
N-Butyrylated Hyaluronic Acid Achieves Anti-Inflammatory Effects In Vitro and in Adjuvant-Induced Immune Activation in Rats



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
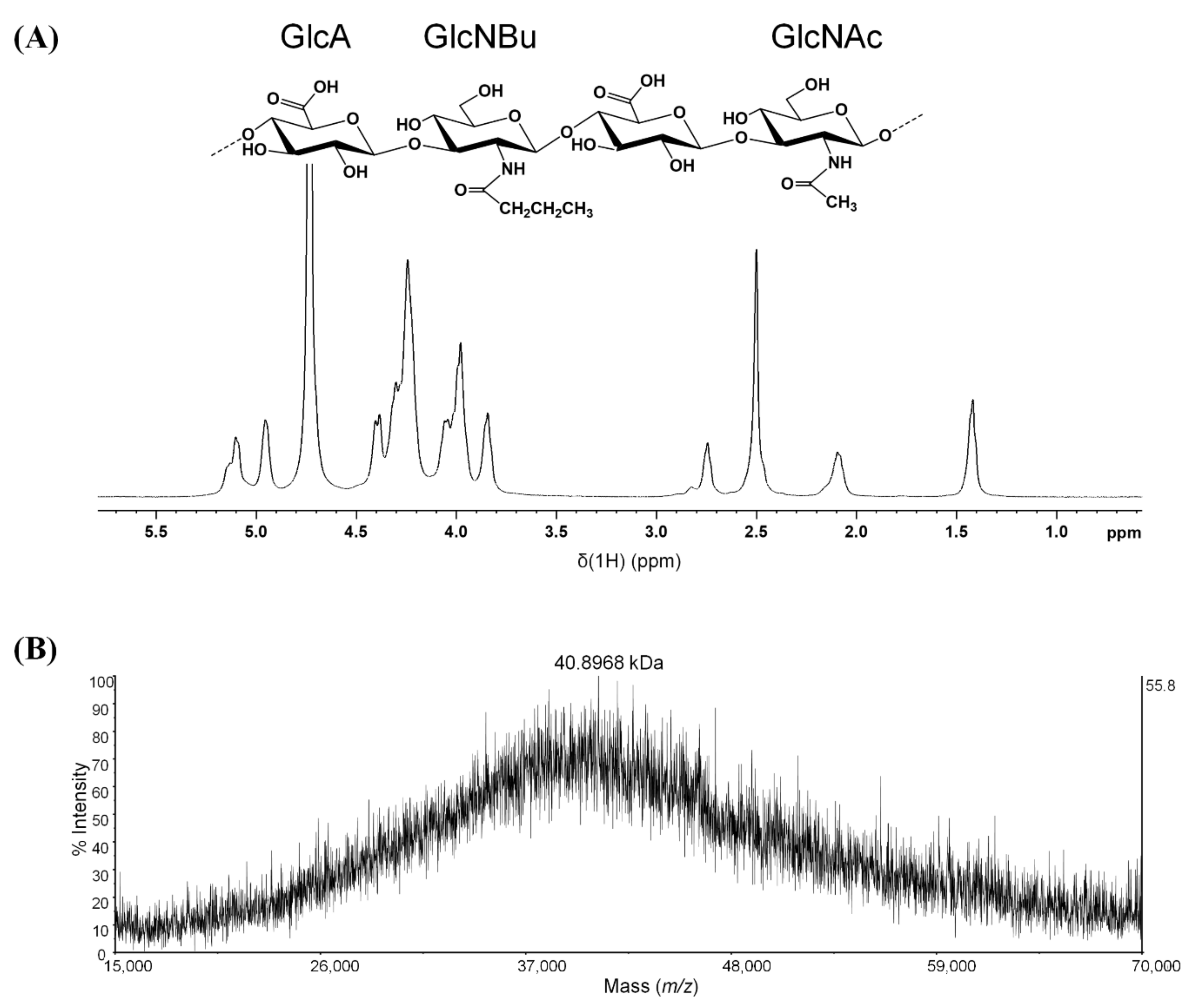
2.1. Characterization of Synthesized BHA
2.2. Effects of BHA on Cell Proliferation
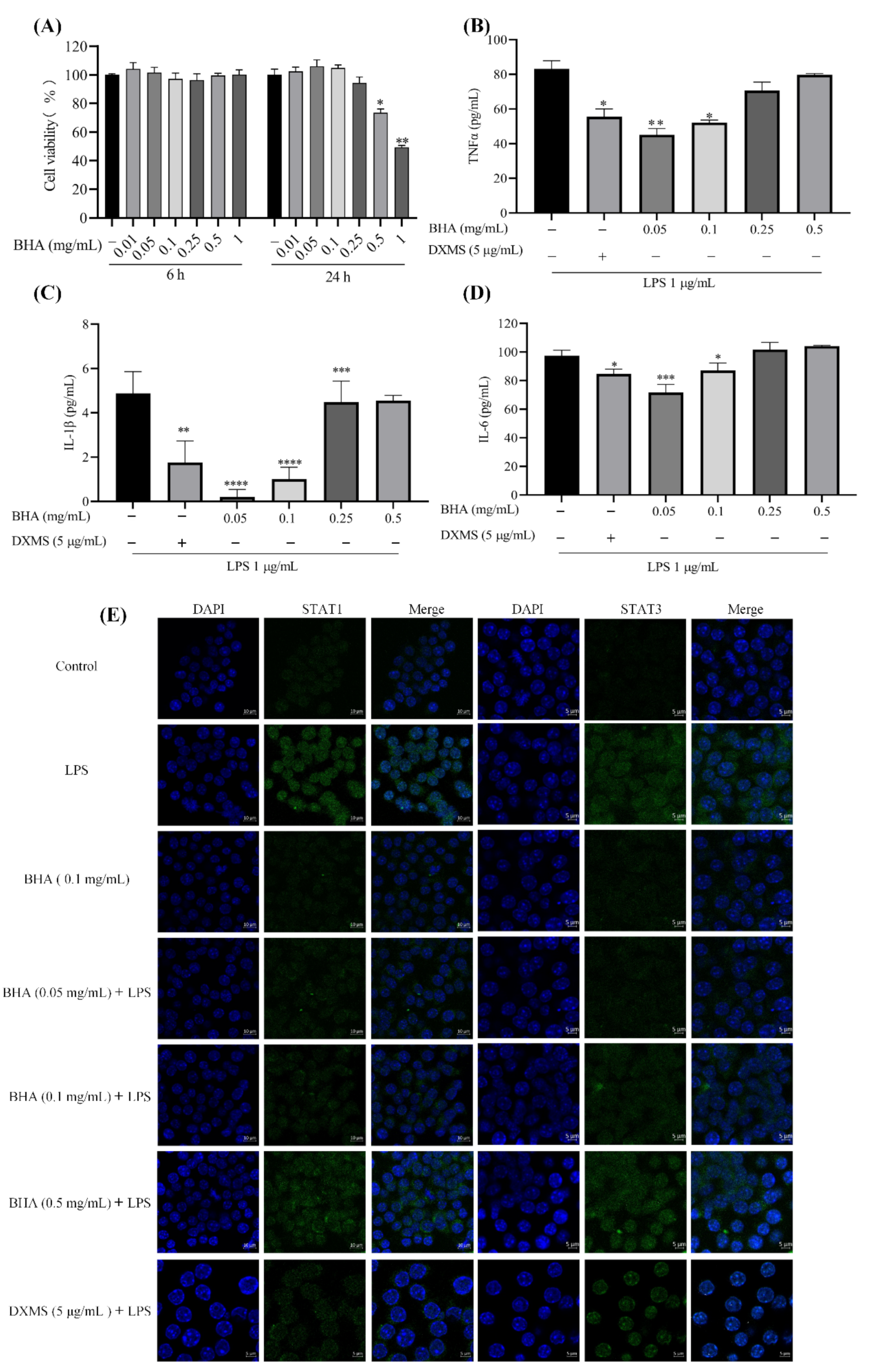
2.3. Effects of BHA on the Expression of Inflammation Factors in LPS-Induced RAW264.7 Marcrophage
2.4. BHA Inhibits LPS-Induced STAT1 and STAT3 Expression
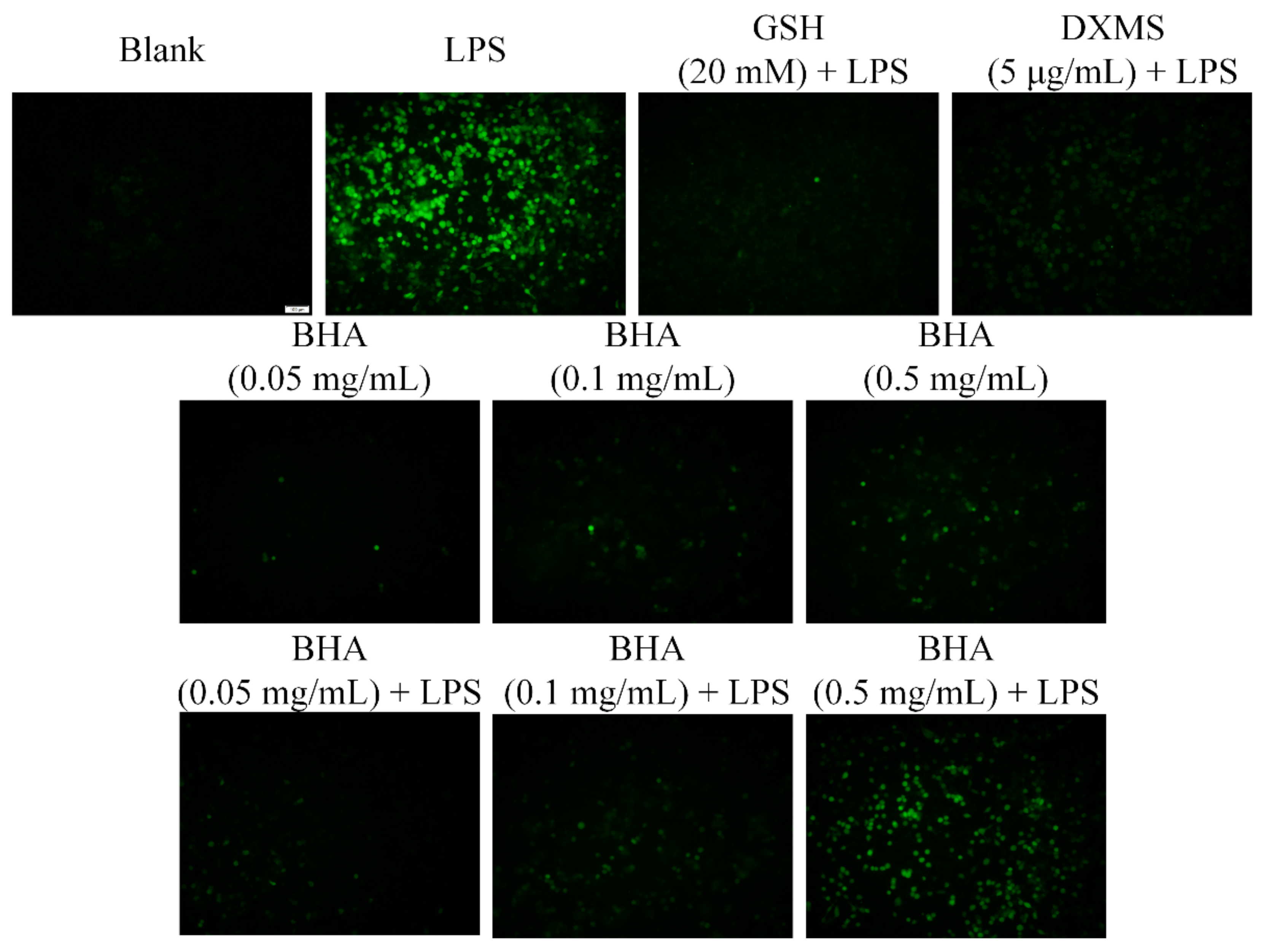
2.5. Effects of BHA on Oxidative Stress
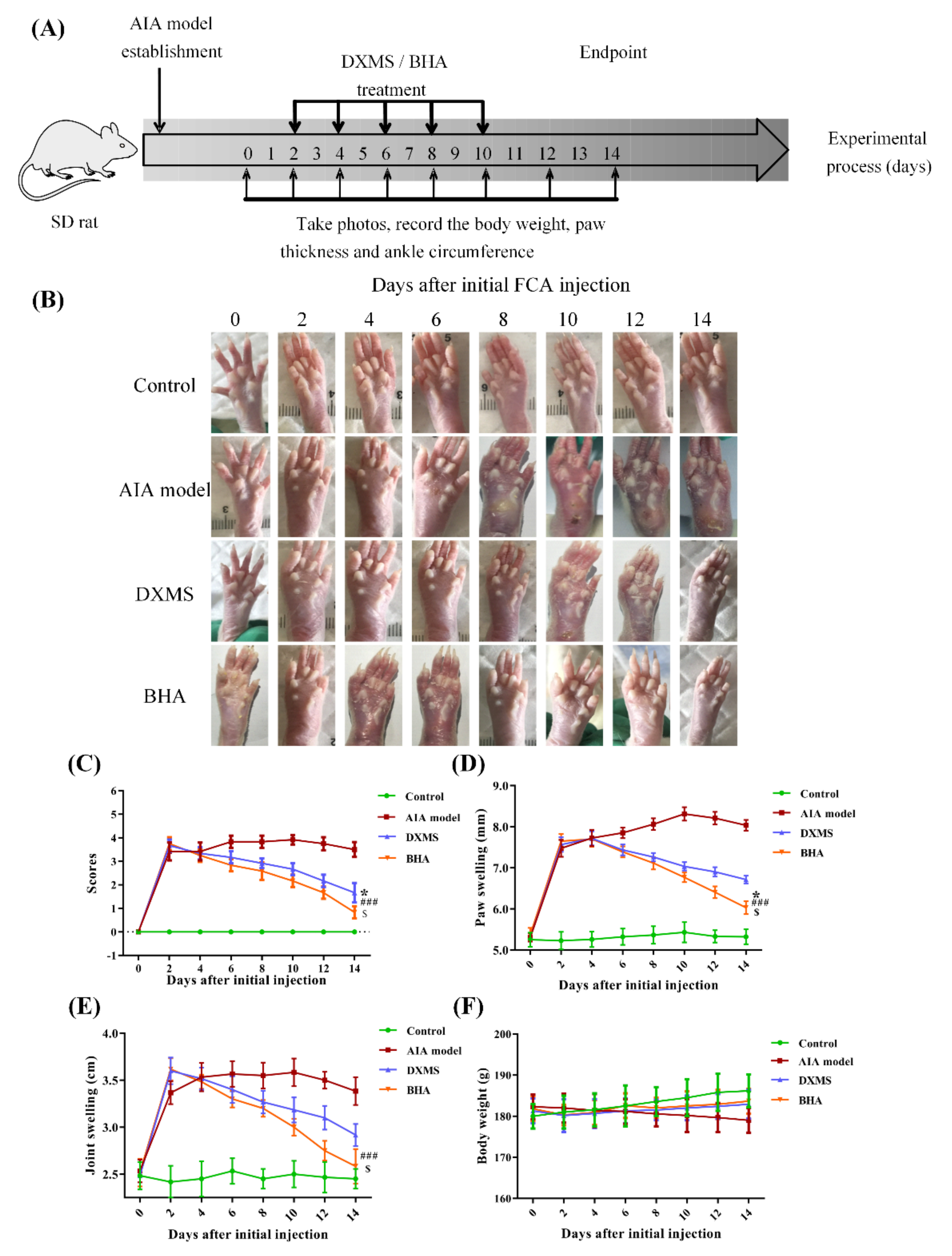
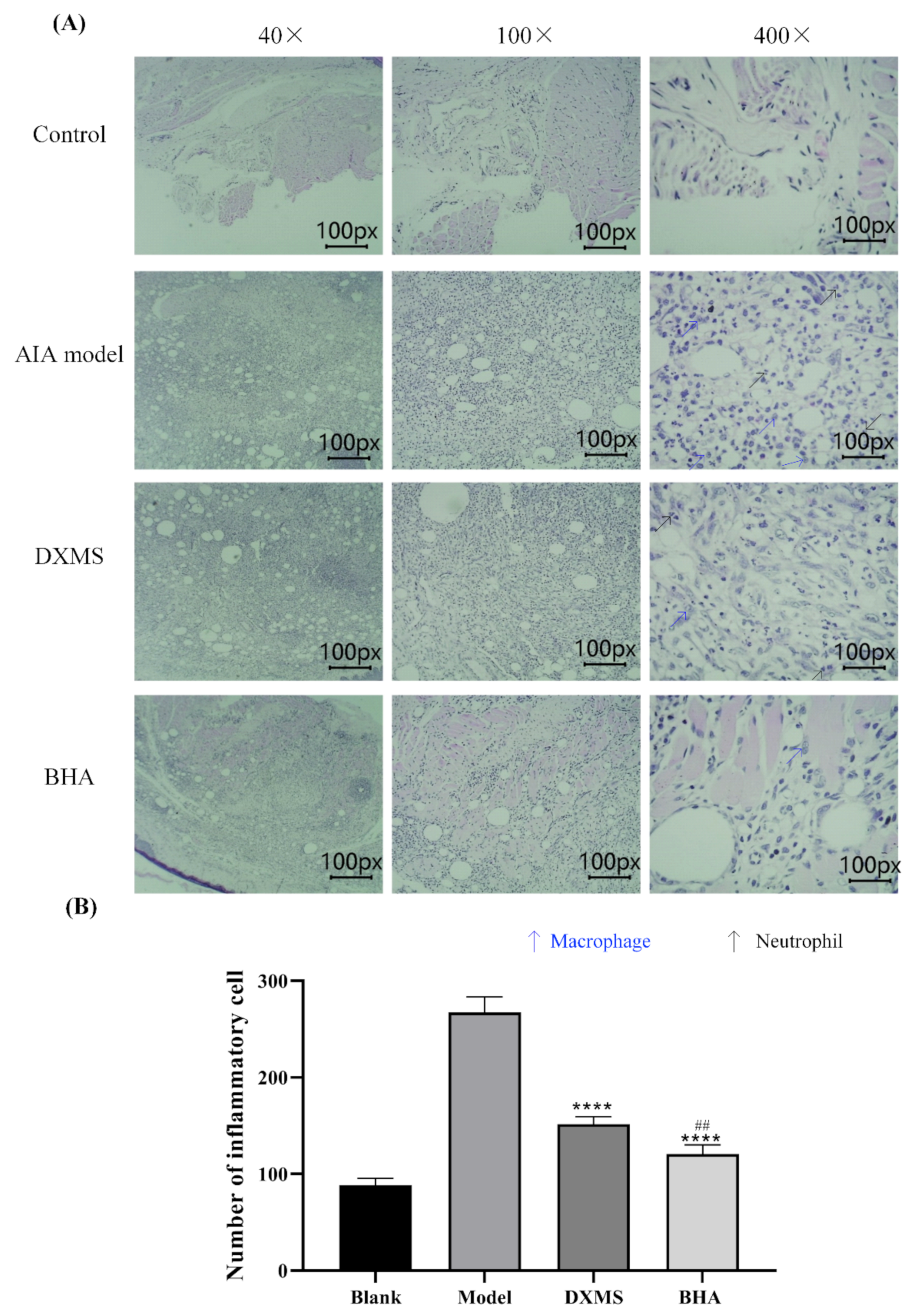
2.6. Therapeutic Effects of BHA on AIA Rats
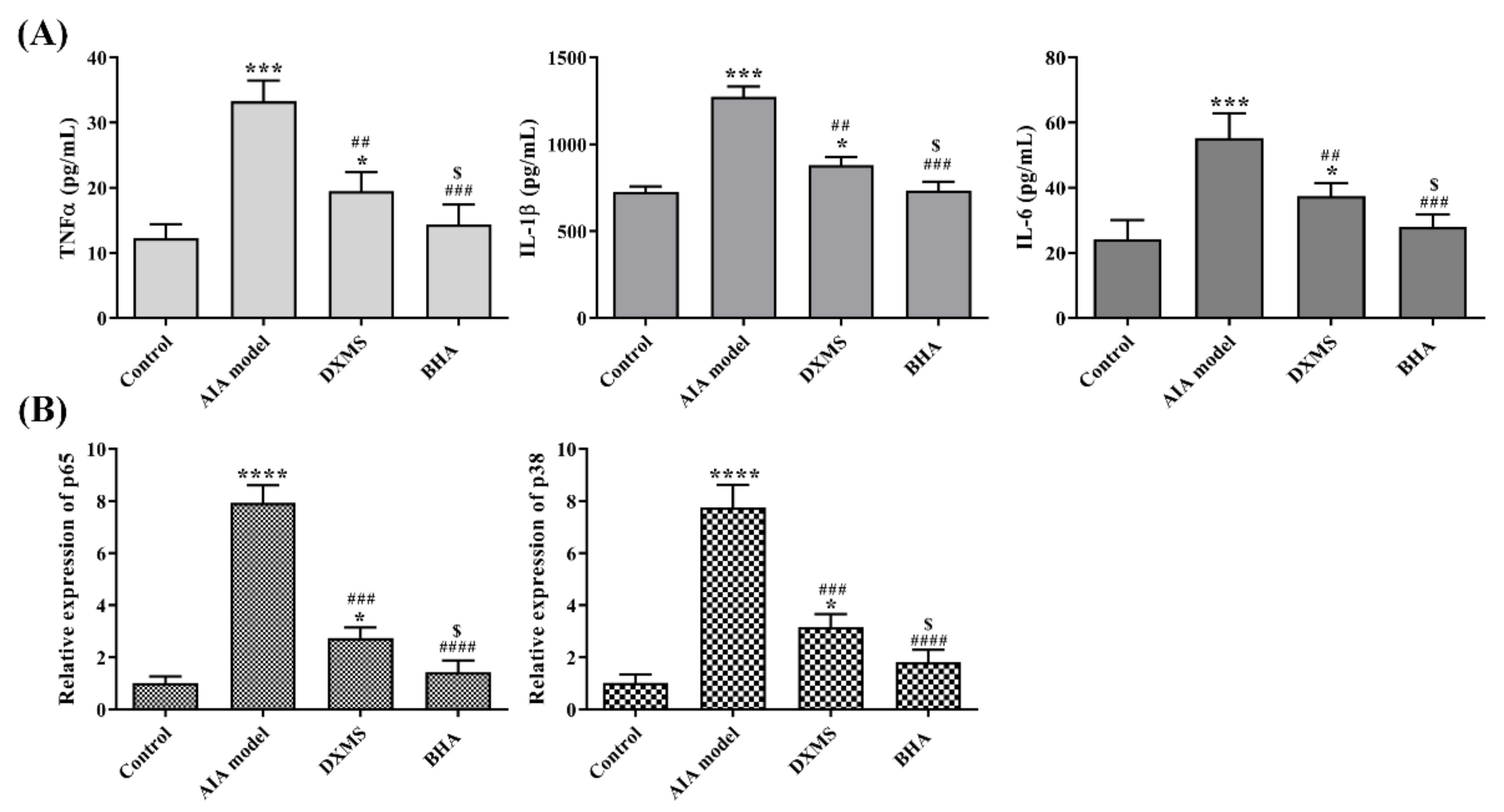
2.7. Effects of BHA on the Expression of Pro-Inflammatory Factors in AIA Rats
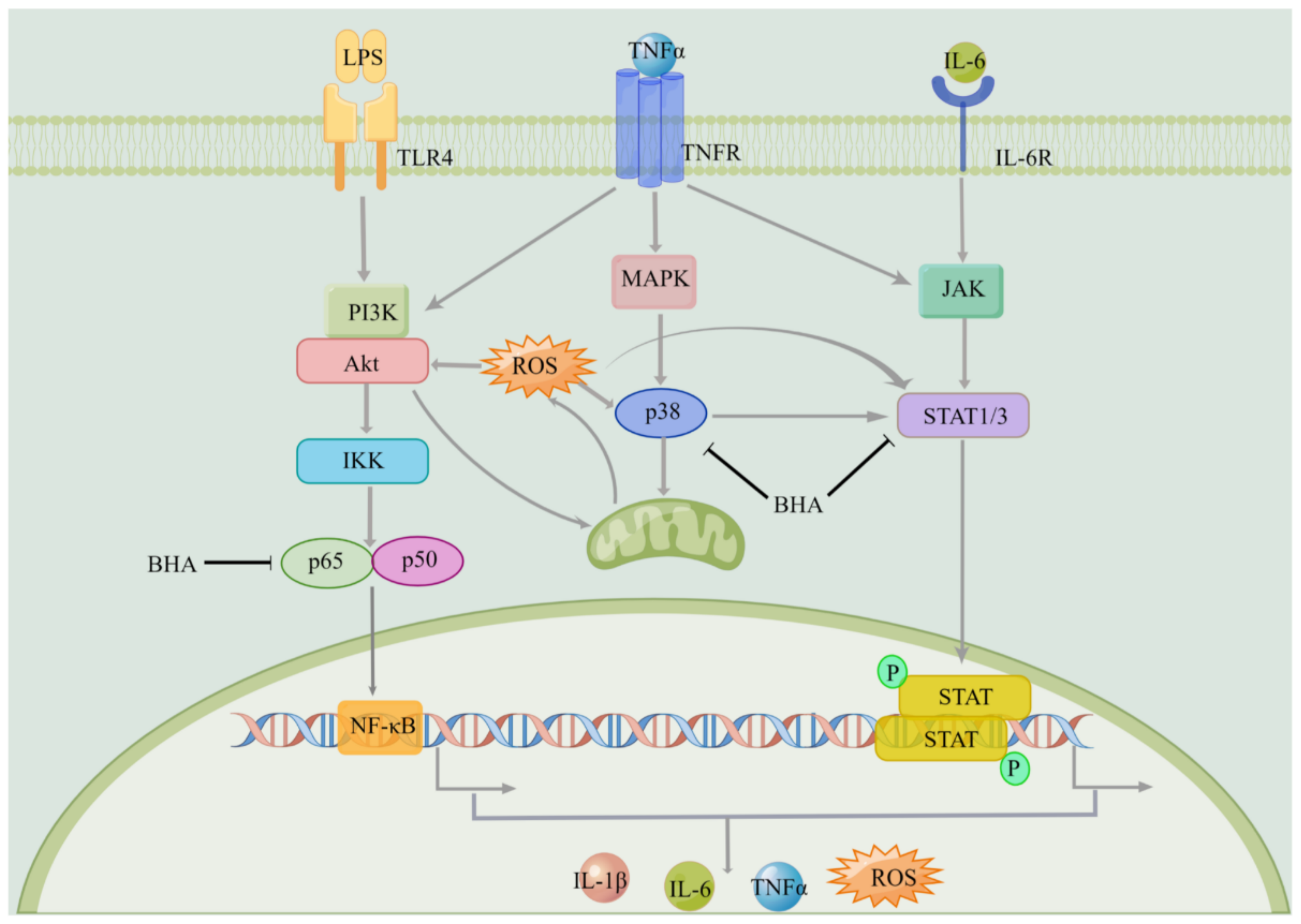
2.8. Effects of BHA on the Regulation of NF-κB and MAPK Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. BHA Synthesis and Characterization
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Immunofluorescence Assay
4.5. ROS Assay
4.6. Adjuvant-Induced Arthritis (AIA) Rat Model
4.7. Administration Method
4.8. Measurement of Paw Thickness and Ankle Circumferences
4.9. Grade Scores Used for Rat Arthritis Assessment
4.10. Enzyme-Linked Immunosorbent Assay
4.11. Histopathological Assessment
4.12. Real-Time Quantitative PCR
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Impellizzeri, D.; Esposito, E.; Di Paola, R.; Ahmad, A.; Campolo, M.; Peli, A.; Morittu, V.M.; Britti, D.; Cuzzocrea, S. Palmitoylethanolamide and luteolin ameliorate development of arthritis caused by injection of collagen type II in mice. Arthritis Res. Ther. 2013, 15, 4055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Chang, C.C.; Yang, Y.; Yuan, L.; Xu, L.; Ho, C.T.; Li, S. Resveratrol Alleviates Rheumatoid Arthritis via Reducing ROS and Inflammation, Inhibiting MAPK Signaling Pathways, and Suppressing Angiogenesis. J. Agric. Food Chem. 2018, 66, 12953–12960. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Lu, J.; Li, W.; Wu, A.; Zhang, X.; Tong, W.; Ho, K.K.; Qin, L.; Song, H.; Mak, K.K. Reciprocal inhibition of YAP/TAZ and NF-κB regulates osteoarthritic cartilage degradation. Nat. Commun. 2018, 9, 4564. [Google Scholar] [CrossRef] [Green Version]
- Mateen, S.; Zafar, A.; Moin, S.; Khan, A.Q.; Zubair, S. Understanding the role of cytokines in the pathogenesis of rheumatoid arthritis. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 455, 161–171. [Google Scholar] [CrossRef]
- Go, D.J.; Shin, K.; Baek, H.J.; Kang, S.W.; Kang, Y.M.; Jun, J.B.; Lee, Y.J.; Park, S.H.; Song, Y.W. Pivotal factors for successful withdrawal of nonsteroidal anti-inflammatory drugs in rheumatoid arthritis patients in remission or with low-disease activity. Clin. Rheumatol. 2018, 37, 307–314. [Google Scholar] [CrossRef]
- Kroot, E.J.; Huisman, A.M.; Van Zeben, J.; Wouters, J.M.; Van Paassen, H.C. Oral pulsed dexamethasone therapy in early rheumatoid arthritis: A pilot study. Ann. N. Y. Acad. Sci. 2006, 1069, 300–306. [Google Scholar] [CrossRef]
- Patrignani, P.; Patrono, C. Cyclooxygenase inhibitors: From pharmacology to clinical read-outs. Biochim. Biophys. Acta 2015, 1851, 422–432. [Google Scholar] [CrossRef]
- Sulaiman, W.; Toib, A.; Chandrashekhar, G.; Arshad, A. The Trends of DMARDS prescribed in Rheumatoid Arthritis Patients in Malaysia. Oman Med. J. 2009, 24, 260. [Google Scholar] [CrossRef]
- Gupta, R.C.; Lall, R.; Srivastava, A.; Sinha, A. Hyaluronic Acid: Molecular Mechanisms and Therapeutic Trajectory. Front. Vet. Sci. 2019, 6, 192. [Google Scholar] [CrossRef] [Green Version]
- Tamer, T.M. Hyaluronan and synovial joint: Function, distribution and healing. Interdiscip. Toxicol. 2013, 6, 111–125. [Google Scholar] [CrossRef]
- Temple-Wong, M.M.; Ren, S.; Quach, P.; Hansen, B.C.; Chen, A.C.; Hasegawa, A.; D’Lima, D.D.; Koziol, J.; Masuda, K.; Lotz, M.K.; et al. Hyaluronan concentration and size distribution in human knee synovial fluid: Variations with age and cartilage degeneration. Arthritis Res. Ther. 2016, 18, 18. [Google Scholar] [CrossRef] [Green Version]
- Gigis, I.; Fotiadis, E.; Nenopoulos, A.; Tsitas, K.; Hatzokos, I. Comparison of two different molecular weight intra-articular injections of hyaluronic acid for the treatment of knee osteoarthritis. Hippokratia 2016, 20, 26–31. [Google Scholar]
- Hermans, J.; Bierma-Zeinstra, S.M.A.; Bos, P.K.; Niesten, D.D.; Verhaar, J.A.N.; Reijman, M. The effectiveness of high molecular weight hyaluronic acid for knee osteoarthritis in patients in the working age: A randomised controlled trial. BMC Musculoskelet. Disord. 2019, 20, 196. [Google Scholar] [CrossRef] [Green Version]
- Roth, A.; Mollenhauer, J.; Wagner, A.; Fuhrmann, R.; Straub, A.; Venbrocks, R.A.; Petrow, P.; Bräuer, R.; Schubert, H.; Ozegowski, J.; et al. Intra-articular injections of high-molecular-weight hyaluronic acid have biphasic effects on joint inflammation and destruction in rat antigen-induced arthritis. Arthritis Res. Ther. 2005, 7, R677–R686. [Google Scholar] [CrossRef] [Green Version]
- Dahl, L.B.; Dahl, I.M.; Engström-Laurent, A.; Granath, K. Concentration and molecular weight of sodium hyaluronate in synovial fluid from patients with rheumatoid arthritis and other arthropathies. Ann. Rheum. Dis. 1985, 44, 817–822. [Google Scholar] [CrossRef] [Green Version]
- Cowman, M.K.; Shortt, C.; Arora, S.; Fu, Y.; Villavieja, J.; Rathore, J.; Huang, X.; Rakshit, T.; Jung, G.I.; Kirsch, T. Role of Hyaluronan in Inflammatory Effects on Human Articular Chondrocytes. Inflammation 2019, 42, 1808–1820. [Google Scholar] [CrossRef] [Green Version]
- Babasola, O.; Rees-Milton, K.J.; Bebe, S.; Wang, J.; Anastassiades, T.P. Chemically modified N-acylated hyaluronan fragments modulate proinflammatory cytokine production by stimulated human macrophages. J. Biol. Chem. 2014, 289, 24779–24791. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, D.; Wang, X.; Bai, R.; Wang, C.; Gao, Y.; Anastassiades, T. N-Butyrylated hyaluronic acid ameliorates gout and hyperuricemia in animal models. Pharm. Biol. 2019, 57, 717–728. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Yan, B.; Zhang, J.; Peng, L.; Ao, X.; Zheng, Z.; Jiang, T.; Zhang, Z. Casticin Attenuates Osteoarthritis-Related Cartilage Degeneration by Inhibiting the ROS-Mediated NF-κB Signaling Pathway in vitro and in vivo. Inflammation 2020, 43, 810–820. [Google Scholar] [CrossRef]
- Brennan, F.M.; McInnes, I.B. Evidence that cytokines play a role in rheumatoid arthritis. J. Clin. Investig. 2008, 118, 3537–3545. [Google Scholar] [CrossRef] [Green Version]
- Rizzardi, K.; Åkerlund, T. High Molecular Weight Typing with MALDI-TOF MS–A Novel Method for Rapid Typing of Clostridium difficile. PLoS ONE 2015, 10, e0122457. [Google Scholar] [CrossRef]
- Locock, K.E.; Meagher, L.; Haeussler, M. Oligomeric cationic polymethacrylates: A comparison of methods for determining molecular weight. Anal. Chem. 2014, 86, 2131–2137. [Google Scholar] [CrossRef]
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Murray, R.Z.; Stow, J.L. Cytokine Secretion in Macrophages: SNAREs, Rabs, and Membrane Trafficking. Front. Immunol. 2014, 5, 538. [Google Scholar] [CrossRef]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- Beconcini, D.; Felice, F.; Zambito, Y.; Fabiano, A.; Piras, A.M.; Macedo, M.H.; Sarmento, B.; Di Stefano, R. Anti-Inflammatory Effect of Cherry Extract Loaded in Polymeric Nanoparticles: Relevance of Particle Internalization in Endothelial Cells. Pharmaceutics 2019, 11, 500. [Google Scholar] [CrossRef] [Green Version]
- Kloep, M.; Güney, N.; Cok, F.; Simsek, O.F. Motives for risk-taking in adolescence: A cross-cultural study. J. Adolesc. 2009, 32, 135–151. [Google Scholar] [CrossRef]
- Hayakawa, M.; Miyashita, H.; Sakamoto, I.; Kitagawa, M.; Tanaka, H.; Yasuda, H.; Karin, M.; Kikugawa, K. Evidence that reactive oxygen species do not mediate NF-kappaB activation. EMBO J. 2003, 22, 3356–3366. [Google Scholar] [CrossRef] [Green Version]
- Castillo, F.; Monasterio, G.; Ibarra, J.P.; Guevara, J.; Cafferata, E.A.; Vicencio, E.; Cortez, C.; Carvajal, P.; Vernal, R. Levels of low-molecular-weight hyaluronan in periodontitis-treated patients and its immunostimulatory effects on CD4(+) T lymphocytes. Clin. Oral Investig. 2021, 25, 4987–5000. [Google Scholar] [CrossRef]
- Rayahin, J.E.; Buhrman, J.S.; Zhang, Y.; Koh, T.J.; Gemeinhart, R.A. High and low molecular weight hyaluronic acid differentially influence macrophage activation. ACS Biomater. Sci. Eng. 2015, 1, 481–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Geng, J.; Du, Y.; Li, S.; Wu, G.; Fu, Y.; Ren, H. Comparative study of the toxicity between three non-steroidal anti-inflammatory drugs and their UV/Na(2)S(2)O(8) degradation products on Cyprinus carpio. Sci. Rep. 2018, 8, 13512. [Google Scholar] [CrossRef] [PubMed]
- Wooley, P.H. The usefulness and the limitations of animal models in identifying targets for therapy in arthritis. Best Pract. Res. Clin. Rheumatol. 2004, 18, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Lepetsos, P.; Papavassiliou, K.A.; Papavassiliou, A.G. Redox and NF-κB signaling in osteoarthritis. Free Radic. Biol. Med. 2019, 132, 90–100. [Google Scholar] [CrossRef]
- Moltó, A.; Dougados, M. Novel DMARD monotherapy in rheumatoid arthritis. Lancet 2019, 393, 2277–2278. [Google Scholar] [CrossRef]
- Miossec, P. The role of interleukin 1 in the pathogenesis of rheumatoid arthritis. Clin. Exp. Rheumatol. 1987, 5, 305–308. [Google Scholar]
- Tas, S.W.; Remans, P.H.; Reedquist, K.A.; Tak, P.P. Signal transduction pathways and transcription factors as therapeutic targets in inflammatory disease: Towards innovative antirheumatic therapy. Curr. Pharm. Des. 2005, 11, 581–611. [Google Scholar] [CrossRef]
- Thalhamer, T.; McGrath, M.A.; Harnett, M.M. MAPKs and their relevance to arthritis and inflammation. Rheumatology 2008, 47, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Schett, G.; Tohidast-Akrad, M.; Smolen, J.S.; Schmid, B.J.; Steiner, C.W.; Bitzan, P.; Zenz, P.; Redlich, K.; Xu, Q.; Steiner, G. Activation, differential localization, and regulation of the stress-activated protein kinases, extracellular signal-regulated kinase, c-JUN N-terminal kinase, and p38 mitogen-activated protein kinase, in synovial tissue and cells in rheumatoid arthritis. Arthritis Rheum. 2000, 43, 2501–2512. [Google Scholar] [CrossRef]
- Chang, J.K.; Li, C.J.; Wu, S.C.; Yeh, C.H.; Chen, C.H.; Fu, Y.C.; Wang, G.J.; Ho, M.L. Effects of anti-inflammatory drugs on proliferation, cytotoxicity and osteogenesis in bone marrow mesenchymal stem cells. Biochem. Pharmacol. 2007, 74, 1371–1382. [Google Scholar] [CrossRef]
- Kalinec, G.; Thein, P.; Park, C.; Kalinec, F. HEI-OC1 cells as a model for investigating drug cytotoxicity. Hear. Res. 2016, 335, 105–117. [Google Scholar] [CrossRef]
- Gao, Y.; Sun, Y.; Yang, H.; Qiu, P.; Cong, Z.; Zou, Y.; Song, L.; Guo, J.; Anastassiades, T.P. A Low Molecular Weight Hyaluronic Acid Derivative Accelerates Excisional Wound Healing by Modulating Pro-Inflammation, Promoting Epithelialization and Neovascularization, and Remodeling Collagen. Int. J. Mol. Sci. 2019, 20, 3722. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, K.; Chu, X.; Li, T.; Shen, N.; Fan, C.; Niu, Z.; Zhang, X.; Hu, L. Intra-articular injection of Botulinum toxin A reduces neurogenic inflammation in CFA-induced arthritic rat model. Toxicon 2017, 126, 70–78. [Google Scholar] [CrossRef]
- Zhang, L.; Li, J.; Yu, S.C.; Jin, Y.; Lv, X.W.; Zou, Y.H.; Li, Y. Therapeutic effects and mechanisms of total flavonoids of Turpinia Arguta Seen on adjuvant arthritis in rats. J. Ethnopharmacol. 2008, 116, 167–172. [Google Scholar] [CrossRef]
- Luo, X.; Zuo, X.; Mo, X.; Zhou, Y.; Xiao, X. Treatment with recombinant Hsp72 suppresses collagen-induced arthritis in mice. Inflammation 2011, 34, 432–439. [Google Scholar] [CrossRef]
- Ma, Y.; Tang, T.; Sheng, L.; Wang, Z.; Tao, H.; Zhang, Q.; Zhang, Y.; Qi, Z. Aloin suppresses lipopolysaccharide-induced inflammation by inhibiting JAK1-STAT1/3 activation and ROS production in RAW264.7 cells. Int. J. Mol. Med. 2018, 42, 1925–1934. [Google Scholar] [CrossRef] [Green Version]
- Singh, C.; Roy-Chowdhuri, S. Quantitative Real-Time PCR: Recent Advances. Methods Mol. Biol. 2016, 1392, 161–176. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luan, X.; Cong, Z.; Anastassiades, T.P.; Gao, Y. N-Butyrylated Hyaluronic Acid Achieves Anti-Inflammatory Effects In Vitro and in Adjuvant-Induced Immune Activation in Rats. Molecules 2022, 27, 3267. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103267
Luan X, Cong Z, Anastassiades TP, Gao Y. N-Butyrylated Hyaluronic Acid Achieves Anti-Inflammatory Effects In Vitro and in Adjuvant-Induced Immune Activation in Rats. Molecules. 2022; 27(10):3267. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103267
Chicago/Turabian StyleLuan, Xue, Zhongcheng Cong, Tassos P. Anastassiades, and Yin Gao. 2022. "N-Butyrylated Hyaluronic Acid Achieves Anti-Inflammatory Effects In Vitro and in Adjuvant-Induced Immune Activation in Rats" Molecules 27, no. 10: 3267. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103267