distribution Measurements
4.3.5. Thermal Denaturation Measurements
UV-vis Spectrophotometer
4.4. Methods Used for Binding Studies
4.4.1. Isothermal Titration Calorimetry Measurements
4.4.2. Computational Studies (In Silico)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Alberts, B. From the national academies. Cell Biol. Educ. 2002, 1, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.E.; Frydman, J. Protein folding in vivo: The importance of molecular chaperones. Curr. Opin. Struct. Biol. 2000, 10, 26–33. [Google Scholar] [CrossRef]
- Zimmerman, S.B.; Minton, A.P. Macromolecular crowding: Biochemical, biophysical, and physiological consequences. Annu Rev. Biophys Biomol. Struct. 1993, 22, 27–65. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J. Macromolecular crowding: Obvious but underappreciated. Trends Biochem. Sci. 2001, 26, 597–604. [Google Scholar] [CrossRef]
- Fulton, A.B. How crowded is the cytoplasm? Cell 1982, 30, 345–347. [Google Scholar] [CrossRef]
- Feig, M.; Sugita, Y. Variable interactions between protein crowders and biomolecular solutes are important in understanding cellular crowding. J. Phys. Chem. B 2012, 116, 599–605. [Google Scholar] [CrossRef]
- Gnutt, D.; Gao, M.; Brylski, O.; Heyden, M.; Ebbinghaus, S. Excluded-volume effects in living cells. Angew. Chem. Int. Ed. Engl. 2015, 54, 2548–2551. [Google Scholar] [CrossRef] [Green Version]
- Hall, D.; Minton, A.P. Macromolecular crowding: Qualitative and semiquantitative successes, quantitative challenges. Biochim. Biophys. Acta. 2003, 1649, 127–139. [Google Scholar] [CrossRef]
- Mondal, S.; Kallianpur, M.V.; Udgaonkar, J.B.; Krishnamoorthy, G. Molecular crowding causes narrowing of population heterogeneity and restricts internal dynamics in a protein. Methods Appl. Fluoresc. 2016, 4, 014003. [Google Scholar] [CrossRef]
- Sarkar, M.; Lu, J.; Pielak, G.J. Protein crowder charge and protein stability. Biochemistry 2014, 53, 1601–1606. [Google Scholar] [CrossRef]
- van den Berg, B.; Ellis, R.J.; Dobson, C.M. Effects of macromolecular crowding on protein folding and aggregation. EMBO J. 1999, 18, 6927–6933. [Google Scholar] [CrossRef]
- Wirth, A.J.; Platkov, M.; Gruebele, M. Temporal variation of a protein folding energy landscape in the cell. J. Am. Chem. Soc. 2013, 135, 19215–19221. [Google Scholar] [CrossRef]
- Kao, H.P.; Abney, J.R.; Verkman, A.S. Determinants of the translational mobility of a small solute in cell cytoplasm. J. Cell Biol. 1993, 120, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Kundu, J.; Mukherjee, S.K.; Chowdhury, P.K. Mixed macromolecular crowding: A protein and solvent perspective. ACS Omega 2018, 3, 4316–4330. [Google Scholar] [CrossRef] [Green Version]
- Parray, Z.A.; Ahmad, F.; Alajmi, M.F.; Hussain, A.; Hassan, M.I.; Islam, A. Interaction of polyethylene glycol with cytochrome c investigated via in vitro and in silico approaches. Sci. Rep. 2021, 11, 6475. [Google Scholar] [CrossRef]
- Ellis, R.J. Macromolecular crowding: An important but neglected aspect of the intracellular environment. Curr. Opin. Struct. Biol. 2001, 11, 114–119. [Google Scholar] [CrossRef]
- Minton, A.P. Excluded volume as a determinant of macromolecular structure and reactivity. Biopolymers 1981, 20, 2093–2120. [Google Scholar] [CrossRef]
- Malik, A.; Kundu, J.; Mukherjee, S.K.; Chowdhury, P.K. Myoglobin unfolding in crowding and confinement. J. Phys. Chem. B 2012, 116, 12895–12904. [Google Scholar] [CrossRef]
- Parray, Z.A.; Shahid, S.; Ahmad, F.; Hassan, M.I.; Islam, A. Characterization of intermediate state of myoglobin in the presence of PEG 10 under physiological conditions. Int. J. Biol. Macromol. 2017, 99, 241–248. [Google Scholar] [CrossRef]
- Shahid, S.; Hassan, M.I.; Islam, A.; Ahmad, F. Size-dependent studies of macromolecular crowding on the thermodynamic stability, structure and functional activity of proteins: In vitro and in silico approaches. Biochim. Biophys. Acta. 2017, 1861, 178–197. [Google Scholar] [CrossRef]
- Nasreen, K.; Ahamad, S.; Ahmad, F.; Hassan, M.I.; Islam, A. Macromolecular crowding induces molten globule state in the native myoglobin at physiological ph. Int. J. Biol. Macromol. 2018, 106, 130–139. [Google Scholar] [CrossRef]
- Parray, Z.A.; Ahamad, S.; Ahmad, F.; Hassan, M.I.; Islam, A. First evidence of formation of pre-molten globule state in myoglobin: A macromolecular crowding approach towards protein folding in vivo. Int. J. Biol. Macromol. 2019, 126, 1288–1294. [Google Scholar] [CrossRef]
- Nasreen, K.; Parray, Z.A.; Ahamad, S.; Ahmad, F.; Ahmed, A.; Freeh Alamery, S.; Hussain, T.; Hassan, M.I.; Islam, A. Interactions under crowding milieu: Chemical-induced denaturation of myoglobin is determined by the extent of heme dissociation on interaction with crowders. Biomolecules 2020, 10, 490. [Google Scholar] [CrossRef] [Green Version]
- Parray, Z.A.; Ahmad, F.; Alajmi, M.F.; Hussain, A.; Hassan, M.I.; Islam, A. Formation of molten globule state in horse heart cytochrome c under physiological conditions: Importance of soft interactions and spectroscopic approach in crowded milieu. Int. J. Biol. Macromol. 2020, 148, 192–200. [Google Scholar] [CrossRef]
- Spitzer, J. From water and ions to crowded biomacromolecules: In vivo structuring of a prokaryotic cell. Microbiol. Mol. Biol. Rev. 2011, 75, 491–506. [Google Scholar] [CrossRef] [Green Version]
- Dyson, H.J.; Wright, P.E. Unfolded proteins and protein folding studied by nmr. Chem. Rev. 2004, 104, 3607–3622. [Google Scholar] [CrossRef]
- Veprintsev, D.B.; Permyakov, S.E.; Permyakov, E.A.; Rogov, V.V.; Cawthern, K.M.; Berliner, L.J. Cooperative thermal transitions of bovine and human apo-alpha-lactalbumins: Evidence for a new intermediate state. FEBS Lett. 1997, 412, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Boesch, C.; Bundi, A.; Oppliger, M.; Wuthrich, K. 1h nuclear-magnetic-resonance studies of the molecular conformation of monomeric glucagon in aqueous solution. Eur. J. Biochem. 1978, 91, 209–214. [Google Scholar] [CrossRef]
- Bhat, M.Y.; Dar, T.A.; Singh, L.R. Casein Proteins: Structural and Functional Aspects, Milk Proteins—From Structure to Biological Properties and Health Aspects; IntechOpen: London, UK, 2016. [Google Scholar]
- Marinko, J.T.; Huang, H.; Penn, W.D.; Capra, J.A.; Schlebach, J.P.; Sanders, C.R. Folding and misfolding of human membrane proteins in health and disease: From single molecules to cellular proteostasis. Chem. Rev. 2019, 119, 5537–5606. [Google Scholar] [CrossRef]
- Parray, Z.A.; Hassan, M.I.; Ahmad, F.; Islam, A. Amphiphilic nature of polyethylene glycols and their role in medical research. Polym. Test. 2020, 82, 106316. [Google Scholar] [CrossRef]
- Richter, K.; Nessling, M.; Lichter, P. Macromolecular crowding and its potential impact on nuclear function. Biochim. Biophys. Acta 2008, 11, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snoussi, K.; Halle, B. Protein self-association induced by macromolecular crowding: A quantitative analysis by magnetic relaxation dispersion. Biophys. J. 2005, 88, 2855–2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Chen, W.; Wu, C.; Chen, H. Peg-protein interaction induced contraction of nald chains. PLoS ONE 2014, 9, e96616. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.B. Macromolecular crowding effects on macromolecular interactions: Some implications for genome structure and function. Biochim. Biophys. Acta 1993, 1216, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Wilf, J.; Minton, A.P. Evidence for protein self-association induced by excluded volume. Myoglobin in the presence of globular proteins. Biochim. Biophys. Acta 1981, 670, 316–322. [Google Scholar] [CrossRef]
- Spitzer, J.J.; Poolman, B. Electrochemical structure of the crowded cytoplasm. Trends Biochem. Sci. 2005, 30, 536–541. [Google Scholar] [CrossRef] [Green Version]
- Parray, Z.A.; Ahmad, F.; Hassan, M.I.; Hasan, I.; Islam, A. Effects of ethylene glycol on the structure and stability of myoglobin using spectroscopic, interaction, and in silico approaches: Monomer is different from those of its polymers. ACS Omega 2020, 5, 13840–13850. [Google Scholar] [CrossRef]
- Parray, Z.A.; Ahmad, F.; Hassan, M.I.; Islam, A. Conformational changes in cytochrome c directed by ethylene glycol accompanying complex formation: Protein-solvent preferential interaction or/and kosmotropic effect. Spectrochim. Acta Part A: Mol. Biomol. Spectrosc. 2020, 242, 118788. [Google Scholar] [CrossRef]
- Makhatadze, G.I.; Privalov, P.L. On the entropy of protein folding. Protein Sci. 1996, 5, 507–510. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.-X. Polymer crowders and protein crowders act similarly on protein folding stability. FEBS Letters 2013, 587, 394–397. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Gierasch, L. Macromolecular crowding remodels the energy landscape of a protein by favoring a more compact unfolded state. J. Am. Chem. Soc. 2010, 132, 10445–10452. [Google Scholar] [CrossRef] [Green Version]
- Johansen, D.; Jeffries, C.M.J.; Hammouda, B.; Trewhella, J.; Goldenberg, D.P. Effects of macromolecular crowding on an intrinsically disordered protein characterized by small-angle neutron scattering with contrast matching. Biophys. J. 2011, 100, 1120–1128. [Google Scholar] [CrossRef] [Green Version]
- McNulty, B.C.; Young, G.B.; Pielak, G.J. Macromolecular crowding in the Escherichia coli periplasm maintains α-synuclein disorder. J. Mol. Biol. 2006, 355, 893–897. [Google Scholar] [CrossRef]
- Sarkar, M.; Li, C.; Pielak, G.J. Soft interactions and crowding. Biophys. Rev. 2013, 5, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.T.; Reeder, B.J. Myoglobin. In Encyclopedia of Respiratory Medicine; Laurent, G.J., Shapiro, S.D., Eds.; Academic Press: Oxford, UK, 2006; pp. 73–76. [Google Scholar]
- Harvey, J.W. Chapter 9—Iron Metabolism and Its Disorders. In Clinical Biochemistry of Domestic Animals (Sixth Edition); Kaneko, J.J., Harvey, J.W., Bruss, M.L., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 259–285. [Google Scholar]
- Shahid, S.; Ahmad, F.; Hassan, M.I.; Islam, A. Relationship between protein stability and functional activity in the presence of macromolecular crowding agents alone and in mixture: An insight into stability-activity trade-off. Arch. Biochem. Biophys. 2015, 584, 42–50. [Google Scholar] [CrossRef]
- Ellis, R.J. Protein misassembly: Macromolecular crowding and molecular chaperones. Adv. Exp. Med. Biol. 2007, 594, 1–13. [Google Scholar]
- Homouz, D.; Perham, M.; Samiotakis, A.; Cheung, M.S.; Wittung-Stafshede, P. Crowded, cell-like environment induces shape changes in aspherical protein. Proc. Natl. Acad. Sci. USA 2008, 105, 11754–11759. [Google Scholar] [CrossRef] [Green Version]
- Rawat, S.; Suri, C.R.; Sahoo, D.K. Molecular mechanism of polyethylene glycol mediated stabilization of protein. Biochem. Biophys. Res. Commun. 2010, 392, 561–566. [Google Scholar] [CrossRef]
- Eaton, W.A.; Hochstrasser, R.M. Single-crystal spectra of ferrimyoglobin complexes in polarized light. J. Chem. Phys. 1968, 49, 985–995. [Google Scholar] [CrossRef]
- Noble, J.E.; Bailey, M.J. Quantitation of protein. Methods Enzym. 2009, 463, 73–95. [Google Scholar]
- Antosiewicz, J.M.; Shugar, D. Uv-vis spectroscopy of tyrosine side-groups in studies of protein structure. Part 2: Selected applications. Biophys. Rev. 2016, 8, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F. Measuring the conformational stability of enzymes. In Thermostablity of Enzymes; Gupta, M.N., Ed.; Springer: New Delhi, India, 1993; pp. 95–112. [Google Scholar]
- Tofani, L.; Feis, A.; Snoke, R.E.; Berti, D.; Baglioni, P.; Smulevich, G. Spectroscopic and interfacial properties of myoglobin/surfactant complexes. Biophys. J. 2004, 87, 1186–1195. [Google Scholar]
- Ross, M.L.; Kunkel, J.; Long, S.; Asuri, P. Combined effects of confinement and macromolecular crowding on protein stability. Int. J. Mol. Sci. 2020, 21, 8516. [Google Scholar] [CrossRef]
- Batra, J.; Xu, K.; Zhou, H.X. Nonadditive effects of mixed crowding on protein stability. Proteins 2009, 77, 133–138. [Google Scholar]
- Shahid, S.; Ahmad, F.; Hassan, M.I.; Islam, A. Mixture of macromolecular crowding agents has a non-additive effect on the stability of proteins. Appl. Biochem. Biotechnol 2019, 188, 927–941. [Google Scholar] [CrossRef]
- Nick Pace, C.; Scholtz, J.M.; Grimsley, G.R. Forces stabilizing proteins. FEBS Lett. 2014, 588, 2177–2184. [Google Scholar]
- Sarkar, M.; Smith, A.E.; Pielak, G.J. Impact of reconstituted cytosol on protein stability. Proc. Natl. Acad. Sci. USA 2013, 110, 19342–19347. [Google Scholar] [CrossRef] [Green Version]
- Mittal, S.; Chowhan, R.K.; Singh, L.R. Macromolecular crowding: Macromolecules friend or foe. Biochim. Biophys. Acta 2015, 1850, 1822–1831. [Google Scholar] [PubMed]
- Qu, P.; Wang, Y.X.; Wu, G.H.; Lu, Z.H.; Xu, M.T. Effect of polyethylene glycols on the alkaline-induced molten globule intermediate of bovine serum albumin. Int. J. Biol. Macromol. 2012, 51, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Breslauer, K.J.; Remeta, D.P.; Chou, W.Y.; Ferrante, R.; Curry, J.; Zaunczkowski, D.; Snyder, J.G.; Marky, L.A. Enthalpy-entropy compensations in drug-DNA binding studies. Proc. Natl. Acad. Sci. USA 1987, 84, 8922–8926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, K.A. Analysis of the size dependence of macromolecular crowding shows that smaller is better. Proc. Natl. Acad. Sci. USA 2015, 112, 7990–7995. [Google Scholar] [CrossRef] [Green Version]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar]
- Kim, Y.C.; Mittal, J. Crowding induced entropy-enthalpy compensation in protein association equilibria. Phys. Rev. Lett. 2013, 110, 208102. [Google Scholar] [CrossRef] [Green Version]
- Goto, Y.; Takahashi, N.; Fink, A.L. Mechanism of acid-induced folding of proteins. Biochemistry 1990, 29, 3480–3488. [Google Scholar] [CrossRef]
- Puett, D. The equilibrium unfolding parameters of horse and sperm whale myoglobin. Effects of guanidine hydrochloride, urea, and acid. J. Biol. Chem. 1973, 248, 4623–4634. [Google Scholar] [CrossRef]
- Nozaki, Y. The preparation of guanidine hydrochloride. Methods Enzym. 1972, 26, 43–50. [Google Scholar]
- Huglin, M.B. Specific refractive index increments of polymer solutions. Part i. Literature values. J. Appl. Polym. Sci. 1965, 9, 3963–4001. [Google Scholar] [CrossRef]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef]
- Tripathi, T. Calculation of thermodynamic parameters of protein unfolding using far-ultraviolet circular dichroism. J. Proteins Proteom. 2013, 4, 85–91. [Google Scholar]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with pymol and autodock/vina. J. Comput Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biovia, D.S. Discovery Studio Modeling Environment; Version 4, Released; Dassault Systemes: San Diego, CA, USA, 2015. [Google Scholar]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. Ligplot: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 1995, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thermodynamic Parameters (Units) | Ka (M−1) | ∆H° (cal mol−1) | ∆S° (cal mol−1 K−1) | ∆G° (cal mol−1) |
|---|---|---|---|---|
| Step 1 | 24,060 (±66) | −22,280 (±649) | −59.2 | −4629 (±649) |
| Step 2 | 996 (±22) | −44,830 (±405) | −137 | −3983 (±405) |
| [PEG 400 +EG], mg mL−1 | Δε409 | |
|---|---|---|
| Tm (°C) | ΔHm (kcal mol−1) | |
| 0 + 0 | 81.5 (±0.5) | 120.20 (±0.92) |
| 0 + 50 | 78.6 (±0.60) | 120.90 (±0.93) |
| 0 + 300 | 65.26 (±0.60) | 106.30 (±0.82) |
| 50 + 0 | 78.5 (±0.61) | 98.70 (±1.4) |
| 300 + 0 | 63.2 (±0.60) | 70.10 (±0.54) |
| 50 + 50 | 77.76 (±0.23) | 85.02 (±0.70) |
| 50 + 300 | 70.69 (±0.63) | 117.27 (±0.25) |
| 300 + 50 | 73.75 (±0.5) | 90.06 (±0.17) |
| Mixture of Crowders (PEG 400 + EG), mg mL−1 | Rh (Å) | ε409, M−1 cm−1 | F335 | [θ] (deg cm2 dmol−1) at Wavelength in nm | |
|---|---|---|---|---|---|
| 222 | 272 | ||||
| No Crowder (0 + 0) | 23.46 (±0.68) | 171,645 (±285) | 29 (±1.24) | −26,190 (±278) | 182 (±12) |
| (0 + 50) | 23.10 (±0.58) | 187,716 (±509) | 33 (±2.4) | −26,015 (±258) | 183 (±14) |
| (0 + 300) | 22.70 (±0.60) | 169,831 (±619) | 33 (±2.0) | −26,065 (±150) | 185 (±10) |
| (50 + 0) | 21.30 (±0.50) | 166,561 (±499) | 43 (±4.1) | −25,119 (±204) | 159 (±7) |
| (300 + 0) | 33.8 (±0.40) | 85,776 (±152) | 120 (±5.2) | −13,987 (±104) | 61 (±4) |
| (50 + 50) | 23.05 (±0.60) | 194,821 (±689) | 38 (±2.3) | −27,547 (±278) | 181 (±12) |
| (50 + 300) | 22.80 (±0.70) | 183,029 (±539) | 42 (±4.4) | −27,278 (±254) | 176 (±10) |
| (300 + 50) | 26.45 (±0.60) | 130,644 (±282) | 98 (±6.4) | −18,908 (±121) | 105 (±6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parray, Z.A.; Ahmad, F.; Hassan, M.I.; Ahmed, A.; Almajhdi, F.N.; Malik, A.; Hussain, T.; Islam, A. Structural Refolding and Thermal Stability of Myoglobin in the Presence of Mixture of Crowders: Importance of Various Interactions for Protein Stabilization in Crowded Conditions. Molecules 2021, 26, 2807. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092807
Parray ZA, Ahmad F, Hassan MI, Ahmed A, Almajhdi FN, Malik A, Hussain T, Islam A. Structural Refolding and Thermal Stability of Myoglobin in the Presence of Mixture of Crowders: Importance of Various Interactions for Protein Stabilization in Crowded Conditions. Molecules. 2021; 26(9):2807. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092807
Chicago/Turabian StyleParray, Zahoor Ahmad, Faizan Ahmad, Md. Imtaiyaz Hassan, Anwar Ahmed, Fahad N. Almajhdi, Ajamaluddin Malik, Tajamul Hussain, and Asimul Islam. 2021. "Structural Refolding and Thermal Stability of Myoglobin in the Presence of Mixture of Crowders: Importance of Various Interactions for Protein Stabilization in Crowded Conditions" Molecules 26, no. 9: 2807. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092807