
Chemical Compounds of Berry-Derived Polyphenols and Their Effects on Gut Microbiota, Inflammation, and Cancer

 , ,
, ,  , , , , , , , , and
, , , , , , , , and 
Abstract
:1. Introduction
2. General Information on Berry-Derived Polyphenols
2.1. Chemical Composition
2.1.1. Flavonols and Flavan-3-ols
2.1.2. Ellagitannins
2.1.3. Phenolic Acids
2.2. Biosynthesis and Regulation
3. Pharmacokinetic Aspects of Berry-Derived Polyphenols
3.1. Bioavailability and Metabolism of Berry-Derived Polyphenols
3.2. Pharmacokinetic of Phenolic Acids and Subgroups
3.3. Pharmacokinetic of Flavonoids and Their Subgroups
3.4. Pharmacokinetics of Other Polyphenolic Compounds
4. Effects of Berry-Derived Polyphenols on Gut Microbiota and Related Pathogenic Bacteria
5. Therapeutic Applications of Berry-Derived Polyphenols
5.1. Effects on Gut Inflammation
5.1.1. In Vitro Studies
5.1.2. In Vivo Studies
5.1.3. Clinical Trials
5.2. Effects on Different Types of Cancer
5.2.1. Colorectal Cancer
In Vitro Studies
In Vivo Studies
Clinical Trials
5.2.2. Breast Cancer
In Vitro Studies
In Vivo Studies
Clinical Trials
5.2.3. Esophageal Cancer
In Vitro Studies
In Vivo Studies
Clinical Trials
5.2.4. Prostate Cancer
5.3. Beneficial Effects on Gut-Microbiota-Induced Metabolic Disorders
5.3.1. In Vivo Analysis
5.3.2. In Vitro Studies
5.3.3. Clinical Trials
6. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| ABC | ATP-binding cassette |
| ACF | Aberrant crypt foci |
| ACI | August Copenhagen Irish |
| Akt | Protein kinase B |
| ANR | Anthocyanidin reductase |
| APC | Adenomatous polyposis coli |
| ATM | Ataxia telangiectasia mutated |
| BE | Blueberry extract |
| BRB | Black raspberry |
| CE | Cranberry extract |
| CETP | Cholesteryl ester transfer protein |
| COX | Cyclooxygenase |
| CRC | Colorectal cancer |
| CRPC | Castration-resistant prostate cancer |
| C3G | Cyanidin-3-glucoside |
| DAI | Disease activity index |
| DMBA | Dimethylbenze(a)anthrazene |
| DNA | Deoxyribonucleic acid |
| DNMT1 | DNA methyltransferase 1 |
| DNMT3B | DNA (cytosine-5-)-methyltransferase 3 beta |
| E2 | 17β-estradiol |
| EC | Esophagus cancer |
| ER | Endoplasmic reticulum |
| ERK: | Extracellular signal-regulated kinase |
| ETs | Ellagitannins |
| FAP | Familial adenomatous polyposis |
| GC/MS | Gas chromatography/mass spectrometry |
| GI50 | 50% cell growth inhibition |
| GIT | Gastrointestinal tract |
| GM-CSF | Granulocyte-macrophage colony-stimulating Factor |
| GPx | Glutathione peroxidase |
| GRO-α | Growth-related oncogene-alpha |
| HbA1c | Glycated hemoglobin |
| HCAs | Hydroxycinnamic acids |
| HCECs | Human colon epithelial cells |
| HDAC | Histone Deacetylase |
| HDL-C | High-density lipoprotein cholesterol |
| HER2 | Human epidermal growth factor receptor 2 |
| HPLC-DAD | High-performance liquid chromatography with diode array detection |
| HPLC-ESI-QTOF-MS | High-performance liquid chromatography coupled to electrospray ionization and quadrupole time-of-flight mass spectrometry |
| HPLC-FT-ICR MS | High-performance liquid chromatography coupled with Fourier-transform ion cyclotron resonance mass spectrometer |
| hsCRP | High-sensitivity C-reactive protein |
| Hsp70 | 70 kDa heat shock protein |
| HS-SPME | Headspace solid-phase microextraction |
| IBD | Inflammatory bowel disease |
| IC50 | Half-maximal inhibitory concentration |
| ICAM-1 | Intercellular adhesion molecule-1 |
| IFN | Interferon |
| IGF1 | Insulin-like growth factor-1 |
| IL | Interleukin |
| IL-1β | Interleukin-1β |
| iNOS | Inducible nitric oxide synthase |
| IP-10 | Interferon-γ-induced protein 10 |
| I-TAC | Interferon-inducible T-cell α chemoattractant |
| LAR | Leucoanthocyanidin reductase |
| LCBP | Lonicera Caerulea L., berry polyphenols |
| LC/ESI-MS | Liquid chromatography–electrospray ionization mass spectrometry |
| LC–MS–MS | Liquid chromatography with tandem mass spectrometry |
| LDL-C | Low-density lipoprotein cholesterol |
| LDL-R | Low-density lipoprotein receptor |
| LED | Light-emitting diode |
| LMWP | Low-molecular-weight phenols |
| LOH | Loss of heterozygosity |
| LPH | Lactase phlorizin hydrolase |
| LPS | Lipopolysaccharide |
| LXRα | Liver X receptor alpha |
| MALDI-TOF-MS | Matrix-assisted laser desorption/ionization-time-of-flight mass spectrometry |
| MAPK | Mitogen-activated protein kinase |
| MBD2 | Methyl-binding domain 2 |
| MCP-1 | Monocyte chemotactic protein-1 |
| MDA | Malondialdehyde |
| MDSC | Myeloid-derived suppressor cells |
| MeJA | Methyl jasmonate |
| MESB | Methanolic extract of strawberry |
| MMPs | Matrix metalloproteinases |
| MPO | Myeloperoxidase |
| mRNA | Messenger ribonucleic acid |
| MRPs | Multidrug-resistance proteins |
| MSI | Microsatellite instability |
| mTOR | Mammalian target of rapamycin |
| NAFLD | Nonalcoholic fatty liver disease |
| NF-κB | Nuclear factor kappa light-chain enhancer of activated B cells |
| NMBA | N-nitroso-methyl benzylamine |
| NO | Nitric oxide |
| PARP | Poly (ADP-ribose) polymerase |
| Pas | Proanthocyanidins |
| PCSK9 | Proprotein convertase subtilisin/kexin type 9 |
| PGC-1 α | Peroxisome proliferator-activated receptor γ coactivator 1-α |
| PGE2 | Prostaglandin E2 |
| pH | Potential of hydrogen |
| PI3-K | Phosphatidylinositol-3 kinase |
| PON-1 | Paraoxonase-1 |
| PPARγ | Peroxisome proliferator-activated receptor-γ |
| PRD | Partial rootzone drying |
| RNIs | Reactive nitrogen intermediates |
| ROS | Reactive oxygen species |
| SFCAs | Short-chain fatty acids |
| SGLT1 | Sodium-dependent glucose transporter |
| sICAM-1 | Soluble intercellular adhesion molecule-1 |
| SIRT1 | Sirtuin 1 |
| SOD | Superoxide dismutase |
| SREBP-1c | Sterol regulatory element-binding protein-1c |
| STAT3 | Signal transducer and activator of transcription 3 |
| TNF | Tumor necrosis factor |
| UPLC-ESI-MS | Ultraperformance liquid chromatography–electrospray ionization mass spectrometry |
| UV-B | Ultraviolet-B |
| VCAM-1 | Vascular cell adhesion molecule-1 |
| VEGF | Vascular endothelial growth factor |
References
- Lavefve, L.; Howard, L.R.; Carbonero, F. Berry Polyphenols Metabolism and Impact on Human Gut Microbiota and Health. Food Funct. 2020, 11, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Garcia, S.N.; Guevara-Gonzalez, R.G.; Miranda-Lopez, R.; Feregrino-Perez, A.A.; Torres-Pacheco, I.; Vazquez-Cruz, M.A. Functional Properties and Quality Characteristics of Bioactive Compounds in Berries: Biochemistry, Biotechnology, and Genomics. Food Res. Int. 2013, 54, 1195–1207. [Google Scholar] [CrossRef]
- Khan, I.; Najeebullah, S.; Ali, M.; Shinwari, Z.K. Phytopharmacological and Ethnomedicinal Uses of the Genus Berberis (Berberidaceae): A Review. Trop. J. Pharm. Res. 2016, 15, 2047–2057. [Google Scholar] [CrossRef] [Green Version]
- Nile, S.H.; Ko, E.Y.; Kim, D.H.; Keum, Y.-S.; Nile, S.H.; Ko, E.Y.; Kim, D.H.; Keum, Y.-S. Screening of Ferulic Acid Related Compounds as Inhibitors of Xanthine Oxidase and Cyclooxygenase-2 with Anti-Inflammatory Activity. Rev. Bras. Farmacogn. 2016, 26, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Gündeşli, M.A.; Korkmaz, N.; Okatan, V. Polyphenol Content and Antioxidant Capacity of Berries: A Review. Int. J. Agric. For. Life Sci. 2019, 3, 350–361. [Google Scholar]
- Nile, S.H.; Park, S.W. Edible Berries: Bioactive Components and Their Effect on Human Health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Vendrame, S.; Del Bo, C.; Ciappellano, S.; Riso, P.; Klimis-Zacas, D. Berry Fruit Consumption and Metabolic Syndrome. Antioxidants 2016, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, D.; Borges, G.; Crozier, A. Berry Flavonoids and Phenolics: Bioavailability and Evidence of Protective Effects. Br. J. Nutr. 2010, 104, S67–S90. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, K.; Olejnik, A. Current Evidence on the Health-Beneficial Effects of Berry Fruits in the Prevention and Treatment of Metabolic Syndrome. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 446–452. [Google Scholar] [CrossRef]
- Puupponen-Pimiä, R.; Nohynek, L.; Hartmann-Schmidlin, S.; Kähkönen, M.; Heinonen, M.; Määttä-Riihinen, K.; Oksman-Caldentey, K.-M. Berry Phenolics Selectively Inhibit the Growth of Intestinal Pathogens. J. Appl. Microbiol. 2005, 98, 991–1000. [Google Scholar] [CrossRef]
- Xiao, X.; Kim, J.; Sun, Q.; Kim, D.; Park, C.-S.; Lu, T.-S.; Park, Y. Preventive Effects of Cranberry Products on Experimental Colitis Induced by Dextran Sulphate Sodium in Mice. Food Chem. 2015, 167, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Jung, H.; Lee, H.; Yi, H.C.; Kwak, H.; Hwang, K.T. Chemopreventive Activity of Ellagitannins and Their Derivatives from Black Raspberry Seeds on HT-29 Colon Cancer Cells. Food Funct. 2015, 6, 1675–1683. [Google Scholar] [CrossRef] [PubMed]
- de Souza, M.O.; Silva, M.; Silva, M.E.; de Paula Oliveira, R.; Pedrosa, M.L. Diet Supplementation with Acai (Euterpe Oleracea Mart.) Pulp Improves Biomarkers of Oxidative Stress and the Serum Lipid Profile in Rats. Nutrition 2010, 26, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Paredes-López, O.; Cervantes-Ceja, M.L.; Vigna-Pérez, M.; Hernández-Pérez, T. Berries: Improving Human Health and Healthy Aging, and Promoting Quality Life—A Review. Plant Foods Hum. Nutr. 2010, 65, 299–308. [Google Scholar] [CrossRef]
- Donovan, J.L.; Manach, C.; Faulks, R.M.; Kroon, P.A. Absorption and Metabolism of Dietary Plant Secondary Metabolites. In Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet; Wiley-Blackwell: Hoboken, NJ, USA, 2006; pp. 303–351. [Google Scholar]
- Blumberg, J.B.; Camesano, T.A.; Cassidy, A.; Kris-Etherton, P.; Howell, A.; Manach, C.; Ostertag, L.M.; Sies, H.; Skulas-Ray, A.; Vita, J.A. Cranberries and Their Bioactive Constituents in Human Health. Adv. Nutr. 2013, 4, 618–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häkkinen, S.H.; Törrönen, A.R. Content of Flavonols and Selected Phenolic Acids in Strawberries and Vaccinium Species: Influence of Cultivar, Cultivation Site and Technique. Food Res. Int. 2000, 33, 517–524. [Google Scholar] [CrossRef]
- Peña-Sanhueza, D.; Inostroza-Blancheteau, C.; Ribera-Fonseca, A.; Reyes-Díaz, M. Anthocyanins in Berries and Their Potential Use in Human Health. In Superfood and Functional Food—The Development of Superfoods and Their Roles as Medicine; Shiomi, N., Waisundara, V., Eds.; IntechOpen: London, UK, 2017; pp. 155–172. [Google Scholar]
- Zanotti, I.; Dall’Asta, M.; Mena, P.; Mele, L.; Bruni, R.; Ray, S.; Del Rio, D. Atheroprotective Effects of (Poly) Phenols: A Focus on Cell Cholesterol Metabolism. Food Funct. 2015, 6, 13–31. [Google Scholar] [CrossRef]
- Tsuda, T. Dietary Anthocyanin-Rich Plants: Biochemical Basis and Recent Progress in Health Benefits Studies. Mol. Nutr. Food Res. 2012, 56, 159–170. [Google Scholar] [CrossRef]
- McGhie, T.K.; Walton, M.C. The Bioavailability and Absorption of Anthocyanins: Towards a Better Understanding. Mol. Nutr. Food Res. 2007, 51, 702–713. [Google Scholar] [CrossRef]
- Ghosh, D.; Konishi, T. Anthocyanins and Anthocyanin-Rich Extracts: Role in Diabetes and Eye Function. Asia Pac. J. Clin. Nutr. 2007, 16, 200–208. [Google Scholar]
- Wu, X.; Prior, R.L. Systematic Identification and Characterization of Anthocyanins by HPLC-ESI-MS/MS in Common Foods in the United States: Fruits and Berries. J. Agric. Food Chem. 2005, 53, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Genskowsky, E.; Puente, L.A.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Determination of Polyphenolic Profile, Antioxidant Activity and Antibacterial Properties of Maqui [Aristotelia Chilensi s (Molina) Stuntz] a Chilean Blackberry. J. Sci. Food Agric. 2016, 96, 4235–4242. [Google Scholar] [CrossRef] [PubMed]
- Marhuenda, J.; Alemán, M.D.; Gironés-Vilaplana, A.; Pérez, A.; Caravaca, G.; Figueroa, F.; Mulero, J.; Zafrilla, P. Phenolic Composition, Antioxidant Activity, and in Vitro Availability of Four Different Berries. J. Chem. 2016, 2016, 5194901. [Google Scholar] [CrossRef] [Green Version]
- Olivas-Aguirre, F.J.; Mendoza, S.; Alvarez-Parrilla, E.; Gonzalez-Aguilar, G.A.; Villegas-Ochoa, M.A.; Quintero-Vargas, J.T.; Wall-Medrano, A. First-Pass Metabolism of Polyphenols from Selected Berries: A High-Throughput Bioanalytical Approach. Antioxidants 2020, 9, 311. [Google Scholar] [CrossRef]
- Debnath-Canning, M.; Unruh, S.; Vyas, P.; Daneshtalab, N.; Igamberdiev, A.U.; Weber, J.T. Fruits and Leaves from Wild Blueberry Plants Contain Diverse Polyphenols and Decrease Neuroinflammatory Responses in Microglia. J. Funct. Foods 2020, 68, 103906. [Google Scholar] [CrossRef]
- Veberic, R.; Jakopic, J.; Stampar, F.; Schmitzer, V. European Elderberry (Sambucus nigra L.) Rich in Sugars, Organic Acids, Anthocyanins and Selected Polyphenols. Food Chem. 2009, 114, 511–515. [Google Scholar] [CrossRef]
- Prencipe, F.P.; Bruni, R.; Guerrini, A.; Rossi, D.; Benvenuti, S.; Pellati, F. Metabolite Profiling of Polyphenols in Vaccinium Berries and Determination of Their Chemopreventive Properties. J. Pharm. Biomed. Anal. 2014, 89, 257–267. [Google Scholar] [CrossRef]
- Cesa, S.; Carradori, S.; Bellagamba, G.; Locatelli, M.; Casadei, M.A.; Masci, A.; Paolicelli, P. Evaluation of processing effects on anthocyanin content and colour modifications of blueberry (Vaccinium spp.) Extracts: Comparison between HPLC-DAD and CIELAB Analyses. Food Chem. 2017, 232, 114–123. [Google Scholar] [CrossRef]
- Levaj, B.; Dragović-Uzelac, V.; Delonga, K.; Kovačević Ganić, K.; Banović, M.; Bursać Kovačević, D. Polyphenols and Volatiles in Fruits of Two Sour Cherry Cultivars, Some Berry Fruits and Their Jams. Food Technol. Biotechnol. 2010, 48, 538–547. [Google Scholar]
- Kajdžanoska, M.; Gjamovski, V.; Stefova, M. HPLC-DAD-ESI-MSn Identification of Phenolic Compounds in Cultivated Strawberries from Macedonia. Maced. J. Chem. Chem. Eng. 2010, 29, 181–194. [Google Scholar] [CrossRef] [Green Version]
- Mikulic-Petkovsek, M.; Slatnar, A.; Stampar, F.; Veberic, R. HPLC–MSn Identification and Quantification of Flavonol Glycosides in 28 Wild and Cultivated Berry Species. Food Chem. 2012, 135, 2138–2146. [Google Scholar] [CrossRef] [PubMed]
- Määttä, K.R.; Kamal-Eldin, A.; Törrönen, A.R. High-Performance Liquid Chromatography (HPLC) Analysis of Phenolic Compounds in Berries with Diode Array and Electrospray Ionization Mass Spectrometric (MS) Detection: Ribes Species. J. Agric. Food Chem. 2003, 51, 6736–6744. [Google Scholar] [CrossRef] [PubMed]
- Mieres-Castro, D.; Schmeda-Hirschmann, G.; Theoduloz, C.; Gómez-Alonso, S.; Pérez-Navarro, J.; Márquez, K.; Jiménez-Aspee, F. Antioxidant Activity and the Isolation of Polyphenols and New Iridoids from Chilean Gaultheria Phillyreifolia and G. Poeppigii Berries. Food Chem. 2019, 291, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Kaume, L.; Howard, L.R.; Devareddy, L. The Blackberry Fruit: A Review on Its Composition and Chemistry, Metabolism and Bioavailability, and Health Benefits. J. Agric. Food Chem. 2012, 60, 5716–5727. [Google Scholar] [CrossRef]
- Beattie, J.; Crozier, A.; Duthie, G.G. Potential Health Benefits of Berries. Curr. Nutr. Food Sci. 2005, 1, 71–86. [Google Scholar] [CrossRef]
- Zorzi, M.; Gai, F.; Medana, C.; Aigotti, R.; Morello, S.; Peiretti, P.G. Bioactive Compounds and Antioxidant Capacity of Small Berries. Foods 2020, 9, 623. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly) Phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects against Chronic Diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landete, J.M. Ellagitannins, Ellagic Acid and Their Derived Metabolites: A Review about Source, Metabolism, Functions and Health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Määttä-Riihinen, K.R.; Kamal-Eldin, A.; Törrönen, A.R. Identification and quantification of phenolic compounds in berries of fragaria and rubus species (Family rosaceae). J. Agric. Food Chem. 2004, 52, 6178–6187. [Google Scholar] [CrossRef]
- Rao, A.V.; Snyder, D.M. Raspberries and Human Health: A Review. J. Agric. Food Chem. 2010, 58, 3871–3883. [Google Scholar] [CrossRef]
- Khallouki, F.; Haubner, R.; Hull, W.E.; Erben, G.; Spiegelhalder, B.; Bartsch, H.; Owen, R.W. Isolation, purification and identification of ellagic acid derivatives, catechins, and procyanidins from the root bark of Anisophyllea dichostyla R. Br. Food Chem. Toxicol. 2007, 45, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Hager, T.J.; Howard, L.R.; Liyanage, R.; Lay, J.O.; Prior, R.L. Ellagitannin Composition of Blackberry as Determined by HPLC-ESI-MS and MALDI-TOF-MS. J. Agric. Food Chem. 2008, 56, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Lazarus, S.A.; Cao, G.; Muccitelli, H.; Hammerstone, J.F. Identification of procyanidins and anthocyanins in blueberries and cranberries (Vaccinium spp.) using high-performance liquid chromatography/mass spectrometry. J. Agric. Food Chem. 2001, 49, 1270–1276. [Google Scholar] [CrossRef] [PubMed]
- Hajazimi, E.; Landberg, R.; Zamaratskaia, G. Simultaneous Determination of Flavonols and Phenolic Acids by HPLC-CoulArray in Berries Common in the Nordic Diet. LWT 2016, 74, 128–134. [Google Scholar] [CrossRef]
- Stanciu, G.; Lupsor, S.; Popescu, A.; Oancea, I.A. Polyphenols Isolation and Determination in Grape Seeds by HPLC/DAD. J. Sci. Arts 2017, 17, 107. [Google Scholar]
- Liang, Z.; Sang, M.; Fan, P.; Wu, B.; Wang, L.; Duan, W.; Li, S. Changes of Polyphenols, Sugars, and Organic Acid in 5 Vitis Genotypes during Berry Ripening. J. Food Sci. 2011, 76, C1231–C1238. [Google Scholar] [CrossRef]
- Radovanović, B.C.; Anđelković, S.M.; Radovanović, A.B.; Anđelković, M.Z. Antioxidant and Antimicrobial Activity of Polyphenol Extracts from Wild Berry Fruits Grown in Southeast Serbia. Trop. J. Pharm. Res. 2013, 12, 813–819. [Google Scholar] [CrossRef]
- Zadernowski, R.; Naczk, M.; Nesterowicz, J. Phenolic Acid Profiles in Some Small Berries. J. Agric. Food Chem. 2005, 53, 2118–2124. [Google Scholar] [CrossRef]
- Matus, J.T.; Loyola, R.; Vega, A.; Peña-Neira, A.; Bordeu, E.; Arce-Johnson, P.; Alcalde, J.A. Post-Veraison Sunlight Exposure Induces MYB-Mediated Transcriptional Regulation of Anthocyanin and Flavonol Synthesis in Berry Skins of Vitis Vinifera. J. Exp. Bot. 2009, 60, 853–867. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Zhao, R.; Zeng, J.; Li, G.; Li, X. Characterization of Destrins with Different Dextrose Equivalents. Molecules 2010, 15, 5162–5173. [Google Scholar] [CrossRef] [Green Version]
- Berli, F.J.; Fanzone, M.; Piccoli, P.; Bottini, R. Solar UV-B and ABA are involved in phenol metabolism of Vitis vinifera L. increasing biosynthesis of berry skin polyphenols. J. Agric. Food Chem. 2011, 59, 4874–4884. [Google Scholar] [CrossRef] [PubMed]
- Koyama, K.; Ikeda, H.; Poudel, P.R.; Goto-Yamamoto, N. Light Quality Affects Flavonoid Biosynthesis in Young Berries of Cabernet Sauvignon Grape. Phytochemistry 2012, 78, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; Tomiyama, H.; Rodyoung, A.; Okawa, K.; Ohara, H.; Sugaya, S.; Terahara, N.; Hirai, N. Abscisic Acid Metabolism and Anthocyanin Synthesis in Grape Skin Are Affected by Light Emitting Diode (LED) Irradiation at Night. J. Plant Physiol. 2014, 171, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Dai, Z.; Wu, B.-H.; Wu, J.; Merlin, I.; Hilbert, G.; Renaud, C.; Gomès, E.; Edwards, E.; Li, S.-H. Anthocyanin Biosynthesis Is Differentially Regulated by Light in the Skin and Flesh of White-Fleshed and Teinturier Grape Berries. Planta 2016, 243, 23–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Tian, D.-D.; Zheng, B.; Wang, D.; Tan, Q.-R.; Wang, C.-Y.; Zhang, Z.-J. Peony-Glycyrrhiza Decoction, an Herbal Preparation, Inhibits Clozapine Metabolism via Cytochrome P450s, but Not Flavin-Containing Monooxygenase in in Vitro Models. Drug Metab. Dispos. 2015, 43, 1147–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, R.; Berli, F.J.; Fontana, A.; Piccoli, P.; Bottini, R. Malbec grape (Vitis vinifera L.) responses to the environment: Berry phenolics as influenced by solar uv-b, water deficit and sprayed abscisic acid. Plant Physiol. Biochem. 2016, 109, 84–90. [Google Scholar] [CrossRef]
- Marfil, C.; Ibañez, V.; Alonso, R.; Varela, A.; Bottini, R.; Masuelli, R.; Fontana, A.; Berli, F. Changes in Grapevine DNA Methylation and Polyphenols Content Induced by Solar Ultraviolet-B Radiation, Water Deficit and Abscisic Acid Spray Treatments. Plant Physiol. Biochem. 2019, 135, 287–294. [Google Scholar] [CrossRef]
- Degu, A.; Ayenew, B.; Cramer, G.R.; Fait, A. Polyphenolic Responses of Grapevine Berries to Light, Temperature, Oxidative Stress, Abscisic Acid and Jasmonic Acid Show Specific Developmental-Dependent Degrees of Metabolic Resilience to Perturbation. Food Chem. 2016, 212, 828–836. [Google Scholar] [CrossRef]
- Wang, J.; Wang, S.; Liu, G.; Edwards, E.J.; Duan, W.; Li, S.; Wang, L. The synthesis and accumulation of resveratrol are associated with veraison and abscisic acid concentration in beihong (Vitis vinifera × Vitis amurensis) berry skin. Front. Plant Sci. 2016, 7, 1605. [Google Scholar] [CrossRef] [Green Version]
- Antolín, M.C.; Santesteban, H.; Santa María, E.; Aguirreolea, J.; Sánchez-Díaz, M. Involvement of abscisic acid and polyamines in berry ripening of Vitis vinifera (L.) subjected to water deficit irrigation. Aust. J. Grape Wine Res. 2008, 14, 123–133. [Google Scholar] [CrossRef]
- Ojeda, H.; Andary, C.; Kraeva, E.; Carbonneau, A.; Deloire, A. Influence of Pre-and Postveraison Water Deficit on Synthesis and Concentration of Skin Phenolic Compounds during Berry Growth of Vitis Vinifera Cv. Shiraz. Am. J. Enol. Vitic. 2002, 53, 261–267. [Google Scholar]
- Sun, R.-Z.; Cheng, G.; Li, Q.; He, Y.-N.; Wang, Y.; Lan, Y.-B.; Li, S.-Y.; Zhu, Y.-R.; Song, W.-F.; Zhang, X. Light-induced variation in phenolic compounds in cabernet sauvignon grapes (Vitis vinifera L.) involves extensive transcriptome reprogramming of biosynthetic enzymes, transcription factors, and phytohormonal regulators. Front. Plant Sci. 2017, 8, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiko, F.; Noriko, S.; Nami, G.-Y.; Akihiro, M.; Kuniaki, K.; Katsumi, H. Effect of Shading on Proanthocyanidin Biosynthesis in the Grape Berry. J. Jpn. Soc. Hortic. Sci. 2007, 76, 112–119. [Google Scholar]
- Koyama, K.; Goto-Yamamoto, N. Bunch Shading during Different Developmental Stages Affects the Phenolic Biosynthesis in Berry Skins of ‘Cabernet Sauvignon’Grapes. J. Am. Soc. Hortic. Sci. 2008, 133, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Quiroga, A.M.; Berli, F.J.; Moreno, D.; Cavagnaro, J.B.; Bottini, R. Abscisic acid sprays significantly increase yield per plant in vineyard-grown wine grape (Vitis vinifera L.) cv. cabernet sauvignon through increased berry set with no negative effects on anthocyanin content and total polyphenol index of both juice and wine. J. Plant Growth Regul. 2009, 28, 28–35. [Google Scholar]
- Shahab, M.; Roberto, S.R.; Ahmed, S.; Colombo, R.C.; Silvestre, J.P.; Koyama, R.; de Souza, R.T. Relationship between Anthocyanins and Skin Color of Table Grapes Treated with Abscisic Acid at Different Stages of Berry Ripening. Sci. Hortic. 2020, 259, 108859. [Google Scholar] [CrossRef]
- Sandhu, A.K.; Gray, D.J.; Lu, J.; Gu, L. Effects of Exogenous Abscisic acid on antioxidant capacities, anthocyanins, and flavonol contents of muscadine grape (Vitis rotundifolia) skins. Food Chem. 2011, 126, 982–988. [Google Scholar] [CrossRef]
- Koyama, R.; de Assis, A.M.; Yamamoto, L.Y.; Borges, W.F.; de Sá Borges, R.; Prudêncio, S.H.; Roberto, S.R. Exogenous abscisic acid increases the anthocyanin concentration of berry and juice from ‘isabel’grapes (Vitis labrusca L.). HortScience 2014, 49, 460–464. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, L.Y.; Koyama, R.; de Assis, A.M.; Borges, W.F.S.; de Oliveira, I.R.; Roberto, S.R. Color of Berry and Juice of’Isabel’grape Treated with Abscisic Acid in Different Ripening Stages. Pesqui. Agropecuária Bras. 2015, 50, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, L.Y.; de Assis, A.M.; Roberto, S.R.; Bovolenta, Y.R.; Nixdorf, S.L.; García-Romero, E.; Gómez-Alonso, S.; Hermosín-Gutiérrez, I. Application of abscisic acid (S-ABA) to cv. isabel grapes (Vitis vinifera × Vitis labrusca) for color improvement: Effects on color, phenolic composition and antioxidant capacity of their grape juice. Food Res. Int. 2015, 77, 572–583. [Google Scholar] [CrossRef]
- Ju, Y.-L.; Liu, M.; Zhao, H.; Meng, J.-F.; Fang, Y.-L. Effect of exogenous abscisic acid and methyl jasmonate on anthocyanin composition, fatty acids, and volatile compounds of cabernet sauvignon (Vitis vinifera L.) grape berries. Molecules 2016, 21, 1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murcia, G.; Fontana, A.; Pontin, M.; Baraldi, R.; Bertazza, G.; Piccoli, P.N. ABA and GA3 Regulate the Synthesis of Primary and Secondary Metabolites Related to Alleviation from Biotic and Abiotic Stresses in Grapevine. Phytochemistry 2017, 135, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, H.; Mancera-Noyola, M.; Zermeño-Gonzalez, A.; Jasso-Cantú, D.; Villarreal-Quintanilla, J.A. Effect of abscisic acid on yield, anthocyanins, polyphenols and procyanidins in grape cv. cabernet franc (Vitis vinífera L.). Int. J. Plant Soil Sci. 2018, 24, 5–7. [Google Scholar] [CrossRef]
- Koyama, R.; Roberto, S.R.; de Souza, R.T.; Borges, W.F.; Anderson, M.; Waterhouse, A.L.; Cantu, D.; Fidelibus, M.W.; Blanco-Ulate, B. Exogenous abscisic acid promotes anthocyanin biosynthesis and increased expression of flavonoid synthesis genes in Vitis vinifera × Vitis labrusca table grapes in a subtropical region. Front. Plant Sci. 2018, 9, 323. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Wang, L.; Belwal, T.; Zhang, X.; Lu, H.; Chen, C.; Li, L. Exogenous melatonin and abscisic acid expedite the flavonoids biosynthesis in grape berry of vitis vinifera cv. kyoho. Molecules 2020, 25, 12. [Google Scholar] [CrossRef] [Green Version]
- Castellarin, S.D.; Di Gaspero, G. Transcriptional Control of Anthocyanin Biosynthetic Genes in Extreme Phenotypes for Berry Pigmentation of Naturally Occurring Grapevines. BMC Plant Biol. 2007, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Deluc, L.; Bogs, J.; Walker, A.R.; Ferrier, T.; Decendit, A.; Merillon, J.-M.; Robinson, S.P.; Barrieu, F. The Transcription Factor VvMYB5b Contributes to the Regulation of Anthocyanin and Proanthocyanidin Biosynthesis in Developing Grape Berries. Plant Physiol. 2008, 147, 2041–2053. [Google Scholar] [CrossRef] [Green Version]
- Villano, C.; Rinaldi, A.; Lanzillo, C.; Moio, L.; Tamburrino, A.J.; Carputo, D.; Frusciante, L.; Aversano, R. Polyphenol Content and Differential Expression of Flavonoid Biosynthetic Pathway Genes in Berries of Aglianico. In Proceedings of the X International Symposium on Grapevine Physiology and Biotechnology, Verona, Italy, 18 June 2016; pp. 141–148. [Google Scholar]
- Davik, J.; Aaby, K.; Buti, M.; Alsheikh, M.; Šurbanovski, N.; Martens, S.; Røen, D.; Sargent, D.J. Major-Effect Candidate Genes Identified in Cultivated Strawberry (Fragaria × ananassa Duch.) for Ellagic Acid Deoxyhexoside and Pelargonidin-3-O-Malonylglucoside Biosynthesis, Key Polyphenolic Compounds. Hortic. Res. 2020, 7, 125. [Google Scholar] [CrossRef]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin Synthesis and Expression of Genes Encoding Leucoanthocyanidin Reductase and Anthocyanidin Reductase in Developing Grape Berries and Grapevine Leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, C.; Zamperin, G.; Ferrarini, A.; Minio, A.; Dal Molin, A.; Venturini, L.; Buson, G.; Tononi, P.; Avanzato, C.; Zago, E. The High Polyphenol Content of Grapevine Cultivar Tannat Berries Is Conferred Primarily by Genes That Are Not Shared with the Reference Genome. Plant Cell 2013, 25, 4777–4788. [Google Scholar] [CrossRef]
- Delgado, R.; Martín, P.; Del Álamo, M.; González, M.-R. Changes in the Phenolic Composition of Grape Berries during Ripening in Relation to Vineyard Nitrogen and Potassium Fertilisation Rates. J. Sci. Food Agric. 2004, 84, 623–630. [Google Scholar] [CrossRef]
- Martins, V.; Billet, K.; Garcia, A.; Lanoue, A.; Gerós, H. Exogenous Calcium Deflects Grape Berry Metabolism towards the Production of More Stilbenoids and Less Anthocyanins. Food Chem. 2020, 313, 126123. [Google Scholar] [CrossRef] [PubMed]
- Griesser, M.; Martinez, S.C.; Eitle, M.W.; Warth, B.; Andre, C.M.; Schuhmacher, R.; Forneck, A. The Ripening Disorder Berry Shrivel Affects Anthocyanin Biosynthesis and Sugar Metabolism in Zweigelt Grape Berries. Planta 2018, 247, 471–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudel, P.R.; Koyama, K.; Goto-Yamamoto, N. Evaluating the Influence of temperature on proanthocyanidin biosynthesis in developing grape berries (Vitis vinifera L.). Mol. Biol. Rep. 2020, 47, 3501–3510. [Google Scholar] [CrossRef] [PubMed]
- Anastasiadi, M.; Pratsinis, H.; Kletsas, D.; Skaltsounis, A.-L.; Haroutounian, S.A. Bioactive Non-Coloured Polyphenols Content of Grapes, Wines and Vinification by-Products: Evaluation of the Antioxidant Activities of Their Extracts. Food Res. Int. 2010, 43, 805–813. [Google Scholar] [CrossRef]
- Bakowska-Barczak, A.M.; Kolodziejczyk, P. Evaluation of saskatoon berry (Amelanchier alnifolia Nutt.) cultivars for their polyphenol content, antioxidant properties, and storage stability. J. Agric. Food Chem. 2008, 56, 9933–9940. [Google Scholar] [CrossRef]
- Barboni, T.; Venturini, N.; Paolini, J.; Desjobert, J.-M.; Chiaramonti, N.; Costa, J. Characterisation of Volatiles and Polyphenols for Quality Assessment of Alcoholic Beverages Prepared from Corsican Myrtus Communis Berries. Food Chem. 2010, 122, 1304–1312. [Google Scholar] [CrossRef]
- Barboni, T.; Cannac, M.; Massi, L.; Perez-Ramirez, Y.; Chiaramonti, N. Variability of polyphenol compounds in Myrtus communis L. (Myrtaceae) berries from corsica. Molecules 2010, 15, 7849–7860. [Google Scholar] [CrossRef] [Green Version]
- Bittová, M.; Krejzová, E.; Roblová, V.; Kubáň, P.; Kubáň, V. Monitoring of HPLC profiles of selected polyphenolic compounds in sea buckthorn (Hippophaë rhamnoides L.) plant parts during annual growth cycle and estimation of their antioxidant potential. Cent. Eur. J. Chem. 2014, 12, 1152–1161. [Google Scholar] [CrossRef]
- Čanadanović-Brunet, J.; Tumbas Šaponjac, V.; Stajčić, S.; Ćetković, G.; Čanadanović, V.; Ćebović, T.; Vulić, J. Polyphenolic Composition, Antiradical and Hepatoprotective Activities of Bilberry and Blackberry Pomace Extracts. J. Berry Res. 2019, 9, 349–362. [Google Scholar] [CrossRef]
- Cavaliere, C.; Foglia, P.; Gubbiotti, R.; Sacchetti, P.; Samperi, R.; Laganà, A. Rapid-Resolution Liquid Chromatography/Mass Spectrometry for Determination and Quantitation of Polyphenols in Grape Berries. Rapid Commun. Mass Spectrom. Int. J. Devoted Rapid Dissem. Minute Res. Mass Spectrom. 2008, 22, 3089–3099. [Google Scholar] [CrossRef]
- De Nisco, M.; Manfra, M.; Bolognese, A.; Sofo, A.; Scopa, A.; Tenore, G.C.; Pagano, F.; Milite, C.; Russo, M.T. Nutraceutical properties and polyphenolic profile of berry skin and wine of Vitis vinifera L. (cv. aglianico). Food Chem. 2013, 140, 623–629. [Google Scholar] [CrossRef] [PubMed]
- De Silva, A.K.H.; Rupasinghe, H.V. Polyphenols composition and anti-diabetic properties in vitro of haskap (Lonicera caerulea L.) berries in relation to cultivar and harvesting date. J. Food Compos. Anal. 2020, 88, 103402. [Google Scholar] [CrossRef]
- Debnath, A.; Kumar, A. Structural and Functional Significance of Niosome and Proniosome in Drug Delivery System. Int. J. Pharm. Eng. 2015, 3, 621–637. [Google Scholar]
- Siriwoharn, T.; Wrolstad, R.E. Polyphenolic Composition of Marion and Evergreen Blackberries. J. Food Sci. 2004, 69, FCT233–FCT240. [Google Scholar] [CrossRef]
- Kapasakalidis, P.G.; Rastall, R.A.; Gordon, M.H. Extraction of polyphenols from processed black currant (Ribes nigrum L.) residues. J. Agric. Food Chem. 2006, 54, 4016–4021. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.A.; Troup, G.J.; Pilbrow, J.R.; Hutton, D.R.; Hewitt, D.; Hunter, C.R.; Ristic, R.; Iland, P.G.; Jones, G.P. Development of seed polyphenols in berries from Vitis vinifera L. cv. shiraz. Aust. J. Grape Wine Res. 2000, 6, 244–254. [Google Scholar] [CrossRef]
- Kusznierewicz, B.; Piekarska, A.; Mrugalska, B.; Konieczka, P.; Namieśnik, J.; Bartoszek, A. Phenolic Composition and Antioxidant Properties of Polish Blue-Berried Honeysuckle Genotypes by HPLC-DAD-MS, HPLC Postcolumn Derivatization with ABTS or FC, and TLC with DPPH Visualization. J. Agric. Food Chem. 2012, 60, 1755–1763. [Google Scholar] [CrossRef]
- Lavola, A.; Karjalainen, R.; Julkunen-Tiitto, R. Bioactive polyphenols in leaves, stems, and berries of saskatoon (Amelanchier alnifolia Nutt.) cultivars. J. Agric. Food Chem. 2012, 60, 1020–1027. [Google Scholar] [CrossRef]
- Montoro, P.; Tuberoso, C.I.; Piacente, S.; Perrone, A.; De Feo, V.; Cabras, P.; Pizza, C. Stability and antioxidant activity of polyphenols in extracts of Myrtus communis L. berries used for the preparation of myrtle liqueur. J. Pharm. Biomed. Anal. 2006, 41, 1614–1619. [Google Scholar] [CrossRef]
- Mozetič, B.; Tomažič, I.; Škvarč, A.; Trebše, P. Determination of polyphenols in white grape berries cv. rebula. Acta Chim. Slov. 2006, 53, 58–64. [Google Scholar]
- Nicoletti, I.; Bello, C.; De Rossi, A.; Corradini, D. Identification and Quantification of Phenolic Compounds in Grapes by HPLC-PDA-ESI-MS on a Semimicro Separation Scale. J. Agric. Food Chem. 2008, 56, 8801–8808. [Google Scholar] [CrossRef] [PubMed]
- Oszmianski, J.; Wojdyl̷o, A.; Gorzelany, J.; Kapusta, I. Identification and Characterization of Low Molecular Weight Polyphenols in Berry Leaf Extracts by HPLC-DAD and LC-ESI/MS. J. Agric. Food Chem. 2011, 59, 12830–12835. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Chiodelli, G.; Giuberti, G.; Ghisoni, S.; Baccolo, G.; Blasi, F.; Montesano, D.; Trevisan, M.; Lucini, L. UHPLC-ESI-QTOF-MS profile of polyphenols in goji berries (Lycium barbarum L.) and its dynamics during in vitro gastrointestinal digestion and fermentation. J. Funct. Foods 2018, 40, 564–572. [Google Scholar] [CrossRef]
- Ruiz, A.; Hermosin-Gutierrez, I.; Mardones, C.; Vergara, C.; Herlitz, E.; Vega, M.; Dorau, C.; Winterhalter, P.; von Baer, D. Polyphenols and antioxidant activity of calafate (Berberis microphylla) fruits and other native berries from southern chile. J. Agric. Food Chem. 2010, 58, 6081–6089. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Hermosín-Gutiérrez, I.; Vergara, C.; von Baer, D.; Zapata, M.; Hitschfeld, A.; Obando, L.; Mardones, C. Anthocyanin Profiles in South Patagonian Wild Berries by HPLC-DAD-ESI-MS/MS. Food Res. Int. 2013, 51, 706–713. [Google Scholar] [CrossRef]
- Sakkiadi, A.-V.; Georgiou, C.A.; Haroutounian, S.A. A Standard Addition Method to Assay the Concentration of Biologically Interesting Polyphenols in Grape Berries by Reversed-Phase HPLC. Molecules 2007, 12, 2259–2269. [Google Scholar] [CrossRef] [Green Version]
- Simirgiotis, M.J.; Bórquez, J.; Schmeda-Hirschmann, G. Antioxidant Capacity, Polyphenolic Content and Tandem HPLC–DAD–ESI/MS Profiling of Phenolic Compounds from the South American Berries Luma Apiculata and L. Chequén. Food Chem. 2013, 139, 289–299. [Google Scholar] [CrossRef]
- Tzanova, M.; Atanassova, S.; Atanasov, V.; Grozeva, N. Content of Polyphenolic Compounds and Antioxidant Potential of Some Bulgarian Red Grape Varieties and Red Wines, Determined by HPLC, UV, and NIR Spectroscopy. Agriculture 2020, 10, 193. [Google Scholar] [CrossRef]
- Wang, L.-J.; Su, S.; Wu, J.; Du, H.; Li, S.-S.; Huo, J.-W.; Zhang, Y.; Wang, L.-S. Variation of Anthocyanins and Flavonols in Vaccinium Uliginosum Berry in Lesser Khingan Mountains and Its Antioxidant Activity. Food Chem. 2014, 160, 357–364. [Google Scholar] [CrossRef]
- Xiao, T.; Guo, Z.; Sun, B.; Zhao, Y. Identification of Anthocyanins from Four Kinds of Berries and Their Inhibition Activity to α-Glycosidase and Protein Tyrosine Phosphatase 1B by HPLC–FT-ICR MS/MS. J. Agric. Food Chem. 2017, 65, 6211–6221. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Byrnes, D.R.; Dinssa, F.F.; Simon, J.E.; Wu, Q. Identification of Polyphenols, Glycoalkaloids, and Saponins in Solanum Scabrum Berries Using HPLC-UV/Vis-MS. J. Food Sci. 2019, 84, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Zorzi, M.; Gai, F.; Medana, C.; Aigotti, R.; Peiretti, P.G. Identification of Polyphenolic Compounds in Edible Wild Fruits Grown in the North-West of Italy by Means of HPLC-DAD-ESI HRMS. Plant Foods Hum. Nutr. 2020, 75, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yuan, B.; Kshatriya, D.; Polyak, A.; Simon, J.E.; Bello, N.T.; Wu, Q. Influence of Diet-Induced Obesity on the Bioavailability and Metabolism of Raspberry Ketone (4-(4-Hydroxyphenyl)-2-Butanone) in Mice. Mol. Nutr. Food Res. 2020, 64, 1900907. [Google Scholar] [CrossRef] [PubMed]
- Mueller, D.; Jung, K.; Winter, M.; Rogoll, D.; Melcher, R.; Richling, E. Human Intervention Study to Investigate the Intestinal Accessibility and Bioavailability of Anthocyanins from Bilberries. Food Chem. 2017, 231, 275–286. [Google Scholar] [CrossRef]
- Annunziata, G.; Maisto, M.; Schisano, C.; Ciampaglia, R.; Daliu, P.; Narciso, V.; Tenore, G.C.; Novellino, E. Colon Bioaccessibility and Antioxidant Activity of White, Green and Black Tea Polyphenols Extract after in Vitro Simulated Gastrointestinal Digestion. Nutrients 2018, 10, 1711. [Google Scholar] [CrossRef] [Green Version]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Németh, K.; Plumb, G.W.; Berrin, J.-G.; Juge, N.; Jacob, R.; Naim, H.Y.; Williamson, G.; Swallow, D.M.; Kroon, P.A. Deglycosylation by Small Intestinal Epithelial Cell β-Glucosidases Is a Critical Step in the Absorption and Metabolism of Dietary Flavonoid Glycosides in Humans. Eur. J. Nutr. 2003, 42, 29–42. [Google Scholar] [CrossRef]
- Crozier, A.; Del Rio, D.; Clifford, M.N. Bioavailability of Dietary Flavonoids and Phenolic Compounds. Mol. Asp. Med. 2010, 31, 446–467. [Google Scholar] [CrossRef]
- Rzepecka-Stojko, A.; Stojko, J.; Kurek-Górecka, A.; Górecki, M.; Kabała-Dzik, A.; Kubina, R.; Moździerz, A.; Buszman, E. Polyphenols from Bee Pollen: Structure, Absorption, Metabolism and Biological Activity. Molecules 2015, 20, 21732–21749. [Google Scholar] [CrossRef] [Green Version]
- Day, A.J.; DuPont, M.S.; Ridley, S.; Rhodes, M.; Rhodes, M.J.; Morgan, M.R.; Williamson, G. Deglycosylation of Flavonoid and Isoflavonoid Glycosides by Human Small Intestine and Liver β-Glucosidase Activity. FEBS Lett. 1998, 436, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Walgren, R.A.; Lin, J.-T.; Kinne, R.K.-H.; Walle, T. Cellular Uptake of Dietary Flavonoid Quercetin 4′-β-Glucoside by Sodium-Dependent Glucose Transporter SGLT1. J. Pharmacol. Exp. Ther. 2000, 294, 837–843. [Google Scholar]
- Ward, N.C.; Croft, K.D.; Puddey, I.B.; Hodgson, J.M. Supplementation with Grape Seed Polyphenols Results in Increased Urinary Excretion of 3-Hydroxyphenylpropionic Acid, an Important Metabolite of Proanthocyanidins in Humans. J. Agric. Food Chem. 2004, 52, 5545–5549. [Google Scholar] [CrossRef] [PubMed]
- Urpi-Sarda, M.; Monagas, M.; Khan, N.; Llorach, R.; Lamuela-Raventós, R.M.; Jáuregui, O.; Estruch, R.; Izquierdo-Pulido, M.; Andrés-Lacueva, C. Targeted Metabolic Profiling of Phenolics in Urine and Plasma after Regular Consumption of Cocoa by Liquid Chromatography–Tandem Mass Spectrometry. J. Chromatogr. A 2009, 1216, 7258–7267. [Google Scholar] [CrossRef] [PubMed]
- Actis-Goretta, L.; Lévèques, A.; Giuffrida, F.; Romanov-Michailidis, F.; Viton, F.; Barron, D.; Duenas-Paton, M.; Gonzalez-Manzano, S.; Santos-Buelga, C.; Williamson, G. Elucidation of (−)-Epicatechin Metabolites after Ingestion of Chocolate by Healthy Humans. Free Radic. Biol. Med. 2012, 53, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Capanoglu, E.; Grootaert, C.; Van Camp, J. Anthocyanin Absorption and Metabolism by Human Intestinal Caco-2 Cells—A Review. Int. J. Mol. Sci. 2015, 16, 21555–21574. [Google Scholar] [CrossRef] [Green Version]
- Rios, L.Y.; Gonthier, M.-P.; Rémésy, C.; Mila, I.; Lapierre, C.; Lazarus, S.A.; Williamson, G.; Scalbert, A. Chocolate Intake Increases Urinary Excretion of Polyphenol-Derived Phenolic Acids in Healthy Human Subjects. Am. J. Clin. Nutr. 2003, 77, 912–918. [Google Scholar] [CrossRef] [Green Version]
- Stalmach, A.; Troufflard, S.; Serafini, M.; Crozier, A. Absorption, Metabolism and Excretion of Choladi Green Tea Flavan-3-Ols by Humans. Mol. Nutr. Food Res. 2009, 53, S44–S53. [Google Scholar] [CrossRef]
- Mattila, P.; Hellström, J.; Törrönen, R. Phenolic Acids in Berries, Fruits, and Beverages. J. Agric. Food Chem. 2006, 54, 7193–7199. [Google Scholar] [CrossRef]
- Godos, J.; Sinatra, D.; Blanco, I.; Mulè, S.; La Verde, M.; Marranzano, M. Association between Dietary Phenolic Acids and Hypertension in a Mediterranean Cohort. Nutrients 2017, 9, 1069. [Google Scholar] [CrossRef] [Green Version]
- Lafay, S.; Gil-Izquierdo, A. Bioavailability of Phenolic Acids. Phytochem. Rev. 2008, 7, 301–311. [Google Scholar] [CrossRef]
- Saibabu, V.; Fatima, Z.; Khan, L.A.; Hameed, S. Therapeutic Potential of Dietary Phenolic Acids. Adv. Pharmacol. Sci. 2015, 2015, 823539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariza, M.T.; Reboredo-Rodríguez, P.; Cervantes, L.; Soria, C.; Martínez-Ferri, E.; González-Barreiro, C.; Cancho-Grande, B.; Battino, M.; Simal-Gándara, J. Bioaccessibility and Potential Bioavailability of Phenolic Compounds from Achenes as a New Target for Strawberry Breeding Programs. Food Chem. 2018, 248, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Edwards, A.; Jiménez-Aspee, F.; Thomas-Valdés, S.; Schmeda-Hirschmann, G.; Theoduloz, C. Qualitative and Quantitative Changes in Polyphenol Composition and Bioactivity of Ribes Magellanicum and R. Punctatum after in Vitro Gastrointestinal Digestion. Food Chem. 2017, 237, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Grootaert, C.; Voorspoels, S.; Jacobs, G.; Pitart, J.; Kamiloglu, S.; Possemiers, S.; Heinonen, M.; Kardum, N.; Glibetic, M. Aronia (Aronia melanocarpa) phenolics bioavailability in a combined in vitro Digestion/Caco-2 cell model is structure and colon region dependent. J. Funct. Foods 2017, 38, 128–139. [Google Scholar] [CrossRef]
- Wu, X.; Pittman III, H.E.; Hager, T.; Hager, A.; Howard, L.; Prior, R.L. Phenolic Acids in Black Raspberry and in the Gastrointestinal Tract of Pigs Following Ingestion of Black Raspberry. Mol. Nutr. Food Res. 2009, 53, S76–S84. [Google Scholar] [CrossRef]
- Khanal, R.; Howard, L.R.; Prior, R.L. Urinary Excretion of Phenolic Acids in Rats Fed Cranberry, Blueberry, or Black Raspberry Powder. J. Agric. Food Chem. 2014, 62, 3987–3996. [Google Scholar] [CrossRef]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human Metabolism and Elimination of the Anthocyanin, Cyanidin-3-Glucoside: A 13C-Tracer Study. Am. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef] [Green Version]
- Stalmach, A.; Edwards, C.A.; Wightman, J.D.; Crozier, A. Gastrointestinal Stability and Bioavailability of (Poly) Phenolic Compounds Following Ingestion of Concord Grape Juice by Humans. Mol. Nutr. Food Res. 2012, 56, 497–509. [Google Scholar] [CrossRef]
- Castañeda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical Studies of Anthocyanins: A Review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Correa-Betanzo, J.; Allen-Vercoe, E.; McDonald, J.; Schroeter, K.; Corredig, M.; Paliyath, G. Stability and biological activity of wild blueberry (Vaccinium angustifolium) polyphenols during simulated in vitro gastrointestinal digestion. Food Chem. 2014, 165, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Yang, J.; Cui, J.; Fan, Y.; Li, N.; Wang, C.; Liu, Y.; Dong, Y. Stability and Mechanism of Phenolic Compounds from Raspberry Extract under in Vitro Gastrointestinal Digestion. LWT 2021, 139, 110552. [Google Scholar] [CrossRef]
- Bermúdez-Soto, M.-J.; Tomás-Barberán, F.-A.; García-Conesa, M.-T. Stability of polyphenols in chokeberry (Aronia melanocarpa) subjected to in vitro gastric and pancreatic digestion. Food Chem. 2007, 102, 865–874. [Google Scholar] [CrossRef]
- He, X.; Li, X.; Lv, Y.; He, Q. Composition and Color Stability of Anthocyanin-Based Extract from Purple Sweet Potato. Food Sci. Technol. 2015, 35, 468–473. [Google Scholar] [CrossRef] [Green Version]
- Van de Velde, F.; Pirovani, M.E.; Drago, S.R. Bioaccessibility Analysis of Anthocyanins and Ellagitannins from Blackberry at Simulated Gastrointestinal and Colonic Levels. J. Food Compos. Anal. 2018, 72, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.; de Pascual-Teresa, S. Metabolism of Anthocyanins by Human Gut Microflora and Their Influence on Gut Bacterial Growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef]
- Kim, I.; Moon, J.K.; Hur, S.J.; Lee, J. Structural changes in mulberry (Morus microphylla. Buckl) and chokeberry (Aronia melanocarpa) anthocyanins during simulated in vitro human digestion. Food Chem. 2020, 318, 126449. [Google Scholar] [CrossRef]
- Vitaglione, P.; Donnarumma, G.; Napolitano, A.; Galvano, F.; Gallo, A.; Scalfi, L.; Fogliano, V. Protocatechuic Acid Is the Major Human Metabolite of Cyanidin-Glucosides. J. Nutr. 2007, 137, 2043–2048. [Google Scholar] [CrossRef]
- Cheng, J.-R.; Liu, X.-M.; Chen, Z.-Y.; Zhang, Y.-S.; Zhang, Y.-H. Mulberry Anthocyanin Biotransformation by Intestinal Probiotics. Food Chem. 2016, 213, 721–727. [Google Scholar] [CrossRef]
- Matsumoto, H.; Inaba, H.; Kishi, M.; Tominaga, S.; Hirayama, M.; Tsuda, T. Orally Administered Delphinidin 3-Rutinoside and Cyanidin 3-Rutinoside Are Directly Absorbed in Rats and Humans and Appear in the Blood as the Intact Forms. J. Agric. Food Chem. 2001, 49, 1546–1551. [Google Scholar] [CrossRef]
- Murkovic, M.; Mülleder, U.; Adam, U.; Pfannhauser, W. Detection of Anthocyanins from Elderberry Juice in Human Urine. J. Sci. Food Agric. 2001, 81, 934–937. [Google Scholar] [CrossRef]
- Milbury, P.E.; Cao, G.; Prior, R.L.; Blumberg, J. Bioavailablility of Elderberry Anthocyanins. Mech. Ageing Dev. 2002, 123, 997–1006. [Google Scholar] [CrossRef]
- Netzel, M.; Strass, G.; Janssen, M.; Bitsch, I.; Bitsch, R. Bioactive Anthocyanins Detected in Human Urine after Ingestion of Blackcurrant Juice. J. Environ. Pathol. Toxicol. Oncol. 2001, 20, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Charron, C.S.; Clevidence, B.A.; Britz, S.J.; Novotny, J.A. Effect of Dose Size on Bioavailability of Acylated and Nonacylated Anthocyanins from Red Cabbage (Brassica oleracea L. var. capitata). J. Agric. Food Chem. 2007, 55, 5354–5362. [Google Scholar] [CrossRef]
- Bitsch, I.; Janssen, M.; Netzel, M.; Strass, G.; Frank, T. Bioavailability of Anthocyanidin-3-Glycosides Following Consumption of Elderberry Extract and Blackcurrant Juice. Int. J. Clin. Pharmacol. Ther. 2004, 42, 293–300. [Google Scholar] [CrossRef]
- Erlund, I.; Freese, R.; Marniemi, J.; Hakala, P.; Alfthan, G. Bioavailability of Quercetin from Berries and the Diet. Nutr. Cancer 2006, 54, 13–17. [Google Scholar] [CrossRef]
- Clifford, M.N.; Scalbert, A. Ellagitannins–Nature, Occurrence and Dietary Burden. J. Sci. Food Agric. 2000, 80, 1118–1125. [Google Scholar] [CrossRef]
- Espín, J.C.; Larrosa, M.; García-Conesa, M.T.; Tomás-Barberán, F. Biological Significance of Urolithins, the Gut Microbial Ellagic Acid-Derived Metabolites: The Evidence so Far. Evid. Based Complement. Altern. Med. 2013, 2013, 270418. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, A.K.; Miller, M.G.; Thangthaeng, N.; Scott, T.M.; Shukitt-Hale, B.; Edirisinghe, I.; Burton-Freeman, B. Metabolic Fate of Strawberry Polyphenols after Chronic Intake in Healthy Older Adults. Food Funct. 2018, 9, 96–106. [Google Scholar] [CrossRef]
- González-Barrio, R.; Edwards, C.A.; Crozier, A. Colonic Catabolism of Ellagitannins, Ellagic Acid, and Raspberry Anthocyanins: In Vivo and in Vitro Studies. Drug Metab. Dispos. 2011, 39, 1680–1688. [Google Scholar] [CrossRef]
- Borges, G.; Roowi, S.; Rouanet, J.-M.; Duthie, G.G.; Lean, M.E.; Crozier, A. The Bioavailability of Raspberry Anthocyanins and Ellagitannins in Rats. Mol. Nutr. Food Res. 2007, 51, 714–725. [Google Scholar] [CrossRef] [PubMed]
- González-Barrio, R.; Borges, G.; Mullen, W.; Crozier, A. Bioavailability of Anthocyanins and Ellagitannins Following Consumption of Raspberries by Healthy Humans and Subjects with an Ileostomy. J. Agric. Food Chem. 2010, 58, 3933–3939. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Wang, X.; Li, L. Human Gut Microbiome: The Second Genome of Human Body. Protein Cell 2010, 1, 718–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, L.V.; Midtvedt, T.; Gordon, J.I. How Host-Microbial Interactions Shape the Nutrient Environment of the Mammalian Intestine. Annu. Rev. Nutr. 2002, 22, 283–307. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of Tea Phenolics and Their Aromatic Fecal Bacterial Metabolites on Intestinal Microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Aura, A.-M. Microbial Metabolism of Dietary Phenolic Compounds in the Colon. Phytochem. Rev. 2008, 7, 407–429. [Google Scholar] [CrossRef]
- Rechner, A.R.; Smith, M.A.; Kuhnle, G.; Gibson, G.R.; Debnam, E.S.; Srai, S.K.S.; Moore, K.P.; Rice-Evans, C.A. Colonic Metabolism of Dietary Polyphenols: Influence of Structure on Microbial Fermentation Products. Free Radic. Biol. Med. 2004, 36, 212–225. [Google Scholar] [CrossRef]
- Salminen, S.; Benno, Y.; De Vos, W. Intestinal Colonisation, Microbiota and Future Probiotics? Asia Pac. J. Clin. Nutr. 2006, 15, 558–562. [Google Scholar]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Xiao, H.; Luo, D.; Zhang, X.; Zhao, S.; Zheng, Q.; Li, Y.; Zhao, Y.; Dong, J.; Li, H. Circadian Rhythm Shapes the Gut Microbiota Affecting Host Radiosensitivity. Int. J. Mol. Sci. 2016, 17, 1786. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Xiao, H.; Li, Y.; Zhou, L.; Zhao, S.; Luo, D.; Zheng, Q.; Dong, J.; Zhao, Y.; Zhang, X. Faecal Microbiota Transplantation Protects against Radiation-Induced Toxicity. EMBO Mol. Med. 2017, 9, 448–461. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Clément, K. The Gut Microbiome, Diet, and Links to Cardiometabolic and Chronic Disorders. Nat. Rev. Nephrol. 2016, 12, 169. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Huang, K.; Zhao, C.; Xu, W.; Sheng, Y.; Luo, Y.; He, X. Procyanidin Attenuates Weight Gain and Modifies the Gut Microbiota in High Fat Diet Induced Obese Mice. J. Funct. Foods 2018, 49, 362–368. [Google Scholar] [CrossRef]
- Blaut, M.; Clavel, T. Metabolic Diversity of the Intestinal Microbiota: Implications for Health and Disease. J. Nutr. 2007, 137, 751S–755S. [Google Scholar] [CrossRef]
- Patterson, E.; Ryan, P.M.; Cryan, J.F.; Dinan, T.G.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Gut Microbiota, Obesity and Diabetes. Postgrad. Med. J. 2016, 92, 286–300. [Google Scholar] [CrossRef]
- Van der Waaij, D. Microbial Ecology of the Intestinal Microflora: Influence of Interactions with the Host Organism. In Nestle Nutrition Workshop Series; Rowen Press: Portland, Oregon, 1999; Volume 42, pp. 1–16. [Google Scholar]
- Bouhnik, Y.; Vahedi, K.; Achour, L.; Attar, A.; Salfati, J.; Pochart, P.; Marteau, P.; Flourie, B.; Bornet, F.; Rambaud, J.-C. Short-Chain Fructo-Oligosaccharide Administration Dose-Dependently Increases Fecal Bifidobacteria in Healthy Humans. J. Nutr. 1999, 129, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Verdu, E.F.; Collins, S.M. Irritable Bowel Syndrome. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 315–321. [Google Scholar] [CrossRef]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of Polyphenols on Gut Microbiota and Implications in Human Health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Hervert-Hernandez, D.; Goñi, I. Dietary Polyphenols and Human Gut Microbiota: A Review. Food Rev. Int. 2011, 27, 154–169. [Google Scholar] [CrossRef]
- Jiao, X.; Wang, Y.; Lin, Y.; Lang, Y.; Li, E.; Zhang, X.; Zhang, Q.; Feng, Y.; Meng, X.; Li, B. Blueberry Polyphenols Extract as a Potential Prebiotic with Anti-Obesity Effects on C57BL/6 J Mice by Modulating the Gut Microbiota. J. Nutr. Biochem. 2019, 64, 88–100. [Google Scholar] [CrossRef]
- Qiao, Y.; Sun, J.; Xia, S.; Tang, X.; Shi, Y.; Le, G. Effects of Resveratrol on Gut Microbiota and Fat Storage in a Mouse Model with High-Fat-Induced Obesity. Food Funct. 2014, 5, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, P.; Kalariya, H.M.; Poulev, A.; Ribnicky, D.M.; Jaja-Chimedza, A.; Roopchand, D.E.; Raskin, I. Grape Polyphenols Reduce Gut-Localized Reactive Oxygen Species Associated with the Development of Metabolic Syndrome in Mice. PLoS ONE 2018, 13, e0198716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overall, J.; Bonney, S.A.; Wilson, M.; Beermann, A.; Grace, M.H.; Esposito, D.; Lila, M.A.; Komarnytsky, S. Metabolic Effects of Berries with Structurally Diverse Anthocyanins. Int. J. Mol. Sci. 2017, 18, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puupponen-Pimiä, R.; Nohynek, L.; Meier, C.; Kähkönen, M.; Heinonen, M.; Hopia, A.; Oksman-Caldentey, K.-M. Antimicrobial Properties of Phenolic Compounds from Berries. J. Appl. Microbiol. 2001, 90, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; DeCoffe, D.; Molcan, E.; Gibson, D.L. Diet-Induced Dysbiosis of the Intestinal Microbiota and the Effects on Immunity and Disease. Nutrients 2012, 4, 1095–1119. [Google Scholar] [CrossRef] [Green Version]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E. A Polyphenol-Rich Cranberry Extract Protects from Diet-Induced Obesity, Insulin Resistance and Intestinal Inflammation in Association with Increased Akkermansia Spp. Population in the Gut Microbiota of Mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Hu, R.; Nakano, H.; Chen, K.; Liu, M.; He, X.; Zhang, H.; He, J.; Hou, D.-X. Modulation of Gut Microbiota by Lonicera caerulea L. Berry Polyphenols in a Mouse Model of Fatty Liver Induced by High Fat Diet. Molecules 2018, 23, 3213. [Google Scholar] [CrossRef] [Green Version]
- Roopchand, D.E.; Carmody, R.N.; Kuhn, P.; Moskal, K.; Rojas-Silva, P.; Turnbaugh, P.J.; Raskin, I. Dietary Polyphenols Promote Growth of the Gut Bacterium Akkermansia Muciniphila and Attenuate High-Fat Diet–Induced Metabolic Syndrome. Diabetes 2015, 64, 2847–2858. [Google Scholar] [CrossRef] [Green Version]
- Hester, S.N.; Mastaloudis, A.; Gray, R.; Antony, J.M.; Evans, M.; Wood, S.M. Efficacy of an Anthocyanin and Prebiotic Blend on Intestinal Environment in Obese Male and Female Subjects. J. Nutr. Metab. 2018, 2018, 7497260. [Google Scholar] [CrossRef]
- Gu, J.; Thomas-Ahner, J.M.; Riedl, K.M.; Bailey, M.T.; Vodovotz, Y.; Schwartz, S.J.; Clinton, S.K. Dietary Black Raspberries Impact the Colonic Microbiome and Phytochemical Metabolites in Mice. Mol. Nutr. Food Res. 2019, 63, 1800636. [Google Scholar] [CrossRef]
- Matziouridou, C.; Marungruang, N.; Nguyen, T.D.; Nyman, M.; Fåk, F. Lingonberries Reduce Atherosclerosis in Apoe−/− Mice in Association with Altered Gut Microbiota Composition and Improved Lipid Profile. Mol. Nutr. Food Res. 2016, 60, 1150–1160. [Google Scholar] [CrossRef] [PubMed]
- Molan, A.-L.; Liu, Z.; Kruger, M. The Ability of Blackcurrant Extracts to Positively Modulate Key Markers of Gastrointestinal Function in Rats. World J. Microbiol. Biotechnol. 2010, 26, 1735–1743. [Google Scholar] [CrossRef]
- Vendrame, S.; Guglielmetti, S.; Riso, P.; Arioli, S.; Klimis-Zacas, D.; Porrini, M. Six-Week Consumption of a Wild Blueberry Powder Drink Increases Bifidobacteria in the Human Gut. J. Agric. Food Chem. 2011, 59, 12815–12820. [Google Scholar] [CrossRef] [PubMed]
- Molan, A.-L.; Liu, Z.; Plimmer, G. Evaluation of the Effect of Blackcurrant Products on Gut Microbiota and on Markers of Risk for Colon Cancer in Humans. Phytother. Res. 2014, 28, 416–422. [Google Scholar] [CrossRef]
- Song, M.; Wang, J.; Eom, T.; Kim, H. Schisandra Chinensis Fruit Modulates the Gut Microbiota Composition in Association with Metabolic Markers in Obese Women: A Randomized, Double-Blind Placebo-Controlled Study. Nutr. Res. 2015, 35, 655–663. [Google Scholar] [CrossRef]
- Sánchez-Patán, F.; Barroso, E.; Van de Wiele, T.; Jiménez-Girón, A.; Martín-Alvarez, P.J.; Moreno-Arribas, M.V.; Martínez-Cuesta, M.C.; Peláez, C.; Requena, T.; Bartolomé, B. Comparative in Vitro Fermentations of Cranberry and Grape Seed Polyphenols with Colonic Microbiota. Food Chem. 2015, 183, 273–282. [Google Scholar] [CrossRef]
- Alqurashi, R.M.; Alarifi, S.N.; Walton, G.E.; Costabile, A.F.; Rowland, I.R.; Commane, D.M. In vitro approaches to assess the effects of acai (Euterpe oleracea) digestion on polyphenol availability and the subsequent impact on the faecal microbiota. Food Chem. 2017, 234, 190–198. [Google Scholar] [CrossRef]
- Yan, Y.; Peng, Y.; Tang, J.; Mi, J.; Lu, L.; Li, X.; Ran, L.; Zeng, X.; Cao, Y. Effects of Anthocyanins from the Fruit of Lycium Ruthenicum Murray on Intestinal Microbiota. J. Funct. Foods 2018, 48, 533–541. [Google Scholar] [CrossRef]
- Kemperman, R.A.; Bolca, S.; Roger, L.C.; Vaughan, E.E. Novel Approaches for Analysing Gut Microbes and Dietary Polyphenols: Challenges and Opportunities. Microbiology 2010, 156, 3224–3231. [Google Scholar] [CrossRef] [Green Version]
- Ferrazzano, G.F.; Amato, I.; Ingenito, A.; Zarrelli, A.; Pinto, G.; Pollio, A. Plant Polyphenols and Their Anti-Cariogenic Properties: A Review. Molecules 2011, 16, 1486–1507. [Google Scholar] [CrossRef] [Green Version]
- Padmini, E.; Valarmathi, A.; Rani, M.U. Comparative Analysis of Chemical Composition and Antibacterial Activities of Mentha Spicata and Camellia Sinensis. Asian J. Exp. Biol. Sci. 2010, 1, 772–781. [Google Scholar]
- Hattori, M.; Kusumoto, I.T.; Namba, T.; Ishigami, T.; Hara, Y. Effect of Tea Polyphenols on Glucan Synthesis by Glucosyltransferase from Streptococcus Mutans. Chem. Pharm. Bull. 1990, 38, 717–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haslam, E.; Lilley, T.H.; Warminski, E.; Liao, H.; Cai, Y.; Martin, R.; Gaffney, S.H.; Goulding, P.N.; Luck, G. Polyphenol Complexation: A Study in Molecular Recognition. ACS Symp. Ser. 1992, 25, 325–338. [Google Scholar]
- Marques, C.; Fernandes, I.; Meireles, M.; Faria, A.; Spencer, J.P.; Mateus, N.; Calhau, C. Gut Microbiota Modulation Accounts for the Neuroprotective Properties of Anthocyanins. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pan, P.; Lam, V.; Salzman, N.; Huang, Y.-W.; Yu, J.; Zhang, J.; Wang, L.-S. Black Raspberries and Their Anthocyanin and Fiber Fractions Alter the Composition and Diversity of Gut Microbiota in F-344 Rats. Nutr. Cancer 2017, 69, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Paturi, G.; Mandimika, T.; Butts, C.A.; Zhu, S.; Roy, N.C.; McNabb, W.C.; Ansell, J. Influence of Dietary Blueberry and Broccoli on Cecal Microbiota Activity and Colon Morphology in Mdr1a−/− Mice, a Model of Inflammatory Bowel Diseases. Nutrition 2012, 28, 324–330. [Google Scholar] [CrossRef]
- Lacombe, A.; Li, R.W.; Klimis-Zacas, D.; Kristo, A.S.; Tadepalli, S.; Krauss, E.; Young, R.; Wu, V.C. Lowbush Wild Blueberries Have the Potential to Modify Gut Microbiota and Xenobiotic Metabolism in the Rat Colon. PLoS ONE 2013, 8, e67497. [Google Scholar] [CrossRef]
- Molan, A.L.; Lila, M.A.; Mawson, J.; De, S. In Vitro and in Vivo Evaluation of the Prebiotic Activity of Water-Soluble Blueberry Extracts. World J. Microbiol. Biotechnol. 2009, 25, 1243–1249. [Google Scholar] [CrossRef]
- Guglielmetti, S.; Fracassetti, D.; Taverniti, V.; Del Bo’, C.; Vendrame, S.; Klimis-Zacas, D.; Arioli, S.; Riso, P.; Porrini, M. Differential modulation of human intestinal bifidobacterium populations after consumption of a wild blueberry (Vaccinium angustifolium) drink. J. Agric. Food Chem. 2013, 61, 8134–8140. [Google Scholar] [CrossRef]
- Kang, Y.; Yang, G.; Zhang, S.; Ross, C.F.; Zhu, M.-J. Goji Berry Modulates Gut Microbiota and Alleviates Colitis in IL-10-Deficient Mice. Mol. Nutr. Food Res. 2018, 62, 1800535. [Google Scholar] [CrossRef]
- Guergoletto, K.B.; Costabile, A.; Flores, G.; Garcia, S.; Gibson, G.R. In vitro fermentation of juçara pulp (Euterpe edulis) by human colonic microbiota. Food Chem. 2016, 196, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamar, G.; Santamarina, A.B.; Mennitti, L.V.; de Cássia Cesar, H.; Oyama, L.M.; de Rosso, V.V.; Pisani, L.P. Bifidobacterium spp. reshaping in the gut microbiota by low dose of juçara supplementation and hypothalamic insulin resistance in wistar rats. J. Funct. Foods 2018, 46, 212–219. [Google Scholar] [CrossRef]
- da Silva-Maia, J.K.; Batista, A.G.; Correa, L.C.; Lima, G.C.; Bogusz Junior, S.; Maróstica Junior, M.R. Aqueous extract of berry (Plinia jaboticaba) byproduct modulates gut microbiota and maintains the balance on antioxidant defense system in rats. J. Food Biochem. 2019, 43, e12705. [Google Scholar] [CrossRef] [PubMed]
- Masumoto, S.; Terao, A.; Yamamoto, Y.; Mukai, T.; Miura, T.; Shoji, T. Non-Absorbable Apple Procyanidins Prevent Obesity Associated with Gut Microbial and Metabolomic Changes. Sci. Rep. 2016, 6, 31208. [Google Scholar] [CrossRef] [PubMed]
- Attri, S.; Sharma, K.; Raigond, P.; Goel, G. Colonic fermentation of polyphenolics from sea buckthorn (Hippophae rhamnoides) berries: Assessment of effects on microbial diversity by principal component analysis. Food Res. Int. 2018, 105, 324–332. [Google Scholar] [CrossRef]
- Attri, S.; Goel, G. Influence of Polyphenol Rich Seabuckthorn Berries Juice on Release of Polyphenols and Colonic Microbiota on Exposure to Simulated Human Digestion Model. Food Res. Int. 2018, 111, 314–323. [Google Scholar] [CrossRef]
- Petersen, C.; Wankhade, U.D.; Bharat, D.; Wong, K.; Mueller, J.E.; Chintapalli, S.V.; Piccolo, B.D.; Jalili, T.; Jia, Z.; Symons, J.D. Dietary Supplementation with Strawberry Induces Marked Changes in the Composition and Functional Potential of the Gut Microbiome in Diabetic Mice. J. Nutr. Biochem. 2019, 66, 63–69. [Google Scholar] [CrossRef]
- Loo, Y.T.; Howell, K.; Chan, M.; Zhang, P.; Ng, K. Modulation of the Human Gut Microbiota by Phenolics and Phenolic Fiber-Rich Foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1268–1298. [Google Scholar] [CrossRef]
- Morais, C.A.; Oyama, L.M.; de Moura Conrado, R.; de Rosso, V.V.; do Nascimento, C.O.; Pisani, L.P. Polyphenols-Rich Fruit in Maternal Diet Modulates Inflammatory Markers and the Gut Microbiota and Improves Colonic Expression of ZO-1 in Offspring. Food Res. Int. 2015, 77, 186–193. [Google Scholar] [CrossRef]
- Gasparrini, M.; Forbes-Hernandez, T.Y.; Cianciosi, D.; Quiles, J.L.; Mezzetti, B.; Xiao, J.; Giampieri, F.; Battino, M. The Efficacy of Berries against Lipopolysaccharide-Induced Inflammation: A Review. Trends Food Sci. Technol. 2021, 22, 6541. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olas, B. Berry Phenolic Antioxidants–Implications for Human Health? Front. Pharmacol. 2018, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Meyskens, F.L.; Szabo, E. Diet and Cancer: The Disconnect between Epidemiology and Randomized Clinical Trials. Cancer Epidemiol. Prev. Biomark. 2005, 14, 1366–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolca, S.; Van de Wiele, T.; Possemiers, S. Gut Metabotypes Govern Health Effects of Dietary Polyphenols. Curr. Opin. Biotechnol. 2013, 24, 220–225. [Google Scholar] [CrossRef]
- Frazier, T.H.; DiBaise, J.K.; McClain, C.J. Gut Microbiota, Intestinal Permeability, Obesity-Induced Inflammation, and Liver Injury. J. Parenter. Enter. Nutr. 2011, 35, 14S–20S. [Google Scholar] [CrossRef]
- Ding, S.; Chi, M.M.; Scull, B.P.; Rigby, R.; Schwerbrock, N.M.; Magness, S.; Jobin, C.; Lund, P.K. High-Fat Diet: Bacteria Interactions Promote Intestinal Inflammation Which Precedes and Correlates with Obesity and Insulin Resistance in Mouse. PLoS ONE 2010, 5, e12191. [Google Scholar] [CrossRef] [Green Version]
- Triebel, S.; Trieu, H.-L.; Richling, E. Modulation of Inflammatory gene expression by a bilberry (Vaccinium myrtillus L.) extract and single anthocyanins considering their limited stability under cell culture conditions. J. Agric. Food Chem. 2012, 60, 8902–8910. [Google Scholar] [CrossRef]
- Esposito, D.; Chen, A.; Grace, M.H.; Komarnytsky, S.; Lila, M.A. Inhibitory Effects of Wild Blueberry Anthocyanins and Other Flavonoids on Biomarkers of Acute and Chronic Inflammation in Vitro. J. Agric. Food Chem. 2014, 62, 7022–7028. [Google Scholar] [CrossRef]
- Pervin, M.; Hasnat, M.A.; Lim, J.-H.; Lee, Y.-M.; Kim, E.O.; Um, B.-H.; Lim, B.O. Preventive and therapeutic effects of blueberry (Vaccinium corymbosum) extract against dss-induced ulcerative colitis by regulation of antioxidant and inflammatory mediators. J. Nutr. Biochem. 2016, 28, 103–113. [Google Scholar] [CrossRef]
- Osman, N.; Adawi, D.; Ahrné, S.; Jeppsson, B.; Molin, G. Probiotics and Blueberry Attenuate the Severity of Dextran Sulfate Sodium (DSS)-Induced Colitis. Dig. Dis. Sci. 2008, 53, 2464–2473. [Google Scholar] [CrossRef]
- Wu, L.-H.; Xu, Z.-L.; Dong, D.; He, S.-A.; Yu, H. Protective Effect of Anthocyanins Extract from Blueberry on TNBS-Induced IBD Model of Mice. Evid. Based Complement. Altern. Med. 2011, 2011, 21785630. [Google Scholar] [CrossRef] [PubMed]
- Montrose, D.C.; Horelik, N.A.; Madigan, J.P.; Stoner, G.D.; Wang, L.-S.; Bruno, R.S.; Park, H.J.; Giardina, C.; Rosenberg, D.W. Anti-Inflammatory Effects of Freeze-Dried Black Raspberry Powder in Ulcerative Colitis. Carcinogenesis 2011, 32, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-S.; Kuo, C.-T.; Stoner, K.; Yearsley, M.; Oshima, K.; Yu, J.; Huang, T.H.-M.; Rosenberg, D.; Peiffer, D.; Stoner, G. Dietary Black Raspberries Modulate DNA Methylation in Dextran Sodium Sulfate (DSS)-Induced Ulcerative Colitis. Carcinogenesis 2013, 34, 2842–2850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piberger, H.; Oehme, A.; Hofmann, C.; Dreiseitel, A.; Sand, P.G.; Obermeier, F.; Schoelmerich, J.; Schreier, P.; Krammer, G.; Rogler, G. Bilberries and Their Anthocyanins Ameliorate Experimental Colitis. Mol. Nutr. Food Res. 2011, 55, 1724–1729. [Google Scholar] [CrossRef]
- Graf, D.; Seifert, S.; Bub, A.; Fröhling, B.; Dold, S.; Unger, F.; Römpp, A.; Watzl, B. Anthocyanin-Rich Juice Does Not Affect Gut-Associated Immunity in F Ischer Rats. Mol. Nutr. Food Res. 2013, 57, 1753–1761. [Google Scholar] [CrossRef]
- Morais, C.A.; de Rosso, V.V.; Estadella, D.; Pisani, L.P. Anthocyanins as Inflammatory Modulators and the Role of the Gut Microbiota. J. Nutr. Biochem. 2016, 33, 1–7. [Google Scholar] [CrossRef]
- Biedermann, L.; Mwinyi, J.; Scharl, M.; Frei, P.; Zeitz, J.; Kullak-Ublick, G.A.; Vavricka, S.R.; Fried, M.; Weber, A.; Humpf, H.-U. Bilberry Ingestion Improves Disease Activity in Mild to Moderate Ulcerative Colitis—An Open Pilot Study. J. Crohns Colitis 2013, 7, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Roth, S.; Spalinger, M.R.; Gottier, C.; Biedermann, L.; Zeitz, J.; Lang, S.; Weber, A.; Rogler, G.; Scharl, M. Bilberry-Derived Anthocyanins Modulate Cytokine Expression in the Intestine of Patients with Ulcerative Colitis. PLoS ONE 2016, 11, e0154817. [Google Scholar] [CrossRef] [Green Version]
- Mertens-Talcott, S.U.; Rios, J.; Jilma-Stohlawetz, P.; Pacheco-Palencia, L.A.; Meibohm, B.; Talcott, S.T.; Derendorf, H. Pharmacokinetics of anthocyanins and antioxidant effects after the consumption of anthocyanin-rich acai juice and pulp (Euterpe oleracea Mart.) in human healthy volunteers. J. Agric. Food Chem. 2008, 56, 7796–7802. [Google Scholar] [CrossRef]
- Hazafa, A.; Rehman, K.-U.; Jahan, N.; Jabeen, Z. The Role of Polyphenol (Flavonoids) Compounds in the Treatment of Cancer Cells. Nutr. Cancer 2020, 72, 386–397. [Google Scholar] [CrossRef]
- Hulvat, M.C. Cancer Incidence and Trends. Surg. Clin. 2020, 100, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Aiyer, H.; Ravoori, S.; Gupta, R. Chemopreventive Effects of Berries and Berry Components in Animal Models: Prevention of Estrogen-Mediated Mammary Tumors in ACI Rats by Berries. In Berries and Cancer Prevention; Springer: Berlin, Germany, 2011; pp. 163–187. [Google Scholar]
- Moloney. Cotter ROS Signalling in the Biology of Cancer—ScienceDirect. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/abs/pii/S1084952116303834 (accessed on 3 March 2021).
- Sharma, A.; Kaur, M.; Katnoria, J.K.; Nagpal, A.K. Polyphenols in Food: Cancer Prevention and Apoptosis Induction. Curr. Med. Chem. 2018, 25, 4740–4757. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary Polyphenols and the Prevention of Diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiecki, A.; Roomi, M.W.; Kalinovsky, T.; Rath, M. Anticancer Efficacy of Polyphenols and Their Combinations. Nutrients 2016, 8, 552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutler, G.J.; Nettleton, J.A.; Ross, J.A.; Harnack, L.J.; Jacobs, D.R., Jr.; Scrafford, C.G.; Barraj, L.M.; Mink, P.J.; Robien, K. Dietary Flavonoid Intake and Risk of Cancer in Postmenopausal Women: The Iowa Women’s Health Study. Int. J. Cancer 2008, 123, 664–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mursu, J.; Robien, K.; Harnack, L.J.; Park, K.; Jacobs, D.R. Dietary Supplements and Mortality Rate in Older Women: The Iowa Women’s Health Study. Arch. Intern. Med. 2011, 171, 1625–1633. [Google Scholar] [CrossRef]
- Mentor-Marcel, R.A.; Bobe, G.; Sardo, C.; Wang, L.-S.; Kuo, C.-T.; Stoner, G.; Colburn, N.H. Plasma Cytokines as Potential Response Indicators to Dietary Freeze-Dried Black Raspberries in Colorectal Cancer Patients. Nutr. Cancer 2012, 64, 820–825. [Google Scholar] [CrossRef] [Green Version]
- Thanikachalam, K.; Khan, G. Colorectal Cancer and Nutrition. Nutrients 2019, 11, 164. [Google Scholar] [CrossRef] [Green Version]
- Haraldsdottir, S.; Einarsdottir, H.M.; Smaradottir, A.; Gunnlaugsson, A.; Halfdanarson, T.R. Colorectal Cancer-Review. Laeknabladid 2014, 100, 75–82. [Google Scholar]
- Castells, A. Hereditary Forms of Colorectal Cancer. Gastroenterol. Hepatol. 2016, 39, 62–67. [Google Scholar] [CrossRef]
- Heinimann, K. Erblicher Darmkrebs: Klinik, Diagnostik Und Management. Ther. Umsch. 2018, 75, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Pozza, A.; Scarpa, M.; Ruffolo, C.; Polese, L.; Erroi, F.; Bridda, A.; Norberto, L.; Frego, M. Colonic Carcinogenesis in IBD: Molecular Events. Ann. Ital. Chir. 2011, 82, 19–28. [Google Scholar] [PubMed]
- Rubin, D.C.; Shaker, A.; Levin, M.S. Chronic Intestinal Inflammation: Inflammatory Bowel Disease and Colitis-Associated Colon Cancer. Front. Immunol. 2012, 3, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeram, N.P.; Adams, L.S.; Zhang, Y.; Lee, R.; Sand, D.; Scheuller, H.S.; Heber, D. Blackberry, Black Raspberry, Blueberry, Cranberry, Red Raspberry, and Strawberry Extracts Inhibit Growth and Stimulate Apoptosis of Human Cancer Cells in Vitro. J. Agric. Food Chem. 2006, 54, 9329–9339. [Google Scholar] [CrossRef]
- McDougall, G.J.; Ross, H.A.; Ikeji, M.; Stewart, D. Berry Extracts Exert Different Antiproliferative Effects against Cervical and Colon Cancer Cells Grown in Vitro. J. Agric. Food Chem. 2008, 56, 3016–3023. [Google Scholar] [CrossRef]
- Duthie, S.J.; Gardner, P.T.; Morrice, P.C.; Wood, S.G.; Pirie, L.; Bestwick, C.C.; Milne, L.; Duthie, G.G. DNA Stability and Lipid Peroxidation in Vitamin E–Deficient Rats in Vivo and Colon Cells in Vitro. Eur. J. Nutr. 2005, 44, 195–203. [Google Scholar] [CrossRef]
- Elisia, I.; Kitts, D.D. Anthocyanins Inhibit Peroxyl Radical-Induced Apoptosis in Caco-2 Cells. Mol. Cell. Biochem. 2008, 312, 139–145. [Google Scholar] [CrossRef]
- Neto, C.C.; Krueger, C.G.; Lamoureaux, T.L.; Kondo, M.; Vaisberg, A.J.; Hurta, R.A.; Curtis, S.; Matchett, M.D.; Yeung, H.; Sweeney, M.I. MALDI-TOF MS Characterization of Proanthocyanidins from Cranberry Fruit (Vaccinium macrocarpon) That Inhibit Tumor Cell Growth and Matrix Metalloproteinase Expression in Vitro. J. Sci. Food Agric. 2006, 86, 18–25. [Google Scholar] [CrossRef]
- Ferguson, P.J.; Kurowska, E.; Freeman, D.J.; Chambers, A.F.; Koropatnick, D.J. A Flavonoid Fraction from Cranberry Extract Inhibits Proliferation of Human Tumor Cell Lines. J. Nutr. 2004, 134, 1529–1535. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Giusti, M.M.; Malik, M.; Moyer, M.P.; Magnuson, B.A. Effects of Commercial Anthocyanin-Rich Extracts on Colonic Cancer and Nontumorigenic Colonic Cell Growth. J. Agric. Food Chem. 2004, 52, 6122–6128. [Google Scholar] [CrossRef]
- Shi, J.B.; Tang, W.J.; Qi, X.B.; Li, R.; Liu, X.H. Novel Pyrazole-5-Carboxamide and Pyrazole–Pyrimidine Derivatives: Synthesis and Anticancer Activity. Eur. J. Med. Chem. 2015, 90, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.K.; Koponen, J.M.; Mykkänen, H.M.; Törrönen, A.R. Berry Phenolic Extracts Modulate the Expression of P21WAF1 and Bax but Not Bcl-2 in HT-29 Colon Cancer Cells. J. Agric. Food Chem. 2007, 55, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Seeram, N.P.; Lee, R.; Feng, L.; Heber, D. Isolation and Identification of Strawberry Phenolics with Antioxidant and Human Cancer Cell Antiproliferative Properties. J. Agric. Food Chem. 2008, 56, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Renis, M.; Calandra, L.; Scifo, C.; Tomasello, B.; Cardile, V.; Vanella, L.; Bei, R.; La Fauci, L.; Galvano, F. Response of Cell Cycle/Stress-Related Protein Expression and DNA Damage upon Treatment of CaCo2 Cells with Anthocyanins. Br. J. Nutr. 2008, 100, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Danbara, N.; Yuri, T.; Tsujita-Kyutoku, M.; Tsukamoto, R.; Uehara, N.; Tsubura, A. Enterolactone Induces Apoptosis and Inhibits Growth of Colo 201 Human Colon Cancer Cells Both in Vitro and in Vivo. Anticancer Res. 2005, 25, 2269–2276. [Google Scholar]
- Lala, G.; Malik, M.; Zhao, C.; He, J.; Kwon, Y.; Giusti, M.M.; Magnuson, B.A. Anthocyanin-Rich Extracts Inhibit Multiple Biomarkers of Colon Cancer in Rats. Nutr. Cancer 2006, 54, 84–93. [Google Scholar] [CrossRef]
- Wang, L.-S.; Arnold, M.; Huang, Y.-W.; Sardo, C.; Seguin, C.; Martin, E.; Huang, T.H.-M.; Riedl, K.; Schwartz, S.; Frankel, W. Modulation of Genetic and Epigenetic Biomarkers of Colorectal Cancer in Humans by Black Raspberries: A Phase I Pilot Study. Clin. Cancer Res. 2011, 17, 598–610. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-S.; Kuo, C.-T.; Cho, S.-J.; Seguin, C.; Siddiqui, J.; Stoner, K.; Weng, Y.-I.; Huang, T.H.-M.; Tichelaar, J.; Yearsley, M. Black Raspberry-Derived Anthocyanins Demethylate Tumor Suppressor Genes through the Inhibition of DNMT1 and DNMT3B in Colon Cancer Cells. Nutr. Cancer 2013, 65, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Pan, P.; Skaer, C.W.; Stirdivant, S.M.; Young, M.R.; Stoner, G.D.; Lechner, J.F.; Huang, Y.-W.; Wang, L.-S. Beneficial Regulation of Metabolic Profiles by Black Raspberries in Human Colorectal Cancer Patients. Cancer Prev. Res. 2015, 8, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Schetter, A.J.; Nguyen, G.H.; Bowman, E.D.; Mathé, E.A.; Yuen, S.T.; Hawkes, J.E.; Croce, C.M.; Leung, S.Y.; Harris, C.C. Association of Inflammation-Related and MicroRNA Gene Expression with Cancer-Specific Mortality of Colon Adenocarcinoma. Clin. Cancer Res. 2009, 15, 5878–5887. [Google Scholar] [CrossRef] [Green Version]
- Gauliard, B.; Grieve, D.; Wilson, R.; Crozier, A.; Jenkins, C.; Mullen, W.D.; Lean, M. The Effects of Dietary Phenolic Compounds on Cytokine and Antioxidant Production by A549 Cells. J. Med. Food 2008, 11, 382–384. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-S.; Burke, C.A.; Hasson, H.; Kuo, C.-T.; Molmenti, C.L.S.; Seguin, C.; Liu, P.; Huang, T.H.-M.; Frankel, W.L.; Stoner, G.D. A Phase Ib Study of the Effects of Black Raspberries on Rectal Polyps in Patients with Familial Adenomatous Polyposis. Cancer Prev. Res. 2014, 7, 666–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Ning, S. Cyanidin-3-Glucoside Attenuates the Angiogenesis of Breast Cancer via Inhibiting STAT3/VEGF Pathway. Phytother. Res. 2019, 33, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dydjow-Bendek, D.; Zagożdżon, P. Selected Dietary Factors and Breast Cancer Risk. Przegl. Epidemiol. 2019, 73, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Terrisse, S.; Derosa, L.; Iebba, V.; Ghiringhelli, F.; Vaz-Luis, I.; Kroemer, G.; Fidelle, M.; Christodoulidis, S.; Segata, N.; Thomas, A.M. Intestinal Microbiota Influences Clinical Outcome and Side Effects of Early Breast Cancer Treatment. Cell Death Differ. 2021, 28, 2778–2796. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xia, Y.; Sun, J. Breast and Gut Microbiome in Health and Cancer. Genes Dis. 2021, 8, 581–589. [Google Scholar] [CrossRef]
- Xu, M.; Bower, K.A.; Wang, S.; Frank, J.A.; Chen, G.; Ding, M.; Wang, S.; Shi, X.; Ke, Z.; Luo, J. Cyanidin-3-Glucoside Inhibits Ethanol-Induced Invasion of Breast Cancer Cells Overexpressing ErbB2. Mol. Cancer 2010, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, J.; Tang, X.; Liu, Y.; Yu, X.; Wang, Z.; Liu, W. Anthocyanins Inhibit Trastuzumab-Resistant Breast Cancer in Vitro and in Vivo. Mol. Med. Rep. 2016, 13, 4007–4013. [Google Scholar] [CrossRef] [Green Version]
- Di Modica, M.; Gargari, G.; Regondi, V.; Bonizzi, A.; Arioli, S.; Belmonte, B.; De Cecco, L.; Fasano, E.; Bianchi, F.; Bertolotti, A. Gut Microbiota Condition the Therapeutic Efficacy of Trastuzumab in HER2-Positive Breast Cancer. Cancer Res. 2021, 81, 2195–2206. [Google Scholar] [CrossRef]
- Cho, E.; Chung, E.Y.; Jang, H.-Y.; Hong, O.-Y.; Chae, H.S.; Jeong, Y.-J.; Kim, S.-Y.; Kim, B.-S.; Yoo, D.J.; Kim, J.-S. Anti-Cancer Effect of Cyanidin-3-Glucoside from Mulberry via Caspase-3 Cleavage and DNA Fragmentation in Vitro and in Vivo. Anti-Cancer Agents Med. Chem. Former. Curr. Med. Chem.-Anti-Cancer Agents 2017, 17, 1519–1525. [Google Scholar] [CrossRef]
- Somasagara, R.R.; Hegde, M.; Chiruvella, K.K.; Musini, A.; Choudhary, B.; Raghavan, S.C. Extracts of Strawberry Fruits Induce Intrinsic Pathway of Apoptosis in Breast Cancer Cells and Inhibits Tumor Progression in Mice. PLoS ONE 2012, 7, e47021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeyabalan, J.; Aqil, F.; Munagala, R.; Annamalai, L.; Vadhanam, M.V.; Gupta, R.C. Chemopreventive and Therapeutic Activity of Dietary Blueberry against Estrogen-Mediated Breast Cancer. J. Agric. Food Chem. 2014, 62, 3963–3971. [Google Scholar] [CrossRef]
- Ravoori, S.; Vadhanam, M.V.; Aqil, F.; Gupta, R.C. Inhibition of Estrogen-Mediated Mammary Tumorigenesis by Blueberry and Black Raspberry. J. Agric. Food Chem. 2012, 60, 5547–5555. [Google Scholar] [CrossRef] [PubMed]
- Peiffer, D.S. Preparing Black Raspberry Components for Their Use as Cancer Therapeutics. J. Berry Res. 2018, 8, 297–306. [Google Scholar] [CrossRef]
- Saarinen, N.M.; Wärri, A.; Dings, R.P.; Airio, M.; Smeds, A.I.; Mäkelä, S. Dietary Lariciresinol Attenuates Mammary Tumor Growth and Reduces Blood Vessel Density in Human MCF-7 Breast Cancer Xenografts and Carcinogen-Induced Mammary Tumors in Rats. Int. J. Cancer 2008, 123, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Adams, L.S.; Chen, S.; Killian, C.; Ahmed, A.; Seeram, N.P. Eugenia Jambolana Lam. Berry Extract Inhibits Growth and Induces Apoptosis of Human Breast Cancer but Not Non-Tumorigenic Breast Cells. J. Agric. Food Chem. 2009, 57, 826–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Chu, Y.-F.; Wu, X.; Liu, R.H. Antioxidant and Antiproliferative Activities of Common Fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar] [CrossRef]
- Sun, J.; Liu, R.H. Cranberry Phytochemical Extracts Induce Cell Cycle Arrest and Apoptosis in Human MCF-7 Breast Cancer Cells. Cancer Lett. 2006, 241, 124–134. [Google Scholar] [CrossRef]
- Braakhuis, A.J.; Campion, P.; Bishop, K.S. Reducing Breast Cancer Recurrence: The Role of Dietary Polyphenolics. Nutrients 2016, 8, 547. [Google Scholar] [CrossRef]
- Abbasi, B.A.; Iqbal, J.; Ahmad, R.; Bibi, S.; Mahmood, T.; Kanwal, S.; Bashir, S.; Gul, F.; Hameed, S. Potential Phytochemicals in the Prevention and Treatment of Esophagus Cancer: A Green Therapeutic Approach. Pharmacol. Rep. 2019, 71, 644–652. [Google Scholar] [CrossRef]
- Chuang-Xin, L.; Wen-Yu, W.; Yao, C.U.I.; Xiao-Yan, L.; Yun, Z. Quercetin Enhances the Effects of 5-Fluorouracil-Mediated Growth Inhibition and Apoptosis of Esophageal Cancer Cells by Inhibiting NF-ΚB. Oncol. Lett. 2012, 4, 775–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faried, A.; Kurnia, D.; Faried, L.S.; Usman, N.; Miyazaki, T.; Kato, H.; Kuwano, H. Anticancer Effects of Gallic Acid Isolated from Indonesian Herbal Medicine, Phaleria macrocarpa (Scheff.) Boerl, on Human Cancer Cell Lines. Int. J. Oncol. 2007, 30, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Stoner, G.D.; Kresty, L.A.; Carlton, P.S.; Siglin, J.C.; Morse, M.A. Isothiocyanates and Freeze-Dried Strawberries as Inhibitors of Esophageal Cancer. Toxicol. Sci. Off. J. Soc. Toxicol. 1999, 52, 95–100. [Google Scholar] [CrossRef]
- Wang, L.-S.; Hecht, S.S.; Carmella, S.G.; Yu, N.; Larue, B.; Henry, C.; McIntyre, C.; Rocha, C.; Lechner, J.F.; Stoner, G.D. Anthocyanins in Black Raspberries Prevent Esophageal Tumors in Rats. Cancer Prev. Res. 2009, 2, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-S.; Dombkowski, A.A.; Seguin, C.; Rocha, C.; Cukovic, D.; Mukundan, A.; Henry, C.; Stoner, G.D. Mechanistic Basis for the Chemopreventive Effects of Black Raspberries at a Late Stage of Rat Esophageal Carcinogenesis. Mol. Carcinog. 2011, 50, 291–300. [Google Scholar] [CrossRef]
- Chen, T.; Yan, F.; Qian, J.; Guo, M.; Zhang, H.; Tang, X.; Chen, F.; Stoner, G.D.; Wang, X. Randomized Phase II Trial of Lyophilized Strawberries in Patients with Dysplastic Precancerous Lesions of the Esophagus. Cancer Prev. Res. 2012, 5, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.-P.; Xu, S.-Q.; Liu, L.; Shi, L.-Y.; Cai, X.-K.; Lu, W.-H.; Lu, B.; Su, Y.-H.; Li, Y.-Y. Cyclooxygenase-2 Expression in Squamous Dysplasia and Squamous Cell Carcinoma of the Esophagus. Cancer Lett. 2003, 198, 193–201. [Google Scholar] [CrossRef]
- Jin, Y.; Zhang, W.; Liu, B.; Wang, H.; Han, Z.; Wang, L.; Liu, Y.; Qu, P.; Li, M.; Ding, Z. Expression of Inducible Nitric Oxide Synthase in Human Esophageal Biopsies from Carcinoma and Precancerous Lesions. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2000, 22, 570–572. [Google Scholar]
- Izzo, J.G.; Malhotra, U.; Wu, T.-T.; Ensor, J.; Luthra, R.; Lee, J.H.; Swisher, S.G.; Liao, Z.; Chao, K.C.; Hittelman, W.N. Association of Activated Transcription Factor Nuclear Factor ΚB with Chemoradiation Resistance and Poor Outcome in Esophageal Carcinoma. J. Clin. Oncol. 2006, 24, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Boone, J.; Ten Kate, F.J.; Offerhaus, G.J.A.; van Diest, P.J.; Rinkes, I.B.; Van Hillegersberg, R. MTOR in Squamous Cell Carcinoma of the Oesophagus: A Potential Target for Molecular Therapy? J. Clin. Pathol. 2008, 61, 909–913. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, Y.; Pienta, K.J. Multiple Roles of Chemokine (CC Motif) Ligand 2 in Promoting Prostate Cancer Growth. J. Natl. Cancer Inst. 2010, 102, 522–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, A.; Santoni, M.; Mollica, V.; Fiorentino, M.; Brandi, G.; Massari, F. Microbiota and Prostate Cancer. In Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Pernigoni, N.; Zagato, E.; Calcinotto, A.; Troiani, M.; Mestre, R.P.; Calì, B.; Attanasio, G.; Troisi, J.; Minini, M.; Mosole, S. Commensal Bacteria Promote Endocrine Resistance in Prostate Cancer through Androgen Biosynthesis. Science 2021, 374, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Romero, J.R.; Chattopadhyay, N. Kaempferol and Quercetin Stimulate Granulocyte-Macrophage Colony-Stimulating Factor Secretion in Human Prostate Cancer Cells. Mol. Cell. Endocrinol. 2008, 287, 57–64. [Google Scholar] [CrossRef]
- Rini, B.I.; Small, E.J. The Potential for Prostate Cancer Immunotherapy. Crit. Rev. Oncol. Hematol. 2003, 46, 117–125. [Google Scholar] [CrossRef]
- Malik, A.; Afaq, S.; Shahid, M.; Akhtar, K.; Assiri, A. Influence of Ellagic Acid on Prostate Cancer Cell Proliferation: A Caspase–Dependent Pathway. Asian Pac. J. Trop. Med. 2011, 4, 550–555. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, M.; Fujita, K.; Hayashi, T.; Kayama, H.; Motooka, D.; Hase, H.; Jingushi, K.; Yamamichi, G.; Yumiba, S.; Tomiyama, E. Gut Microbiota–Derived Short-Chain Fatty Acids Promote Prostate Cancer Growth via IGF1 Signaling. Cancer Res. 2021, 81, 4014–4026. [Google Scholar] [CrossRef] [PubMed]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Aboonabi, A.; Meyer, R.R.; Gaiz, A.; Singh, I. Anthocyanins in Berries Exhibited Anti-Atherogenicity and Antiplatelet Activities in a Metabolic Syndrome Population. Nutr. Res. 2020, 76, 82–93. [Google Scholar] [CrossRef]
- Wu, T.; Tang, Q.; Yu, Z.; Gao, Z.; Hu, H.; Chen, W.; Zheng, X.; Yu, T. Inhibitory Effects of Sweet Cherry Anthocyanins on the Obesity Development in C57BL/6 Mice. Int. J. Food Sci. Nutr. 2014, 65, 351–359. [Google Scholar] [CrossRef]
- Takahashi, A.; Shimizu, H.; Okazaki, Y.; Sakaguchi, H.; Taira, T.; Suzuki, T.; Chiji, H. Anthocyanin-Rich Phytochemicals from Aronia Fruits Inhibit Visceral Fat Accumulation and Hyperglycemia in High-Fat Diet-Induced Dietary Obese Rats. J. Oleo Sci. 2015, 64, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Heyman, L.; Axling, U.; Blanco, N.; Sterner, O.; Holm, C.; Berger, K. Evaluation of Beneficial Metabolic Effects of Berries in High-Fat Fed C57BL/6J Mice. J. Nutr. Metab. 2014, 2014, 403041. [Google Scholar] [CrossRef] [PubMed]
- Brader, L.; Overgaard, A.; Christensen, L.P.; Jeppesen, P.B.; Hermansen, K. Polyphenol-Rich Bilberry Ameliorates Total Cholesterol and LDL-Cholesterol When Implemented in the Diet of Zucker Diabetic Fatty Rats. Rev. Diabet. Stud. RDS 2013, 10, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benn, T.; Kim, B.; Park, Y.-K.; Yang, Y.; Pham, T.X.; Ku, C.S.; Farruggia, C.; Harness, E.; Smyth, J.A.; Lee, J.-Y. Polyphenol-Rich Blackcurrant Extract Exerts Hypocholesterolaemic and Hypoglycaemic Effects in Mice Fed a Diet Containing High Fat and Cholesterol. Br. J. Nutr. 2015, 113, 1697–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eid, H.M.; Ouchfoun, M.; Brault, A.; Vallerand, D.; Musallam, L.; Arnason, J.T.; Haddad, P.S. Lingonberry (Vaccinium vitis-idaea L.) Exhibits Antidiabetic Activities in a Mouse Model of Diet-Induced Obesity. Evid. Based Complement. Altern. Med. 2014, 2014, 645812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badescu, M.; Badulescu, O.; Badescu, L.; Ciocoiu, M. Effects of Sambucus Nigra and Aronia Melanocarpa Extracts on Immune System Disorders within Diabetes Mellitus. Pharm. Biol. 2015, 53, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Chaplin, A.; Carpéné, C.; Mercader, J. Resveratrol, Metabolic Syndrome, and Gut Microbiota. Nutrients 2018, 10, 1651. [Google Scholar] [CrossRef] [Green Version]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P. Resveratrol Improves Mitochondrial Function and Protects against Metabolic Disease by Activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K. Resveratrol Improves Health and Survival of Mice on a High-Calorie Diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef]
- Lee, B.; Lee, M.; Lefevre, M.; Kim, H.-R. Anthocyanins Inhibit Lipogenesis during Adipocyte Differentiation of 3T3-L1 Preadipocytes. Plant Foods Hum. Nutr. 2014, 69, 137–141. [Google Scholar] [CrossRef]
- Noguchi, A.; Moriyama, A.; Mineo, S.; Fujisawa, Y.; Sugiyama, M.; Sakaguchi, E. Effects of Long-Term Boysenberry Juice Intake on Lipid Accumulation in Rats. J. Nutr. Sci. Vitaminol. 2011, 64, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Boath, A.S.; Grussu, D.; Stewart, D.; McDougall, G.J. Berry Polyphenols Inhibit Digestive Enzymes: A Source of Potential Health Benefits? Food Dig. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Boath, A.S.; Stewart, D.; McDougall, G.J. Berry Components Inhibit α-Glucosidase in Vitro: Synergies between Acarbose and Polyphenols from Black Currant and Rowanberry. Food Chem. 2012, 135, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Martineau, L.C.; Couture, A.; Spoor, D.; Benhaddou-Andaloussi, A.; Harris, C.; Meddah, B.; Leduc, C.; Burt, A.; Vuong, T.; Le, P.M. Anti-Diabetic Properties of the Canadian Lowbush Blueberry Vaccinium Angustifolium Ait. Phytomedicine 2006, 13, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.W.; Tomlinson, B. Effects of Bilberry Supplementation on Metabolic and Cardiovascular Disease Risk. Molecules 2020, 25, 1653. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Miao, Y.; Meng, Z.; Zhong, Y. Effects of Vaccinium Berries on Serum Lipids: A Meta-Analysis of Randomized Controlled Trials. Evid. Based Complement. Altern. Med. 2015, 2015, 790329. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Ling, W.; Guo, H.; Song, F.; Ye, Q.; Zou, T.; Li, D.; Zhang, Y.; Li, G.; Xiao, Y. Anti-Inflammatory Effect of Purified Dietary Anthocyanin in Adults with Hypercholesterolemia: A Randomized Controlled Trial. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 843–849. [Google Scholar] [CrossRef]
- Qin, Y.; Xia, M.; Ma, J.; Hao, Y.; Liu, J.; Mou, H.; Cao, L.; Ling, W. Anthocyanin Supplementation Improves Serum LDL-and HDL-Cholesterol Concentrations Associated with the Inhibition of Cholesteryl Ester Transfer Protein in Dyslipidemic Subjects. Am. J. Clin. Nutr. 2009, 90, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Du, M.; Leyva, M.J.; Sanchez, K.; Betts, N.M.; Wu, M.; Aston, C.E.; Lyons, T.J. Blueberries Decrease Cardiovascular Risk Factors in Obese Men and Women with Metabolic Syndrome. J. Nutr. 2010, 140, 1582–1587. [Google Scholar] [CrossRef] [Green Version]
- Riso, P.; Klimis-Zacas, D.; Del Bo, C.; Martini, D.; Campolo, J.; Vendrame, S.; Møller, P.; Loft, S.; De Maria, R.; Porrini, M. Effect of a Wild Blueberry (Vaccinium angustifolium) Drink Intervention on Markers of Oxidative Stress, Inflammation and Endothelial Function in Humans with Cardiovascular Risk Factors. Eur. J. Nutr. 2013, 52, 949–961. [Google Scholar] [CrossRef] [Green Version]
- Stull, A.J.; Cash, K.C.; Johnson, W.D.; Champagne, C.M.; Cefalu, W.T. Bioactives in Blueberries Improve Insulin Sensitivity in Obese, Insulin-Resistant Men and Women. J. Nutr. 2010, 140, 1764–1768. [Google Scholar] [CrossRef]
- Shidfar, F.; Heydari, I.; Hajimiresmaiel, S.J.; Hosseini, S.; Shidfar, S.; Amiri, F. The Effects of Cranberry Juice on Serum Glucose, ApoB, ApoA-I, Lp (a), and Paraoxonase-1 Activity in Type 2 Diabetic Male Patients. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2012, 17, 355. [Google Scholar]
- Novotny, J.A.; Baer, D.J.; Khoo, C.; Gebauer, S.K.; Charron, C.S. Cranberry Juice Consumption Lowers Markers of Cardiometabolic Risk, Including Blood Pressure and Circulating C-Reactive Protein, Triglyceride, and Glucose Concentrations in Adults. J. Nutr. 2015, 145, 1185–1193. [Google Scholar] [CrossRef] [Green Version]
- Chambers, B.K.; Camire, M.E. Can Cranberry Supplementation Benefit Adults with Type 2 Diabetes? Diabetes Care 2003, 26, 2695–2696. [Google Scholar] [CrossRef] [Green Version]
- Amani, R.; Moazen, S.; Shahbazian, H.; Ahmadi, K.; Jalali, M.T. Flavonoid-Rich Beverage Effects on Lipid Profile and Blood Pressure in Diabetic Patients. World J. Diabetes 2014, 5, 962. [Google Scholar] [CrossRef]
- Törrönen, R.; Kolehmainen, M.; Sarkkinen, E.; Mykkänen, H.; Niskanen, L. Postprandial Glucose, Insulin, and Free Fatty Acid Responses to Sucrose Consumed with Blackcurrants and Lingonberries in Healthy Women. Am. J. Clin. Nutr. 2012, 96, 527–533. [Google Scholar] [CrossRef]
- Moazen, S.; Amani, R.; Rad, A.H.; Shahbazian, H.; Ahmadi, K.; Jalali, M.T. Effects of Freeze-Dried Strawberry Supplementation on Metabolic Biomarkers of Atherosclerosis in Subjects with Type 2 Diabetes: A Randomized Double-Blind Controlled Trial. Ann. Nutr. Metab. 2013, 63, 256–264. [Google Scholar] [CrossRef]
- Tjelle, T.E.; Holtung, L.; Bøhn, S.K.; Aaby, K.; Thoresen, M.; Wiik, S.Å.; Paur, I.; Karlsen, A.S.; Retterstøl, K.; Iversen, P.O. Polyphenol-Rich Juices Reduce Blood Pressure Measures in a Randomised Controlled Trial in High Normal and Hypertensive Volunteers. Br. J. Nutr. 2015, 114, 1054–1063. [Google Scholar] [CrossRef] [Green Version]
- McAnulty, L.S.; Collier, S.R.; Landram, M.J.; Whittaker, D.S.; Isaacs, S.E.; Klemka, J.M.; Cheek, S.L.; Arms, J.C.; McAnulty, S.R. Six Weeks Daily Ingestion of Whole Blueberry Powder Increases Natural Killer Cell Counts and Reduces Arterial Stiffness in Sedentary Males and Females. Nutr. Res. 2014, 34, 577–584. [Google Scholar] [CrossRef]
- Pantuso, T. Blueberry Consumption Improves Blood Pressure and Arterial Stiffness in Postmenopausal Women with Hypertension. Available online: https://0-www-proquest-com.brum.beds.ac.uk/docview/1986673835?pq-origsite=gscholar&fromopenview=true (accessed on 3 March 2021).
- Johnson, S.A.; Figueroa, A.; Navaei, N.; Wong, A.; Kalfon, R.; Ormsbee, L.T.; Feresin, R.G.; Elam, M.L.; Hooshmand, S.; Payton, M.E. Daily Blueberry Consumption Improves Blood Pressure and Arterial Stiffness in Postmenopausal Women with Pre-and Stage 1-Hypertension: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. J. Acad. Nutr. Diet. 2015, 115, 369–377. [Google Scholar] [CrossRef]






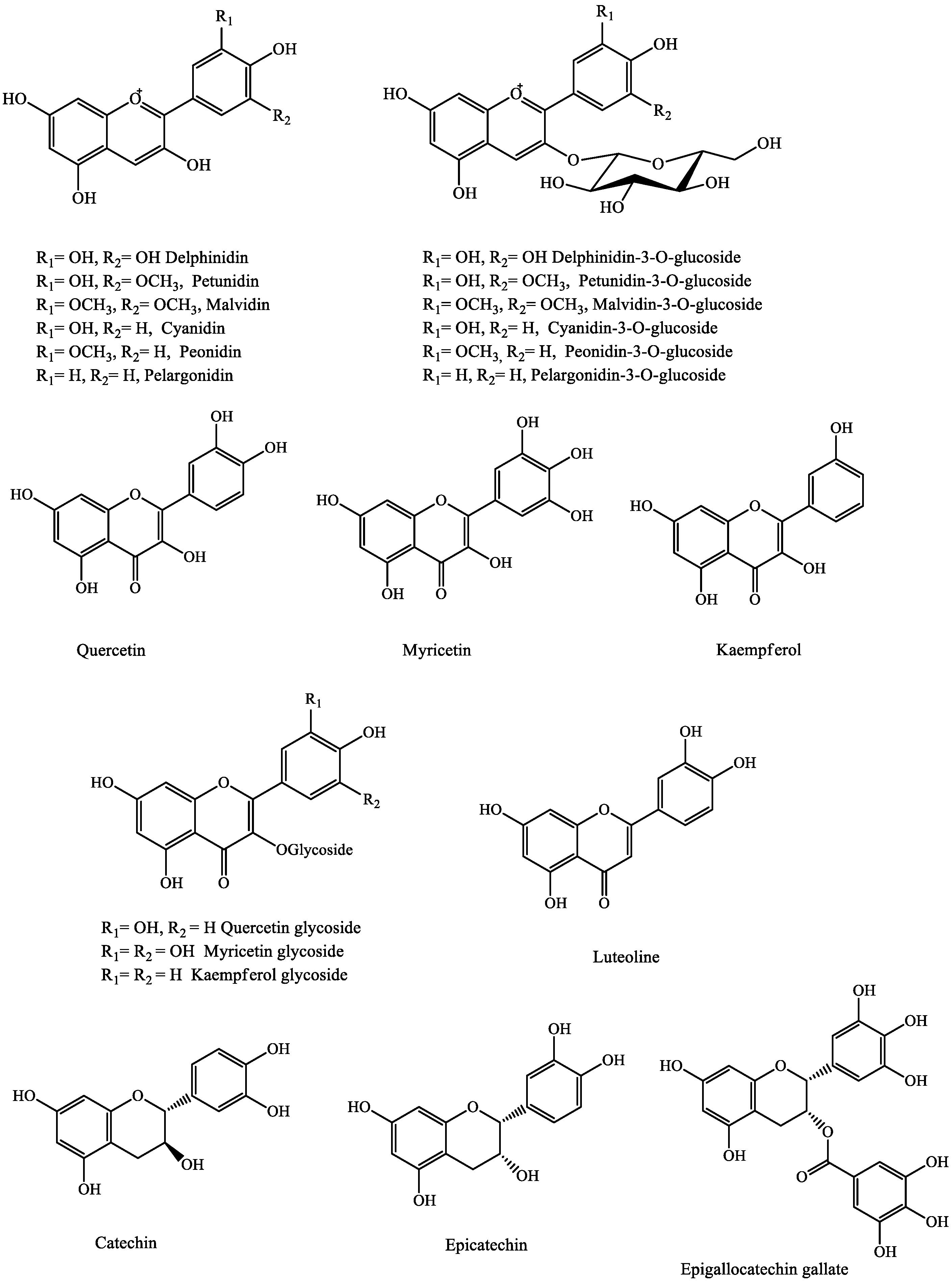{kind=link}
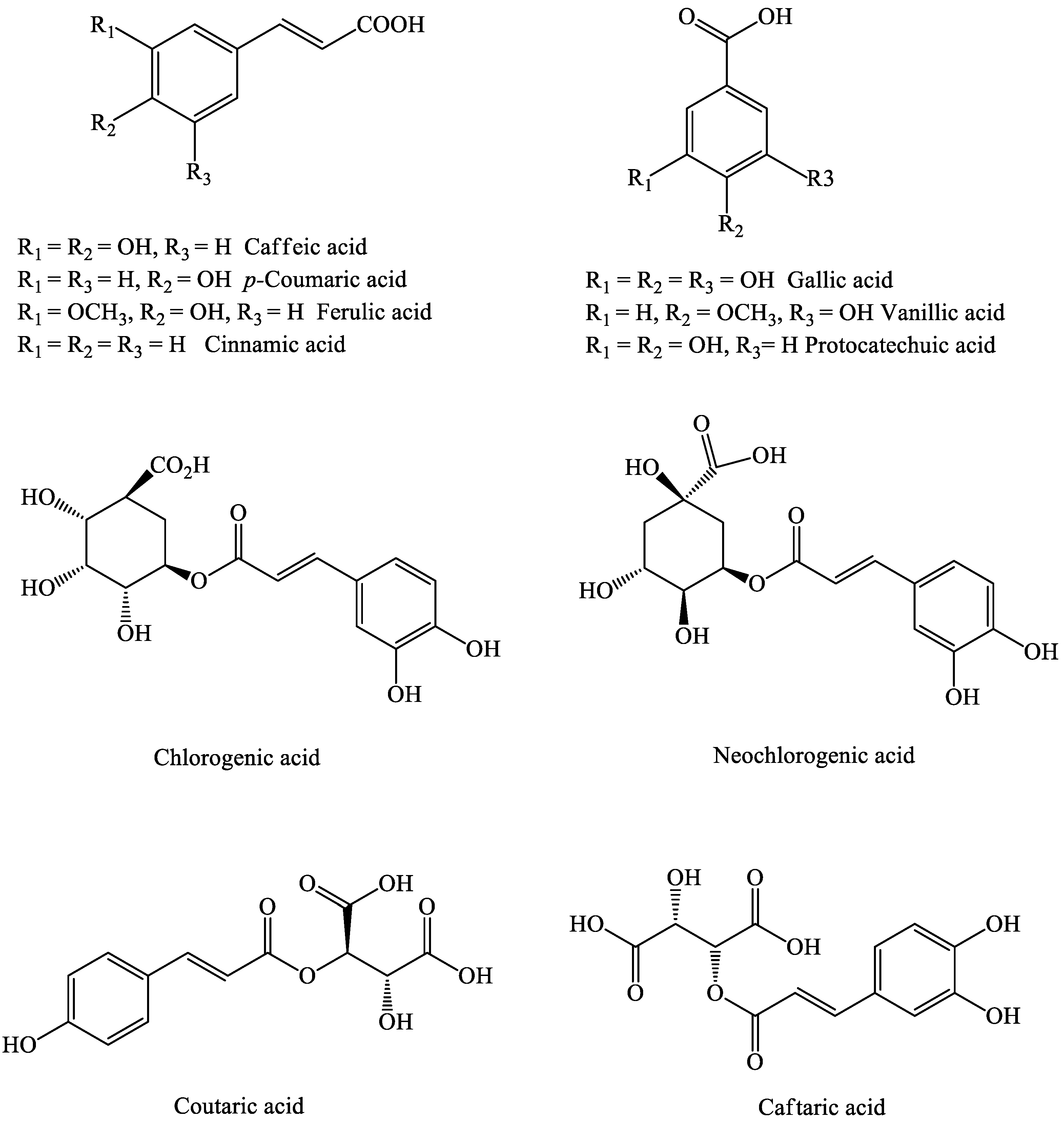{kind=link}
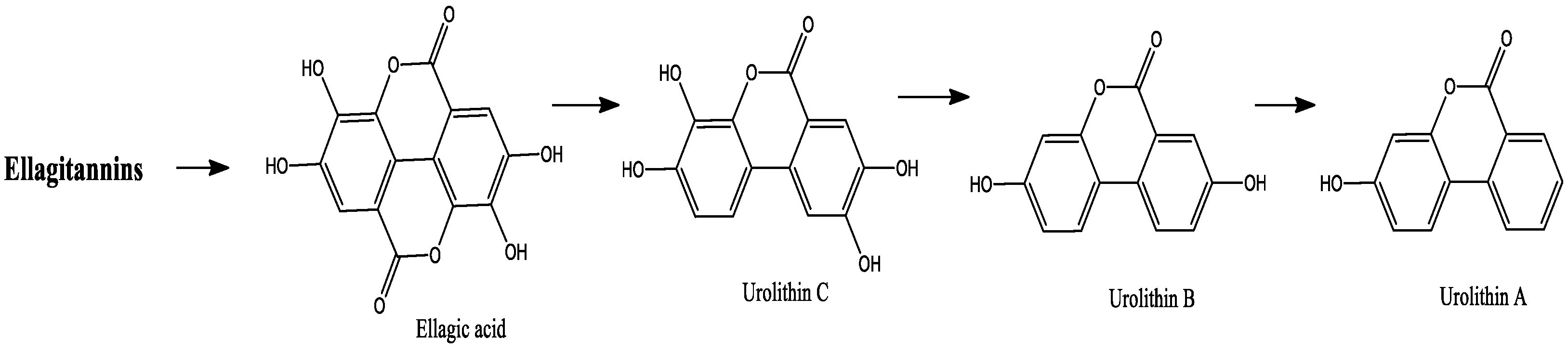{kind=link}
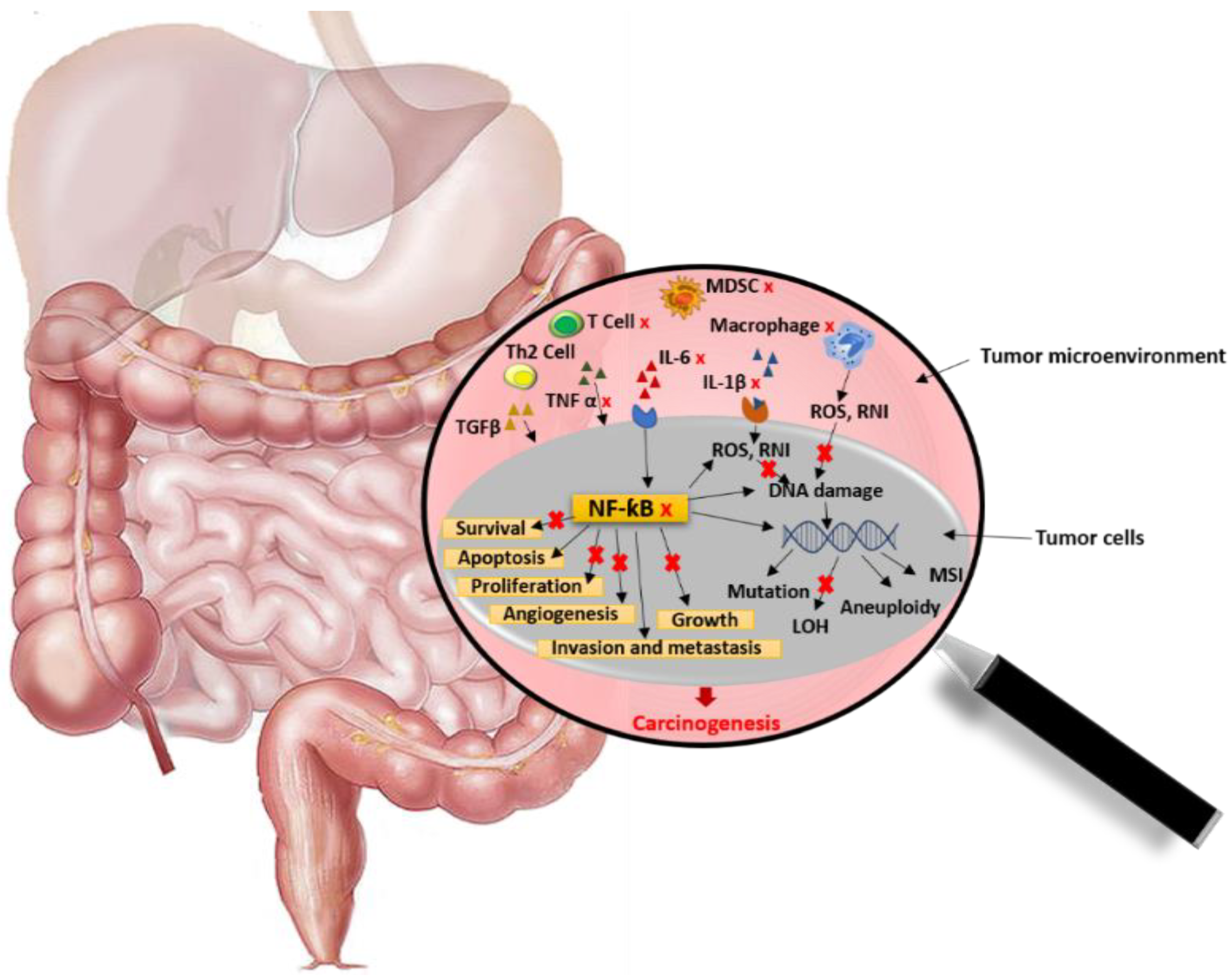{kind=link}
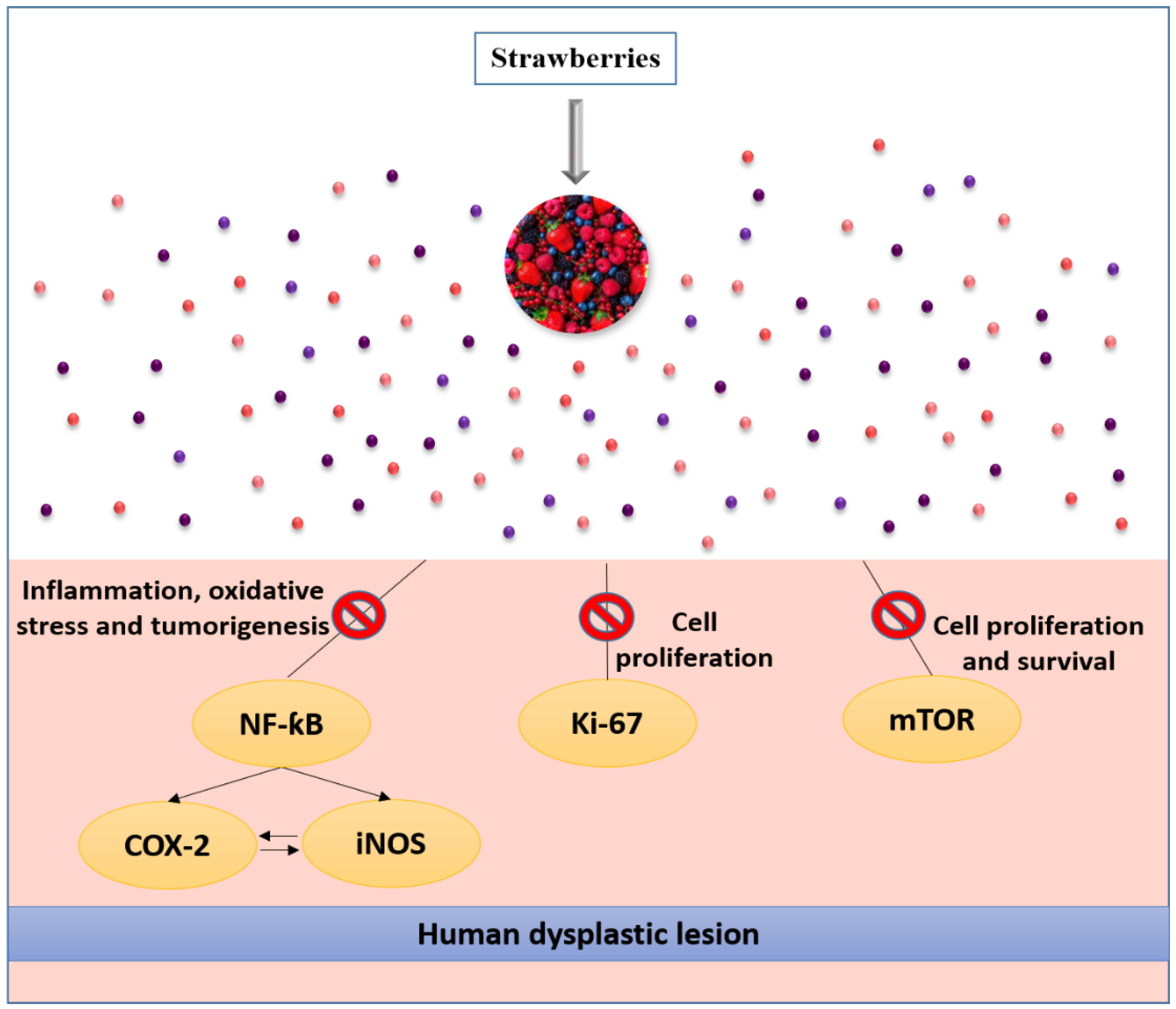{kind=link}
| Plants | Characterization Methods | Major Polyphenols | References |
|---|---|---|---|
| Vitis vinifera cultivar: Mandilaria | HPLC–DAD | (+)-Catechin, (−)-epicatechin, procyanidin B2, and procyanidin B3 | [88] |
| Vitis vinifera cultivar: Voidomatis | HPLC–DAD | (+)-Catechin, (−)-epicatechin, procyanidin B2, and procyanidin B3 | |
| Vitis vinifera cultivar: Asyrtiko | HPLC–DAD | (+)-Catechin, (−)-epicatechin, procyanidin B3, and epicatechin gallate | |
| Vitis vinifera cultivar: Aidani | HPLC–DAD | (+)-Catechin, (−)-epicatechin, procyanidin B2, and procyanidin B3 | |
| Amelanchier alnifolia Nutt. cultivar: Nelson | HPLC-ESI-MS/MS | Cyanidin-3-galactoside and chlorogenic acid | [89] |
| Myrtus communis from Canari | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | Myricetin, myricetin-3-O-arabinoside, and myricetin-3-O-galactoside | [90] |
| Myrtus communis from Bastia | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | Myricetin, myricetin-3-O-arabinoside, and myricetin-3-O-galactoside | |
| Myrtus communis from Agriate | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | Myricetin, myricetin-3-O-arabinoside, and myricetin-3-O-galactoside | |
| Myrtus communis from Corte | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | Myricetin, myricetin-3-O-arabinoside, and (−)epigallocatechin | |
| Myrtus communis from Ajaccio | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | Myricetin, quercetin-3-O-rhamnoside, and (−)epigallocatechin | |
| Myrtus communis from Morta | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | Myricetin, quercetin-3-O-rhamnoside, and (−)epigallocatechin | |
| Myrtus communis from Bonifacio | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | Myricetin, quercetin-3-O-glucoside, and myricetin-3-O-arabinoside | |
| Myrtus communis from Abbazzia | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | Myricetin-3-O-arabinoside, myricetin, and (−)epigallocatechin | |
| Myrtus communis from Travo | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | (−)Epigallocatechin, quercetin-3-O-rhamnoside, and myricetin | |
| Myrtus communis from Aleria | HPLC–DAD HS–SPME, GC and GC/MS LC–MS–MS | Myricetin, myricetin-3-O-arabinoside, and (−)epigallocatechin | |
| Myrtus communis L. | HPLC–DAD | Myricetin-3-O-arabinoside and myricetin-3-O-galactoside | [91] |
| Hippophaërhamnoides L. | HPLC | Catechin, epicatechin, gallic acid, p-coumaric acid, caffeic acid, ferulic acid, rutin (quercetin 3-rutinoside), and quercitrin (quercetin 3-rhamnoside) | [92] |
| Wild bilberry and blackberry cv. Čačanskabestrnaand cv. Thornfree | HPLC | Protocatechuic acid and gallic acid | [93] |
| Vitis vinifera cv. Italia (white) | LC/ESI-MS | Procyanidin B1, quercetin glucoronide, and epicatechin | [94] |
| Vitis vinifera cv. Michele Palieri (red) | LC/ESI-MS | Peonidin-3-O-glucoside, procyanidin B1, quercetin glucoronide, cis-resveratrol, and epicatechin | |
| Vitis vinifera cv. Red Globe (red) | LC/ESI-MS | Peonidin-3-O-glucoside, procyanidin B1, quercetin glucoside, cis-resveratrol, and epicatechin | |
| Vaccinium spp. | CIELAB and HPLC-DAD | Delphinidin-3- O-galactoside, malvidin-3-O-galactoside, malvidin-3-O-arabinoside, and delphinidin-3-O-arabinoside | [30] |
| Vitis vinifera cv. Aglianico | HPLC coupled with LC-ESI/MS/MS | Malvidin-3-(6-O-coumaroyl)-glucoside, malvidin-3-O-glucoside, trans-resveratrol, and petunidin-3-O-glucoside | [95] |
| Lonicera caerulea L. | UPLC-ESI-MS | Cyanidin-3-O-glucoside, quercetin-3-O-rutinoside, and catechin | [96] |
| Vaccinium angustifolium | HPLC and MS | Cyanidin-3-galactoside, delphinidin 3-glucoside, malvidin-3-galactoside, and petunidin-3-galactoside | [97] |
| Aristotelia chilensis (Molina) Stuntz | HPLC | Delphinidin-3-glucoside, delphinidin-3,5-diglucoside, delphinidin-3-sambubioside, and ellagic acid | [24] |
| Vaccinium myrtillus | HPLC | Gallic acid, vanillic acid, ferulic acid, and quercetin | [46] |
| Vaccinium vitis-idaea | HPLC | Vanillic acid, ferulic acid, caffeic acid, and p-coumaric acid | |
| Rubus chamaemorus | HPLC | Gallic acid, caffeic acid, ferulic acid, and p-coumaric acid | |
| Hippophae rhamnoides L. | HPLC | Quercetin, gallic acid, p-coumaric acid, and isorhamnetin | |
| Rubus laciniatus and Rubus sp. Hyb | HPLC | Epicatechin, ellagic acid, and rutin | [98] |
| 16 strawberry cultivars | LC/DAD/ESI-MS | Ellagic acid, feruloyl, caffeoyl, coumaroyl hexose, quercetin, kaempferol, pelargonidin-3-O-glucoside, (epi)catechin, and (epi)afzelechin | [32] |
| Ribes nigrum L. | HPLC | Delphinidin-3-O-glucoside, delphinidin-3-O-rutinoside, cyanidin-3-O-glucoside, and cyanidin-3-O-rutinoside | [99] |
| Vitis vinifera L. cv. Shiraz | HPLC | (+)-Catechin, (–)-epicatechin, (–)-epicatechin-3-O-gallate | [100] |
| Lonicera caerulea L. | HPLC-DAD-MS | Cyanidin-3-glucoside | [101] |
| Amelanchier alnifolia Nutt. | HPLC-DAD and HPLC- ESI/MS | Chlorogenic acid, cinnamic acid deriv 2, cyanidin 3-galactoside, quercetin 3-galactoside, catechin deriv 1, and procyanidin deriv 2 | [102] |
| Prunus cerasus cv. Marasca | HPLC | (–)-Epicatechin, quercetin 3-rutinoside, and chlorogenic acid | [31] |
| Prunus cerasus cv. Oblačinska | HPLC | (–)-Epicatechin, quercetin 3-rutinoside, and chlorogenic acid | |
| Fragaria× ananassa cv. Maya | HPLC | (–)-Epicatechin, kaempferol derivative, chlorogenic acid, and ellagic acid | |
| Rubus idaeus cv. Willamette | HPLC | (–)-Epicatechin, quercetin 3-rutinoside, chlorogenic acid, and ellagic acid | |
| Vaccinium myrtilus | HPLC | (–)-Epicatechin, kaempferol derivative, chlorogenic acid, and ellagic acid | |
| Vitis labrusca, Vitis vinifera, hybrid of Vitis labrusca and Vitis vinifera, hybrid of Vitis vinifera and Vitis amurensis, and hybrid of Vitis thunbergii and Vitis vinifera | HPLC and HPLC-MS | Delphinidin, cyanidin, petunidin, peonidin, malvidin, caftaric acid, coutaric acid, procyanindin B1, epicatechin, rutin, myricetin 3-O-glucoside, quercetin 3-O-glucuronide, and quercetin 3-O- glucoside | [48] |
| Ribes nigrum cv. Öjebyn, Ribes nigrum cv. Vertti, Ribes pallidum cv. Red Dutch, and Ribes × pallidum cv. White Dutch | LC-DAD and LC-MS | Caffeoylglucose, p-coumaroylglucose and hexose, feruloylhexose, myricetin 3-O-rutinoside, myricetin 3-O-glucoside, rutin, quercetin 3-O-glucoside, kaempferol hexoside−malonate, delphinidin 3-O-rutinoside, and cyanidin 3-O-rutinoside | [34] |
| Strawberry | HPLC-DAD | Pelargonidin 3-glucoside and pelargonidin 3-rutinoside | [25] |
| Blackberry | HPLC-DAD | Delphinidin 3-glucoside and cyanidin 3-glucoside | |
| Blueberry | HPLC-DAD | Petunidin 3-glucoside and delphinidin 3-glucoside | |
| Raspberry | HPLC-DAD | Cyanidin 3-glucoside and petunidin 3-glucoside | |
| Gaultheria phillyreifolia and Gaultheria poeppigii | HPLC-DAD-ESI-MS | Cyanidin galactoside, cyanidin arabinoside, delphinidin galactoside, delphinidin arabinoside, quercetin-3-arabinoside, quercetin-3-rutinoside, quercetin-3-rhamnoside and 3-caffeoylquinic acid, monotropein-10-trans-coumarate, monotropein-10-trans-cinnamate, and 6α-hydroxy-dihydromonotropein-10-trans-cinnamate | [35] |
| Blueberry, bilberry, cranberry, lingonberry, eastern shadbush, Japanese wineberry, black mulberry, chokeberry, red, black and white currants, jostaberry, red and white gooseberry, goji berry, and rowan; and wild and cultivated species of blackberry, raspberry, strawberry, and elderberry | HPLC–MS | Glycosides of quercetin, myricetin, kaempferol, isorhamnetin, syringetin, and laricitrin | [33] |
| Myrtus communis L. | HPLC-ESI-MS and HPLC-UV/VIS | Delphinidin-3-O-glucoside, cyanidin-3-O-glucoside, petunidin-3-O-glucoside, peonidin-3-O-glucoside, malvidin-3-O-glucoside, delphinidin-3-O-arabinoside, petunidin-3-O-arabinoside, malvidin-3-O-arabinoside, myricetin-3-O-galactoside, myricetin-3-O-rhamnoside, myricetin-3-O-arabinoside, quercetin-3-O-glucoside, quercetin-3-O-rhamnoside, and myricetin | [103] |
| White grapes cv. Rebula | HPLC-DAD | Trans-caftaric, trans-coutaric, cis-coutaric, and trans-fertaric acid | [104] |
| Vitis vinifera (Primitivo) | HPLC-PDA-ESI-MS | Catechin, petunidin-3-O-glucoside, peonidin 3-O-glucoside, and malvidin 3-O-glucoside | [105] |
| Vitis vinifera (Negroamaro) | HPLC-PDA-ESI-MS | Caftaric acid, delphinidin 3-O-glucoside, petunidin-3-O-glucoside, and malvidin 3-O-glucoside | |
| Vitis vinifera (Susumaniello) | HPLC-PDA-ESI-MS | Caftaricacid, catechin, delphinidin 3-O-glucoside, cyanidin 3-O-glucoside, petunidin-3-O-glucoside, peonidin 3-O-glucoside, trans-piceid, and malvidin 3-O-glucoside | |
| Vitis vinifera (Uva di Troia) | HPLC-PDA-ESI-MS | Catechin, delphinidin 3-O-glucoside, cyanidin 3-O-glucoside, petunidin-3-O-glucoside, peonidin 3-O-glucoside, and malvidin 3-O-glucoside | |
| Vitis vinifera (Malvasia Nera) | HPLC-PDA-ESI-MS | Catechin, epicatechin, petunidin-3-O-glucoside, peonidin 3-O-glucoside, and malvidin 3-O-glucoside | |
| Vitis vinifera (Aglianico) | HPLC-PDA-ESI-MS | Catechin, epicatechin, malvidin 3-O-glucoside, petunidin-3-O-glucoside, delphinidin 3-O-glucoside, and peonidin 3-O-glucoside | |
| Vitis vinifera (Cesanese) | HPLC-PDA-ESI-MS | Delphinidin 3-O-glucoside, cyanidin 3-O-glucoside, petunidin-3-O-glucoside, peonidin 3-O-glucoside, and malvidin 3-O-glucoside | |
| Vitis vinifera (Merlot) | HPLC-PDA-ESI-MS | Caftaric acid, epicatechin, delphinidin 3-O-glucoside, malvidin 3-O-glucoside, and catechin | |
| Red Globe grape (Vitis vinifera L.) | HPLC-ESI-QTOF-MS | Catechin, pelargonidin, and cyanidin-3,5-O-diglucoside | [26] |
| Raspberry (Rubus idaeus L.) | HPLC-ESI-QTOF-MS | Epicatechin, cyanidin-3,5-O-diglucoside, and pelargonidin | |
| Blackberry (Rubus spp.) | HPLC-ESI-QTOF-MS | Epicatechin, cyanidin-3-O-β-glucoside, and cyanidin-3,5-O-diglucoside | |
| Black currant (Ribes nigrum) cultivar ‘Titania’ | HPLC-DAD and LC-ESI/MS | Chlorogenic acid, quercetin-3-O-galactoside, and quercetin-3-O-glucosyl-6’’-acetate | [106] |
| Raspberry (Rubus ideaus) cultivar ‘Polka’ | HPLC-DAD and LC-ESI/MS | Chlorogenic acid, quercetin-3-O-rutinoside, and quercetin-3-O-glucuronide | |
| Honeysuckle (Lonicera kamtschatica) cultivar ‘Zielona’ | HPLC-DAD and LC-ESI/MS | Chlorogenic acid, quercetin-3-O-glucoside, and 3,5-di-O-caffeoylquinic acid | |
| Bilberry (Vaccinium myrtillus L.) | HPLC-DAD and LC-ESI/MS | Chlorogenic acid, 3-O-p-coumaroylquinic acid, and quercetin-3-O-glucuronide | |
| Strawberry (Fragaria × ananassa Duch.) cultivar ‘Elkat’ | HPLC-DAD and LC-ESI/MS | Quercetin-3-O-rutinoside, quercetin-3-O-glucuronide, and kaempferol-3-O-rutinoside | |
| Vaccinium floribundum Kunth | HPLC-UV/DAD HPLC-ESI-MS and MS | Chlorogenic acid, quercetin-3-O-galactoside, quercetin-3-O-arabinofuranoside, quercetin-3-O-rhamnoside, cyanidin-3-O-galactoside, cyanidin-3-O-arabinoside, and delphinidin-3-O-arabinoside | [29] |
| Vaccinium myrtillus L. | HPLC-UV/DAD HPLC-ESI-MS and MS | Chlorogenic acid, quercetin-3-O-galactoside, quercetin-3-O-glucuronide, delphinidin-3-O-galactoside, delphinidin-3-O-glucoside, cyanidin-3-O-galactoside, and petunidin-3-O-glucoside | |
| Wild blackberry (Rubus fruticosus) | HPLC | Gallic acid, t-caftaric acid, caffeic acid, and sirginic acid | [49] |
| Blackthorn (Prunus spinosa L.) | HPLC | Gallic acid, t-caftaric acid, t-coutaric acid, caffeic acid, and sirginic acid | |
| European cornel (Cornus mas) | HPLC | Gallic acid, t-caftaric acid, caffeic acid, p-coumaric acid, and sirginic acid | |
| Lycium barbarum L. | UHPLC-ESI-QTOF-MS | Cyanidin, ferulic acid, catechin, and luteolin | [107] |
| Calafate (Berberis microphylla) | HPLC-DAD-MS/MS | Delphinidin-3-glucoside, cyanidin-3-glucoside, petunidin-3-glucoside, peonidin-3-glucoside, malvidin-3-glucoside, delphinidin-3-rutinoside, cyanidin-3-rutinoside, petunidin-3-rutinoside, peonidin-3-rutinoside, malvidin-3-rutinoside, delphinidin-3,5-dihexoside, cyanidin-3,5-dihexoside, petunidin-3,5-dihexoside, peonidin-3,5-dihexoside, malvidin-3,5-dihexoside, and petunidin-3-rutinoside-5-glucoside | [108] |
| Maqui (Aristotelia chilensis) | HPLC-DAD-MS/MS | Delphinidin-3-glucoside, cyanidin-3-glucoside, delphinidin-3-sambubioside-5-glucoside, cyanidin-3-sambubioside-5-glucoside, delphinidin-3-sambubioside, cyanidin-3-sambubioside, delphinidin-3,5-diglucoside, and cyanidin-3,5-diglucoside | |
| Murtilla (Ugni molinae) | HPLC-DAD-MS/MS | Cyanidin-3-glucoside and peonidin-3-glucoside | |
| Berberis microphylla | HPLC-DAD-ESI-MS/MS | Delphinidin-3-glucoside, petunidin-3-glucoside, malvidin-3-glucoside, and petunidin-3.5-dihexoside | [109] |
| Berberis ilicifolia | HPLC-DAD-ESI-MS/MS | Delphinidin-3-glucoside, petunidin-3-glucoside, malvidin-3-glucoside, and cyanidin-3-glucoside | |
| Berberis empetrifolia | HPLC-DAD-ESI-MS/MS | Delphinidin-3-glucoside, petunidin-3-glucoside, malvidin-3-glucoside, and petunidin-3-rutinoside | |
| Ribes magellanicum | HPLC-DAD-ESI-MS/MS | Delphinidin-3-glucoside, delphinidin-3-rutinoside, cyanidin-3-glucoside, and cyanidin-3-rutinoside | |
| Ribes cucullatum | HPLC-DAD-ESI-MS/MS | Delphinidin-3-glucoside, delphinidin-3-rutinoside, cyanidin-3-glucoside, and cyanidin-3-rutinoside | |
| Gaultheria mucronate | HPLC-DAD-ESI-MS/MS | Cyanidin-pentoside and delphinidin-pentoside + cyanidin-3-galactoside | |
| Gaultheria antarctica | HPLC-DAD-ESI-MS/MS | Cyanidin-pentoside, delphinidin-3-galactoside, and cyanidin-3-galactoside + delphinidin-pentoside | |
| Rubus geoides | HPLC-DAD-ESI-MS/MS | Cyanidin-3-sambubioside | |
| Myrteola nummularia | HPLC-DAD-ESI-MS/MS | Delphinidin-3-glucoside and cyanidin-3-glucoside | |
| Fuchsia magellanica | HPLC-DAD-ESI-MS/MS | Cyanidin-3-glucoside | |
| Greek grape (Vitis vinifera) samples | HPLC | (+)-Catechin, (–)-epicatechin, trans-resveratrol, quercetin, and quercetin glycosides | [110] |
| Luma apiculata | HPLC-DAD-ESI/MS | Delphinidin 3-O-hexose, epigallocatechin gallate, cyanidin-3-O-glucose, petunidin 3-O-glucose, peonidin 3-O-glucose, malvidin 3-O-glucose, isoquercitrin, and quercitrin | [111] |
| Luma chequén | HPLC-DAD-ESI/MS | Procyanidin B1, delphinidin 3-O-hexose, cyanidin-3-O-glucose, petunidin 3-O-glucose, peonidin 3-O-glucose, malvidin 3-O-glucose, isoquercitrin, and syringetin-3-O-glucose | |
| Vitis vinifera L. ssp. Sativa | HPLC-DAD | Chlorogenic acid, caffeic acid, gallic acid, vanillin, feluric acid, ellagic acid, and E-resveratrol | [47] |
| Grape varieties (cabernet sauvignon, syrah, malbec, and merlot) | HPLC and UV | Quercetin and trans-resveratrol | [112] |
| Black elderberry (Sambucus nigra L.) | HPLC-DAD | Anthocyanins: cyanidin 3-sambubioside-5-glucoside, cyanidin 3,5-diglucoside, cyanidin 3-sambubioside, cyanidin 3-glucoside, and cyanidin 3-rutinoside Quercetins:quercetin, quercetin 3-rutinoside, and quercetin 3-glucoside | [28] |
| Vaccinium uliginosum berry | HPLC-DAD and HPLC-ESI-MS | Delphinidin 3-O-glucoside, malvidin 3-O-glucoside, myricetin 3-O-hexoside, and quercetin 3-O-galactoside | [113] |
| Lonicera caerulea berry | HPLC–DAD–EIS–MS | Cyanidin-3,5-dihexoside, cyanidin-3-hexoside-catechin, dimer of cyanidin-hexoside, peonidin-3,5-dihexoside, cyanidin- 3-glucoside, cyanidin-3-rutinoside, peonidin- 3-glucoside, peonidin-3-rutinoside, and dimer of cyanidin-3-hexoside | [61] |
| Black raspberry and marionberry | HPLC-DAD-ESI/MS/MS | Delphinidin, cyanidin, pelargonidin, petunidin, peonidin, and malvidin, | [23] |
| Blueberries (Vaccinium uliginosum L.) | HPLC-FT-ICR MS/MS | Cyanidin-3-O-glucoside, cyanidin-3-O-rutinoside, catechin, and myricetin 3-O-hexose | [114] |
| Bilberries (Vaccinium myrtillus L.) | HPLC-FT-ICR MS/MS | Cyanidin-3-O-rutinoside, catechin, quercetin-3-O-galactoside, and quercetin-3-O-arabinoside | |
| Mulberries (Morus alba L.) | HPLC-FT-ICR MS/MS | Malvidin-3-O-glucoside, myricetin 3-O-hexose, quercetin-3-O-rutinoside, and chlorogenic acid | |
| Cranberries (Vaccinium oxycoccos L.) | HPLC-FT-ICR MS/MS | Cyanidin-3-O-rutinoside, catechin, quercetin-3-O-rutinoside, and chlorogenic acid | |
| Solanum scabrum | HPLC/UV–visible/MS | Chlorogenic acid, neochlorogenic acid, quercetin, isorhamnetin, quercetin-rhamnosyl-hexoside, tigogenin, diosgenin, petunidin, delphinidin, malvidin, and petunidin-3-(p-coumaroyl-rutinoside)-5-O-glucoside | [115] |
| Black mulberry (Morus nigra L.) | HPLC-DAD-ESI HRMS | Quercetin-dirhamnosyl-hexoside, quercetin-rhamnosyl-dihexoside, and procyanidin trimer 1 | [116] |
| Cornelian cherry (Cornus mas L.) | HPLC-DAD-ESI HRMS | Rhamnetin-rhamnosyl-hexoside, kaempferol-dirhamnosyl-hexoside, and procyanidin trimer (2 and 3) | |
| Elderberry (Sambucus nigra L.) | HPLC-DAD-ESI HRMS | Rhamnetin-rhamnosyl-hexoside, myricetin-rhamnosyl-hexoside, quercetin-acetyl-pentosyl-hexoside, kaempferol-rhamnosyl-dihexoside, quercetin-rhamnosyl-dihexoside, and procyanidin trimer 1 | |
| Hawthorn (Crataegus monogyna L.) | HPLC-DAD-ESI HRMS | Rhamnetin-rhamnosyl-hexoside, quercetin-dihexoside, and procyanidin trimer (2 and 3) | |
| Lingonberry (Vaccinium vitis-idaea L.) | HPLC-DAD-ESI HRMS | Rhamnetin-rhamnosyl-hexoside and quercetin-dirhamnosyl-hexoside | |
| Rose hip (Rosa canina L.) | HPLC-DAD-ESI HRMS | Quercetin-galloyl-hexoside and procyanidin trimer 1 | |
| Rubus fruticosus, Ribes nigrum, Vaccinium corymbosum, Lyciumbarbarum L., Rubus idaeus, Ribes rubrum, Ribes grossularia L., Ribes pallidum, and Ribes grossularia L. | HPLC-DAD-UV-ESI HRMS | Ferulic acid, coumaric acid, cyanidin, and delphinidin Myricetin, kaempferol, catechin, quercetin, chlorogenic acid, malvidin, and petunidin | [38] |
| Berries Polyphenols | Type of Study | Dosage | Gut Microbiota Modulation | Ref. |
|---|---|---|---|---|
| Açai | In vitro | 1 g of dry berries reduced in powder (24 h) | Inhibited the growth of Clostridium histolyticum, Bacteroides-Prevotella spp. Bifidobacterium spp., and Lactobacillus/Enterococcus spp.; Clostridium coccoides-Eubacterium rectale were not affected | [201] |
| Bilberry, black currant, buckthorn berry cloudberry, lingonberry, raspberry, and strawberry | In vitro | 2 and 10 mg/mL of dry berries reduced in powder (24 h) | Inhibited the growth of Salmonella enterica and Staphylococcus aureus Lactobacillus rhamnosus and Listeria monocytogenes were not affected | [10] |
| Blackberry, black raspberry, black currant, maqui berry, Concord grape, and blueberry | In vivo (mice; high-fat diet) | 400 μg berry anthocyanins (12 weeks) | Reduced gut luminal oxygenation and promoted abundance of obligate anaerobic bacteria from Bacteroidetes and Actinobacteria phyla at these sites | [187] |
| Black raspberry | In vivo (healthy mice) | 10% of dry berry powder (6 weeks) | Increased the abundance of Barnesiella Reduced Clostridium and Lactobacillus | [140] |
| Blackberry | In vivo (rats; normal diet, and high-fat diet) | 25 mg/kg/day (17 weeks) | Normal diet: promoted abundance of Pseudoflavonifractor and Oscillobacter High-fat diet: promoted abundance of Oscillobacter and reduced Rumminococcus | [208] |
| Black currant | In vivo (healthy rats) | 2 mL (extracts; 4 weeks) | Promoted abundance of Bifidobacteria and lactobacilli Reduced Bacteroides and clostridia | [196] |
| Black currant | Clinical study (healthy volunteers) | 672 mg/day of dry berry powder (2 weeks) | Promoted abundance of Lactobacillus spp. and Bifidobacterium spp. Reduced Clostriudium spp. and Bacterioides spp. | [198] |
| Black raspberry | In vivo (healthy rats) | 5% of dry berry powder (6 weeks) | Promoted abundance of Akkermansia, Desulfovibrio, and Anaerostipes | [209] |
| Blueberry polyphenols | Altered gut bacteria composition, such as the abundance of Proteobacteria, Deferribacteres, Actinobacteria, Bifidobacterium, Desulfovibrio, Adlercreutzia, Helicobacter, Flexispira, and Prevotella | [184] | ||
| Blueberry | In vivo (mice; inflammatory bowel disease model) | 10% of dry berry powder (21 weeks) | Reduced Clostridium perfringens, Enterococcus spp., Lactobacillus spp., and Escherichia coli | [210] |
| Blueberry | In vivo (healthy rats) | 8% of dry berry powder (21 weeks) | Reduced Lactobacillus and Enterococcus Promoted abundance of bacteria from the orders Actinomycetales, Bifidobacteriales, and Coriobacteriales | [211] |
| Blueberry | In vitro | 10 and 25% (berry extracts) for 5 days | Promoted abundance of Lactobacillus rhamnosus and Bifidobacterium breve | [212] |
| Blueberry | In vitro | 5, 10, and 25% (berry extracts) for 48 h | Promoted abundance of Lactobacilli and Bifidobacteria populations | |
| Blueberry | Animal study (healthy rats) | 4 mL berry extracts per kg per day for 6 days | Promoted abundance of Lactobacilli and Bifidobacteria populations | [212] |
| Blueberry | Human study (healthy volunteers) | 250 mL drink (10% dry berry powder in water) for 6 weeks | Promoted abundance of Bifidobacterium spp. and Lactobacillus acidophilus The consumption of blueberry led to an increase in Bifidobacterium spp. and Lactobacillus acidophilus | [197] |
| Blueberry | Human study (healthy volunteers) | 250 mL drink (10% dry berry powder in water) for 6 weeks | Promoted abundance of Bifidobacterium longum subsp. infantis | [213] |
| Cranberry | In vitro | (−) Salmonella enterica, Staphylococcus aureus, Lactobacillus rhamnosus, and (−) Listeria monocytogenes | Inhibited the growth of Salmonella enterica, Staphylococcus aureus, and Listeria monocytogenes Lactobacillus rhamnosus was not affected | [10] |
| Cranberry and grapeseeds | In vitro | 500 mg/L of the extracts (48 h) | Inhibited the growth of Bacteroides, Prevotella, Blautiacoccoides-Eubacterium rectale, Lactobacillus, Bifidobacterium, Enterobacteriaceae, Clostridium leptum, and Ruminococcus | [200] |
| Cranberry extract polyphenols | Increased the abundance of Akkermansia species | [190] | ||
| Cranberry | Animal study (mice; high-fat diet) | 200 mg/kg (berry extracts) for 9 weeks | Pyrosequencing (+) Akkermansia | [190] |
| Goji berry | Animal study (mice; colitis model) | 1% (dry berry powder) for 10 weeks | Bifidobacteria, Clostridium leptum, Fecalibacterium prazusnitzii | [214] |
| Grape polyphenols | Increased the beneficial anaerobic gut bacteria, such as Akkermansia muciniphila | [186] | ||
| Grape polyphenols | Animal study (mice; high-fat diet) | Significantly reduced the Firmicutes/Bacteroidetes ratio Promoted the growth of A. muciniphila | [192] | |
| Juçara | In vitro | 1% (dry berry powder) for 24 h | Promoted abundance of Bifidobacterium, Eubacterium rectale-Clostridium coccoide, and Bacteroides spp. | [215] |
| Juçara | Animal study (rats; high-fat diet) | 0.5 and 0.25% (dry berry powder) for 7 days | Promoted abundance of Bifidobacterium | [216] |
| Lingonberry | Animal study (mice; high-fat diet) | 44% (dry berry powder) for 8 weeks | Promoted abundance of Bacteroides, Parabacteroides, Clostridium 106, (−) Mucispirillum, and Oscillospira | [195] |
| Lonicera cerula L. | Animal study (mice; high-fat diet) | 1% (dry berry powder for 45 days) | Promoted abundance of Bacteroides, Parabacteroides, two genera from the order Bacteroidales (−) Staphylococcus, Lactobacillus, Ruminococcus, and Oscillospira | [191] |
| Lycium ruthenicum | In vitro (batch-culture fermentation) | 1 g/L anthocyanins for 24 h | Promoted abundance of Bifidobacterium, Allisonella (−) Prevotella, Dialister, Megamonas, and Clostridium | [202] |
| Plinia jaboticaba | Animal study (healthy rats) | Juice for 2 and 7 weeks | Promoted abundance of Lactobacillus, Bifidobacterium, and Enterobacteriaceae | [217] |
| Procyanidin supplement | Significantly increased the β-diversity of intestinal microbiota and the Bacteroidetes abundance Reduced the Firmicutes/Bacteroidetes ratio Decreased the abundance of Lachnospiraceae | [218] | ||
| Resveratrol | Inhibited the growth of Enterococcus faecalis, and increased the growth of Bifidobacterium and Lactobacillus | [185] | ||
| Raspberry, cloudberry, and strawberry | In vitro | 0.5–5 mg/mL of berries extracts (24–48 h) | Inhibited the growth of Gram-negative but not Gram-positive bacteria | [188] |
| Schisandra chinensis | Human study (obese volunteers) | 6.7 g dry berry powder per day for 12 weeks | Promoted abundance of Bacteroides, Akkermansia, Roseburia, Prevotella Bifidobacterium, and (−) Ruminococcus | [199] |
| Sea buckthorn | In vitro | 250 mg lyophilized fraction of small intestine digested berries for 72 h | Promoted abundance of Bacteroides/Prevotella and Bifidobacteria | [219] |
| 10% berry juice for 1 week (continuous gut model) | Promoted abundance of Lactobacilli, Bacteroides/Prevotella, and Bifidobacteria | [220] | ||
| Strawberry | Animal study (mice; diabetic model) | 2.35% (dry berry powder) for 10 weeks | Promoted abundance of Bifidobacterium, (−) Bacteroides, and Akkermansia | [221] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouyahya, A.; Omari, N.E.; EL Hachlafi, N.; Jemly, M.E.; Hakkour, M.; Balahbib, A.; El Menyiy, N.; Bakrim, S.; Naceiri Mrabti, H.; Khouchlaa, A.; et al. Chemical Compounds of Berry-Derived Polyphenols and Their Effects on Gut Microbiota, Inflammation, and Cancer. Molecules 2022, 27, 3286. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103286
Bouyahya A, Omari NE, EL Hachlafi N, Jemly ME, Hakkour M, Balahbib A, El Menyiy N, Bakrim S, Naceiri Mrabti H, Khouchlaa A, et al. Chemical Compounds of Berry-Derived Polyphenols and Their Effects on Gut Microbiota, Inflammation, and Cancer. Molecules. 2022; 27(10):3286. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103286
Chicago/Turabian StyleBouyahya, Abdelhakim, Nasreddine El Omari, Naoufal EL Hachlafi, Meryem El Jemly, Maryam Hakkour, Abdelaali Balahbib, Naoual El Menyiy, Saad Bakrim, Hanae Naceiri Mrabti, Aya Khouchlaa, and et al. 2022. "Chemical Compounds of Berry-Derived Polyphenols and Their Effects on Gut Microbiota, Inflammation, and Cancer" Molecules 27, no. 10: 3286. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103286