
Pharmacokinetics, Prostate Distribution and Metabolic Characteristics of Four Representative Flavones after Oral Administration of the Aerial Part of Glycyrrhiza uralensis in Rats

Abstract
:1. Introduction
2. Results
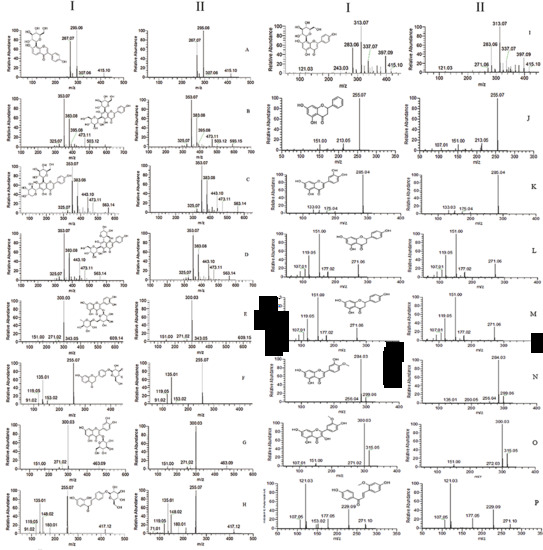
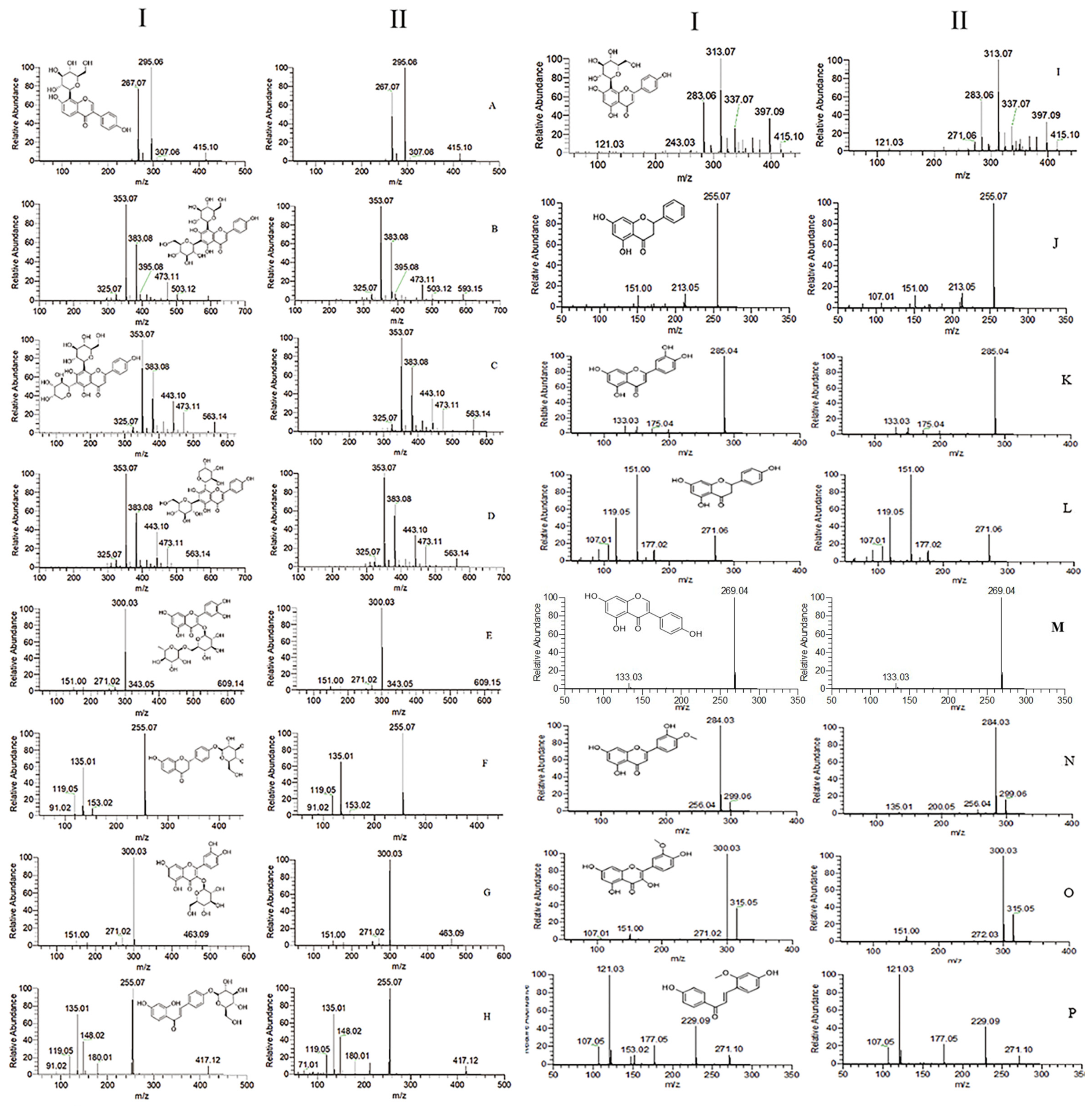
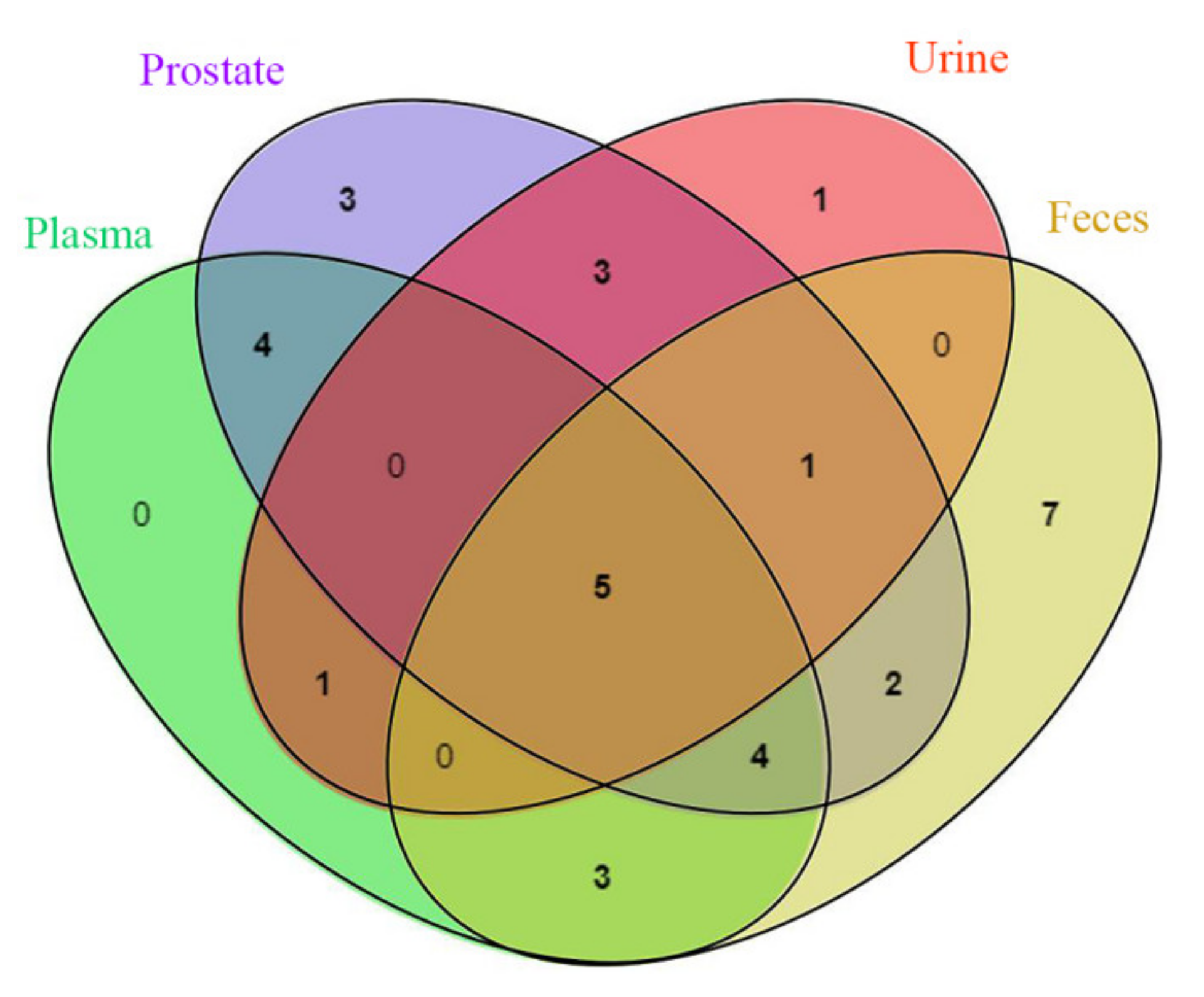
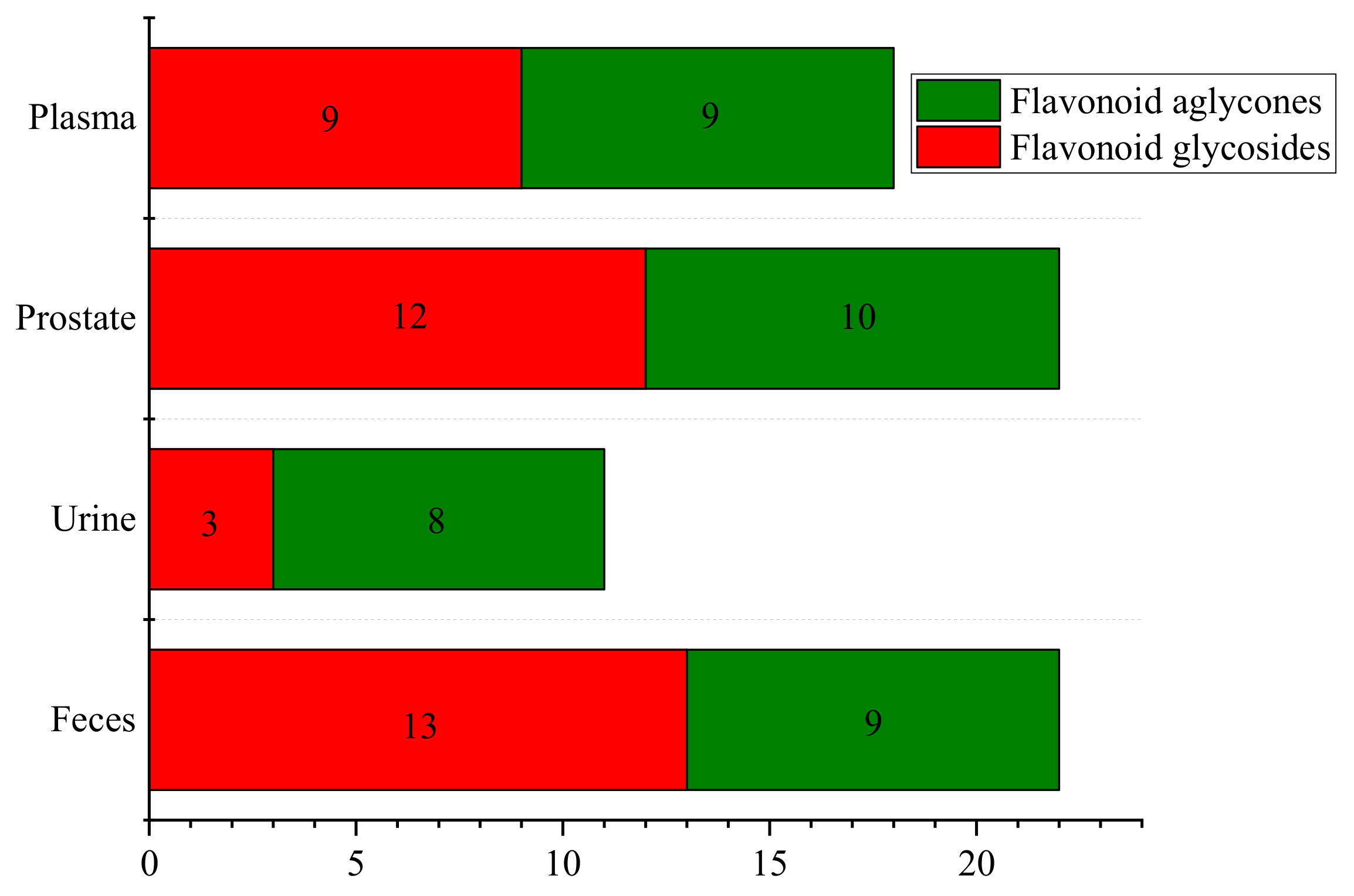
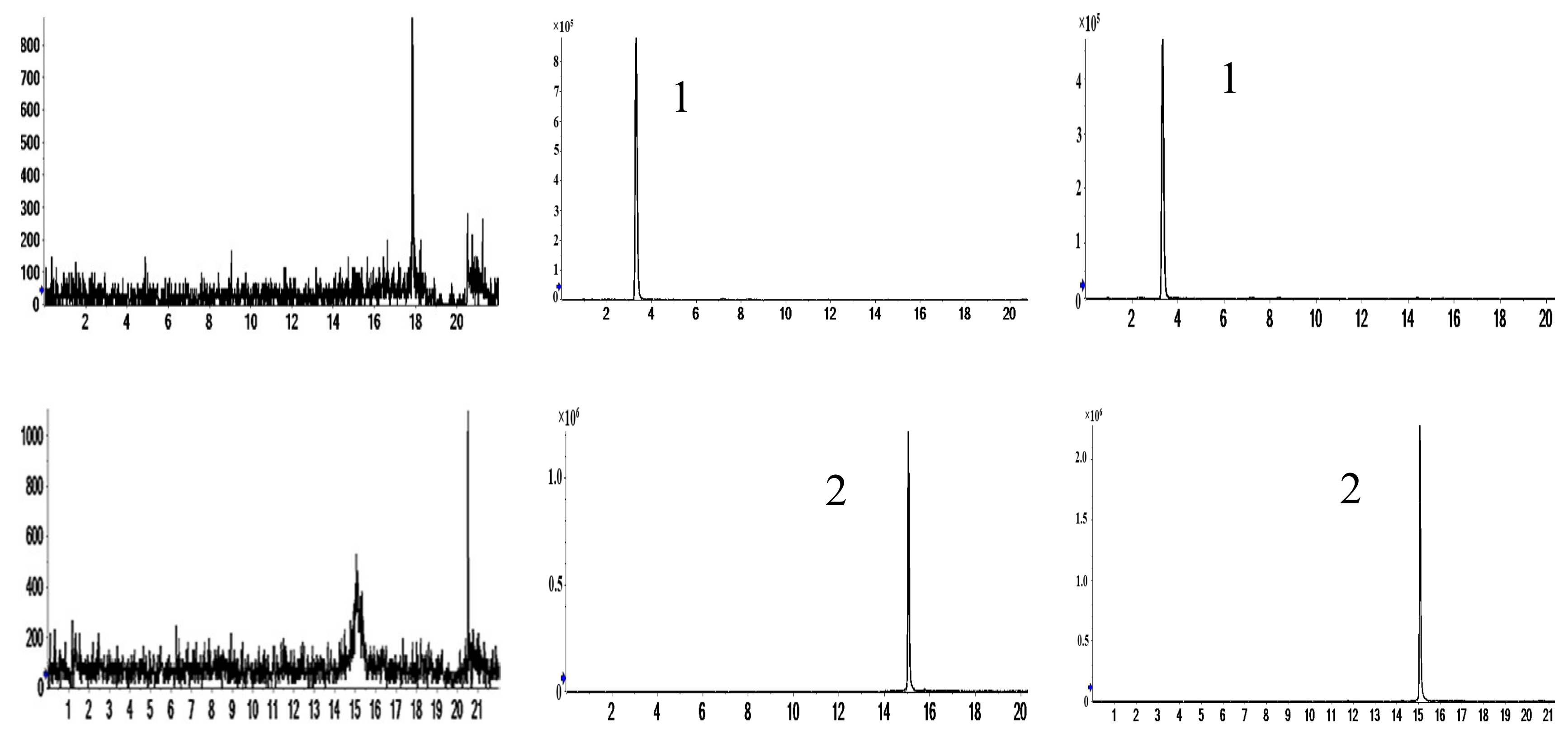
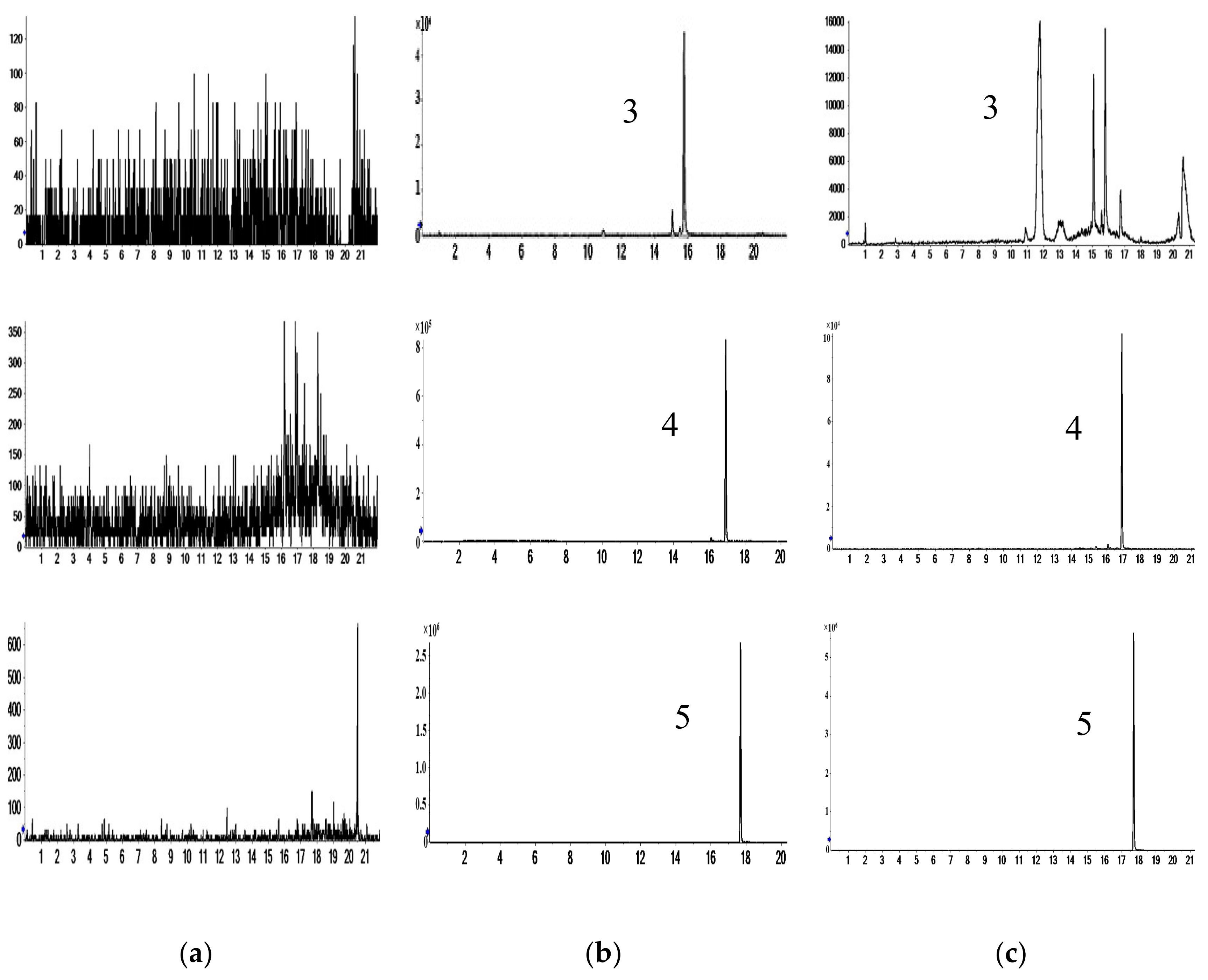
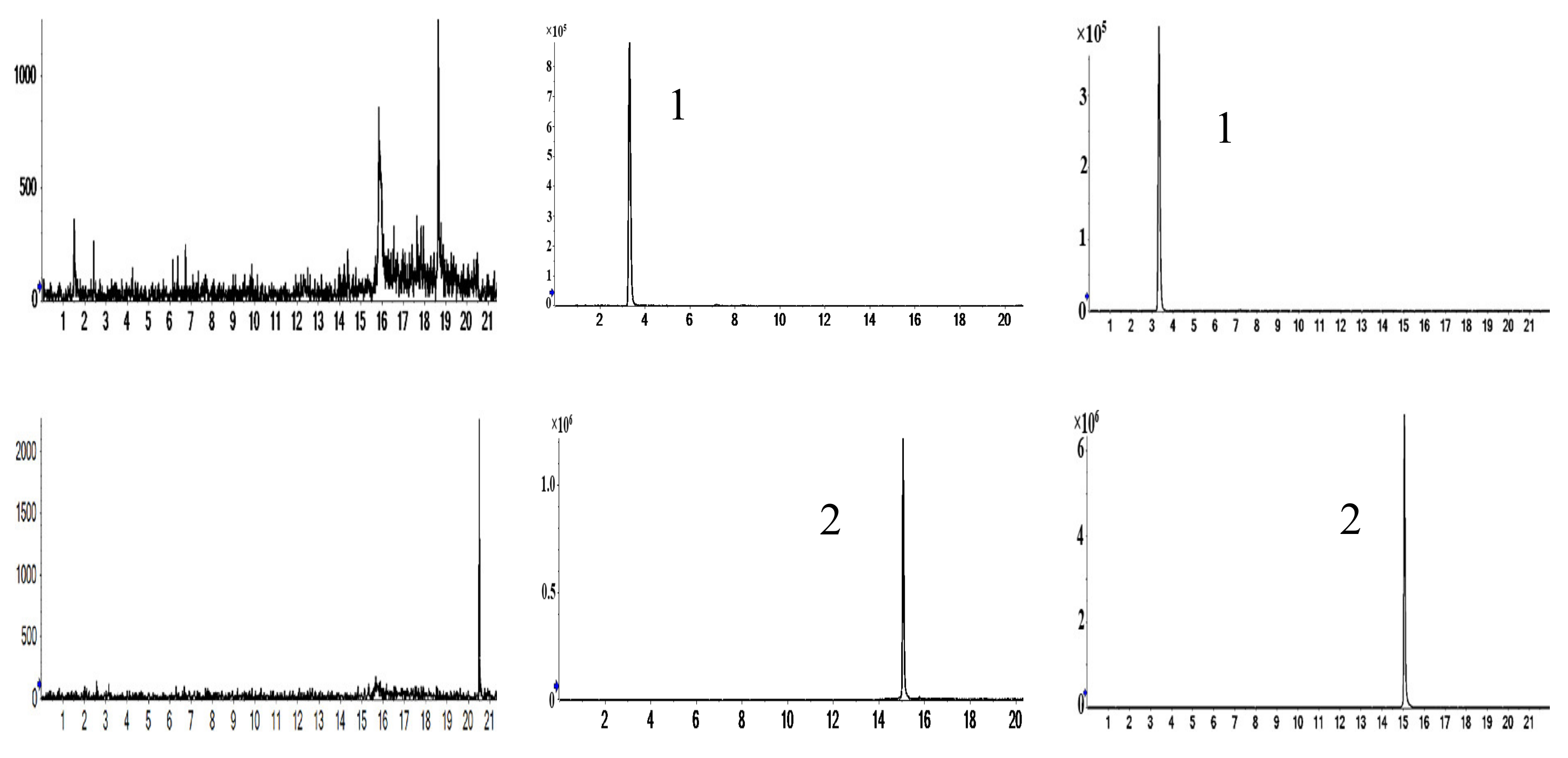
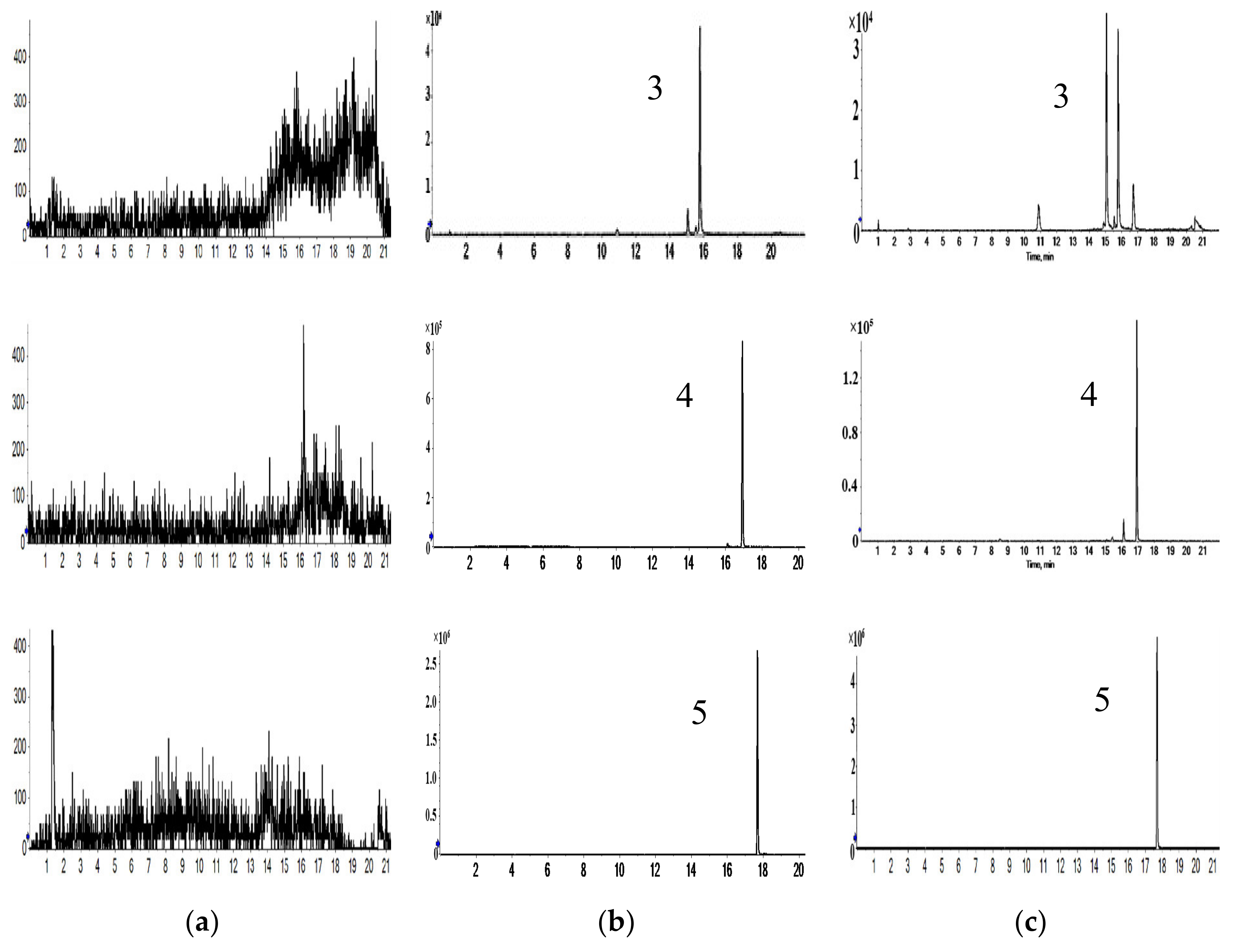
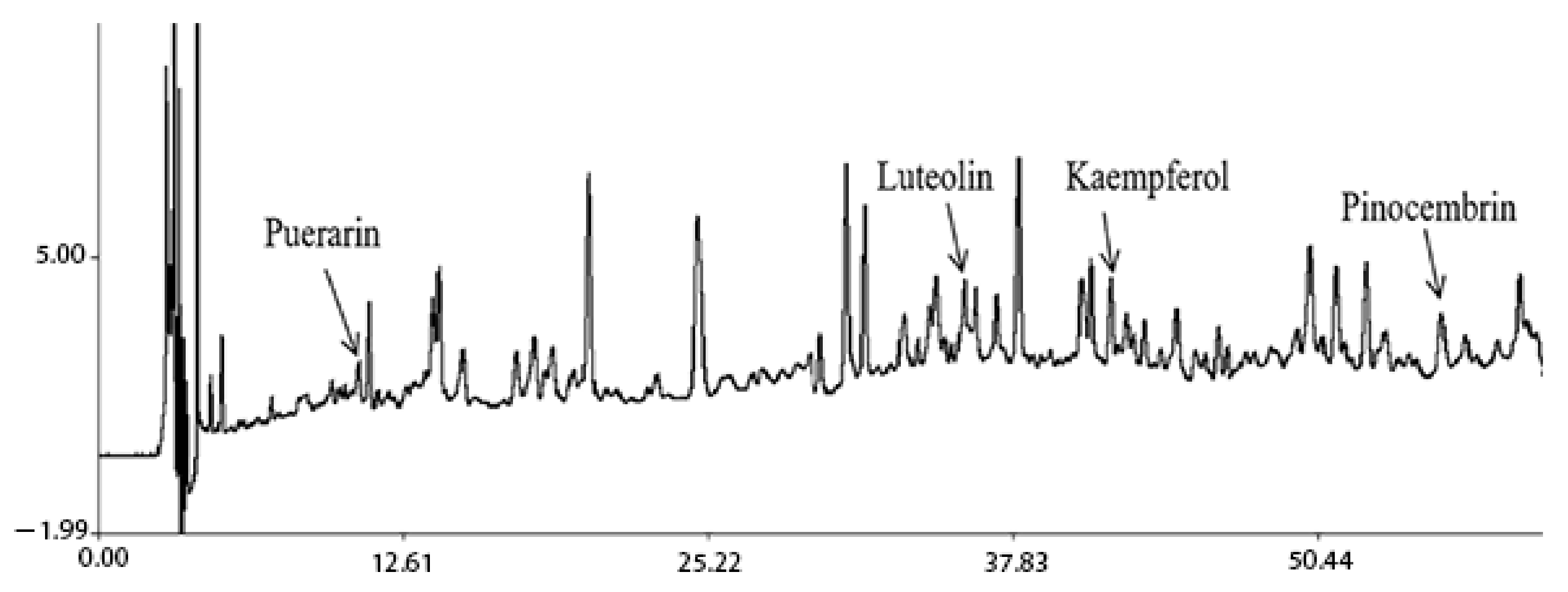
2.1. Characterization of Prototype Flavonoids in Biological Sample
2.2. Metabolism of Four Representative Flavonoids
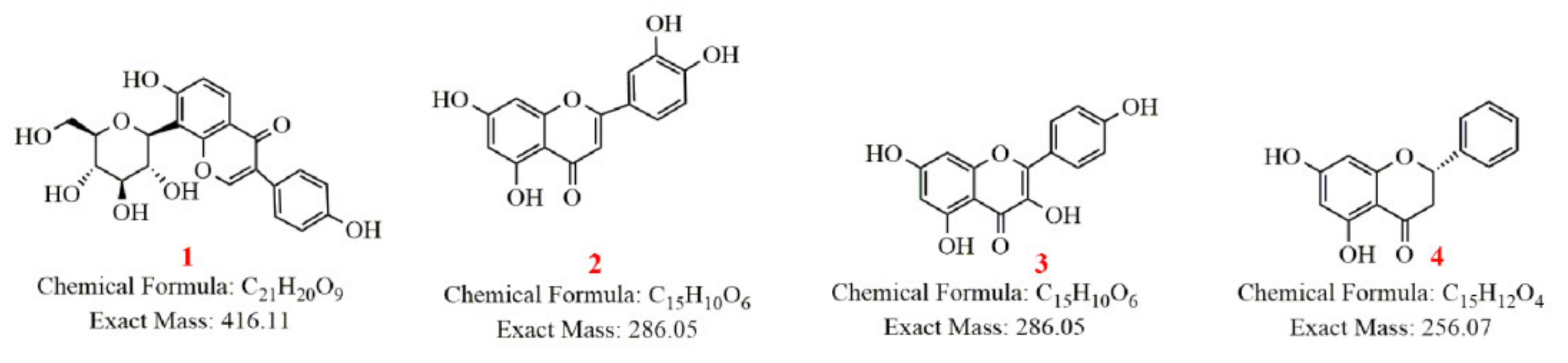
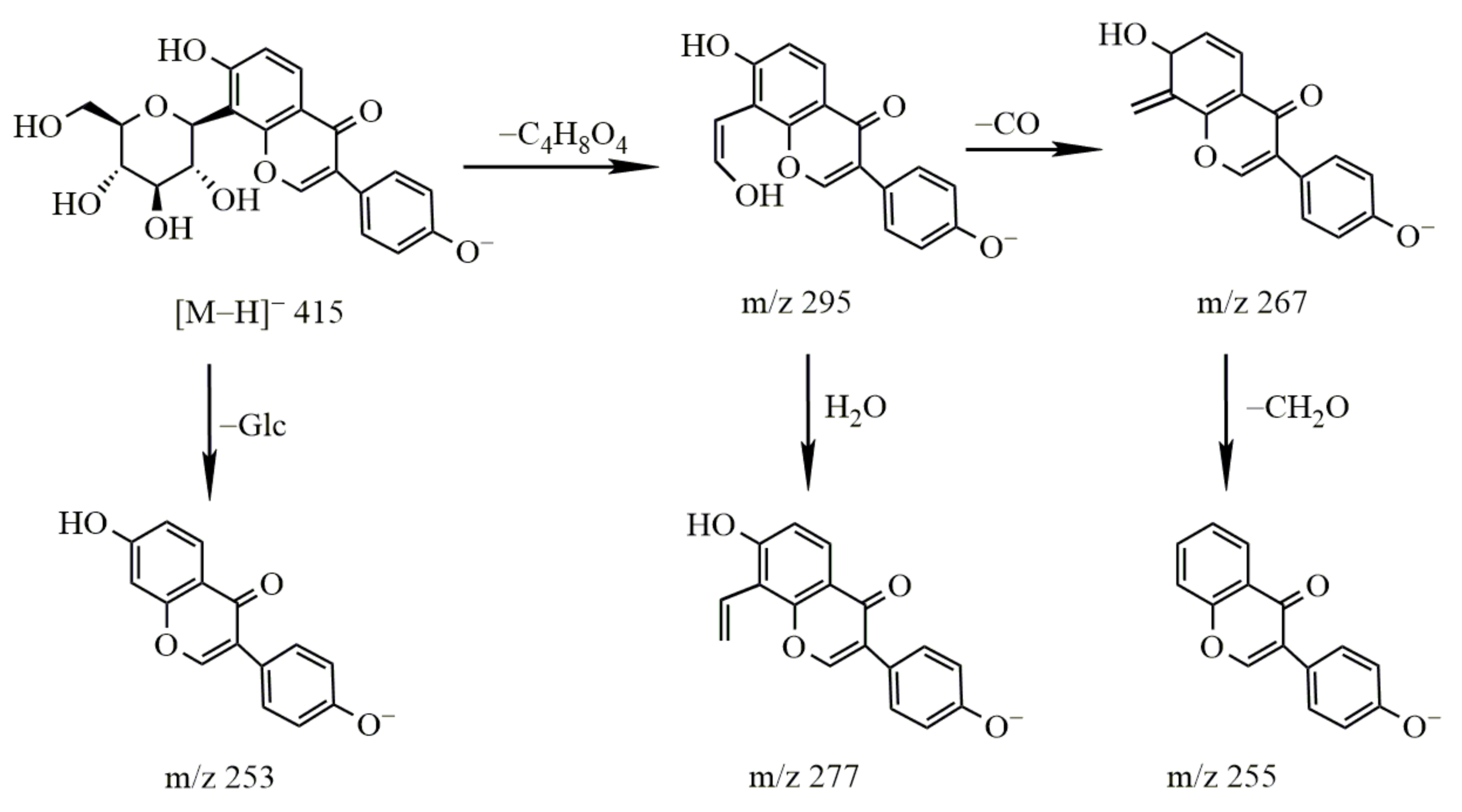
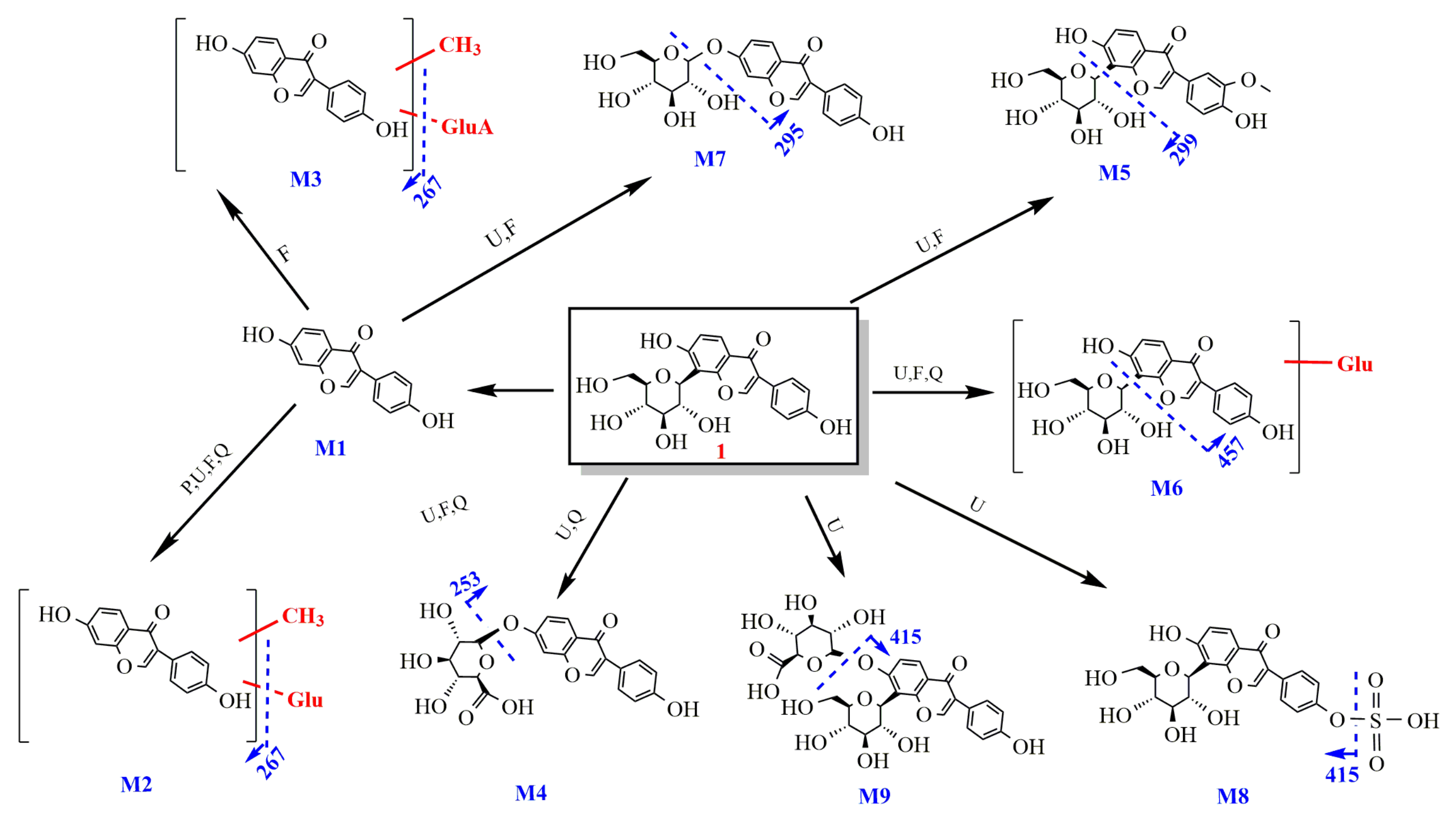
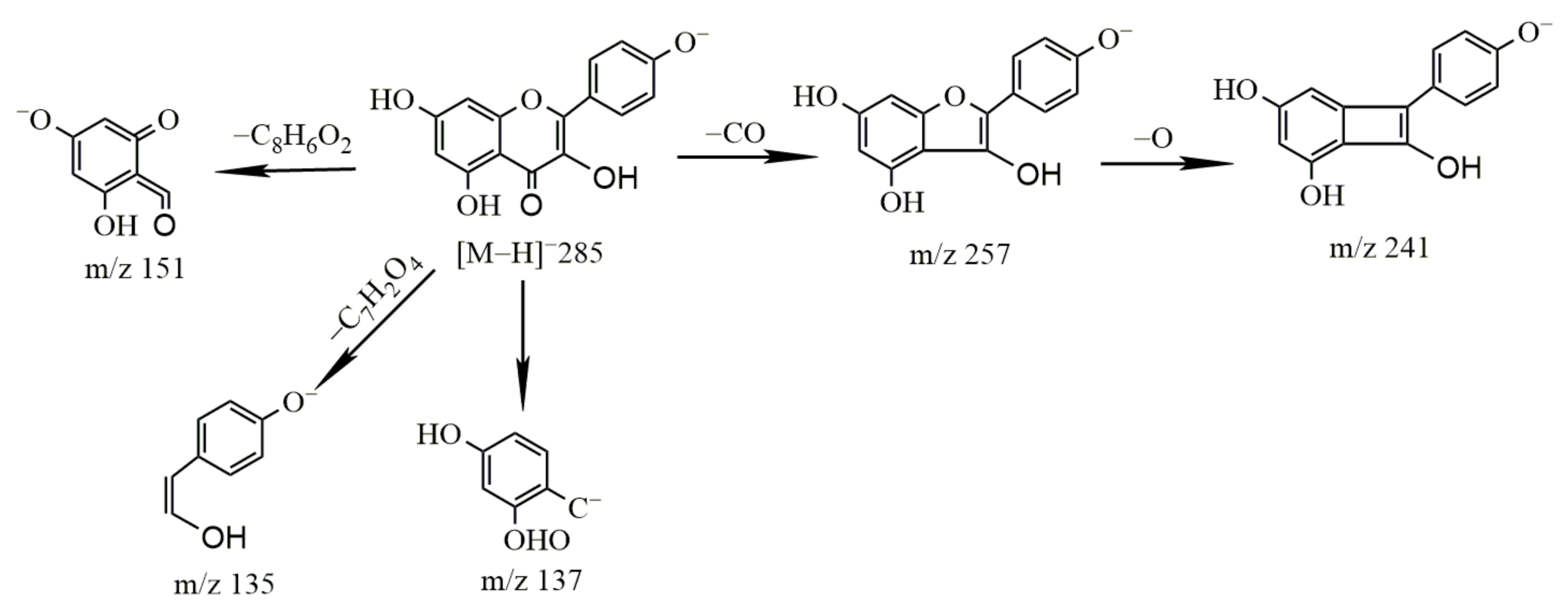
2.2.1. Study on the Metabolism of Puerarin
- Identification of puerarin
- 2.
- Metabolism of puerarin
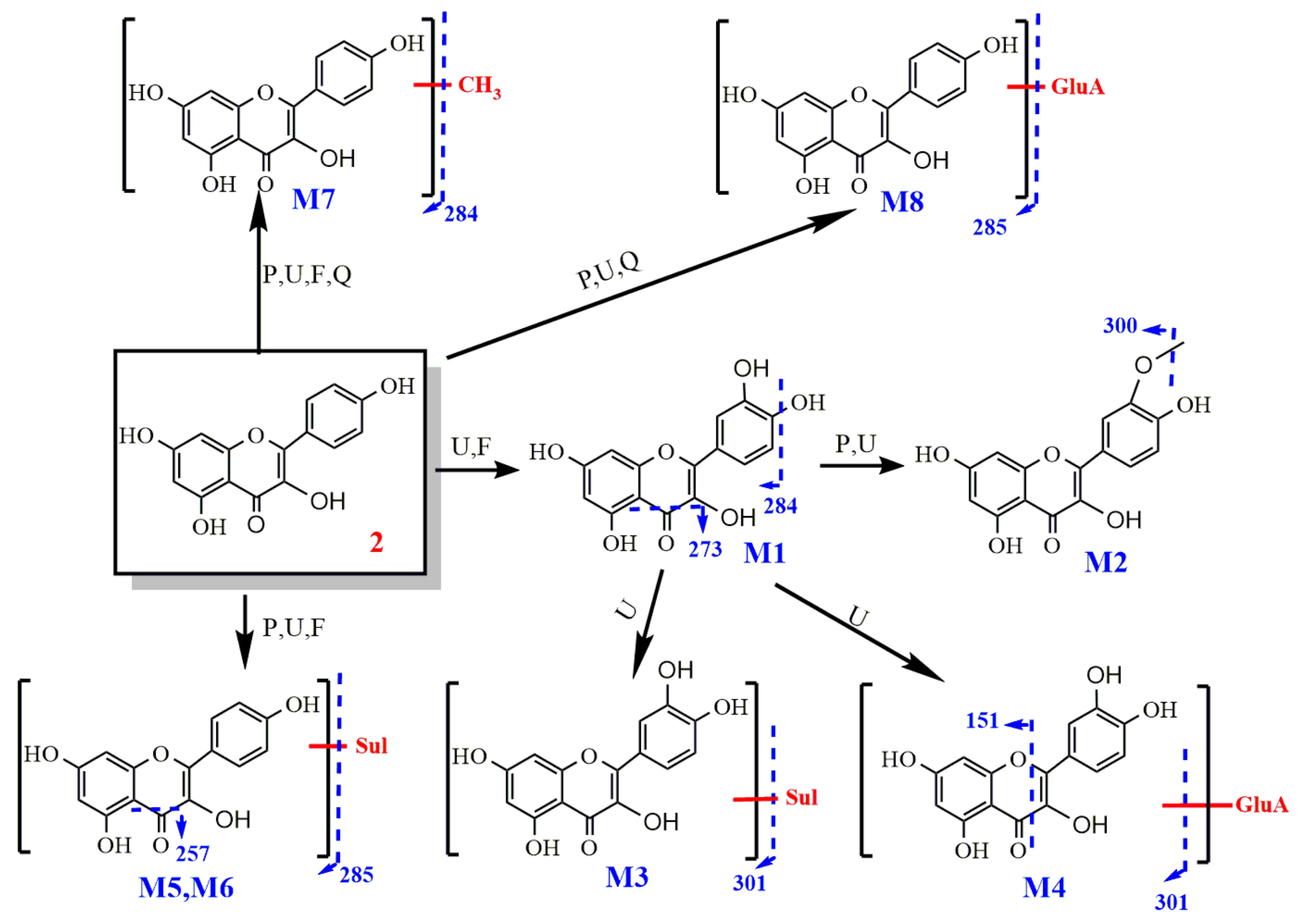
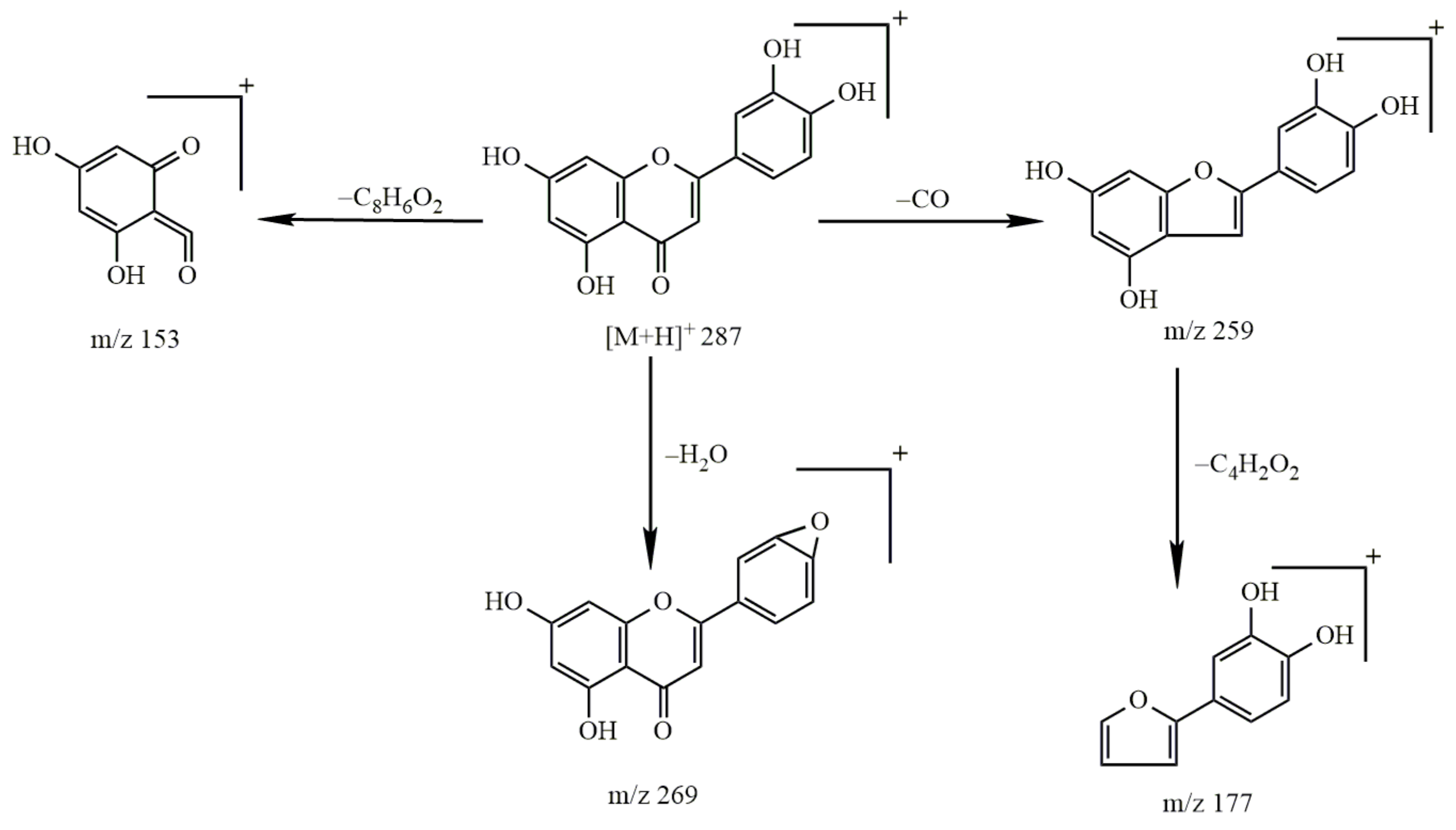
2.2.2. Study on the Metabolism of Kaempferol
- Identification of kaempferol
- 2.
- Metabolism of kaempferol
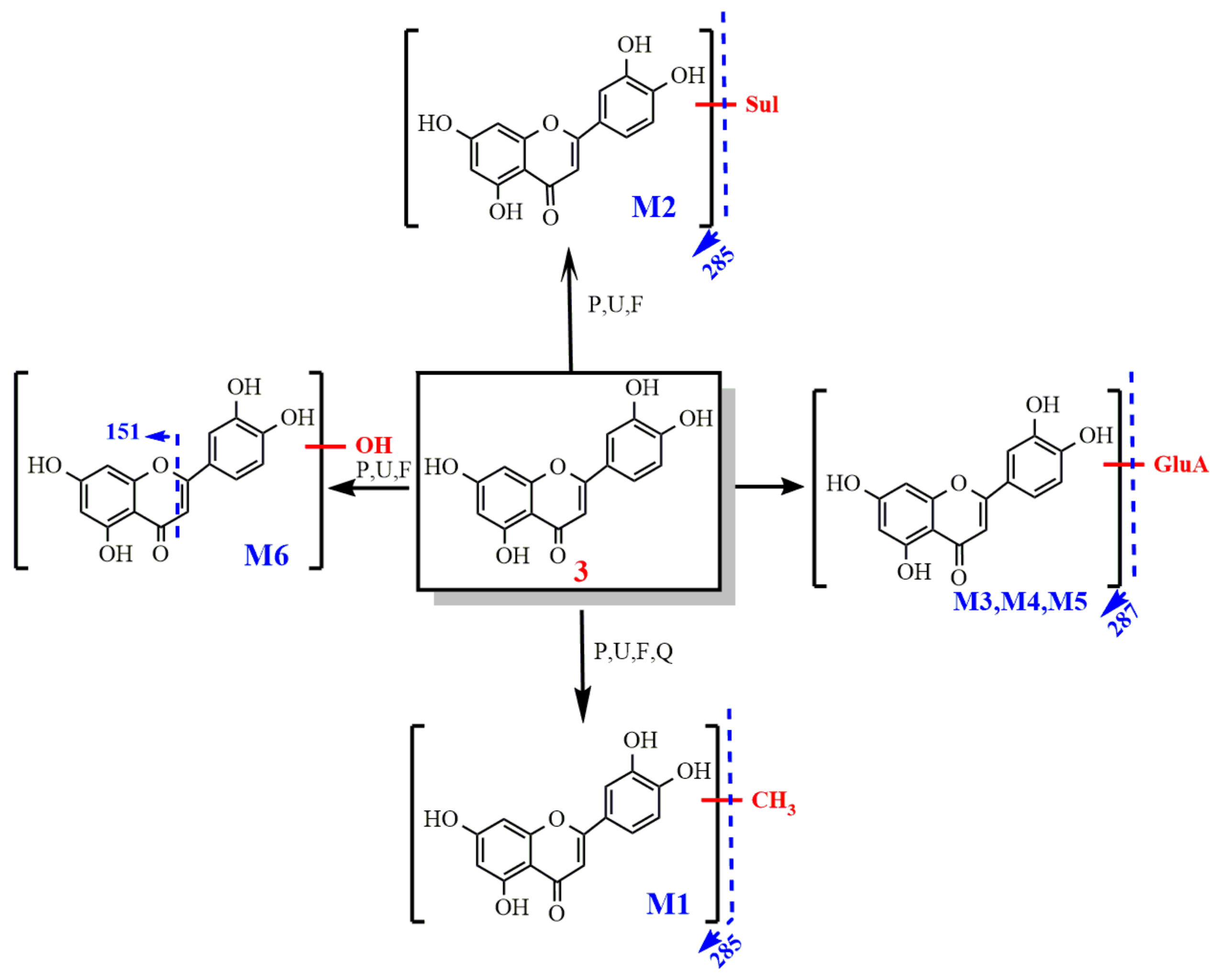
2.2.3. Study on the Metabolism of Luteolin
- Identification of luteolin
- 2.
- Metabolism of luteolin
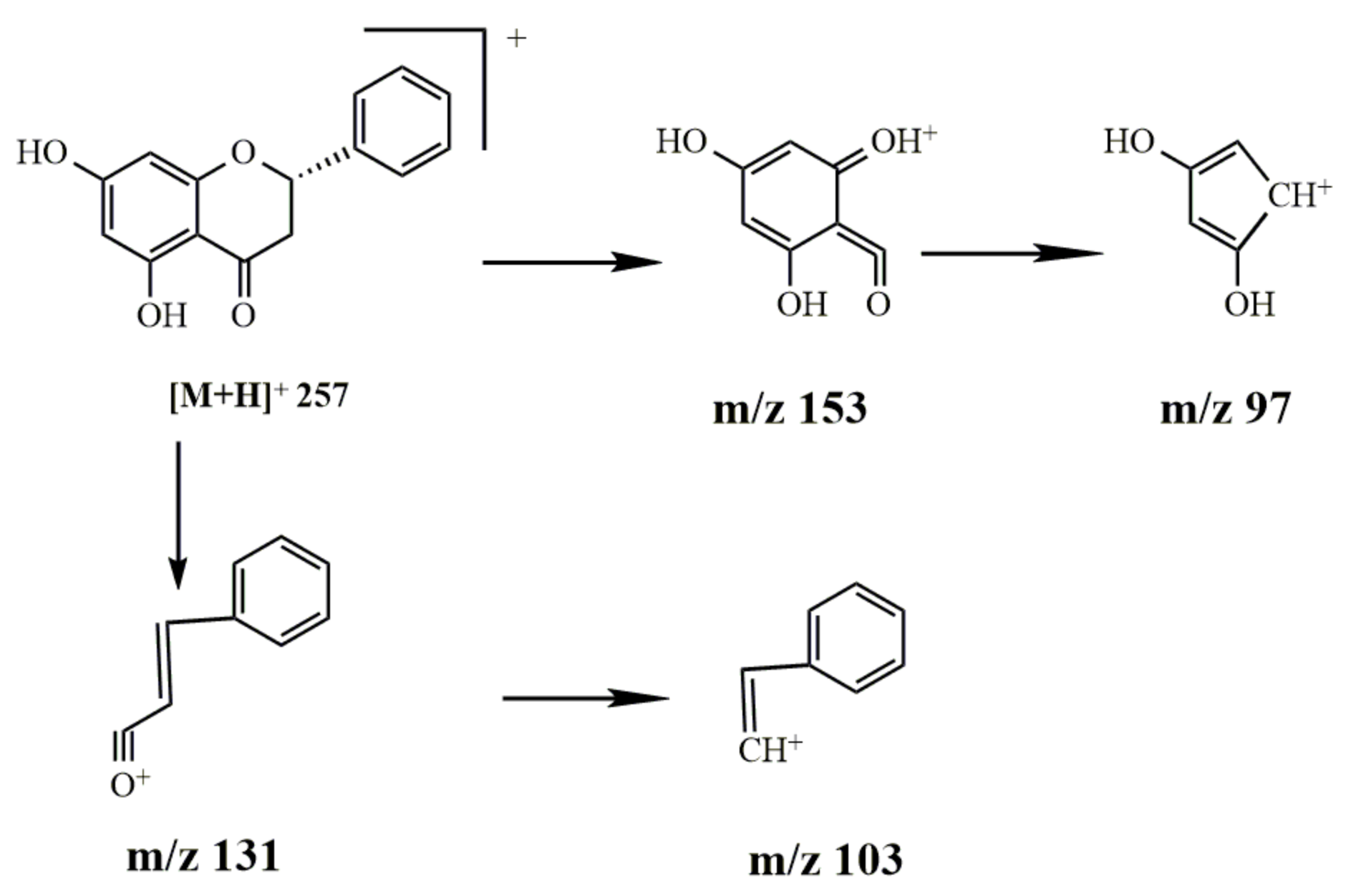
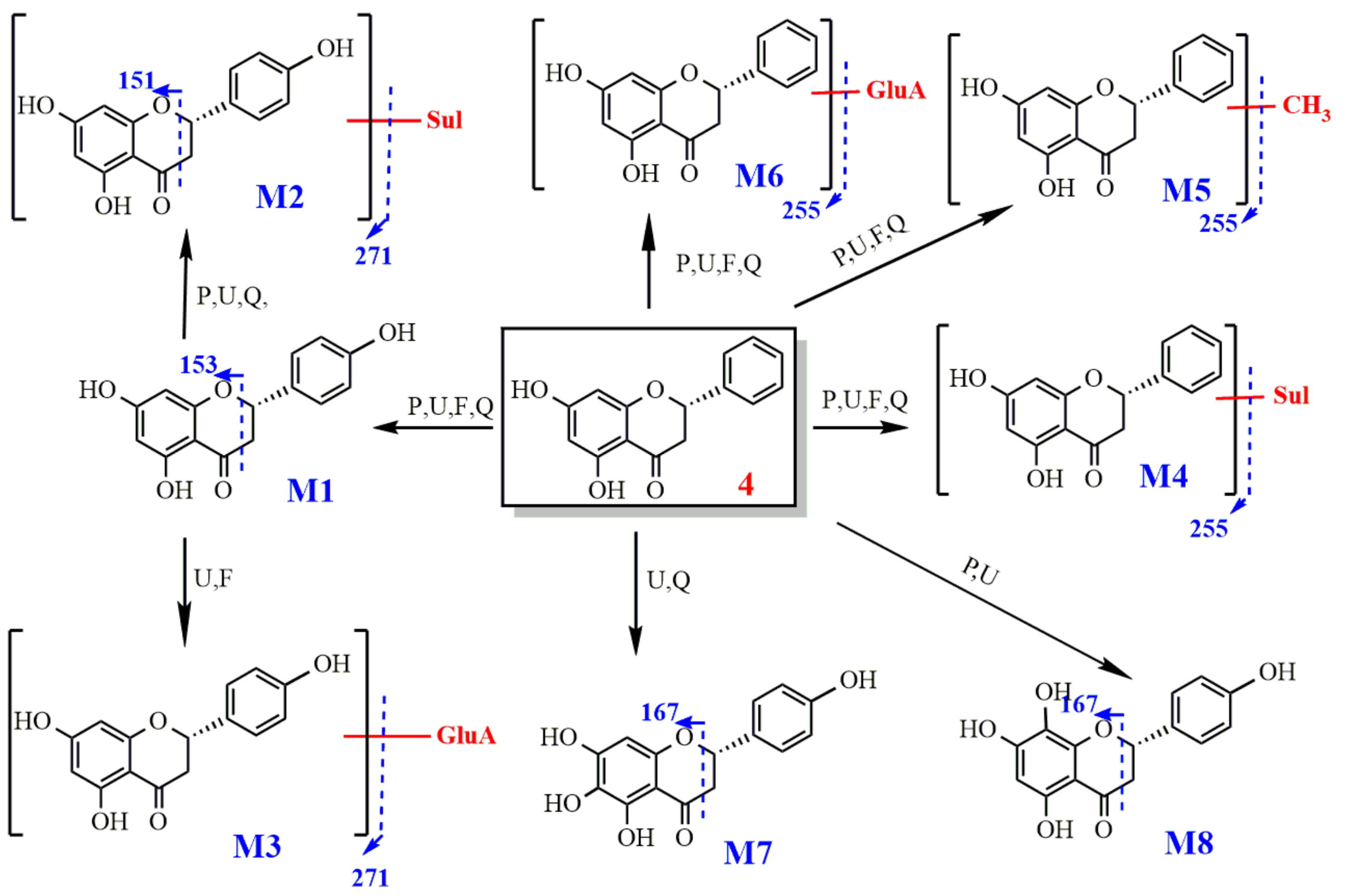
2.2.4. Study on the Metabolism of Pinocembrin
- Identification of pinocembrin
- 2.
- Metabolism of pinocembrin
2.3. Method Validation on Pharmacokinetics and Prostate Distribution
2.3.1. Selectivity
2.3.2. Calibration Curves
2.3.3. Matrix Effect and Extraction Recovery
2.3.4. Accuracy and Precision
2.3.5. Stability
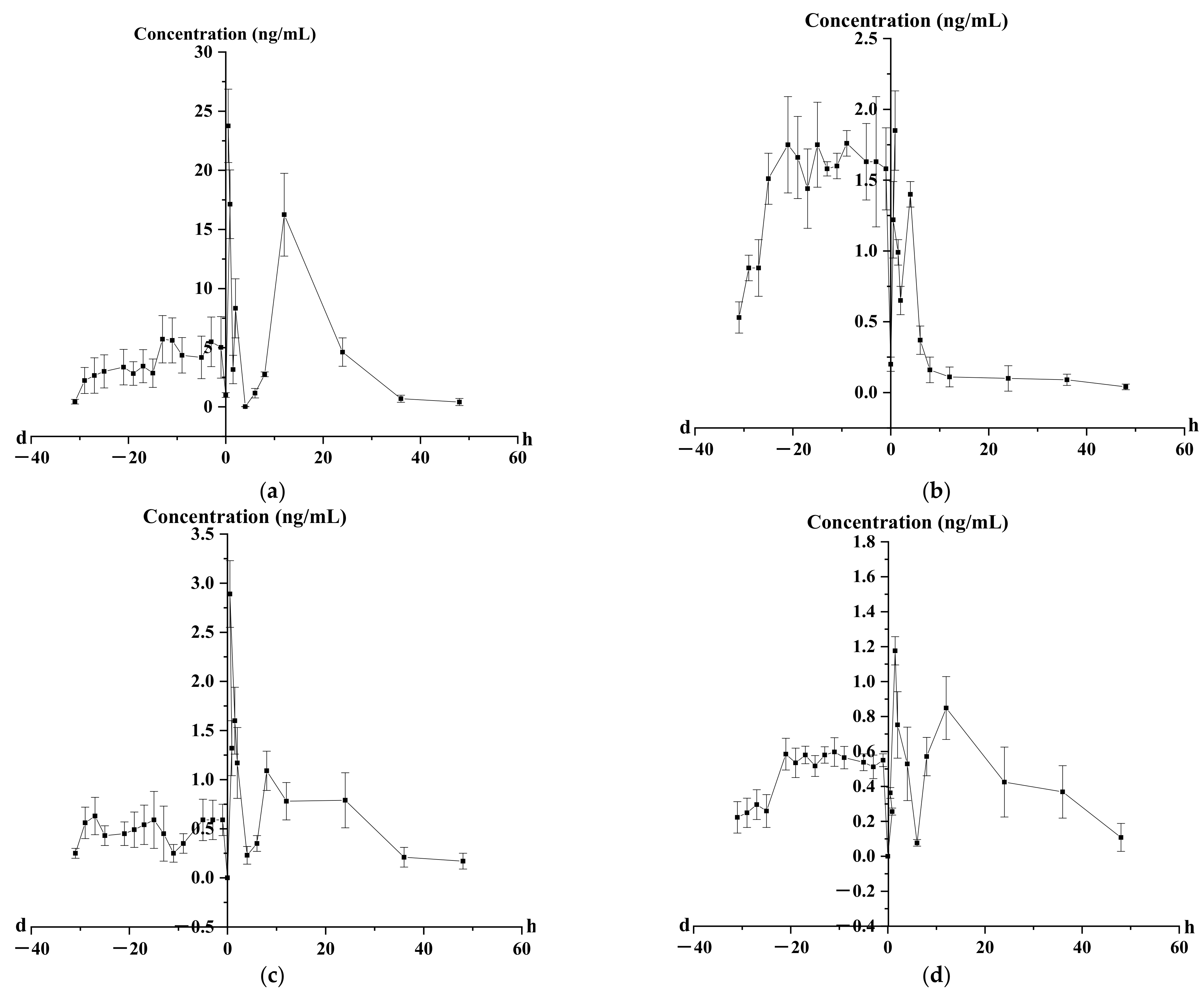
2.4. Pharmacokinetics Analysis of Four Representative Flavonoids
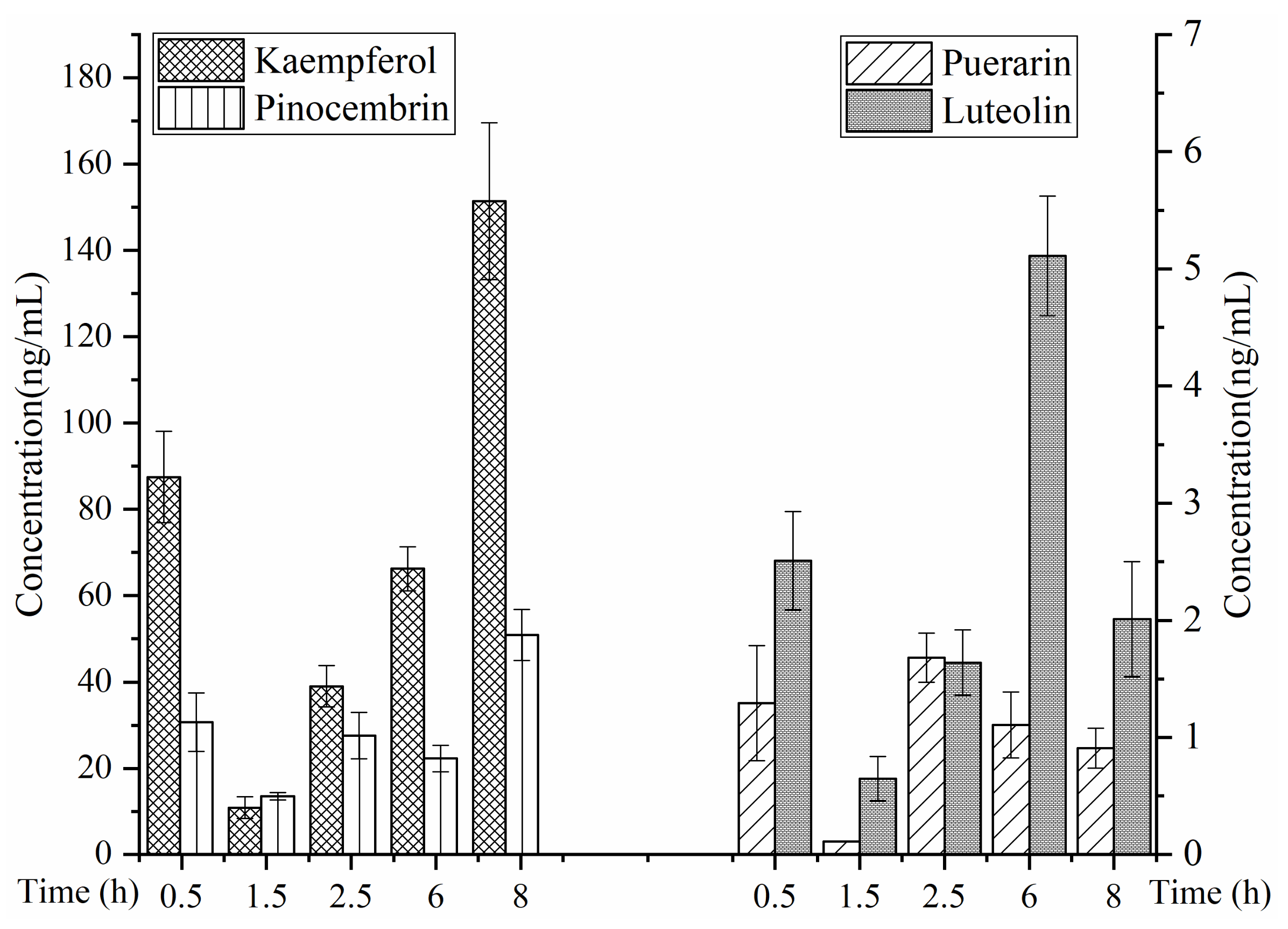
2.5. Prostate Distribution of Four Representative Flavonoids
3. Discussion
3.1. Characterization of Prototype Flavonoids in Biological Sample
3.2. Metabolism of Four Representative Flavonoids
3.2.1. Metabolism of Puerarin
3.2.2. Metabolism of Kaempferol
3.2.3. Metabolism of Luteolin
3.2.4. Metabolism of Pinocembrin
3.2.5. Metabolism of Flavonoids
3.3. Pharmacokinetic and Prostate Distribution
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation of the Aerial Part of Glycyrrhiza Uralensis Sample
4.3. Animals
4.4. Preparation of Plasma, Urine, Fecal and Prostate Samples
4.4.1. Metabolic Characteristics of Four Representative Flavonoids
4.4.2. Content Determination of Four Representative Flavonoids
4.5. Preparation of Calibration Standards and Quality Control (QC) Samples
4.6. Instrumentation and Analytical Conditions
4.6.1. Metabolic Characteristics of Four Representative Flavonoids
4.6.2. Content Determination of Four Representative Flavonoids
4.7. Method Validation on Pharmacokinetics and Prostate Distribution
4.7.1. Selectivity
4.7.2. Calibration Curves
4.7.3. Extraction Recovery and Matrix Effect
4.7.4. Accuracy and Precision
4.7.5. Stability
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Liu, H.; Cui, J.; Zhang, L.; Chang, G.; Wang, W. Screening of anti-chronic nonbacterial prostatitis activity of different extractions of the aerial part of Glycyrrhiza uralensis, and network pharmacology research. Biomed. Rep. 2021, 15, 99. [Google Scholar] [CrossRef] [PubMed]
- Akbar, E.; Roghayeh, R. Preparation and biological activity of nanocapsulated Glycyrrhiza glabra L. var. glabra. Flavour Fragr. J. 2015, 30, 113–119. [Google Scholar]
- Andrea, S.; Jonas, T.; Hubertus, K.; Annegret, S. Efficacy of leaf extract fractions of Glycyrrhiza glabra L. against downy mildew of cucumber (Pseudoperonospora cubensis). Eur. J. Plant Pathol. 2012, 134, 755–762. [Google Scholar]
- Abolfazl, S.; Javad, A.; Vahid, S.; Seyedeh, F.; Amirhossein, S.; Rezvan, S.; Javad, A. Identification and biological activity of the volatile compounds of Glycyrrhiza triphylla Fisch. & C.A.Mey. Microb. Pathog. 2017, 109, 39–44. [Google Scholar]
- Fan, J.; Kuang, Y.; Dong, Z.; Yi, Y.; Zhou, Y.; Li, B.; Qiao, X.; Ye, M. Prenylated Phenolic Compounds from the Aerial Parts of Glycyrrhiza uralensis as PTP1B and alpha-Glucosidase Inhibitors. J. Nat. Prod. 2020, 83, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, L.; Saija, A.; Cristani, M.; Cimino, F.; D’Arrigo, M.; Trombetta, D.; Rao, F.; Ruberto, G. Phytocomplexes from liquorice (Glycyrrhiza glabra L.) leaves—Chemical characterization and evaluation of their antioxidant, anti-genotoxic and anti-inflammatory activity. Fitoterapia 2011, 82, 546–556. [Google Scholar] [CrossRef]
- Xiao, X.; Xu, L.; Hu, H.; Yang, Y.; Zhang, X.; Peng, Y.; Xiao, P. DPPH Radical Scavenging and Postprandial Hyperglycemia Inhibition Activities and Flavonoid Composition Analysis of Hawk Tea by UPLC-DAD and UPLC-Q/TOF MS(E). Molecules 2017, 22, 1622. [Google Scholar] [CrossRef]
- Chang, G.; Bo, Y.; Cui, J.; Zhao, Z.; Wang, W.; Hou, J. Main chemical constituents in aerial parts of Glycyrrhiza uralensis by UPLC-Q-Exactive Orbitrap-MS. China J. Chin. Mater. Med. 2021, 46, 1449–1459. [Google Scholar]
- Fan, Y. Isolation, Identification, Quantification and Bioactivity Assessment of the Chemical Constituents in Glycyrrhiza Uralensis Leaves; Inner Mongolia University: Inner Mongolia, China, 2018. [Google Scholar]
- Zeng, N.; Fan, X.; Jiang, F. Determination of six soybean isoflavones in vegetable protein drinks by UPLC-MS/MS. Food Sci. Technol. 2018, 43, 303–307. [Google Scholar]
- Ma, Z.; Chen, K.; Li, Y. Determination of 25 characteristic components in Danhe Granules by LC-MS and consistency analysis of preparations. Chin. Tradit. Herb. Drugs 2019, 50, 5970–5979. [Google Scholar]
- Yuan, J.; Li, L.; Cai, Z.; Wu, N.; Chen, C.; Yin, S.; Liu, S.; Wang, W.; Mei, Y.; Wei, L.; et al. Qualitative Analysis and Componential Differences of Chemical Constituents in Taxilli Herba from Different Hosts by UFLC-Triple TOF-MS/MS. Molecules 2021, 26, 6373. [Google Scholar] [CrossRef]
- Ji, X.; Wang, Z.; Dong, B. Simultaneous determination of typhaneoside and isorhamnetin-3-O-neohesperidoside in Tianqi Tongjing capsules by UPLC-MS/MS. Chin. J. Pharm. Anal. 2018, 38, 1055–1060. [Google Scholar]
- Singh, P.; Bajpai, V.; Gond, V.; Kumar, A.; Tadigoppula, N.; Kumar, B. Determination of Bioactive Compounds of Fenugreek (Trigonella foenum-graecum) Seeds Using LC-MS Techniques. In Legume Genomics; Humana: New York, NY, USA, 2020; Volume 2107, pp. 377–393. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, H.; Shi, H. Simultaneous quantification of nine constituents in Ziziphi Spinosae Semen by HPLC-MS. Chin. Tradit. Herb. Drugs 2015, 46, 123–127. [Google Scholar]
- Zhao, X.; Zhao, Y.-L.; Liu, X.-M.; Han, W.; Yu, Z.-G. Simultaneous determination of six isoflavonoids in rat plasma after administration of total flavonoid from Gegen by ultra-HPLC-MS/MS. J. Sep. Sci. 2012, 35, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Dong, Y.; Xu, A. Rapid analysis on flavonoids in Gegen Zhiju Soft Capsule by UPLC/Q-TOF-MS. Chin. Tradit. Herb. Drugs 2015, 46, 970–976. [Google Scholar]
- Wu, Y.; Wang, X.; Yan, W.; Hua, H.; Xu, G.; Lei, M. Analysis of flavonoids in acacia honey and locust flower by UPLC-MS/MS. Chin. J. Pharm. Anal. 2017, 37, 796–802. [Google Scholar]
- Meng, X.; Le, S.; Yang, Z. Investigation on biotransformation characteristics of astragalosides in human intestinal microbiota. Food Drug 2018, 20, 161–167. [Google Scholar]
- Manach, C.; Donovan, J.L. Pharmacokinetics and metabolism of dietary flavonoids in humans. Free Radic. Res. 2004, 38, 771–785. [Google Scholar] [CrossRef]
- Xie, Y.; Zeng, X.; Li, G.; Cai, Z.; Ding, N.; Ji, G. Assessment of intestinal absorption of total flavones of Hippophae rhamnoides L. in rat using in situ absorption models. Drug Dev. Ind. Pharm. 2010, 36, 787–794. [Google Scholar] [CrossRef]
- Zeng, X.; Xie, Y.; Yuan, X. Research progress in disposition of flavonoids in vivo. Chin. J. New Drugs 2010, 19, 750–754. [Google Scholar]
- Chen, L.; Fan, H.; Bian, Y.; Yang, N.; Jing, S.; Wang, Q. Study on Metabolites of Gegen Qinlian Dispensing Granule in Rat. Chin. J. Exp. Tradit. Med. Formulae 2011, 17, 101–104. [Google Scholar]
- Yang, F.; Wu, D.; Chen, L.; Yang, N.; Bian, Y. Studies on Metabolites of Main Components of Gegen Qinlian Decoction in Rats. Chin. J. Exp. Tradit. Med. Formulae 2013, 19, 164–168. [Google Scholar]
- Yuan, Y.; Zheng, S.; Zeng, L.; Deng, Z.; Zhang, B.; Li, H. The Phenolic Compounds, Metabolites, and Antioxidant Activity of Propolis Extracted by Ultrasound-Assisted Method. J. Food Sci. 2019, 84, 3850–3865. [Google Scholar] [CrossRef] [PubMed]
- Sonmezdag, A.S.; Kelebek, H.; Selli, S. Characterization of Aroma-Active Compounds, Phenolics, and Antioxidant Properties in Fresh and Fermented Capers (Capparis spinosa) by GC-MS-Olfactometry and LC-DAD-ESI-MS/MS. J. Food Sci. 2019, 84, 2449–2457. [Google Scholar] [CrossRef] [PubMed]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [Green Version]
- Geng, P.; Zhang, R.; Aisa, H.A.; He, J.; Qu, K.; Zhu, H.; Abliz, Z. Fast profiling of the integral metabolism of flavonols in the active fraction of Gossypium herbaceam L. using liquid chromatography/multi-stage tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 1877–1888. [Google Scholar] [CrossRef]
- Han, X.; Wang, S.; Yang, X.X.; Li, T.; Zhao, H.; Zhou, L.; Zhao, L.; Bao, Y.; Meng, X. Analysis of plasma migration components in Patrinia villosa (Thunb.) Juss. effective parts by UPLC-Q-TOF-MS. Biomed. Chromatogr. 2020, 34, e4701. [Google Scholar] [CrossRef]
- Yang, L. Research on the MetaboliteIdentification of Luteolin in Rats by HPLC-MS/MS; Dalian University of Technology: Dalian, China, 2016. [Google Scholar]
- Li, J.; He, M.; Feng, Y.; Li, T.; Li, Y.; Wu, B.; Ou, Y.; Yang, S. In-vivo Studies on Metabolism of Luteolin and Luteoloside in Rats. Tradit. Chin. Drug Res. Clin. Pharmacol. 2017, 28, 61–68. [Google Scholar]
- Li, Y.; Shangguan, Y. UPLC-MS/MS Analysis of the Metabolites of Pinocembrin in Human Liver Microsome. Chin. J. Mod. Appl. Pharm. 2012, 29, 159–163. [Google Scholar]
- Liu, H.; Cui, J.; Wang, W. Overview of study on metabolites and pharmacokinetic parameters of flavonoids in theaerial part of Glycyrrhiza uralensis. J. Tianjin Univ. Tradit. Chin. Med. 2022, 41, 90–101. [Google Scholar]
- Dong, L.; Liu, J.; Li, L.; Chen, X.; Shan, W.; Zhong, D. Simultaneous determination of flavonol glycoside and its major metabolites by LC-MS/MS in rat plasma. Acta Pharm. Sin. 2018, 53, 1345–1351. [Google Scholar]
- Gu, H.; Jiang, J.; Zhou, H.; Liu, Z. Pharmacokinetic study on six active ingredients of Qikui Sustained-release Tablets in rabbit plasma. Chin. Tradit. Herb. Drugs 2019, 50, 2891–2898. [Google Scholar]
- Zhang, L.; Ji, T.; Su, S.; Shang, E.; Guo, S.; Guo, J.; Qian, D.; Duan, J. Pharmacokinetics of Mori Folium flavones and alkaloids in normal and diabetic rats. China J. Chin. Mater. Med. 2017, 42, 4218–4225. [Google Scholar]
- Zhou, C. Studies on the Quality Control and Pharmacokinetics of Apocynum venetum L.; Shenyang Pharmaceutical University: Shenyang, China, 2011. [Google Scholar]
- Wang, Y.; Yuan, J.; Xiao, J.; Cai, Q.; Wang, X.; An, R.; Ma, Y. Pharmacokinetics of active ingredients of Radix Glycyrrhizae in rats with oral administration of Banxia Xiexin decoction and its different compatibilities. Chin. J. Pharm. Anal. 2012, 32, 1331–1338. [Google Scholar]
- Zhang, R.; Wang, S.; Lu, M.; Zhang, Z.; Tian, X.; Lv, D. Simultaneous determination and pharmacokinetic study of three isoflavones from Trifolium pratense extract in rat plasma by LC-MS/MS. Biomed. Chromatogr. 2015, 29, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Jin, H. Study on Serum Pharmacochemistry of Gegen Qinlian Decoction; Shanghai Jiao Tong University: Shanghai, China, 2008. [Google Scholar]
- Liu, T.; Jiang, X. Investigation of the absorption mechanisms of baicalin and baicalein in rats. J. Pharm. Sci. 2006, 95, 1326–1333. [Google Scholar] [CrossRef]
- Akao, T.; Kawabata, K.; Yanagisawa, E.; Ishihara, K.; Mizuhara, Y.; Wakui, Y.; Sakashita, Y.; Kobashi, K. Baicalin, the predominant flavone glucuronide of scutellariae radix, is absorbed from the rat gastrointestinal tract as the aglycone and restored to its original form. J. Pharm. Pharmacol. 2000, 52, 1563–1568. [Google Scholar] [CrossRef]
- Abe, K.; Inoue, O.; Yumioka, E. Biliary excretion of metabolites of baicalin and baicalein in rats. Chem. Pharm. Bull. 1990, 38, 209–211. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xiang, C.; Wang, B.; Zhao, Y.; Zhang, Q. Metabolism and excretion of total flavonoids of Semen Ziziphi Spinosae in rat urine and feces. Zhongguo Zhong Yao Za Zhi 2009, 34, 340–343. [Google Scholar]
- Liu, S.; Fen, L.; Dai, G.; Huang, G. Excretion Pathway Analysis of Limonin in Rats. Chin. J. Exp. Tradit. Med. Formulae 2017, 23, 91–94. [Google Scholar]
- Chen, Y.; Wang, J.; Jia, X.; Xin, R.; Bin, X. Advances in studies on absorption, metabolism, and drug interaction of dihydroflavones in Aurantii Fructus Immaturus. Chin. Tradit. Herb. Drugs 2010, 41, 1564–1566. [Google Scholar]
- Mccullough, M.L.; Peterson, J.J.; Patel, R.; Jacques, P.F.; Shah, R.; Dwyer, J.T. Flavonoid intake and cardiovascular disease mortality in a prospective cohort of US adults. Am. J. Clin. Nutr. 2012, 95, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Chahar, M.K.; Sharma, N.; Dobhal, M.P.; Joshi, Y.C. Flavonoids: A versatile source of anticancer drugs. Pharmacogn. Rev. 2011, 5, 1–12. [Google Scholar] [PubMed] [Green Version]
- Izumi, T.; Piskula, M.K.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kubota, Y.; Kikuchi, M. Soy Isoflavone Aglycones Are Absorbed Faster and in Higher Amounts than Their Glucosides in Humans. J. Nutr. 2000, 130, 1695–1699. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zhang, L.; Yang, L.; Cui, J.; Hou, J.; Wang, W.; Li, D. Safety Evaluation of Over-ground Parts of Glycyrrhiza uralensis. Chin. J. Exp. Tradit. Med. Formulae 2019, 25, 74–80. [Google Scholar]
- Soares, V.C.; Varanda, E.A.; Raddi, M.S. In vitro basal and metabolism-mediated cytotoxicity of flavonoids. Food Chem. Toxicol. 2006, 44, 835–838. [Google Scholar] [CrossRef]
- Perez-Coll, C.S.; Herkovits, J. Lethal and teratogenic effects of naringenin evaluated by means of an amphibian embryo toxicity test (AMPHITOX). Food Chem. Toxicol. 2004, 42, 299–306. [Google Scholar] [CrossRef]
- Matsuo, M.; Sasaki, N.; Saga, K.; Kaneko, T. Cytotoxicity of Flavonoids toward Cultured Normal Human Cells. Biol. Pharm. Bull. 2005, 28, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Walle, T.; Vincent, T.S.; Walle, U.K. Evidence of covalent binding of the dietary flavonoid quercetin to DNA and protein in human intestinal and hepatic cells. Biochem. Pharmacol. 2003, 65, 1603–1610. [Google Scholar] [CrossRef]
- Prasain, J.K.; Jones, K.; Brissie, N.; Moore, R.; Wyss, J.M.; Barnes, S. Identification of puerarin and its metabolites in rats by liquid chromatography-tandem mass spectrometry. J. Agric. Food Chem. 2004, 52, 3708–3712. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Q.; Zhu, L.; Li, Q.; Zeng, X.; Lu, L.; Hu, M.; Wang, X.; Liu, Z. Metabolic Disposition of Luteolin Is Mediated by the Interplay of UDP-Glucuronosyltransferases and Catechol-O-Methyltransferases in Rats. Drug Metab. Dispos. 2017, 45, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Kulling, S.E.; Honig, D.M.; Metzler, M. Oxidative metabolism of the soy isoflavones daidzein and genistein in humans in vitro and in vivo. J. Agric. Food Chem. 2001, 49, 3024–3033. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H.; van der Wildt, J.; Kinzel, J.; Attalla, H.; Wähäla, K.; Mäkelä, T.; Hase, T.; Fotsis, T. Lignan and isoflavonoid conjugates in human urine. J. Steroid Biochem. Mol. Biol. 1995, 52, 97–103. [Google Scholar] [CrossRef]
- Wang, S.; Gong, T.; Lu, J.; Kano, Y.; Yuan, D. Simultaneous determination of tectorigenin and its metabolites in rat plasma by ultra performance liquid chromatography/quadrupole time-of-flight mass spectrometry. J. Chromatogr. B 2013, 933, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lin, H.; Hu, M. Metabolism of Flavonoids via Enteric Recycling: Role of Intestinal Disposition. J. Pharmacol. Exp. Ther. 2002, 304, 1228–1235. [Google Scholar] [CrossRef]
- Zhang, L.; Zuo, Z.; Lin, G. Intestinal and Hepatic Glucuronidation of Flavonoids. Mol. Pharm. 2007, 4, 833–845. [Google Scholar] [CrossRef]
- Breinholt, V.M.; Offord, E.A.; Brouwer, C.; Nielsen, S.E.; Brosen, K.; Friedberg, T. In vitro investigation of cytochrome P450-mediated metabolism of dietary flavonoids. Food Chem. Toxicol. 2002, 40, 609–616. [Google Scholar] [CrossRef]
- de Vries, J.H.; Hollman, P.C.; Meyboom, S.; Buysman, M.N.; Zock, P.L.; van Staveren, W.A.; Katan, M.B. Plasma concentrations and urinary excretion of the antioxidant flavonols quercetin and kaempferol as biomarkers for dietary intake. Am. J. Clin. Nutr. 1998, 68, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Marsh, C.A.; Alexander, F.; Levvy, G.A. Glucuronide decomposition in the digestive tract. Nature 1952, 170, 163–164. [Google Scholar] [CrossRef]
- Yan, X.; Jiang, H.; Zeng, S. Advances in the study of metabolism and drug-drug interactions of flavonoids in prmiary cultured hepatocytes. Acta Pharm. Sin. 2006, 41, 1130–1135. [Google Scholar]
- Serra, A.; Rubió, L.; Borràs, X.; Macià, A.; Romero, M.-P.; Motilva, M.-J. Distribution of olive oil phenolic compounds in rat tissues after administration of a phenolic extract from olive cake. Mol. Nutr. Food Res. 2011, 56, 486–496. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, S.; Manna, C.; Migliardi, V.; Mazzoni, O.; Morrica, P.; Capasso, G.; Pontoni, G.; Galletti, P.; Zappia, V. Pharmacokinetics and metabolism of hydroxytyrosol, a natural antioxidant from olive oil. Drug Metab. Dispos. 2001, 29, 1492–1498. [Google Scholar] [PubMed]
- Pike, A.C.; Brzozowski, A.M.; Hubbard, R.E.; Bonn, T.; Thorsell, A.G.; Engström, O.; Ljunggren, J.; Gustafsson, J.A.; Carlquist, M. Structure of the ligand-binding domain of oestrogen receptor beta in the presence of a partial agonist and a full antagonist. EMBO J. 1999, 18, 4608–4618. [Google Scholar] [CrossRef] [Green Version]
- Halabalaki, M.; Alexi, X.; Aligiannis, N.; Lambrinidis, G.; Pratsinis, H.; Florentin, I.; Mitakou, S.; Mikros, E.; Skaltsounis, A.-L.; Alexis, M.N. Estrogenic Activity of Isoflavonoids fromOnobrychis ebenoides. Planta Medica 2006, 72, 488–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.; Walle, T. Methylated Flavonoids Have Greatly Improved Intestinal Absorption and Metabolic Stability. Drug Metab. Dispos. 2006, 34, 1786–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebert, D.W.; Dieter, M.Z. The evolution of drug metabolism. Pharmacology 2000, 61, 124–135. [Google Scholar] [CrossRef] [PubMed]
- van der Weide, J.; Steijns, L.S. Cytochrome P450 enzyme system: Genetic polymorphisms and impact on clinical pharmacology. Ann. Clin. Biochem. 1999, 36, 722–729. [Google Scholar] [CrossRef] [Green Version]
- Ciolino, H.P.; Wang, T.T.; Yeh, G.C. Diosmin and diosmetin are agonists of the aryl hydrocarbon receptor that differentially affect cytochrome P450 1A1 activity. Cancer Res. 1998, 58, 2754–2760. [Google Scholar]
- Ciolino, H.P.; Yeh, G.C. The flavonoid galangin is an inhibitor of CYP1A1 activity and an agonist/antagonist of the aryl hydrocarbon receptor. Br. J. Cancer 1999, 79, 1340–1346. [Google Scholar] [CrossRef] [Green Version]
- Stopper, H.; Schmitt, E.; Kobras, K. Genotoxicity of phytoestrogens. Mutat. Res. Mol. Mech. Mutagen. 2005, 574, 139–155. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, J.; Yan, H.; Shi, M.; Zheng, Q.; Wang, Y.; Zhu, Y.; Miao, L.; Gao, X. Kaempferol inhibits benign prostatic hyperplasia by resisting the action of androgen. Eur. J. Pharmacol. 2021, 907, 174251. [Google Scholar] [CrossRef] [PubMed]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
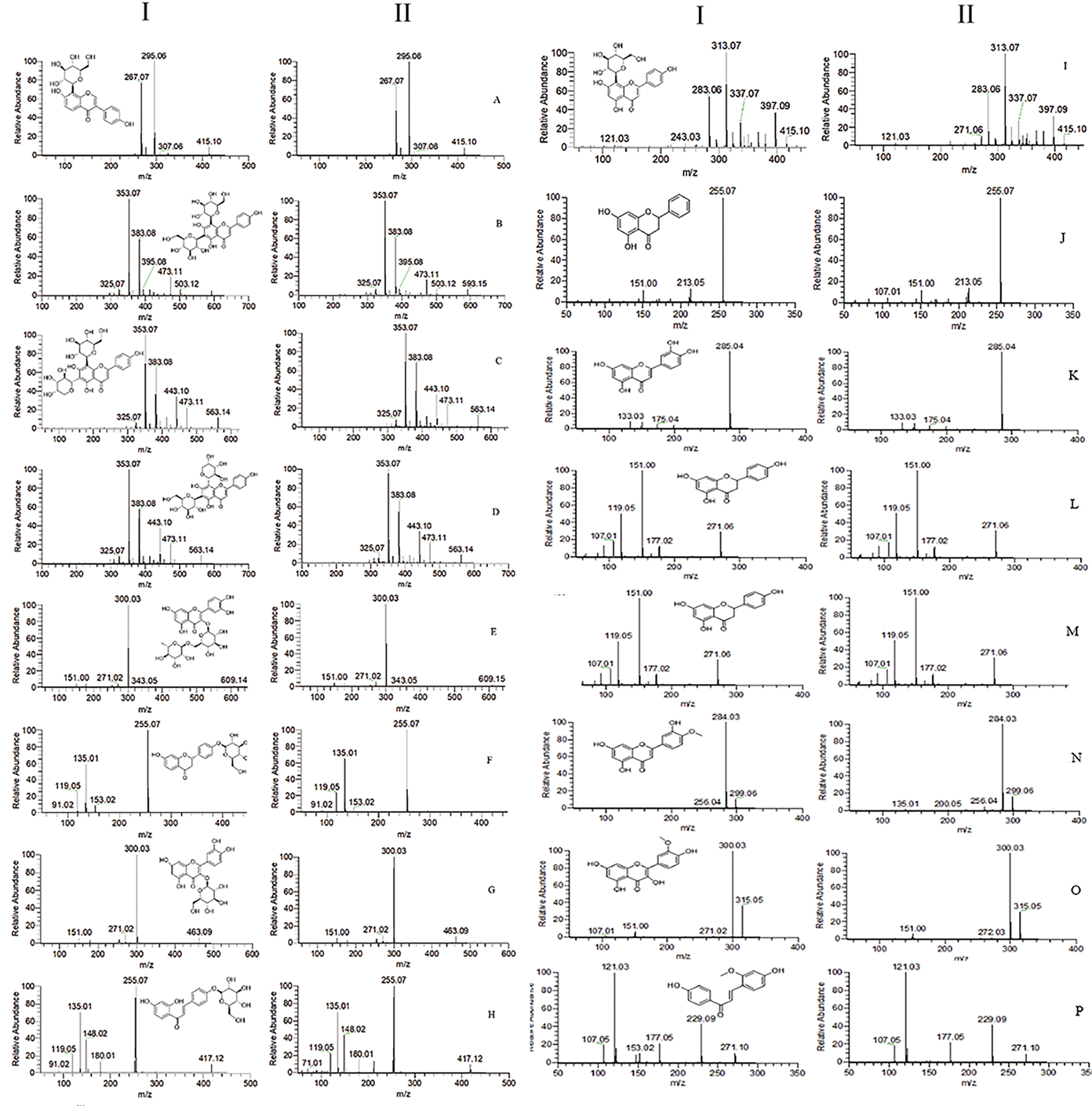
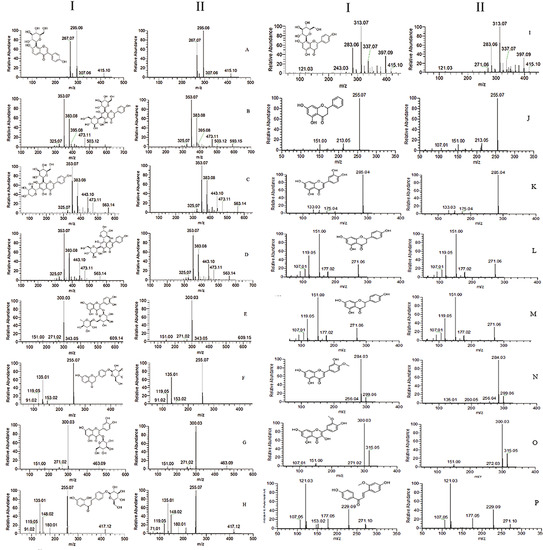
| No. | RT [min] | Name | Formula | Predicted | Measured | HR-MS | MS2 | ∆ (ppm) | Plasma | Prostate | Urine | Feces |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 33.922 | 8-Prenylnaringenin [9] | C20H20O5 | 339.1227 | 339.1231 | [M − H]− | 219, 133, 119 | −0.44 | + | |||
| 2 | 22.566 | Astragalin [7] | C21H20O11 | 447.0922 | 447.0925 | [M − H]− | 285, 284, 255, 227, 151 | 0.21 | + | + | + | |
| 3 | 23.924 | Baicalin [7] | C15H10O5 | 269.0445 | 269.0451 | [M − H]− | 241, 223, 197, 169, 136 | 1.14 | + | + | ||
| 4 | 25.602 | Calycosin [10] | C16H12O5 | 285.0758 | 285.0764 | [M + H]+ | 270, 253, 225, 137 | −0.78 | + | |||
| 5 | 27.92 | Diosmetin * | C16H12O6 | 299.0550 | 299.0555 | [M − H]− | 284, 256, 227, 180, 151 | −0.63 | + | + | + | + |
| 6 | 22.633 | Diosmin [11] | C28H32O15 | 609.1814 | 609.1833 | [M + H]+ | 463, 301, 286, 258, 85 | 0.85 | + | + | ||
| 7 | 27.498 | Genistein * | C15H10O5 | 269.0451 | 269.0444 | [M − H]− | 225, 201, 181, 151, 133 | −0.49 | + | + | + | |
| 8 | 22.733 | Hesperidin [11] | C28H34O15 | 609.1814 | 609.1818 | [M − H]− | 325, 301, 286, 242, 164, 151, 125 | 0.6 | + | |||
| 9 | 21.784 | Hyperoside [12] | C21H20O12 | 463.0871 | 463.0894 | [M − H]− | 300, 271, 255, 243, 151 | 0.52 | + | + | ||
| 10 | 21.476 | Isoliquiritigenin * | C15H12O4 | 257.0808 | 257.0812 | [M + H]+ | 242, 211, 147, 137, 119, 91, 81 | −1.03 | + | + | ||
| 11 | 36.151 | Isoliquiritin * | C21H22O9 | 417.1180 | 417.1183 | [M − H]− | 255, 180, 148, 135, 119, 108, 91 | 0.16 | + | + | ||
| 12 | 23.188 | Isoquercitrin * | C21H20O12 | 463.0871 | 463.0875 | [M − H]− | 300, 271, 179, 151 | 2.62 | + | + | + | |
| 13 | 21.683 | Isorhamnetin * | C16H12O7 | 315.0499 | 315.0505 | [M − H]− | 300, 283, 255, 151, 107 | 0.15 | + | + | ||
| 14 | 22.252 | Isorhamnetin-3-O-nehesperidine [13] | C28H32O16 | 625.1763 | 625.1777 | [M + H]+ | 317, 302, 85, 71 | 0.35 | + | |||
| 15 | 22.257 | Isorhamnetin-3-O-rutinoside [13] | C28H32O16 | 623.1607 | 623.1608 | [M − H]− | 417, 314, 299, 255 | 0.27 | + | |||
| 16 | 20.61 | Isoschaftoside * | C26H28O14 | 563.1395 | 563.1396 | [M − H]− | 473, 443, 413, 383, 353, 325, 191 | 0.21 | + | |||
| 17 | 18.738 | Isovitexin [14] | C21H20O10 | 433.1129 | 433.1138 | [M + H]+ | 415, 397, 379, 367, 337, 313, 283 | 0.12 | + | + | ||
| 18 | 22.557 | Kaempferol [14] | C15H10O6 | 287.0550 | 287.0556 | [M + H]+ | 258, 165, 153, 121 | −0.14 | + | + | + | + |
| 19 | 21.864 | Kaempferol3-glucorhamnoside [14] | C27H30O15 | 593.1501 | 593.1505 | [M − H]− | 285, 284, 255, 229, 227, 151 | 0.05 | + | |||
| 20 | 45.936 | Liquiritin * | C21H22O9 | 417.1180 | 417.1185 | [M − H]− | 255, 153, 135, 119, 91 | 0.3 | + | + | ||
| 21 | 3.812 | Luteolin * | C15H10O6 | 285.0394 | 285.0399 | [M − H]− | 257, 241, 199, 151 | −0.4 | + | + | + | + |
| 22 | 29.342 | Naringenin * | C15H12O5 | 271.0601 | 271.0607 | [M − H]− | 177, 165, 151, 119, 107, 93, 83, 65 | 0.08 | + | + | ||
| 23 | 22.511 | Naringin [15,16] | C27H32O14 | 579.1708 | 579.1711 | [M − H]− | 313, 271, 151, 119, 107 | −0.28 | + | |||
| 24 | 33.203 | Ononin [17] | C22H22O9 | 431.1337 | 431.1346 | [M + H]+ | 269, 254, 237, 213, 118, 107 | −0.37 | + | + | ||
| 25 | 20.822 | Orientin [17] | C21H20O11 | 447.0922 | 447.0926 | [M − H]− | 357, 327, 297, 285, 269, 217, 151 | 0.18 | + | |||
| 26 | 21.68 | Pinocembrin * | C15H12O4 | 255.0652 | 255.0657 | [M − H]− | 213, 151, 107, 83 | −1.08 | + | + | + | + |
| 27 | 19.604 | Puerarin * | C21H20O9 | 415.1024 | 415.1027 | [M − H]− | 361, 307, 295, 277, 267, 253, 109 | −0.03 | + | + | + | |
| 28 | 21.779 | Quercetin [18] | C15H10O7 | 301.0343 | 301.0347 | [M − H]− | 273, 178, 151, 121 | −1.02 | + | + | + | |
| 29 | 27.347 | Retrochalcone * | C16H14O4 | 271.0965 | 271.0967 | [M + H]+ | 229, 177, 121, 107 | −0.8 | + | + | + | + |
| 30 | 21.345 | Rutin * | C27H30O16 | 609.1450 | 609.1452 | [M − H]− | 300, 271, 255, 244, 178, 151 | 0.07 | + | + | ||
| 31 | 20.398 | Schaftoside * | C26H28O14 | 563.1395 | 563.1396 | [M − H]− | 473, 443, 413, 383, 353, 325, 191 | −0.12 | + | + | ||
| 32 | 19.745 | Vicenin-2 * | C27H30O15 | 593.1501 | 593.1502 | [M − H]− | 503, 473, 383, 353, 325, 297 | 0.11 | + | |||
| 33 | 17.75 | Vitexin * | C21H20O10 | 433.1129 | 433.1128 | [M + H]+ | 415, 397, 337, 323, 313, 283, 121 | 0.08 | + | |||
| 34 | 25.965 | Wogonin [19] | C16H12O5 | 285.0758 | 285.0764 | [M − H]− | 270, 253, 177, 150 | −0.77 | + | + | + |
| No. | Formula | RT [min] | Δ (ppm) | Calc. MW | HR-MS | MS | Identification | Plasma | Urine | Feces | Prostate |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 * | C21H20O9 | 19.592 | 0.53 | 416.11095 | [M − H]− | 295, 277, 267, 253 | Puerarin | + | + | + | + |
| 1-M1 | C15H10O4 | 24.857 | 0.18 | 254.238 | [M + H]+ | 145, 137 | Daidzein | ||||
| 1-M2 [23] | C21H20O10 | 21.683 | −0.28 | 432.10552 | [M + H]+ | 415, 313, 283, 267 | D-Me-Glu | + | + | + | + |
| 1-M3 [23] | C22H20O10 | 24.136 | −0.19 | 444.10556 | [M − H]− | 267, 253 | D-Me-GluA | + | |||
| 1-M4 [23] | C21H18O10 | 20.618 | −0.15 | 430.08993 | [M − H]− | 253 | Daidzein-7-O-glucuronid | + | + | ||
| 1-M5 [24] | C22H22O10 | 19.913 | 0.26 | 446.12141 | [M + H]+ | 327, 299 | 3′-Methoxy Puerarin | + | + | ||
| 1-M6 [24] | C27H30O14 | 21.253 | 0.06 | 578.16359 | [M − H]− | 457 | Puerarin-Glu | + | + | + | |
| 1-M7 [25] | C21H20O9 | 19.673 | 0.31 | 416.11068 | [M − H]− | 295 | Daidzin | + | + | ||
| 1-M8 [25] | C21H20O12S | 23.896 | 0.19 | 496.06764 | [M − H]− | 415 | puerarin 4′-O-β-sulfate | + | |||
| 1-M9 [25] | C27H28O15 | 19.177 | 0.72 | 592.14325 | [M − H]− | 415, 295, 253 | puerarin-7-O-glucuronide | + | |||
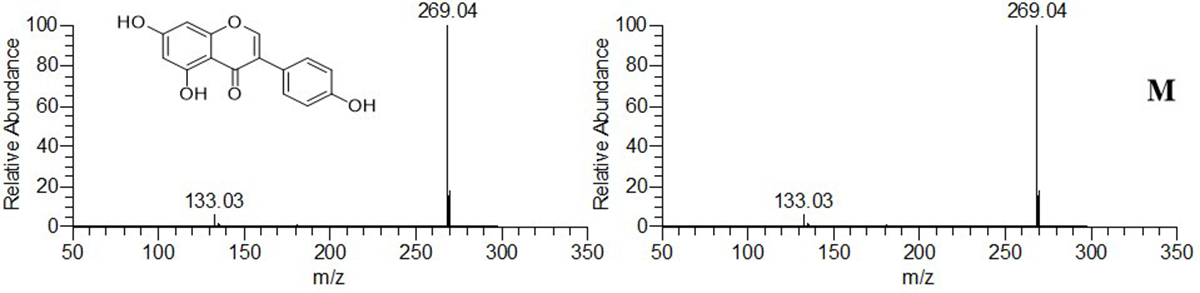
| 2 * | C15H10O6 | 24.739 | −0.52 | 286.04759 | [M − H]− | 257, 241, 151, 137, 135 | Kaempferol | + | + | + | |
| 2-M1 [26] | C15H10O7 | 25.616 | −0.72 | 302.04244 | [M + H]+ | 284, 273 | Quercetin | + | + | ||
| 2-M2 [27] | C16H12O7 | 27.27 | 0.2 | 316.262 | [M + H]+ | 300 | Isorhamnetin | + | + | ||
| 2-M3 [28] | C15H10O10S | 24.754 | 0.15 | 381.99953 | [M − H]− | 301, 284, 151 | Q-S | + | |||
| 2-M4 [28] | C21H18O13 | 21.710 | 1.37 | 478.07539 | [M − H]− | 301, 284, 151 | Q-GluA | + | |||
| 2-M5 [29] | C15H10O9S | 24.349 | −1.23 | 366.0041 | [M − H]− | 285, 257, 151, 137 | K-S | + | + | + | |
| 2-M6 [29] | C15H10O9S | 24.643 | −0.14 | 366.0045 | [M − H]− | 285, 257, 151, 137 | K-S | + | + | + | |
| 2-M7 [30] | C16H12O6 | 27.960 | −1.28 | 300.06301 | [M − H]− | 284, 151 | K-Me | + | + | + | + |
| 2-M8 [29] | C21H18O12 | 22.864 | −0.37 | 462.07966 | [M − H]− | 285, 268, 240 | K-GluA | + | + | + | + |
| 3 * | C15H10O6 | 27.34 | −0.72 | 286.04753 | [M + H]+ | 269, 259, 177, 153 | Luteolin | + | + | + | + |
| 3-M1 [31] | C16H12O6 | 29.310 | −0.62 | 300.0632 | [M − H]− | 285, 256 | L-Me | + | + | + | + |
| 3-M2 [31] | C15H10O9S | 24.599 | −0.16 | 366.00449 | [M − H]− | 285 | L-S | + | + | + | |
| 3-M3 [31] | C21H18O12 | 24.919 | 1.14 | 462.07964 | [M + H]+ | 287 | L-7-GluA | + | + | ||
| 3-M4 [31] | C21H18O12 | 22.864 | −0.81 | 462.07945 | [M + H]+ | 287 | L-4′-GluA | + | + | + | |
| 3-M5 [31] | C21H18O12 | 22.861 | 1.44 | 462.08049 | [M + H]+ | 287 | L-3′-GluA | + | + | ||
| 3-M6 | C15H10O7 | 25.656 | −0.66 | 302.04245 | [M − H]− | 273, 151 | L+O | + | + | + | |
| 4 * | C15H12O4 | 21.68 | −1.08 | 256.07328 | [M + H]+ | 153, 131, 103, 97 | Pinocembrin | + | + | + | + |
| 4-M1 [32] | C15H12O5 | 33.179 | −0.34 | 272.06838 | [M − H]− | 151, 119 | Naringenin | + | + | + | + |
| 4-M2 | C15H12O8S | 27.385 | −0.28 | 352.02519 | [M − H]− | 271, 151, 119 | N+S | + | + | + | |
| 4-M3 | C21H20O11 | 22.978 | 0.08 | 448.1006 | [M − H]− | 271, 135, 115 | N+GluA | + | + | ||
| 4-M4 | C15H12O7S | 23.124 | 0.38 | 336.0305 | [M − H]− | 255, 135, 119 | P+S | + | + | + | + |
| 4-M5 | C16H14O4 | 25.508 | −0.53 | 270.08907 | [M − H]− | 254 | P+Me | + | + | + | + |
| 4-M6 [32] | C21H20O10 | 21.630 | −0.46 | 432.10545 | [M − H]− | 255, 135, 119 | P+GluA | + | + | + | + |
| 4-M7 [32] | C15H12O7 | 23.172 | 2.9 | 306.07484 | [M − H]− | 167 | 5,6,7-Trihydroxyflavanone | + | + | ||
| 4-M8 [32] | C15H12O7 | 25.459 | −0.37 | 306.07384 | [M − H]− | 167 | 5,7,8-Trihydroxyflavanone | + | + |
| Sample | Components | Test Range (ng·mL−1/ng·g−1) | Regression Equation | r2 | Lower Limit of Quantification (ng·mL−1/ng·g−1) |
|---|---|---|---|---|---|
| Plasma | Puerarin | 0.5–100 | y = 0.006x + 0.0121 | 0.9994 | 0.5 |
| Kaempferol | 0.2–5 | y = 0.0012x + 0.0003 | 0.9981 | 0.2 | |
| Luteolin | 0.2–10 | y = 0.1701x − 0.0248 | 0.9994 | 0.2 | |
| Pinocembrin | 0.1–2 | y = 0.0137x + 0.001 | 1 | 0.1 | |
| Prostate | Puerarin | 0.1–5 | y = 0.0151x + 0.0028 | 0.9996 | 0.1 |
| Kaempferol | 1–200 | y = 0.0003x + 0.0004 | 0.9999 | 1 | |
| Luteolin | 0.2–20 | y = 0.0072x + 0.0021 | 0.9998 | 0.2 | |
| Pinocembrin | 0.2–100 | y = 0.0026x + 0.0035 | 0.9984 | 0.2 |
| Sample | Component | Concentration (μg·mL−1/μg·g−1) | Matrix Effect ( ± s, %) | Precision (%, RSD) | ( ± s, %) | Precision (%, RSD) |
|---|---|---|---|---|---|---|
| Plasma | Puerarin | 0.5 | 102.703 ± 3.096 | 3.015 | 105.855 ± 9.174 | 8.666 |
| 10 | 93.878 ± 2.903 | 3.092 | 95.308 ± 6.054 | 6.352 | ||
| 50 | 94.895 ± 1.737 | 1.831 | 99.462 ± 1.576 | 1.584 | ||
| Luteolin | 0.2 | 105.091 ± 1.373 | 1.306 | 96.367 ± 7.273 | 7.547 | |
| 2 | 94.444 ± 0.275 | 0.291 | 109.244 ± 0.77 | 0.705 | ||
| 5 | 96.414 ± 8.307 | 8.616 | 107.082 ± 2.178 | 2.034 | ||
| Kaempferol | 0.1 | 103.623 ± 3.494 | 3.372 | 100.6999 ± 3.782 | 3.756 | |
| 0.5 | 109.066 ± 4.723 | 4.330 | 99 ± 2.218 | 2.304 | ||
| 2 | 88.801 ± 3.221 | 3.627 | 102.664 ± 2.461 | 2.397 | ||
| Pinocembrin | 0.1 | 82.308 ± 1.538 | 2.128 | 99.291 ± 5.258 | 5.285 | |
| 0.5 | 101.507 ± 1.087 | 1.071 | 99.865 ± 1.039 | 1.040 | ||
| 2 | 95.215 ± 1.031 | 1.082 | 106.759 ± 2.344 | 2.196 | ||
| Prostate | Puerarin | 0.100 1.000 5.000 | 92.691 ± 3.453 90.890 ± 5.368 96.419 ± 1.076 | 3.725 5.906 1.116 | 98.925 ± 2.845 104.914 ± 6.361 99.325 ± 1.116 | 2.876 6.063 1.123 |
| Luteolin | 0.200 5.000 10.000 | 98.342 ± 5.458 101.592 ± 1.914 100.336 ± 0.302 | 5.550 1.884 0.301 | 101.686 ± 4.141 96.951 ± 1.258 98.963 ± 0.380 | 4.072 1.298 0.384 | |
| Kaempferol | 2.000 100.000 200.000 | 99.660 ± 1.640 98.652 ± 0.969 105.671 ± 1.823 | 1.646 0.983 1.725 | 104.266 ± 3.086 96.681 ± 0.538 96.512 ± 1.562 | 2.960 0.556 1.618 | |
| Pinocembrin | 2.000 50.000 100.000 | 98.142 ± 2.285 96.251 ± 1.169 93.291 ± 1.108 | 2.328 1.214 1.188 | 92.255 ± 3.320 96.911 ± 2.216 97.388 ± 0.619 | 3.598 2.287 0.636 |
| Sample | Component | Concentration (μg·mL−1/μg·g−1) | Intra-Day | Inter-Day | ||||
|---|---|---|---|---|---|---|---|---|
| Mean ± S.D. (μg·mL−1/μg·g−1) | Accuracy (%, RE) | Precision (%, RSD) | Mean ± S.D. (μg·mL−1/μg·g−1) | Accuracy | Precision | |||
| (%, RE) | (%, RSD) | |||||||
| Plasma | Puerarin | 0.500 | 0.480 ± 0.03 | −0.042 | 6.250 | 0.477 ± 0.047 | −0.049 | 9.914 |
| 10.000 | 10.493 ± 0.92 | 0.047 | 8.770 | 11.9 ± 0.436 | 0.160 | 3.663 | ||
| 50.000 | 49.033 ± 2.957 | −0.020 | 6.030 | 48.2 ± 1.179 | −0.037 | 2.446 | ||
| Luteolin | 0.200 | 0.233 ± 0.045 | 0.143 | 19.325 | 0.213 ± 0.026 | 0.061 | 12.034 | |
| 2.000 | 2.273 ± 0.118 | 0.120 | 5.211 | 2.137 ± 0.222 | 0.064 | 10.385 | ||
| 5.000 | 4.877 ± 0.172 | −0.025 | 3.530 | 5.217 ± 0.259 | 0.042 | 4.963 | ||
| Kaempferol | 0.100 | 0.126 ± 0.039 | 0.206 | 18.615 | 0.127 ± 0.003 | 0.215 | 2.399 | |
| 0.500 | 0.523 ± 0.023 | 0.045 | 4.413 | 0.49 ± 0.012 | −0.020 | 2.355 | ||
| 2.000 | 1.973 ± 0.012 | −0.014 | 0.585 | 2.05 ± 0.07 | 0.024 | 3.415 | ||
| Pinocembrin | 0.100 | 0.137 ± 0.012 | 0.272 | 8.778 | 0.94 ± 0.044 | 0.894 | 4.637 | |
| 0.500 | 5.493 ± 0.224 | 0.909 | 4.072 | 0.497 ± 0.032 | −0.007 | 6.472 | ||
| 2.000 | 2.127 ± 0.061 | 0.060 | 2.873 | 2.203 ± 0.085 | 0.092 | 3.860 | ||
| Prostate | Puerarin | 0.100 1.000 5.000 | 0.088 ± 0.004 1.073 ± 0.142 4.973 ± 0.125 | −0.132 0.068 −0.005 | 4.286 13.220 2.514 | 0.107 ± 0.014 1.070 ± 0.123 4.873 ± 0.134 | 0.068 0.065 −0.026 | 13.220 11.484 2.756 |
| Luteolin | 0.200 5.000 10.000 | 0.197 ± 0.011 5.330 ± 0.085 9.490 ± 0.070 | −0.014 0.062 −0.054 | 5.651 1.603 0.738 | 0.192 ± 0.003 4.590 ± 0.087 11.100 ± 0.700 | −0.042 −0.089 0.099 | 1.378 1.899 6.306 | |
| Kaempferol | 2.000 100.000 200.000 | 2.300 ± 0.200 96.533 ± 2.658 203.467 ± 0.503 | 0.130 −0.036 0.017 | 8.696 2.753 0.247 | 2.000 ± 0.087 102.700 ± 3.161 202.633 ± 2.303 | 0.000 0.026 0.013 | 4.359 3.078 1.136 | |
| Pinocembrin | 2.000 50.000 100.000 | 2.087 ± 0.078 50.867 ± 1.484 103.867 ± 1.185 | 0.042 0.017 0.037 | 3.722 2.918 1.141 | 1.940 ± 0.044 49.933 ± 3.166 102.900 ± 0.608 | −0.031 −0.001 0.028 | 2.247 6.340 0.591 | |
| Sample | Component | Concentration (μg·mL−1/μg·g−1) | 24 h at Room Temperature Freeze-Thaw Cycles | |||||
|---|---|---|---|---|---|---|---|---|
| Mean ± S.D. | Accuracy | Precision | Mean ± S.D. | Accuracy | Precision | |||
| (μg·mL−1/μg·g−1) | (%, RE) | (%, RSD) | (μg·mL−1/μg·g−1) | (%, RE) | (%, RSD) | |||
| Plasma | Puerarin | 0.500 | 0.493 ± 0.015 | −0.014 | 3.096 | 0.517 ± 0.01 | 0.033 | 2.019 |
| 10.000 | 10.217 ± 0.07 | 0.021 | 0.687 | 10.333 ± 0.902 | 0.032 | 8.728 | ||
| 50.000 | 49.967 ± 1.159 | −0.001 | 2.320 | 50.667 ± 1.266 | 0.013 | 2.499 | ||
| Luteolin | 0.200 | 0.24 ± 0.026 | 0.167 | 11.024 | 0.2 ± 0.018 | 0.000 | 8.789 | |
| 2.000 | 2.13 ± 0.0171 | 0.061 | 8.023 | 2.29 ± 0.098 | 0.127 | 4.301 | ||
| 5.000 | 4.903 ± 0.091 | −0.020 | 1.851 | 4.927 ± 0.055 | −0.015 | 1.118 | ||
| Kaempferol | 0.100 | 0.102 ± 0.015 | 0.023 | 14.959 | 0.088 ± 0.004 | −0.131 | 3.960 | |
| 0.500 | 0.473 ± 0.035 | −0.056 | 7.419 | 0.511 ± 0.019 | 0.022 | 3.624 | ||
| 2.000 | 2.2 ± 0.056 | 0.091 | 2.531 | 2.047 ± 0.144 | 0.023 | 7.018 | ||
| Pinocembrin | 0.100 | 0.117 ± 0.012 | 0.143 | 9.897 | 0.103 ± 0.015 | 0.032 | 14.495 | |
| 0.500 | 0.51 ± 0.02 | 0.020 | 3.922 | 0.503 ± 0.021 | 0.007 | 4.136 | ||
| 2.000 | 2.01 ± 0.082 | 0.005 | 4.072 | 1.920 ± 0.026 | −0.042 | 1.378 | ||
| Prostate | Puerarin | 0.100 1.000 5.000 | 0.973 ± 0.049 1.097 ± 0.138 4.940 ± 0.056 | 0.897 0.088 −0.012 | 5.068 12.580 1.127 | 0.100 ± 0.005 0.973 ± 0.042 5.057 ± 0.224 | 0.000 −0.027 0.011 | 5.292 4.277 4.423 |
| Luteolin | 0.200 5.000 10.000 | 0.194 ± 0.005 4.963 ± 0.118 9.900 ± 0.458 | −0.033 −0.007 −0.010 | 2.328 2.387 4.629 | 0.213 ± 0.031 4.833 ± 0.058 10.433 ± 0.551 | 0.062 −0.034 0.042 | 14.321 1.195 5.279 | |
| Kaempferol | 2.000 100.000 200.000 | 2.000 ± 0.265 99.133 ± 3.557 201.667 ± 3.215 | 0.000 −0.009 0.008 | 13.229 3.588 1.594 | 1.833 ± 0.058 96.667 ± 2.082 202.333 ± 2.082 | −0.091 −0.034 0.012 | 3.149 2.153 1.029 | |
| Pinocembrin | 2.000 50.000 100.000 | 1.923 ± 0.076 51.000 ± 3.000 102.000 ± 2.646 | −0.040 0.020 0.020 | 3.971 5.882 2.594 | 1.867 ± 0.153 51.333 ± 1.528 102.333 ± 2.887 | −0.071 0.026 0.023 | 8.183 2.976 2.821 | |
| Parameters | t1/2 | Tmax | Cmax | AUC0–t | AUC0–∞ | MRT0–t | MRT0–∞ |
|---|---|---|---|---|---|---|---|
| (h) | (h) | (ng/mL) | (h×ng/mL) | (h×ng/mL) | (h) | (h) | |
| Puerarin | 6.43 ± 0.20 | 0.50 ± 0.04 | 23.76 ± 1.05 | 238.05 ± 23.35 | 241.86 ± 2.45 | 14.40 ± 1.21 | 15.08 ± 1.71 |
| Luteolin | 31.08 ± 1.17 | 0.87 ± 0.05 | 185 ± 0.12 | 35.01 ± 0.81 | 51.63 ± 1.98 | 20.74 ± 1.91 | 43.95 ± 1.72 |
| Kaempferol | 18.98 ± 1.46 | 4.00 ± 0.17 | 1.27 ± 0.06 | 18.75 ± 1.18 | 23.79 ± 0.86 | 20.12 ± 0.84 | 31.82 ± 1.22 |
| Pinocembrin | 13.18 ± 0.72 | 1.50 ± 0.05 | 1.18 ± 0.06 | 21.78 ± 0.73 | 23.84 ± 1.29 | 19.12 ± 0.82 | 23.26 ± 0.83 |
| Name | Q1 | Q3 | DP | CE |
|---|---|---|---|---|
| Puerarin | 415.1 | 267.1 | −150 | −46 |
| Kaempferol | 285.1 | 117 | −150 | −54 |
| Luteolin | 285 | 133.1 | −135 | −46 |
| Pinocembrin | 255 | 213 | −120 | −27 |
| NEG-IS-NI | 417 | 122 | −100 | −30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Chang, G.; Wang, W.; Ji, Z.; Cui, J.; Peng, Y. Pharmacokinetics, Prostate Distribution and Metabolic Characteristics of Four Representative Flavones after Oral Administration of the Aerial Part of Glycyrrhiza uralensis in Rats. Molecules 2022, 27, 3245. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103245
Liu H, Chang G, Wang W, Ji Z, Cui J, Peng Y. Pharmacokinetics, Prostate Distribution and Metabolic Characteristics of Four Representative Flavones after Oral Administration of the Aerial Part of Glycyrrhiza uralensis in Rats. Molecules. 2022; 27(10):3245. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103245
Chicago/Turabian StyleLiu, Haifan, Guanhua Chang, Wenquan Wang, Zuen Ji, Jie Cui, and Yifeng Peng. 2022. "Pharmacokinetics, Prostate Distribution and Metabolic Characteristics of Four Representative Flavones after Oral Administration of the Aerial Part of Glycyrrhiza uralensis in Rats" Molecules 27, no. 10: 3245. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103245