
Potential Therapeutic Benefits of Honey in Neurological Disorders: The Role of Polyphenols

 ,
,  ,
,  ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Composition of Honey
3. Medicinal Properties of Honey
4. Honey and Neurological Disorders
4.1. Alzheimer’s Disease
4.2. Parkinson’s Disease
4.3. Huntington’s Disease
4.4. Amyotrophic Lateral Sclerosis
4.5. Epilepsy
4.6. Schizophrenia
4.7. Depression
5. Conclusions
6. Limitations and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grossman, R. The Other Medicines: The Penicillin of Bees; Pan Books: London, UK, 1986; p. 177. [Google Scholar]
- Wootton, M.; Edwards, R.A.; Faraji-Haremi, R.; Williams, P.J. Effect of accelerated storage conditions on the chemical composition and properties of Australian honeys. J. Apic. Res. 1978, 17, 167–172. [Google Scholar] [CrossRef]
- Majno, G. The Healing Hand: Man and Wound in the Ancient World; Harvard University Press: Cambridge, MA, USA, 1991. [Google Scholar]
- Asadi-Pooya, A.A.; Pnjehshahin, M.R.; Beheshti, S. The antimycobacterial effect of honey: An in vitro study. Riv. Biol. 2003, 96, 491–495. [Google Scholar] [PubMed]
- Hippocrates. On the articulations. The genuine works of Hippocrates. Clin. Orthop. Relat. Res. 2002, 400, 19–25. [Google Scholar]
- Molan, P.C. Why honey is effective as a medicine. Its use in modern medicine. Bee World 1999, 80, 80–92. [Google Scholar] [CrossRef]
- White, J.W., Jr.; Hoban, N. Composition of honey. IV. Identification of the disaccharides. Arch. Biochem. Biophys. 1959, 80, 386–392. [Google Scholar] [CrossRef]
- White, J.W., Jr. The composition of honey. Bee World 1957, 80, 386–392. [Google Scholar] [CrossRef]
- Eteraf-Oskouei, T.; Najafi, M. Traditional and modern uses of natural honey in human diseases: A review. Iran. J. Basic. Med. Sci. 2013, 16, 731–742. [Google Scholar]
- Michiu, D.; Socaciu, M.I.; Fogarasi, M.; Jimborean, A.M.; Ranga, F.; Mureşan, V.; Semeniuc, C.A. Implementation of an Analytical Method for Spectrophotometric Evaluation of Total Phenolic Content in Essential Oils. Molecules 2022, 27, 1345. [Google Scholar] [CrossRef]
- Biesaga, M.; Pyrzyńska, K. Stability of bioactive polyphenols from honey during different extraction methods. Food Chem. 2013, 136, 46–54. [Google Scholar] [CrossRef]
- Pyrzynska, K.; Biesaga, M. Analysis of phenolic acids and flavonoids in honey. TrAC-Trend Anal. Chem. 2009, 28, 893–902. [Google Scholar] [CrossRef]
- Khalil, M.I.; Sulaiman, S.A. The potential role of honey and its polyphenols in preventing heart diseases: A review. Afr. J. Tradit. Complement. Altern. Med. 2010, 7, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossen, M.S.; Ali, M.Y.; Jahurul, M.H.; Abdel-Daim, M.M.; Gan, S.H.; Khalil, M.I. Beneficial roles of honey polyphenols against some human degenerative diseases: A review. Pharmacol. Rep. 2017, 69, 1194–1205. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Datta, N.; Tomas-Barberan, F.A.; Ferreres, F.; Martos, I.; Singanusong, R. Flavonoids, phenolic acids and abscisic acid in Australian and New Zealand Leptospermum honeys. Food Chem. 2003, 81, 159–168. [Google Scholar] [CrossRef]
- Deng, J.; Liu, R.; Lu, Q.; Hao, P.; Xu, A.; Zhang, J.; Tan, J. Biochemical properties, antibacterial and cellular antioxidant activities of buckwheat honey in comparison to manuka honey. Food Chem. 2018, 252, 243–249. [Google Scholar] [CrossRef]
- Stephens, J.M.; Schlothauer, R.C.; Morris, B.D.; Yang, D.; Fearnley, L.; Greenwood, D.R.; Loomes, K.M. Phenolic compounds and methylglyoxal in some New Zealand manuka and kanuka honeys. Food Chem. 2010, 120, 78–86. [Google Scholar] [CrossRef]
- Šarić, G.; Vahčić, N.; Bursać Kovačević, D.; Putnik, P. The changes of flavonoids in honey during storage. Processes 2020, 8, 943. [Google Scholar] [CrossRef]
- Marghitas, L.A.; Dezmirean, D.S.; Pocol, C.B.; Marioara, I.L.; Bobis, O.; Gergen, I. The development of a biochemical profile of acacia honey by identifying biochemical determinants of its quality. Not. Bot. Horti. Agrobot. Cluj-Napoca 2010, 38, 84–90. [Google Scholar]
- Martos, I.; Ferreres, F.; Yao, L.; D’Arcy, B.; Caffin, N.; Tomás-Barberán, F.A. Flavonoids in monospecific eucalyptus honeys from Australia. J. Agric. Food. Chem. 2000, 48, 4744–4748. [Google Scholar] [CrossRef]
- Perna, A.; Intaglietta, I.; Simonetti, A.; Gambacorta, E. A comparative study on phenolic profile, vitamin C content and antioxidant activity of Italian honeys of different botanical origin. Int. J. Food Sci. Technol. 2013, 48, 1899–1908. [Google Scholar] [CrossRef]
- Kıvrak, Ş.; Kıvrak, İ. Assessment of phenolic profile of Turkish honeys. Int. J. Food Prop. 2017, 20, 864–876. [Google Scholar] [CrossRef] [Green Version]
- Kečkeš, S.; Gašić, U.; Veličković, T.Ć.; Milojković-Opsenica, D.; Natić, M.; Tešić, Ž. The determination of phenolic profiles of Serbian unifloral honeys using ultra-high-performance liquid chromatography/high resolution accurate mass spectrometry. Food Chem. 2013, 138, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Hamdy, A.A.; Ismail, H.M.; Al-Ahwal, A.-M.; Gomaa, N.F. Determination of flavonoid and phenolic Acid contents of clover, cotton and citrus floral honeys. J. Egypt. Public Health Assoc. 2009, 84, 245–259. [Google Scholar] [PubMed]
- Zumla, A.; Lulat, A. Honey—A remedy rediscovered. J. R. Soc. Med. 1989, 82, 384–385. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, M.; Sirajudeen, K.N.; Swamy, M.; Yaacob, M.; Sulaiman, S. Studies on the antioxidant properties of Tualang honey of Malaysia. Afr. J. Tradit. Complement. Altern. Med. 2010, 7, 59–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abeshu, M.A.; Geleta, B. Medicinal uses of honey. Biol. Med. 2016, 8, 1. [Google Scholar]
- Swellam, T.; Miyanaga, N.; Onozawa, M.; Hattori, K.; Kawai, K.; Shimazui, T.; Akaza, H. Antineoplastic activity of honey in an experimental bladder cancer implantation model: In vivo and in vitro studies. Int. J. Urol. 2003, 10, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.S.; Bhowmik, D.; Biswajit, C.; Chandira, M.R. Medicinal uses and health benefits of honey: An overview. J. Chem. Pharm. Res. 2010, 2, 385–395. [Google Scholar]
- Dustmann, J.H. Antibacterial effect of honey. Apiacta 1979, 14, 7–11. [Google Scholar]
- Johnston, M.; McBride, M.; Dahiya, D.; Owusu-Apenten, R.; Nigam, P.S. Antibacterial activity of Manuka honey and its components: An overview. AIMS Microbiol. 2018, 4, 655–664. [Google Scholar] [CrossRef]
- Albaridi, N.A. Antibacterial Potency of Honey. Int. J. Microbiol. 2019, 2019, 2464507. [Google Scholar] [CrossRef]
- Onifade, A.A.; Jewell, A.P.; Ajadi, T.A.; Rahamon, S.K.; Ogunrin, O.O. Effectiveness of a herbal remedy in six HIV patients in Nigeria. J. Herb. Med. 2013, 3, 99–103. [Google Scholar] [CrossRef]
- Fingleton, J.; Corin, A.; Sheahan, D.; Cave, N.; Braithwaite, I.; Weatherall, M.; Beasley, R. Randomised controlled trial of topical kanuka honey for the treatment of cold sores. Adv. Integr. Med. 2014, 1, 119–123. [Google Scholar] [CrossRef]
- Anyanwu, C.U. Investigation of in vitro antifungal activity of honey. J. Med. Plants Res. 2012, 6, 3512–3516. [Google Scholar] [CrossRef]
- Ahmad, K.; Khali, A.T.; Somayya, R.; Khan, F.N.; Shah, A.R.; Ovais, M.; Shinwari, Z.K. Potential antifungal activity of different honey brands from Pakistan: A quest for natural remedy. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Hau-Yama, N.E.; Magaña-Ortiz, D.; Oliva, A.I.; Ortiz-Vázquez, E. Antifungal activity of honey from stingless bee Melipona beecheii against Candida albicans. J. Apic. Res. 2020, 59, 12–18. [Google Scholar] [CrossRef]
- Erejuwa, O.O.; Sulaiman, S.A.; Ab Wahab, M.S. Honey: A novel antioxidant. Molecules 2012, 17, 4400–4423. [Google Scholar] [CrossRef]
- Gül, A.; Pehlivan, T. Antioxidant activities of some monofloral honey types produced across Turkey. Saudi. J. Biol. Sci. 2018, 25, 1056–1065. [Google Scholar] [CrossRef]
- Dżugan, M.; Tomczyk, M.; Sowa, P.; Grabek-Lejko, D. Antioxidant Activity as Biomarker of Honey Variety. Molecules 2018, 23, 2069. [Google Scholar] [CrossRef] [Green Version]
- Almasaudi, S.B.; Abbas, A.T.; Al-Hindi, R.R.; El-Shitany, N.A.; Abdel-Dayem, U.A.; Ali, S.S.; Saleh, R.M.; Al Jaouni, S.K.; Kamal, M.A.; Harakeh, S.M. Manuka Honey Exerts Antioxidant and Anti-Inflammatory Activities That Promote Healing of Acetic Acid-Induced Gastric Ulcer in Rats. Evid. Based. Complement. Altern. Med. 2017, 2017, 5413917. [Google Scholar] [CrossRef]
- Ghazali, W.S.; Romli, A.C.; Mohamed, M. Effects of honey supplementation on inflammatory markers among chronic smokers: A randomized controlled trial. BMC Complement. Altern. Med. 2017, 17, 175. [Google Scholar] [CrossRef]
- Minden-Birkenmaier, B.A.; Cherukuri, K.; Smith, R.A.; Radic, M.Z.; Bowlin, G.L. Manuka honey modulates the inflammatory behavior of a dHL-60 neutrophil Model under the cytotoxic limit. Int. J. Biomater. 2019, 2019, 6132581. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.I.; Mabrouk, G.M.; Shehata, H.H.; Aboelhussein, M.M. Antineoplastic effects of bee honey and Nigella sativa on hepatocellular carcinoma cells. Integr. Cancer. Ther. 2012, 11, 354–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efem, S.E.; Udoh, K.T.; Iwara, C.I. The antimicrobial spectrum of honey and its clinical significance. Infection 1992, 20, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Almasaudi, S.B.; Al-Nahari, A.A.M.; Abd El-Ghany, E.S.M.; Barbour, E.; Al Muhayawi, S.M.; Al-Jaouni, S.; Azhar, E.; Qari, M.; Qari, Y.A.; Harakeh, S. Antimicrobial effect of different types of honey on Staphylococcus aureus. Saudi J. Biol. Sci. 2017, 24, 1255–1261. [Google Scholar] [CrossRef] [Green Version]
- Bouzo, D.; Cokcetin, N.N.; Li, L.; Ballerin, G.; Bottomley, A.L.; Lazenby, J.; Whitchurch, C.B.; Paulsen, I.T.; Hassan, K.A.; Harry, E.J. Characterizing the mechanism of action of an ancient antimicrobial, Manuka honey, against Pseudomonas aeruginosa using modern transcriptomics. MSystems 2020, 5, e00106-20. [Google Scholar] [CrossRef]
- Ahmed, S.; Othman, N.H. The anti-cancer effects of Tualang honey in modulating breast carcinogenesis: An experimental animal study. BMC Complement. Altern. Med. 2017, 17, 208. [Google Scholar] [CrossRef] [Green Version]
- Aryappalli, P.; Al-Qubaisi, S.S.; Attoub, S.; George, J.A.; Arafat, K.; Ramadi, K.B.; Mohamed, Y.A.; Al-Dhaheri, M.M.; Al-Sbiei, A.; Fernandez-Cabezudo, M.J.; et al. The IL-6/STAT3 signaling pathway is an early target of manuka honey-induced suppression of human breast cancer cells. Front. Oncol. 2017, 7, 167. [Google Scholar] [CrossRef] [Green Version]
- Taban, Q.; Mumtaz, P.T.; Ali, A. Honey in Anticancer Drug Toxicity. In Therapeutic Applications of Honey and Its Phytochemicals; Springer: Berlin, Germany, 2020; pp. 307–324. [Google Scholar]
- Najafi, M.; Mahdizadeh, A.E.; Rafiei, F.; Eteraf, O.T. Effects of pharmacologic preconditioning by natural honey on arrhythmias and infarct size in isolated heart. Pharm. Sci. 2008, 4, 1–11. [Google Scholar]
- Najafi, M.; Shaseb, E.; Ghaffary, S.; Fakhrju, A.; Eteraf Oskouei, T. Effects of chronic oral administration of natural honey on ischemia/reperfusion-induced arrhythmias in isolated rat heart. Iran. J. Basic Med. Sci. 2011, 14, 75–81. [Google Scholar]
- Zeina, B.; Zohra, B.I.; Al-Assad, S. The effects of honey on Leishmania parasites: An in vitro study. Trop. Doct. 1997, 27 (Suppl. S1), 36–38. [Google Scholar]
- Gharirvand Eskandari, E.; Setorki, M.; Doudi, M. Medicinal plants with antileishmanial properties: A review study. Pharm. Biomed. Res. 2020, 6, 1–16. [Google Scholar] [CrossRef]
- Ahmed, A.; Khan, R.A.; Azim, M.K.; Saeed, S.A.; Mesaik, M.A.; Ahmed, S.; Imran, I. Effect of natural honey on human platelets and blood coagulation proteins. Pak. J. Pharm. Sci. 2011, 24, 389–397. [Google Scholar] [PubMed]
- Olas, B. Honey and its phenolic compounds as an effective natural medicine for cardiovascular diseases in humans? Nutrients 2020, 12, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.H.; Andrae, L.; Engeseth, N.J. Antimutagenic effect of various honeys and sugars against Trp-p-1. J. Agric. Food. Chem. 2002, 50, 6923–6928. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Gautam, S.; Maru, G.; Kawle, D.; Sharma, A. Suppression of error prone pathway is responsible for antimutagenic activity of honey. Food. Chem. Toxicol. 2012, 50, 625–633. [Google Scholar] [CrossRef]
- Azim, M.K.; Perveen, H.; Mesaik, M.A.; Simjee, S.U. Antinociceptive activity of natural honey in thermal-nociception models in mice. Phytother. Res. 2007, 21, 194–197. [Google Scholar] [CrossRef]
- Geißler, K.; Schulze, M.; Inhestern, J.; Meißner, W.; Guntinas-Lichius, O. The effect of adjuvant oral application of honey in the management of postoperative pain after tonsillectomy in adults: A pilot study. PLoS ONE 2020, 15, e0228481. [Google Scholar] [CrossRef]
- Hannan, A.; Munir, S.; Arshad, M.U.; Bashir, N. In Vitro Antimycobacterial Activity of Pakistani Beri Honey Using BACTEC MGIT 960. Int. Sch. Res. Not. 2014, 2014, 490589. [Google Scholar] [CrossRef] [Green Version]
- Ghashm, A.A.; Othman, N.H.; Khattak, M.N.; Ismail, N.M.; Saini, R. Antiproliferative effect of Tualang honey on oral squamous cell carcinoma and osteosarcoma cell lines. BMC Complement. Altern. Med. 2010, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Mumtaz, P.T.; Bashir, S.M.; Rather, M.A.; Dar, K.B.; Taban, Q.; Sajood, S.; Ali, A.; Rather, Z.A.; Amin, I.; Dar, M.A. Antiproliferative and Apoptotic Activities of Natural Honey. In Therapeutic Applications of Honey and Its Phytochemicals; Springer: Singapore, 2020; pp. 345–360. [Google Scholar]
- Kassim, M.; Mansor, M.; Achoui, M.; Yan, O.S.; Devi, S.; Yusoff, K.M. Honey as an immunomodulator during sepsis in animal model. Critical Care 2009, 13, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Babaei, S.; Rahimi, S.; Torshizi, M.A.; Tahmasebi, G.; Miran, S.N. Effects of propolis, royal jelly, honey and bee pollen on growth performance and immune system of Japanese quails. Vet. Res. Forum. 2016, 7, 13–20. [Google Scholar] [PubMed]
- Wusiman, A.; Xu, S.; Ni, H.; Gu, P.; Liu, Z.; Zhang, Y.; Qiu, T.; Hu, Y.; Liu, J.; Wu, Y.; et al. Immunomodulatory effects of Alhagi honey polysaccharides encapsulated into PLGA nanoparticles. Carbohydr. Polym. 2019, 211, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Ariantari, N.P.; Yowani, S.C.; Swastini, D.A. UJji aktivitas penurunan kolesterol produk madu herbal yang beredar di pasaran pada tikus putih diet lemak tinggi [hypocholesterolemic activity of marketed herbal honey products in albino rats with hypercholesterolemic diet]. J. Kimia 2010, 4, 15–19. [Google Scholar]
- Mohamed, Z.B.; Alfarisi, H.A.; Wahab, A.Y.; binti Abd Fuaat, A.; Mohamad, C.A.; Ibrahim, M. Hypocholesterolemic and anti-inflammatory effects of trihoney in hypercholesterolemic rabbit model. Int. J. Allied Health Sci. 2019, 3, 846. [Google Scholar]
- Khalil, M.I.; Tanvir, E.M.; Afroz, R.; Sulaiman, S.A.; Gan, S.H. Cardioprotective Effects of Tualang Honey: Amelioration of Cholesterol and Cardiac Enzymes Levels. Biomed. Res. Int. 2015, 2015, 286051. [Google Scholar] [CrossRef] [Green Version]
- Bt Hj Idrus, R.; Sainik, N.Q.; Nordin, A.; Saim, A.B.; Sulaiman, N. Cardioprotective Effects of Honey and Its Constituent: An Evidence-Based Review of Laboratory Studies and Clinical Trials. Int. J. Environ. Res. Public Health 2020, 17, 3613. [Google Scholar] [CrossRef]
- Erejuwa, O.O.; Sulaiman, S.A.; Ab Wahab, M.S.; Sirajudeen, K.N.; Salleh, S.; Gurtu, S. Honey supplementation in spontaneously hypertensive rats elicits antihypertensive effect via amelioration of renal oxidative stress. Oxid. Med. Cell. Longev. 2012, 2012, 374037. [Google Scholar] [CrossRef] [Green Version]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.; Sirajudeen, K.N.; Salleh, M.S.; Gurtu, S. Hepatoprotective effect of tualang honey supplementation in streptozotocin-induced diabetic rats. Int. J. Appl. Res. Nat. Prod. 2012, 4, 37–41. [Google Scholar]
- Zhao, H.; Cheng, N.; He, L.; Peng, G.; Liu, Q.; Ma, T.; Cao, W. Hepatoprotective effects of the honey of apis cerana fabricius on bromobenzene-induced liver damage in mice. J. Food. Sci. 2018, 83, 509–516. [Google Scholar] [CrossRef]
- Gharzouli, K.; Amira, S.; Gharzouli, A.; Hennouf, S. Gastroprotective effects of honey and glucose-fructose-sucrose-maltose mixture against ethanol-, indomethacin-, and acidified aspirin-induced lesions in the rat. Exp. Toxicol. Pathol. 2002, 54, 217–221. [Google Scholar] [CrossRef]
- Qaid, A.; Yaseen, E. Neuroprotective Effects of Tualang Honey in Male Rats Exposed to Normobaric Hypoxia. Ph.D. Thesis, Kampus Kesihatan, Universiti Sains Malaysia, Penang, Malaysia, 2017. [Google Scholar]
- Azman, K.F.; Zakaria, R.; Othman, Z.; Abdul Aziz, C.B. Neuroprotective effects of Tualang honey against oxidative stress and memory decline in young and aged rats exposed to noise stress. J. Taibah Univ. Sci. 2018, 12, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.; Eldaim, M.A.; Abdel-Daim, M.M. Nephroprotective effect of bee honey and royal jelly against subchronic cisplatin toxicity in rats. Cytotechnology 2016, 68, 1039–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obia, O.; Odum, J.E.; Chuemere, A.N. Nephroprotective and antihyperlipidemic activity honey in alloxan-induced diabetic rats. Int. J. Biochem. Res. Rev. 2018, 22, 1–7. [Google Scholar] [CrossRef]
- Erejuwa, O.O.; Gurtu, S.; Sulaiman, S.A.; Ab Wahab, M.S.; Sirajudeen, K.N.; Salleh, M.S. Hypoglycemic and antioxidant effects of honey supplementation in streptozotocin-induced diabetic rats. Int. J. Vitam. Nutr. Res. 2010, 80, 74–82. [Google Scholar] [PubMed]
- El-Haskoury, R.; Al-Waili, N.; El-Hilaly, J.; Al-Waili, W.; Lyoussi, B. Antioxidant, hypoglycemic, and hepatoprotective effect of aqueous and ethyl acetate extract of carob honey in streptozotocin-induced diabetic rats. Vet. World 2019, 12, 1916. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.; Sulaiman, S.A.; Jaafar, H.; Sirajudeen, K.N. Antioxidant protective effect of honey in cigarette smoke-induced testicular damage in rats. Int. J. Mol. Sci. 2011, 12, 5508–5521. [Google Scholar] [CrossRef] [Green Version]
- Sharifah, D. Effects of nicotine and Gelam honey on testis parameters and sperm qualities of juvenile rats. Sci. Res. Essays. 2011, 6, 5471–5474. [Google Scholar]
- Zaid, S.S.; Sulaiman, S.A.; Sirajudeen, K.N.; Othman, N.H. The effects of Tualang honey on female reproductive organs, tibia bone and hormonal profile in ovariectomised rats—Animal model for menopause. BMC Complement. Altern. Med. 2010, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Zaid, S.S.; Othman, S.; Kassim, N.M. Potential protective effect of Tualang honey on BPA-induced ovarian toxicity in prepubertal rat. BMC Complement. Altern. Med. 2014, 14, 509. [Google Scholar] [CrossRef] [Green Version]
- Carroll, W.M. The global burden of neurological disorders. Lancet Neurol. 2019, 18, 418–419. [Google Scholar] [CrossRef] [Green Version]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, J.; Pietrzak, M.; Pomianowski, J.; Wieczorek, Z. Honey as a source of bioactive compounds. Pol. J. Nat. Sci. 2014, 29, 275–285. [Google Scholar]
- Mijanur Rahman, M.; Gan, S.H.; Khalil, M.I. Neurological effects of honey: Current and future prospects. Evid. Based Complement. Altern. Med. 2014, 2014, 958721. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi, T.; Kanzaki, N.; Otsuka, Y.; Izumo, T.; Shibata, H.; Machida, S. Quercetin inhibits adipogenesis of muscle progenitor cells in vitro. Biochem. Biophys. Rep. 2017, 13, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tian, Q.; Li, Z.; Dang, M.; Lin, Y.; Hou, X. Activation of Nrf2 signaling by sitagliptin and quercetin combination against β-amyloid induced Alzheimer’s disease in rats. Drug. Dev. Res. 2019, 80, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Zaplatic, E.; Bule, M.; Shah, S.Z.A.; Uddin, M.S.; Niaz, K. Molecular mechanisms underlying protective role of quercetin in attenuating Alzheimer’s disease. Life Sci. 2019, 224, 109–119. [Google Scholar] [CrossRef]
- Sabogal-Guáqueta, A.M.; Munoz-Manco, J.I.; Ramírez-Pineda, J.R.; Lamprea-Rodriguez, M.; Osorio, E.; Cardona-Gómez, G.P. The flavonoid quercetin ameliorates Alzheimer’s disease pathology and protects cognitive and emotional function in aged triple transgenic Alzheimer’s disease model mice. Neuropharmacology 2015, 93, 134–145. [Google Scholar] [CrossRef] [Green Version]
- Moreno, L.C.G.E.I.; Puerta, E.; Suárez-Santiago, J.E.; Santos-Magalhães, N.S.; Ramirez, M.J.; Irache, J.M. Effect of the oral administration of nanoencapsulated quercetin on a mouse model of Alzheimer’s disease. Int. J. Pharm. 2017, 517, 50–57. [Google Scholar] [CrossRef]
- Ramezani, M.; Darbandi, N.; Khodagholi, F.; Hashemi, A. Myricetin protects hippocampal CA3 pyramidal neurons and improves learning and memory impairments in rats with Alzheimer’s disease. Neural Regen. Res. 2016, 11, 1976. [Google Scholar] [CrossRef]
- Semwal, D.K.; Semwal, R.B.; Combrinck, S.; Viljoen, A. Myricetin: A Dietary Molecule with Diverse Biological Activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhong, Y.; Gao, C.; Li, J. Myricetin ameliorates scopolamine-induced memory impairment in mice via inhibiting acetylcholinesterase and down-regulating brain iron. Biochem. Biophys. Res. Commun. 2017, 490, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lin, H.; Xie, S.; Huang, B.; Qian, Y.; Chen, K.; Niu, Y.; Shen, H.M.; Cai, J.; Li, P.; et al. Myricetin inhibits NLRP3 inflammasome activation via reduction of ROS-dependent ubiquitination of ASC and promotion of ROS-independent NLRP3 ubiquitination. Toxicol. Appl. Pharmacol. 2019, 365, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Krueyos, N.; Ritthiruangdej, P. Anti-oxidative assays as markers for anti-inflammatory activity of flavonoids. Int. Immunopharmacol. 2016, 40, 170–175. [Google Scholar] [CrossRef]
- Park, K.S.; Chong, Y.; Kim, M.K. Myricetin: Biological activity related to human health. Appl. Biol. Chem. 2016, 59, 259–269. [Google Scholar] [CrossRef]
- Beg, T.; Jyoti, S.; Naz, F.; Rahul Ali, F.; Ali, S.K.; Reyad, A.M.; Siddique, Y.H. Protective Effect of Kaempferol on the Transgenic Drosophila Model of Alzheimer’s Disease. CNS Neurol. Disord. Drug Targets. 2018, 17, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Kouhestani, S.; Jafari, A.; Babaei, P. Kaempferol attenuates cognitive deficit via regulating oxidative stress and neuroinflammation in an ovariectomized rat model of sporadic dementia. Neural Regen. Res. 2018, 13, 1827. [Google Scholar]
- Kim, J.H.; Lee, S.; Cho, E.J.; Kim, H.Y. Neuroprotective effects of kaempferol, quercetin, and its glycosides by regulation of apoptosis. J. Korea Acad.-Ind. Corp. Soc. 2019, 20, 286–293. [Google Scholar]
- Darbandi, N.; Ramezani, M.; Khodagholi, F.; Noori, M. Kaempferol promotes memory retention and density of hippocampal CA1 neurons in intra-cerebroventricular STZ-induced experimental AD model in Wistar rats. Biologija 2016, 62. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, H.Y.; Cho, E.J. Protective effects of kaempferol, quercetin, and its glycosides on amyloid beta-induced neurotoxicity in C6 glial cell. J. Appl. Biol. Chem. 2019, 62, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Pate, K.M.; Rogers, M.; Reed, J.W.; van der Munnik, N.; Vance, S.Z.; Moss, M.A. Anthoxanthin polyphenols attenuate aβ oligomer-induced neuronal responses associated with Alzheimer’s disease. CNS Neurosci. Ther. 2017, 23, 135–144. [Google Scholar] [CrossRef]
- Ali, F.; Siddique, Y.H. Bioavailability and Pharmaco-therapeutic Potential of Luteolin in Overcoming Alzheimer’s Disease. CNS Neurol. Disord. Drug Targets 2019, 18, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, H.; Cheng, H.; Che, Z. Ameliorating effect of luteolin on memory impairment in an Alzheimer’s disease model. Mol. Med. Rep. 2016, 13, 4215–4220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Lin, M.; Zhong, X.; Yang, H.; Deng, M. Galangin decreases p-tau, Aβ42 and β-secretase levels, and suppresses autophagy in okadaic acid-induced PC12 cells via an Akt/GSK3β/mTOR signaling-dependent mechanism. Mol. Med. Rep. 2019, 19, 1767–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilic, F.S.; Kaygisiz, B.; Aydin, S.; Yildirim, E.; Oner, S.; Erol, K. The effects and mechanisms of the action of galangin on spatial memory in rats. Bratisl. Lek. Listy 2019, 120, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Li, J.; Hua, L.; Han, B.; Zhang, Y.; Yang, X.; Zeng, Z.; Bai, H.; Yin, H.; et al. Effects of caffeic acid on learning deficits in a model of Alzheimer’s disease. Int. J. Mol. Med. 2016, 38, 869–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, S.; Roy, A.; Jana, M.; Pahan, K. Cinnamic acid activates PPARα to stimulate Lysosomal biogenesis and lower Amyloid plaque pathology in an Alzheimer’s disease mouse model. Neurobiol. Dis. 2019, 124, 379–395. [Google Scholar] [CrossRef]
- Ay, M.; Luo, J.; Langley, M.; Jin, H.; Anantharam, V.; Kanthasamy, A.; Kanthasamy, A.G. Molecular mechanisms underlying protective effects of quercetin against mitochondrial dysfunction and progressive dopaminergic neurodegeneration in cell culture and MitoPark transgenic mouse models of Parkinson’s Disease. J. Neurochem. 2017, 141, 766–782. [Google Scholar] [CrossRef]
- Ghaffari, F.; Moghaddam, A.H.; Zare, M. Neuroprotective effect of quercetin nanocrystal in a 6-hydroxydopamine model of parkinson disease: Biochemical and behavioral evidence. Basic Clin. Neurosci. 2018, 9, 317. [Google Scholar] [CrossRef]
- El-Horany, H.E.; El-latif, R.N.; ElBatsh, M.M.; Emam, M.N. Ameliorative effect of quercetin on neurochemical and behavioral deficits in rotenone rat model of Parkinson’s disease: Modulating autophagy (quercetin on experimental Parkinson’s disease). J. Biochem. Mol. Toxicol. 2016, 30, 360–369. [Google Scholar] [CrossRef]
- Ara, G.; Afzal, M.; Jyoti, S.; Naz, F.; Siddique, Y.H. Effect of Myricetin on the Loss of Dopaminergic Neurons in the Transgenic Drosophila Model of Parkinson’s Disease. Curr. Drug Ther. 2019, 14, 58–64. [Google Scholar] [CrossRef]
- Huang, B.; Liu, J.; Ma, D.; Chen, G.; Wang, W.; Fu, S. Myricetin prevents dopaminergic neurons from undergoing neuroinflammation-mediated degeneration in a lipopolysaccharide-induced Parkinson’s disease model. J. Funct. Foods 2018, 45, 452–461. [Google Scholar] [CrossRef]
- Guo, B.; Zheng, C.; Cai, W.; Cheng, J.; Wang, H.; Li, H.; Sun, Y.; Cui, W.; Wang, Y.; Han, Y.; et al. Multifunction of Chrysin in Parkinson’s Model: Anti-Neuronal Apoptosis, Neuroprotection via Activation of MEF2D, and Inhibition of Monoamine Oxidase-B. J. Agric. Food Chem. 2016, 64, 5324–5333. [Google Scholar] [CrossRef] [PubMed]
- Del Fabbro, L.; Rossito Goes, A.; Jesse, C.R.; de Gomes, M.G.; Cattelan Souza, L.; Lobo Ladd, F.V.; Lobo Ladd, A.A.B.; Nunes Arantes, R.V.; Reis Simionato, A.; Oliveira, M.S.; et al. Chrysin protects against behavioral, cognitive and neurochemical alterations in a 6-hydroxydopamine model of Parkinson’s disease. Neurosci. Lett. 2019, 706, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.R.; Shaikh, M.A.; Ul Haq, S.H.I.; Nazir, S. Neuroprotective role of chrysin in attenuating loss of dopaminergic neurons and improving motor, learning and memory functions in rats. Int. J. Health Sci. 2018, 12, 35–43. [Google Scholar]
- Khatri, D.K.; Juvekar, A.R. Kinetics of Inhibition of Monoamine Oxidase Using Curcumin and Ellagic Acid. Pharmacogn. Mag. 2016, 12 (Suppl. S2), S116–S120. [Google Scholar]
- Sarkaki, A.; Farbood, Y.; Dolatshahi, M.; Mansouri, S.M.; Khodadadi, A. Neuroprotective Effects of Ellagic Acid in a Rat Model of Parkinson’s Disease. Acta Med. Iran. 2016, 54, 494–502. [Google Scholar]
- Baluchnejadmojarad, T.; Rabiee, N.; Zabihnejad, S.; Roghani, M. Ellagic acid exerts protective effect in intrastriatal 6-hydroxydopamine rat model of Parkinson’s disease: Possible involvement of ERβ/Nrf2/HO-1 signaling. Brain Res. 2017, 1662, 23–30. [Google Scholar] [CrossRef]
- Prorok, T.; Jana, M.; Patel, D.; Pahan, K. Cinnamic acid protects the nigrostriatum in a mouse model of Parkinson’s disease via peroxisome proliferator-activated receptorα. Neurochem. Res. 2019, 44, 751–762. [Google Scholar] [CrossRef]
- Chen, G.; Liu, J.; Jiang, L.; Ran, X.; He, D.; Li, Y.; Huang, B.; Wang, W.; Fu, S. Galangin Reduces the Loss of Dopaminergic Neurons in an LPS-Evoked Model of Parkinson’s Disease in Rats. Int. J. Mol Sci. 2017, 19, 12. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.J.; Lee, E.J.; Park, J.S.; Kim, S.N.; Park, E.M.; Kim, H.S. Anti-inflammatory mechanism of galangin in lipopolysaccharide-stimulated microglia: Critical role of PPAR-γ signaling pathway. Biochem. Pharmacol. 2017, 144, 120–131. [Google Scholar] [CrossRef]
- Chaturvedi, R.K.; Beal, M.F. PPAR: A therapeutic target in Parkinson’s disease. J. Neurochem. 2008, 106, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Carta, A.R. PPAR-γ: Therapeutic prospects in Parkinson’s disease. Curr. Drug. Targets 2013, 14, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Siddique, Y.H.; Jyoti, S. Alteration in biochemical parameters in the brain of transgenic Drosophila melanogaster model of Parkinson’s disease exposed to apigenin. Integr. Med. Res. 2017, 6, 245–253. [Google Scholar] [CrossRef]
- Sandhir, R.; Mehrotra, A. Quercetin supplementation is effective in improving mitochondrial dysfunctions induced by 3-nitropropionic acid: Implications in Huntington’s disease. Biochim. Biophys. Acta. Mol. Basis Dis. 2013, 1832, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, J.; Singh, R.; Dutta, D.; Naskar, A.; Rajamma, U.; Mohanakumar, K.P. Quercetin improves behavioral deficiencies, restores astrocytes and microglia, and reduces serotonin metabolism in 3-nitropropionic acid-induced rat model of Huntington’s disease. CNS Neurosci. Ther. 2014, 20, 10–19. [Google Scholar] [CrossRef]
- Joshi, V.; Mishra, R.; Upadhyay, A.; Amanullah, A.; Poluri, K.M.; Singh, S.; Kumar, A.; Mishra, A. Polyphenolic flavonoid (Myricetin) upregulated proteasomal degradation mechanisms: Eliminates neurodegenerative proteins aggregation. J. Cell. Physiol. 2019, 234, 20900–20914. [Google Scholar] [CrossRef] [PubMed]
- Khan, E.; Tawani, A.; Mishra, S.K.; Verma, A.K.; Upadhyay, A.; Kumar, M.; Sandhir, R.; Mishra, A.; Kumar, A. Myricetin Reduces Toxic Level of CAG Repeats RNA in Huntington’s Disease (HD) and Spino Cerebellar Ataxia (SCAs). ACS Chem. Biol. 2018, 13, 180–188. [Google Scholar] [CrossRef]
- Thangarajan, S.; Ramachandran, S.; Krishnamurthy, P. Chrysin exerts neuroprotective effects against 3-Nitropropionic acid induced behavioral despair—Mitochondrial dysfunction and striatal apoptosis via upregulating Bcl-2 gene and downregulating Bax—Bad genes in male wistar rats. Biomed. Pharmacother. 2016, 84, 514–525. [Google Scholar] [CrossRef]
- Haider, M.; Salman, M.; Kaushik, P.; Bharadwaj, N.; Aggarwal, N.B.; Tabassum, H.; Parvez, S. Chrysin ameliorates 3 nitropropinoic acid induced neurotoxicity targeting behavioural, biochemical and histological alterations. Int. J. Neurosci. 2020, 1–9. [Google Scholar] [CrossRef]
- Alarcón-Herrera, N.; Flores-Maya, S.; Bellido, B.; García-Bores, A.M.; Mendoza, E.; Ávila-Acevedo, G.; Hernández-Echeagaray, E. Protective effects of chlorogenic acid in 3-nitropropionic acid induced toxicity and genotoxicity. Food Chem. Toxicol. 2017, 109, 1018–1025. [Google Scholar] [CrossRef]
- Ip, P.; Sharda, P.R.; Cunningham, A.; Chakrabartty, S.; Pande, V.; Chakrabartty, A. Quercitrin and quercetin 3-β-d-glucoside as chemical chaperones for the A4V SOD1 ALS-causing mutant. Protein Eng. Des. Sel. 2017, 30, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, N.K.; Modi, P.; Sharma, S.; Deep, S. Quercetin and Baicalein Act as Potent Antiamyloidogenic and Fibril Destabilizing Agents for SOD1 Fibrils. ACS Chem. Neurosci. 2020, 11, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Inden, M.; Shirai, K.; Sekine, S.I.; Masaki, Y.; Kurita, H.; Ichihara, K.; Inuzuka, T.; Hozumi, I. The effects of Brazilian green propolis that contains flavonols against mutant copper-zinc superoxide dismutase-mediated toxicity. Sci. Rep. 2017, 7, 2882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, T.; Inden, M.; Kiuchi, M.; Asaka, Y.; Kurita, H.; Hozumi, I. The effects of kaempferol against mutant copper-zinc superoxide dismutase-mediated toxicity via autophagy. In Proceedings of the Annual Meeting of the Japanese Pharmacological Society WCP2018 (The 18th World Congress of Basic and Clinical Pharmacology), Kyoto, Japan, 1–6 July 2018. [Google Scholar]
- Ueda, T.; Ito, T.; Kurita, H.; Inden, M.; Hozumi, I. p-Coumaric acid has protective effects against mutant copper–zinc superoxide dismutase 1 via the activation of autophagy in N2a cells. Int. J. Mol. Sci. 2019, 20, 2942. [Google Scholar] [CrossRef] [Green Version]
- Maya, S.; Prakash, T.; Goli, D. Evaluation of neuroprotective effects of wedelolactone and gallic acid on aluminium-induced neurodegeneration: Relevance to sporadic amyotrophic lateral sclerosis. Eur. J. Pharmacol. 2018, 835, 41–51. [Google Scholar] [CrossRef]
- Maya, S.; Prakash, T.; Goli Daya, S. Effect of wedelolactone and gallic acid on quinolinic acid-induced neurotoxicity and impaired motor function: Significance to sporadic amyotrophic lateral sclerosis. Neurotoxicology 2018, 68, 1–12. [Google Scholar]
- Aaron, C.; Beaudry, G.; Parker, J.A.; Therrien, M. Maple Syrup Decreases TDP-43 Proteotoxicity in a Caenorhabditis elegans Model of Amyotrophic Lateral Sclerosis (ALS). J. Agric. Food Chem. 2016, 64, 3338–3344. [Google Scholar] [CrossRef]
- Wu, D.; Zheng, Z.; Fan, S.; Wen, X.; Han, X.; Wang, S.; Wang, Y.; Zhang, Z.; Shan, Q.; Li, M.; et al. Ameliorating effect of quercetin on epilepsy by inhibition of inflammation in glial cells. Exp. Ther. Med. 2020, 20, 854–859. [Google Scholar] [CrossRef]
- Moghbelinejad, S.; Rashvand, Z.; Khodabandehloo, F.; Mohammadi, G.; Nassiri-Asl, M. Modulation of the Expression of the GABAA Receptor β1 and β3 Subunits by Pretreatment with Quercetin in the KA Model of Epilepsy in Mice: The Effect of Quercetin on GABAA Receptor Beta Subunits. J. Pharmacopunct. 2016, 19, 163–166. [Google Scholar]
- Singh, T.; Kaur, T.; Goel, R.K. Adjuvant quercetin therapy for combined treatment of epilepsy and comorbid depression. Neurochem. Int. 2017, 104, 27–33. [Google Scholar] [CrossRef]
- Parihar, G.; Dehariya, B.; Ghule, S.; Dixit, P.; Balekar, N. Quercetin Exerts Anti-convulsant Effects in Animal Model of Grand Mal Epilepsy: Modulation of GABA and Glycinergic Pathways. J. Drug Deliv. Ther. 2017, 7, 194–196. [Google Scholar]
- Hashemian, M.; Ghasemi-Kasman, M.; Ghasemi, S.; Akbari, A.; Moalem-Banhangi, M.; Zare, L.; Ahmadian, S.R. Fabrication and evaluation of novel quercetin-conjugated Fe3O4–β-cyclodextrin nanoparticles for potential use in epilepsy disorder. Int. J. Nanomedicine. 2019, 14, 6481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.Q.; Meng, F.H.; Tu, L.X.; Sun, L. Myricetin attenuates the severity of seizures and neuroapoptosis in pentylenetetrazole kindled mice by regulating the of BDNF-TrkB signaling pathway and modulating matrix metalloproteinase-9 and GABAA. Exp. Ther. Med. 2019, 17, 3083–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, J.; Jiang, Y. Antiepileptic potential of matrine via regulation the levels of gamma-aminobutyric acid and glutamic acid in the brain. Int. J. Mol. Sci. 2013, 14, 23751–23761. [Google Scholar] [CrossRef] [Green Version]
- Zhen, J.L.; Chang, Y.N.; Qu, Z.Z.; Fu, T.; Liu, J.Q.; Wang, W.P. Luteolin rescues pentylenetetrazole-induced cognitive impairment in epileptic rats by reducing oxidative stress and activating PKA/CREB/BDNF signaling. Epilepsy Behav. 2016, 57, 177–184. [Google Scholar] [CrossRef]
- Tambe, R.; Patil, A.; Jain, P.; Sancheti, J.; Somani, G.; Sathaye, S. Assessment of luteolin isolated from Eclipta alba leaves in animal models of epilepsy. Pharm. Biol. 2017, 55, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Zhen, J.; Chang, Y.; Tao, F.U.; Zhenzhen, Q.U.; Liu, J.; Wang, W. Effects of luteolin on CaM-CaMPK signaling pathway in hippocampus in epileptic rats. Int. J. Tradit. Chin. Med. 2016, 38, 232–237. [Google Scholar]
- Sharma, P.; Kumari, A.; Gulati, A.; Krishnamurthy, S.; Hemalatha, S. Chrysin isolated from Pyrus pashia fruit ameliorates convulsions in experimental animals. Nutr. Neurosci. 2019, 22, 569–577. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, J.; Afzal, O.; Kazmi, I.; Al-Abbasi, F.A.; Altamimi, A.S.A.; Yang, Z. Neuroprotective role of chrysin-loaded poly (lactic-co-glycolic acid) nanoparticle against kindling-induced epilepsy through Nrf2/ARE/HO-1 pathway. J. Biochem. Mol. Toxicol. 2020, 35, e22634. [Google Scholar] [CrossRef]
- Hashemi, P.; Babaei, J.F.; Vazifekhah, S.; Nikbakht, F. Evaluation of the neuroprotective, anticonvulsant, and cognition-improvement effects of apigenin in temporal lobe epilepsy: Involvement of the mitochondrial apoptotic pathway. Iran. J. Basic Med. Sci. 2019, 22, 752. [Google Scholar]
- Zhang, Y.; Seeburg, D.P.; Pulli, B.; Wojtkiewicz, G.R.; Bure, L.; Atkinson, W.; Schob, S.; Iwamoto, Y.; Ali, M.; Zhang, W.; et al. Myeloperoxidase nuclear imaging for epileptogenesis. Radiology 2016, 278, 822–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.; Yuan, J.; Liu, Y.; Qin, Y.; Wang, X.; Gu, J.; Chen, G.; Zhang, B.; Liu, H.K.; Zhao, J.; et al. Epileptic brain fluorescent imaging reveals apigenin can relieve the myeloperoxidase-mediated oxidative stress and inhibit ferroptosis. Proc. Natl. Acad. Sci. USA 2020, 117, 10155–10164. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Sharma, S.; Singh, D. Apigenin reverses behavioural impairments and cognitive decline in kindled mice via CREB-BDNF upregulation in the hippocampus. Nutr. Neurosci. 2020, 23, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Kaur, T.; Goel, R.K. Ferulic Acid Supplementation for management of depression in epilepsy. Neurochem. Res. 2017, 42, 2940–2948. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.M.; Abbas, K.M.; Abulseoud, O.A.; El-Hussainy, E.M.A. Effects of ferulic acid on oxidative stress, heat shock protein 70, connexin 43, and monoamines in the hippocampus of pentylenetetrazole-kindled rats. Can. J. Physiol. Pharmacol. 2017, 95, 732–742. [Google Scholar] [CrossRef]
- Hassanzadeh, P.; Arbabi, E.; Atyabi, F.; Dinarvand, R. Ferulic acid exhibits antiepileptogenic effect and prevents oxidative stress and cognitive impairment in the kindling model of epilepsy. Life Sci. 2017, 179, 9–14. [Google Scholar] [CrossRef]
- Zhang, S.H.; Liu, D.; Hu, Q.; Zhu, J.; Wang, S.; Zhou, S. Ferulic acid ameliorates pentylenetetrazol-induced seizures by reducing neuron cell death. Epilepsy Res. 2019, 156, 106183. [Google Scholar] [CrossRef]
- Shakeel, S.; Rehman, M.U.; Tabassum, N.; Amin, U. Effect of naringenin (a naturally occurring flavanone) against pilocarpine-induced status epilepticus and oxidative stress in mice. Pharmacogn. Mag. 2017, 13 (Suppl. S1), S154–S160. [Google Scholar]
- Park, J.; Jeong, K.H.; Shin, W.H.; Bae, Y.S.; Jung, U.J.; Kim, S.R. Naringenin ameliorates kainic acid-induced morphological alterations in the dentate gyrus in a mouse model of temporal lobe epilepsy. Neuroreport 2016, 27, 1182–1189. [Google Scholar] [CrossRef]
- Khodayar, M.J.; Salehi, S.; Rezaei, M.; Siahpoosh, A.; Khazaei, A.; Houshmand, G. Evaluation of the effect of naringenin on pentylenetetrazole and maximal electroshock-induced convulsions in mice. Jundishapur J. Nat. Pharm. Prod. 2017, 12, e31384. [Google Scholar]
- Mert, D.G.; Turgut, N.H.; Arslanbas, E.; Gungor, H.; Kara, H. The influence of quercetin on recognition memory and brain oxidative damage in a ketamine model of schizophrenia. Psychiatry Clin. Psychopharmacol. 2019, 29, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hajizadeh, M.A.; Valizadegan, F. Antidepressant effects of quercetin and its nanocrystal on schizophrenia animal model with using forced swimming test. J. Anim. Res. 2017, 30, 365–376. [Google Scholar]
- Hosseiny, R.; Hajizadeh, M.A.; Zare, M.; Mermohammad, R.F. The effect of quercetin and its nanocrystalin on behavioral impairment induced by ketamine injection in an animal model of schizophrenia. Daneshvar Med. 2016, 23, 11–20. [Google Scholar]
- Schwartz, D.L. Quercetin as an Augmentation Agent in Schizophrenia. J. Clin. Psychopharmacol. 2016, 36, 282–283. [Google Scholar] [CrossRef]
- Samad, N.; Saleem, A.; Yasmin, F.; Shehzad, M.A. Quercetin protects against stress-induced anxiety- and depression-like behavior and improves memory in male mice. Physiol. Res. 2018, 67, 795–808. [Google Scholar] [CrossRef]
- Khan, K.; Najmi, A.K.; Akhtar, M. A natural phenolic compound quercetin showed the usefulness by targeting inflammatory, oxidative stress markers and augment 5-HT levels in one of the animal models of depression in mice. Drug Res. 2019, 69, 392–400. [Google Scholar] [CrossRef]
- Fang, K.; Li, H.R.; Chen, X.X.; Gao, X.R.; Huang, L.L.; Du, A.Q.; Jiang, C.; Li, H.; Ge, J.F. Quercetin Alleviates LPS-Induced Depression-Like Behavior in Rats via Regulating BDNF-Related Imbalance of Copine 6 and TREM1/2 in the Hippocampus and PFC. Front. Pharmacol. 2020, 10, 1544. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, G.; Cui, L.; Wang, Q. Myricetin attenuates depressant-like behavior in mice subjected to repeated restraint stress. Int. J. Mol. Sci. 2015, 16, 28377–28385. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.M.; Wang, G.L.; Ma, Z.G. Protective effects of myricetin on chronic stress-induced cognitive deficits. Neuroreport 2016, 27, 652–658. [Google Scholar] [CrossRef]
- Gao, W.; Wang, W.; Peng, Y.; Deng, Z. Antidepressive effects of kaempferol mediated by reduction of oxidative stress, proinflammatory cytokines and up-regulation of AKT/β-catenin cascade. Metab. Brain Dis. 2019, 34, 485–494. [Google Scholar] [CrossRef]
- Filho, C.B.; Jesse, C.R.; Donato, F.; Giacomeli, R.; Del Fabbro, L.; da Silva Antunes, M.; de Gomes, M.G.; Goes, A.T.; Boeira, S.P.; Prigol, M.; et al. Chronic unpredictable mild stress decreases BDNF and NGF levels and Na+, K+-ATPase activity in the hippocampus and prefrontal cortex of mice: Antidepressant effect of chrysin. Neuroscience 2015, 289, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Borges Filho, C.; Jesse, C.R.; Donato, F.; Del Fabbro, L.; de Gomes, M.G.; Goes, A.T.; Souza, L.C.; Boeira, S.P. Chrysin promotes attenuation of depressive-like behavior and hippocampal dysfunction resulting from olfactory bulbectomy in mice. Chem. Biol. Interact. 2016, 260, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Bansal, Y.; Singh, R.; Saroj, P.; Sodhi, R.K.; Kuhad, A. Naringenin protects against oxido-inflammatory aberrations and altered tryptophan metabolism in olfactory bulbectomized-mice model of depression. Toxicol. Appl. Pharmacol. 2018, 355, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Umukoro, S.; Kalejaye, H.A.; Ben-Azu, B.; Ajayi, A.M. Naringenin attenuates behavioral derangements induced by social defeat stress in mice via inhibition of acetylcholinesterase activity, oxidative stress and release of pro-inflammatory cytokines. Biomed. Pharmacother. 2018, 105, 714–723. [Google Scholar] [CrossRef]
- Tayyab, M.; Farheen, S.; Khanam, N.; Mobarak Hossain, M.; Shahi, M.H. Antidepressant and Neuroprotective Effects of Naringenin via Sonic Hedgehog-GLI1 Cell Signaling Pathway in a Rat Model of Chronic Unpredictable Mild Stress. Neuromolecular. Med. 2019, 21, 250–261. [Google Scholar] [CrossRef]
- Lee, S.; Kim, H.B.; Hwang, E.S.; Kim, E.S.; Kim, S.S.; Jeon, T.D.; Song, M.C.; Lee, J.S.; Chung, M.C.; Maeng, S.; et al. Antidepressant-like Effects of p-Coumaric Acid on LPS-induced Depressive and Inflammatory Changes in Rats. Exp. Neurobiol. 2018, 27, 189–199. [Google Scholar] [CrossRef]
- Zheng, X.; Cheng, Y.; Chen, Y.; Yue, Y.; Li, Y.; Xia, S.; Li, Y.; Deng, H.; Zhang, J.; Cao, Y. Ferulic acid improves depressive-like behavior in prenatally stressed offspring rats via anti-inflammatory activity and HPA axis. Int. J. Mol. Sci. 2019, 20, 493. [Google Scholar] [CrossRef] [Green Version]
- Bo, X.M.; Yu, R.B.; Du, S.J.; Zhang, R.L.; He, L. Ferulic acid alleviates lipopolysaccharide-induced depression-like behavior by inhibiting inflammation and apoptosis. Asian Pac. J. Trop. Biomed. 2020, 10, 523. [Google Scholar]
- Zhao, L.; Zhu, L.; Guo, X. Valproic acid attenuates Aβ25-35-induced neurotoxicity in PC12 cells through suppression of mitochondria-mediated apoptotic pathway. Biomed. Pharmacother. 2018, 106, 77–82. [Google Scholar] [CrossRef]
- Ahmad, L.; Mujahid, M.; Mishra, A.; Rahman, M.A. Protective role of hydroalcoholic extract of Cajanus cajan Linn leaves against memory impairment in sleep deprived experimental rats. J. Ayurveda. Integr. Med. 2020, 11, 471–477. [Google Scholar] [CrossRef]
- Zhang, L.; Xia, R.; Jia, J.; Wang, L.; Li, K.; Li, Y.; Zhang, J. Oleanolic acid protects against cognitive decline and neuroinflammation-mediated neurotoxicity by blocking secretory phospholipase A2 IIA-activated calcium signals. Mol. Immunol. 2018, 99, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.A.; Abo El-Ella, D.M.; El-Emam, S.Z.; Shahat, A.S.; El-Sayed, R.M. Physical & mental activities enhance the neuroprotective effect of vinpocetine & coenzyme Q10 combination against Alzheimer & bone remodeling in rats. Life Sci. 2019, 229, 21–35. [Google Scholar] [PubMed]
- Yuan, H.; Jiang, C.; Zhao, J.; Zhao, Y.; Zhang, Y.; Xu, Y.; Gao, X.; Guo, L.; Liu, Y.; Liu, K.; et al. Euxanthone Attenuates Aβ1-42-Induced Oxidative Stress and Apoptosis by Triggering Autophagy. J. Mol. Neurosci. 2018, 66, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Aminyavari, S.; Zahmatkesh, M.; Farahmandfar, M.; Khodagholi, F.; Dargahi, L.; Zarrindast, M.R. Protective role of Apelin-13 on amyloid β25–35-induced memory deficit; involvement of autophagy and apoptosis process. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 89, 322–334. [Google Scholar] [CrossRef]
- Aykac, A.; Ozbeyli, D.; Uncu, M.; Ertaş, B.; Kılınc, O.; Şen, A.; Orun, O.; Sener, G. Evaluation of the protective effect of Myrtus communis in scopolamine-induced Alzheimer model through cholinergic receptors. Gene 2019, 689, 194–201. [Google Scholar] [CrossRef]
- Ali, A.M.; Kunugi, H. Bee honey protects astrocytes against oxidative stress: A preliminary in vitro investigation. Neuropsychopharmacol. Rep. 2019, 39, 312–314. [Google Scholar] [CrossRef] [Green Version]
- Arranz, A.M.; De Strooper, B. The role of astroglia in Alzheimer’s disease: Pathophysiology and clinical implications. Lancet Neurol. 2019, 18, 406–414. [Google Scholar] [CrossRef]
- Baranowska-Wójcik, E.; Szwajgier, D.; Winiarska-Mieczan, A. Honey as the potential natural source of cholinesterase inhibitors in Alzheimer’s disease. Plant Foods Hum. Nutr. 2020, 75, 30–32. [Google Scholar] [CrossRef]
- Şahin, B. Can sunflower honey have a protective effect against Alzheimer’s disease? J. Ongoing Chem. Res. 2021, 6, 6–9. [Google Scholar]
- Shati, A.A.; Elsaid, F.G.; Hafez, E.E. Biochemical and molecular aspects of aluminium chloride-induced neurotoxicity in mice and the protective role of Crocus sativus L. extraction and honey syrup. Neuroscience 2011, 175, 66–74. [Google Scholar] [CrossRef]
- Saxena, A.K.; Phyu, H.P.; Al-Ani, I.M.; Talib, N.A. Potential protective effect of honey against chronic cerebral hypoperfusion-induced neurodegeneration in rats. J. Anat. Soc. India 2014, 63, 151–155. [Google Scholar] [CrossRef]
- Cheng, W.; Chen, W.; Wang, P.; Chu, J. Asiatic acid protects differentiated PC12 cells from Aβ25–35-induced apoptosis and tau hyperphosphorylation via regulating PI3K/Akt/GSK-3β signaling. Life Sci. 2018, 208, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Fawzy Fahim, V.; Wadie, W.; Shafik, A.N.; Ishak Attallah, M. Role of simvastatin and insulin in memory protection in a rat model of diabetes mellitus and dementia. Brain. Res. Bull. 2019, 144, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.P.; Hoppe, J.B.; Saccomori, A.B.; Dos Santos, B.G.; Sagini, J.P.; Crestani, M.S.; August, P.M.; Hözer, R.M.; Grings, M.; Parmeggiani, B.; et al. Physical exercise during pregnancy prevents cognitive impairment induced by amyloid-β in adult offspring rats. Mol. Neurobiol. 2019, 56, 2022–2038. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, P.; Zheng, Y.; Yang, C.; Du, T.; Ge, M.; Chang, X.; Duan, R.; Ma, G. Effects of NMDAR Antagonist on the Regulation of P-MARCKS Protein to Aβ1-42 Oligomers Induced Neurotoxicity. Neurochem. Res. 2018, 43, 2008–2015. [Google Scholar] [CrossRef]
- Altenhöfer, S.; Witte, I.; Teiber, J.F.; Wilgenbus, P.; Pautz, A.; Li, H.; Daiber, A.; Witan, H.; Clement, A.M.; Förstermann, U.; et al. One enzyme, two functions pon2 prevents mitochondrial superoxide formation and apoptosis independent from its lactonase activity. J. Biol. Chem. 2010, 285, 24398–24403. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, S.M.; Moreira, P.I.; Agostinho, P.; Pereira, C.; Oliveira, C.R. Neurodegenerative pathways in Parkinson’s disease: Therapeutic strategies. Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 405–419. [Google Scholar] [CrossRef] [Green Version]
- Schapira, A.H. Mitochondria in the aetiology and pathogenesis of Parkinson’s disease. Lancet Neurol. 2008, 7, 97–109. [Google Scholar] [CrossRef]
- Shim, J.S.; Kim, H.G.; Ju, M.S.; Choi, J.G.; Jeong, S.Y.; Oh, M.S. Effects of the hook of Uncaria rhynchophylla on neurotoxicity in the 6-hydroxydopamine model of Parkinson’s disease. J. Ethnopharmacol. 2009, 126, 361–365. [Google Scholar] [CrossRef]
- Chaudhuri, K.R.; Schapira, A.H. Non-motor symptoms of Parkinson’s disease: Dopaminergic pathophysiology and treatment. Lancet Neurol. 2009, 8, 464–474. [Google Scholar] [CrossRef]
- Martin, L.J. Mitochondrial and cell death mechanisms in neurodegenerative diseases. Pharmaceuticals 2010, 3, 839–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, F.; Vivacqua, G.; Yu, S. The role of α-synuclein in neurotransmission and synaptic plasticity. J. Chem. Neuroanat. 2011, 42, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef]
- In, S.; Hong, C.W.; Choi, B.; Jang, B.G.; Kim, M.J. Inhibition of mitochondrial clearance and Cu/Zn-SOD activity enhance 6-hydroxydopamine-induced neuronal apoptosis. Mol. Neurobiol. 2016, 53, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Sarrafchi, A.; Bahmani, M.; Shirzad, H.; Rafieian-Kopaei, M. Oxidative stress and Parkinson’s disease: New hopes in treatment with herbal antioxidants. Curr. Pharm. Des. 2016, 22, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, O.K.T.A.Y.; Karahalil, F.A.T.M.A.; Can, Z.; Sahin, H.; Kolayli, S.E.V.G.İ. Total monoamine oxidase (MAO) inhibition by chestnut honey, pollen and propolis. J. Enzym. Inhib. Med. Chem. 2014, 29, 690–694. [Google Scholar] [CrossRef]
- Topal, N.; Bulduk, I.; Mut, Z.; Bozoğlu, H.; Tosun, Y.K. Flowers, Pollen and Honey for Use in the Treatment of Parkinson’s Disease. Rev. Chim. 2020, 71, 308–319. [Google Scholar] [CrossRef]
- Vonsattel, J.P.; DiFiglia, M. Huntington disease. J. Neuropathol. Exp. Neurol. 1998, 57, 369–384. [Google Scholar] [CrossRef] [Green Version]
- Wexler, A.; Wild, E.J.; Tabrizi, S.J. George Huntington: A legacy of inquiry, empathy and hope. Brain 2016, 139 Pt 8, 2326–2333. [Google Scholar] [CrossRef] [Green Version]
- McColgan, P.; Tabrizi, S.J. Huntington’s disease: A clinical review. Eur. J. Neurol. 2018, 25, 24–34. [Google Scholar] [CrossRef]
- A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. The Huntington’s Disease Collaborative Research Group. Cell 1993, 72, 971–983.
- Jin, Y.N.; Johnson, G.V. The interrelationship between mitochondrial dysfunction and transcriptional dysregulation in Huntington disease. J. Bioenerg. Biomembr. 2010, 42, 199–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Es, M.A.; Hardiman, O.; Chio, A.; Al-Chalabi, A.; Pasterkamp, R.J.; Veldink, J.H.; van den Berg, L.H. Amyotrophic lateral sclerosis. Lancet 2017, 390, 2084–2098. [Google Scholar] [CrossRef]
- Hardiman, O.; Al-Chalabi, A.; Chio, A.; Corr, E.M.; Logroscino, G.; Robberecht, W.; Shaw, P.J.; Simmons, Z.; van den Berg, L.H. Amyotrophic lateral sclerosis. Nat. Rev. Dis. Primers. 2017, 3, 17085. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.H.; Al-Chalabi, A. Amyotrophic Lateral Sclerosis. N. Engl. J. Med. 2017, 377, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Bond, L.; Bernhardt, K.; Madria, P.; Sorrentino, K.; Scelsi, H.; Mitchell, C.S. A Metadata Analysis of Oxidative Stress Etiology in Preclinical Amyotrophic Lateral Sclerosis: Benefits of Antioxidant Therapy. Front. Neurosci. 2018, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Massey, L.K.; Mah, A.L.; Ford, D.L.; Miller, J.; Liang, J.; Doong, H.; Monteiro, M.J. Overexpression of ubiquilin decreases ubiquitination and degradation of presenilin proteins. J. Alzheimer’s Dis. 2004, 6, 79–92. [Google Scholar] [CrossRef]
- Phokasem, P.; Jantrapirom, S.; Karinchai, J.; Yoshida, H.; Yamaguchi, M.; Chantawannakul, P. Honeybee products and edible insect powders improve locomotive and learning abilities of Ubiquilin-knockdown Drosophila. BMC Complement. Med. Ther. 2020, 20, 267. [Google Scholar] [CrossRef]
- Thijs, R.D.; Surges, R.; O’Brien, T.J.; Sander, J.W. Epilepsy in adults. Lancet 2019, 393, 689–701. [Google Scholar] [CrossRef]
- Beghi, E. The Epidemiology of Epilepsy. Neuroepidemiology 2020, 54, 185–191. [Google Scholar] [CrossRef]
- Hashemian, M.; Anissian, D.; Ghasemi-Kasman, M.; Akbari, A.; Khalili-Fomeshi, M.; Ghasemi, S.; Ahmadi, F.; Moghadamnia, A.A.; Ebrahimpour, A. Curcumin-loaded chitosan-alginate-STPP nanoparticles ameliorate memory deficits and reduce glial activation in pentylenetetrazol-induced kindling model of epilepsy. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Nieoczym, D.; Socała, K.; Raszewski, G.; Wlaź, P. Effect of quercetin and rutin in some acute seizure models in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 54, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Garlich, F.M.; Balakrishnan, K.; Shah, S.K.; Howland, M.A.; Fong, J.; Nelson, L.S. Prolonged altered mental status and bradycardia following pediatric donepezil ingestion. Clin. Toxicol. 2014, 52, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Vaidya, V.A. Cyclic AMP response element binding protein and brain-derived neurotrophic factor: Molecules that modulate our mood? J. Biosci. 2006, 31, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.G.; Jin, S.L.; Li, G.Y.; Li, Q.Q.; Li, Z.R.; Ma, H.X.; Zhuo, C.J.; Jiang, R.H.; Ye, M.J. Serotonin regulates brain-derived neurotrophic factor expression in select brain regions during acute psychological stress. Neural Regen. Res. 2016, 11, 1471. [Google Scholar]
- Yu, L.; Zhang, Y.; Ma, R.; Bao, L.; Fang, J.; Yu, T. Potent protection of ferulic acid against excitotoxic effects of maternal intragastric administration of monosodium glutamate at a late stage of pregnancy on developing mouse fetal brain. Eur. Neuropsychopharmacol. 2006, 16, 170–177. [Google Scholar] [CrossRef]
- Xie, W.; Cai, L.; Yu, Y.; Gao, L.; Xiao, L.; He, Q.; Ren, Z.; Liu, Y. Activation of brain indoleamine 2,3-dioxygenase contributes to epilepsy-associated depressive-like behavior in rats with chronic temporal lobe epilepsy. J. Neuroinflamm. 2014, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Waldbaum, S.; Patel, M. Mitochondrial dysfunction and oxidative stress: A contributing link to acquired epilepsy? J. Bioenerg. Biomembr. 2010, 42, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Na, M.; Liu, Y.; Shi, C.; Gao, W.; Ge, H.; Wang, Y.; Wang, H.; Long, Y.; Shen, H.; Shi, C.; et al. Prognostic value of CA4/DG volumetry with 3 T magnetic resonance imaging on postoperative outcome of epilepsy patients with dentate gyrus pathology. Epilepsy Res. 2014, 108, 1315–1325. [Google Scholar] [CrossRef]
- Schultz, S.H.; North, S.W.; Shields, C.G. Schizophrenia: A review. Am. Fam. Physician. 2007, 75, 1821–1829. [Google Scholar]
- Lewis, D.A.; Lieberman, J.A. Catching up on schizophrenia: Natural history and neurobiology. Neuron 2000, 28, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, P.V.; Dean, O.; Andreazza, A.C.; Berk, M.; Kapczinski, F. Antioxidant treatments for schizophrenia. Cochrane Database Syst. Rev. 2016, 10, CD008919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahadik, S.P.; Mukherjee, S. Free radical pathology and antioxidant defense in schizophrenia: A review. Schizophr. Res. 1996, 19, 1–17. [Google Scholar] [CrossRef]
- Yahaya, R.; Zahary, M.N.; Othman, Z.; Ismail, R.; Him, N.A.S.N.; Abd Aziz, A.; Dahlan, R.; Jusoh, A.F. Tualang honey supplementation as cognitive enhancer in patients with schizophrenia. Heliyon 2020, 6, e03948. [Google Scholar] [CrossRef] [PubMed]
- Perez-Caballero, L.; Torres-Sanchez, S.; Romero-López-Alberca, C.; González-Saiz, F.; Mico, J.A.; Berrocoso, E. Monoaminergic system and depression. Cell Tissue Res. 2019, 377, 107–113. [Google Scholar] [CrossRef]
- Racagni, G.; Popoli, M. The pharmacological properties of antidepressants. Int. Clin. Psychopharmacol. 2010, 25, 117–131. [Google Scholar] [CrossRef]
- Schildkraut, J.J.; Kety, S.S. Biogenic amines and emotion. Science 1967, 156, 21–37. [Google Scholar] [CrossRef]
- Dell’Osso, L.; Carmassi, C.; Mucci, F.; Marazziti, D. Depression, Serotonin and Tryptophan. Curr. Pharm. Des. 2016, 22, 949–954. [Google Scholar] [CrossRef]
- Nautiyal, K.M.; Hen, R. Serotonin receptors in depression: From A to B. F1000Research 2017, 6, 123. [Google Scholar] [CrossRef] [Green Version]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S.; Deyama, S.; Fogaça, M.V. Role of BDNF in the pathophysiology and treatment of depression: Activity-dependent effects distinguish rapid-acting antidepressants. Eur. J. Neurosci. 2019, 53, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro) degenerative processes in that illness. Prog. NeuroPsychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar] [PubMed]
- Bakunina, N.; Pariante, C.M.; Zunszain, P.A. Immune mechanisms linked to depression via oxidative stress and neuroprogression. Immunology 2015, 144, 365–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindqvist, D.; Dhabhar, F.S.; James, S.J.; Hough, C.M.; Jain, F.A.; Bersani, F.S.; Reus, V.I.; Verhoeven, J.E.; Epel, E.S.; Mahan, L.; et al. Oxidative stress, inflammation and treatment response in major depression. Psychoneuroendocrinology 2017, 76, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Liu, H.; Suzuki, K.; Ma, S.; Liu, C. Linking What We Eat to Our Mood: A Review of Diet, Dietary Antioxidants, and Depression. Antioxidants 2019, 8, 376. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, S.; Nagappa, A.N.; Patil, C.R. Role of oxidative stress in depression. Drug Discov. Today 2020, 25, 1270–1276. [Google Scholar] [CrossRef]
- Sheas, M.N.; Rasool, H.; Rafique, M.N.; Tariq, M.R.; Muhammad, A.; Ali, K. Exploring the Potential of Honey and Curcumin as Antidepressent. Punjab Univ. J. Zool. 2019, 34, 89–95. [Google Scholar] [CrossRef]


{kind=link}
| Active Ingredient | Honey Type | Quantification Method | ||||||
|---|---|---|---|---|---|---|---|---|
| Manuka [15,16,17] | Acacia [18,19] | Eucalyptus [20] | Chestnut [21] | Cedar [22] | Sunflower [23] | Clover [24] | ||
| Quercetin | 43 | 200 | 105 | 46 | - | 200 | 200 | HPLC |
| Myricetin | 70 | - | 780 | 138 | - | - | - | UHPLC |
| Galangin | 35 | 550 | - | 149 | - | 220 | - | UPLC |
| Apigenin | 40 | 290 | 140 | 100 | 470 | 90 | 260 | HPLC |
| Naringenin | - | 60 | - | 350 | 107 | 169 | 368 | RP-HPLC |
| Chrysin | 30 | 130 | 190 | 680 | 170 | - | 470 | UHPLC |
| Luteolin | 38 | 140 | 66 | 80 | 464 | 320 | 550 | HPLC-DAD |
| Pinocembrin | 180 | 640 | 142 | - | - | 440 | - | HPLC |
| Pinobanksin | 290 | 148 | 319 | - | - | - | - | HPLC |
| Isorhamnetin | 40 | 90 | - | - | - | - | - | HPLC |
| Kaempferol | 150 | 90 | 147 | 250 | 670 | 120 | 172 | HPLC-UV |
| Gallic acid | 300 | 101 | 531 | 610 | 354 | 160 | 760 | HPLC-UV |
| Caffeic acid | 50 | 110 | 900 | 997 | 255 | 350 | 772 | HPLC-UV |
| Coumaric acid | 103 | 170 | 101 | 786 | 915 | 110 | 147 | HPLC-UV |
| Chlorogenic acid | 60 | 40 | 284 | 552 | - | 360 | - | HPLC-DAD |
| Ferulic acid | - | 740 | 368 | 166 | 220 | - | 531 | HPLC-PDA |
| Syringic acid | 400 | 300 | 366 | 202 | 394 | - | 235 | HPLC-UV |
| Cinnamic acid | - | 50 | - | 450 | 60 | - | 570 | HPLC-UV |
| Benzoic acid | 51 | - | 870 | 150 | - | - | - | HPLC |
| Total Active Ingredients | 1880 | 3849 | 5309 | 5706 | 3476 | 2539 | 5035 | - |
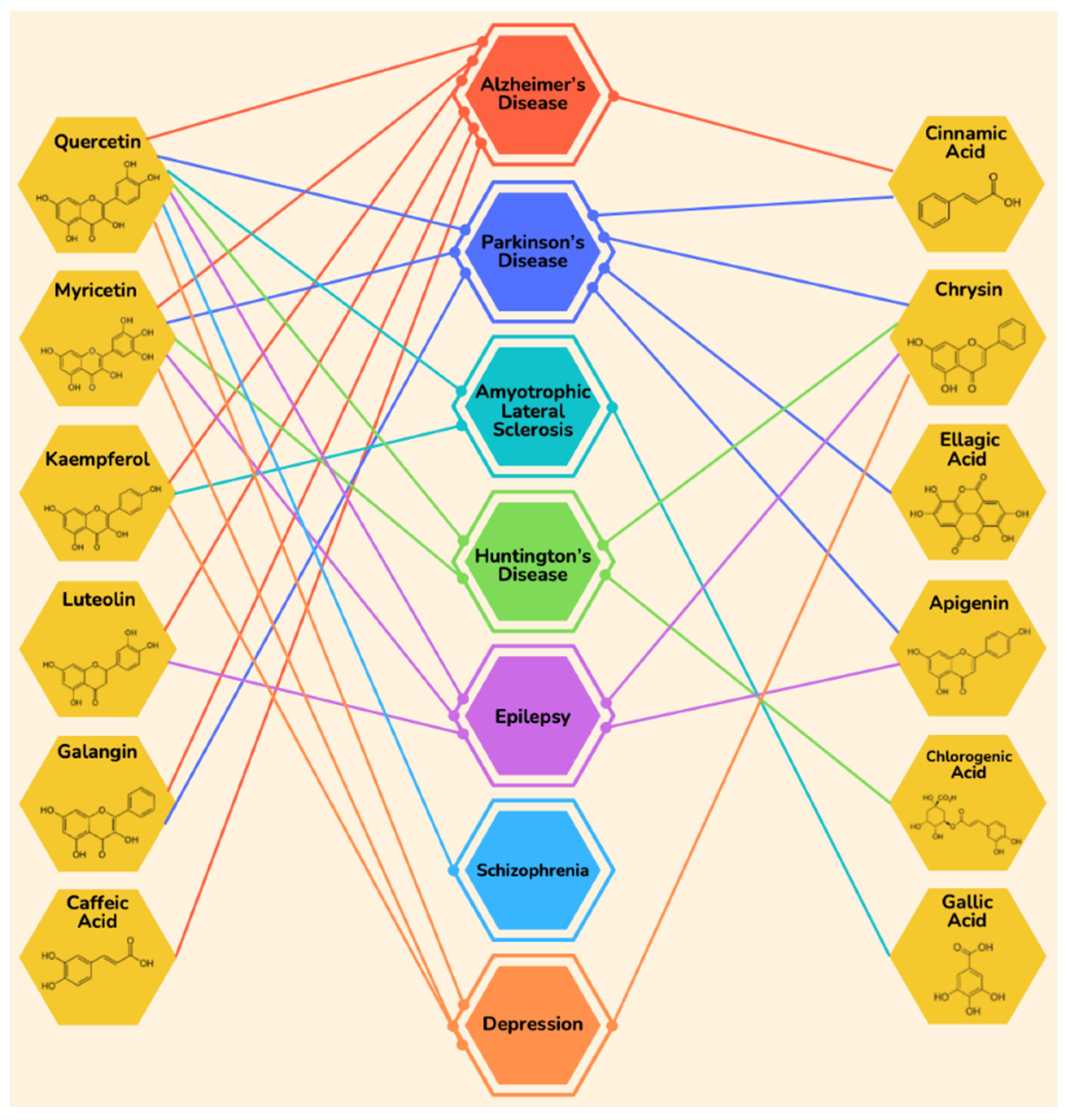
| Disease | Component | Effect | References |
|---|---|---|---|
| Alzheimer’s Disease | Quercetin | Improves mitochondrial activity | [89] |
| Activates Nrf-2 signaling | [90] | ||
| Deceases oxidative stress | |||
| Reduces oxidative stress via PON2 activity | [91] | ||
| Reduces neuroinflammation | [92,93] | ||
| Decreases astrogliosis | |||
| Prevents neurodegeneration | |||
| Recovers cognitive disabilities | |||
| Myricetin | Improves memory and cognitive function | [94] | |
| Prevents the formation of fibrils as well as oligomers of Aβ | [95] | ||
| Anti-tau protein effect | |||
| ↓ Acetylcholinesterase | [96] | ||
| ↑ Acetylcholine | |||
| Exhibits anti-inflammatory activity | [97,98] | ||
| Inhibition of the NF-κB pathway and NLRP3 inflammasome | |||
| Prevents the formation of fibrils as well as oligomers of Aβ | [99] | ||
| Kaempferol | Decreases lipid peroxidation and senile plaque formation | [100] | |
| Protects from apoptotic damage | |||
| Regulates concentrations of antioxidative enzymes | [101] | ||
| ↓ TNF-α and inhibits inflammation | |||
| Exhibits anti-apoptotic activity | [102] | ||
| Downregulates Bcl-2-associated X protein (Bax) and cleaved caspase-9 | |||
| Increases neuronal density in the hippocampus | [103] | ||
| Downregulates inflammatory proteins | [104] | ||
| Protects from oxidative damage | [105] | ||
| Luteolin | Alleviates oxidative stress | [106] | |
| Scavenges free radicals | |||
| Downregulates inflammatory and apoptotic proteins | |||
| Improves memory deficits | [107] | ||
| Maintains neuronal density | |||
| Galangin | Diminishes autophagy | [108] | |
| Decreases levels of p-tau, β-secretase, and Aβ42 | |||
| ↑ Acetylcholine | [109] | ||
| Improves cognitive functions | |||
| Caffeic acid | ↓ Acetylcholinesterase | [110] | |
| ↓ Inflammation | |||
| ↓ Oxidative stress | |||
| Cinnamic acid | Attenuates the formation of amyloid plaques | [111] | |
| Activates PPARα | |||
| Improves memory functions | |||
| Parkinson’s Disease | Quercetin | Protects from degeneration | [112] |
| ↑ Dopamine levels | |||
| ↑ Brain-Derived Neurotrophic Factor expression | |||
| ↑ Energy production of mitochondria | |||
| Restores activities of antioxidant enzymes | [113] | ||
| Repairs cognitive deficits | |||
| ↓ Endoplasmic reticulum stress | [114] | ||
| ↓ C/EBP homologous protein (CHOP) | |||
| ↑ Autophagy | |||
| ↑ Beclin-1 expression | |||
| Myricetin | ↓ Degeneration | [115] | |
| ↑ Dopamine levels | |||
| Inhibits inflammation | [116] | ||
| Prevents the activation of microglia | |||
| ↓ Expression of inflammatory cytokines | |||
| Chrysin | ↑ Expression of a survival factor MEF2D | [117] | |
| Inhibits MAO-B | |||
| Alleviates oxidative stress | [118] | ||
| Prevents inflammation and dysfunction of Na+, K+-ATPase pump | |||
| Ameliorates neuronal loss | [119] | ||
| Improves memory | |||
| Ellagic acid | Inhibits MAO activity | [120] | |
| Prevents the loss of neurons | |||
| ↓ Oxidative stress | |||
| Modulates levels of antioxidant enzymes | [121] | ||
| Protects from oxidative insult | |||
| Diminishes apoptosis | [122] | ||
| Inhibits MAO-B | |||
| Modulates ERβ/Nrf2/HO-1 signaling cascade | |||
| Cinnamic acid | Protects neurodegeneration | [123] | |
| Activates Peroxisome Proliferator Activating Receptor α (PPARα) | |||
| Galangin | Inhibits microglial activation | [124] | |
| Suppresses inflammatory factors | |||
| Activates PPAR-γ | [125,126,127] | ||
| Inhibits inflammation | |||
| Prevents apoptosis | |||
| Protects from oxidative damage | |||
| Apigenin | Exhibits antioxidative function | [128] | |
| Inhibits MAO | |||
| Prevents apoptosis | |||
| Inhibits caspase-3 activation | |||
| Huntington’s Disease | Quercetin | Improves motor functions | [129] |
| Regulates peroxisome proliferator-activated receptor gamma, coactivator (PGC-1), or sirtuins (SIRT1) | |||
| ↑ Energy production of mitochondria | |||
| Ameliorates behavioral malfunctions | [130] | ||
| Exhibits anxiolytic effect | |||
| Myricetin | Reduces aggregation of polyglutamine | [131] | |
| ↓ Proteo toxicity | [132] | ||
| Repairs behavioral changes | |||
| Chrysin | Upregulates anti-apoptotic factor | [133] | |
| Downregulates pro-apoptotic factor | |||
| Restores neurobehavioral functions | [134] | ||
| ↑ Serotonin | |||
| Chlorogenic acid | Protects from genotoxicity | [135] | |
| Amyotrophic Lateral Sclerosis | Quercetin | Inhibits aggregation of Cu-Zn superoxide dismutase (SOD) | [136,137] |
| Kaempferol | Prevents cell death | [138,139] | |
| Reduces aggregation of superoxide dismutase 1 (SOD1) | |||
| ↑ AMPK phosphorylation | |||
| Inhibits mTOR phosphorylation | |||
| Boosts up autophagy | |||
| Coumaric acid | Ameliorates oxidative stress and endoplasmic reticulum stress | [140] | |
| ↑ Autophagy | |||
| Gallic acid | ↑ Levels of antioxidant enzymes | [141] | |
| ↓ Lipid peroxidation | |||
| Downregulates inflammatory cytokines such as TNF-α, IL-6, IL-β, and NF-κB | |||
| Improves motor functions | |||
| Prevents glutamate excitotoxicity | [142] | ||
| Inhibits the formation of neurofibrillary tangles | |||
| Improves motor skills | |||
| ↓ TDP-43 proteotoxicity | [143] | ||
| Attenuates seizures | |||
| Epilepsy | Quercetin | Inhibits activation of microglial cells and inflammatory cytokines | [144] |
| Attenuates neurodegeneration | |||
| Inhibits expression of the gene for GABA receptors | [145] | ||
| Reduces depression | |||
| Restores tryptophan levels | [146] | ||
| Exerts anticonvulsant effect | |||
| Modulates Glycinergic and GABAergic ion channels | [147] | ||
| Reduces behavioral signs of seizures | |||
| ↓ Neuronal loss | [148] | ||
| ↓ Astrocyte activation | |||
| Myricetin | Ameliorates intensity of seizures | [149,150] | |
| Inhibits apoptosis | |||
| Downregulates Bad, Bax, and cleaved caspase 3 | |||
| Upregulates Bcl-2 and Bcl-xL | |||
| Normalizes glutamate/GABA | |||
| Luteolin | Improves cognitive deficits | [151] | |
| ↑ Expression of Brain-Derived Neurotrophic Factor (BDNF) | |||
| Exhibits anticonvulsant effect | [152] | ||
| ↑ Activation of receptors for GABAA | |||
| Reduces oxidative stress | |||
| Improves cognition impairments | [153] | ||
| Modulates CaM-CaMPK signaling pathway | |||
| Attenuates seizures | |||
| Chrysin | Modulates GABAA receptors | [154] | |
| Abrogates convulsion-induced oxidative damage | |||
| ↓ The severity of epileptic seizures | [155] | ||
| ↓ Apoptosis | |||
| Boosts the expression of Nrf2, NQO-1, and HO-1 | |||
| Apigenin | Reduces neuronal loss | [156] | |
| ↓ Release of cytochrome c from mitochondria | |||
| Alleviates apoptosis | |||
| Inhibits overexpression of hypochlorite (HClO) | [157,158] | ||
| ↓ Oxidative damage | |||
| Averts cognitive impairments | [159] | ||
| Exerts antidepressant and anti-anxiolytic effects | |||
| ↑ Expression of Brain-Derived Neurotrophic Factor (BDNF) | |||
| Ferulic acid | Palliates depression | [160] | |
| ↓ Levels of proinflammatory cytokines such as TNF-α and IL-1β | |||
| Inhibits Cyclooxygenase 2 (COX2) activity | |||
| Modulates corticosterone levels | |||
| Ameliorates oxidative stress | [161] | ||
| Upregulates neuroprotective Heat shock protein 70 (Hsp70) and neurotransmitters such as Serotonin (5-HT) and norepinephrine | |||
| Diminishes oxidative stress | [162] | ||
| Repairs cognitive deficits and seizure activity | |||
| Improves memory functions and learning capacities | [163] | ||
| Inhibits apoptotic process | |||
| Scavenges free radicles | |||
| Naringenin | ↑ Antioxidant enzymes | [164] | |
| Restores neuronal morphology | |||
| ↓ Neurodegeneration | |||
| Impedes occurrence of seizures | [165] | ||
| ↓ Granule cell disruption (GCD) in the hippocampus | |||
| Ameliorates generation of proinflammatory cytokines | |||
| Exerts anticonvulsant effect | [166] | ||
| Agonist effect on GABAA receptors | |||
| ↓ Glutamate transmission | |||
| Schizophrenia | Quercetin | Scavenges free radicals | [167] |
| ↑ Levels of antioxidant enzymes | |||
| Reduces depressive behaviors | [168] | ||
| Improves behavioral impairments | [169] | ||
| Boosts up antipsychotic therapy | [170] | ||
| Depression | Quercetin | ↑ Levels of antioxidant enzymes | [171,172] |
| ↓ Decrease levels of inflammatory cytokines | |||
| Accrues serotonin level | |||
| Mitigates depressive behaviors | [173] | ||
| Modulates levels of BDNF | |||
| Myricetin | Recovers hopeless behaviors | [174,175] | |
| Regulates BDNF levels | |||
| Exerts antioxidant effect | |||
| Kaempferol | Exert anti-depressive effects | [176] | |
| ↑ Levels of antioxidant enzymes | |||
| Upregulates AKT/β-catenin cascade | |||
| Chrysin | Upregulates nerve growth factor (NGF) and Brain-Derived Neurotrophic Factor (BDNF) | [177] | |
| Normalizes Na+, K+-ATPase activity | |||
| Employs anti-depressant effects | [178] | ||
| Inhibits kynurenine pathway | |||
| Elevates the levels of serotonin (5HT) | |||
| Naringenin | Performs anti-depressive activity | [179] | |
| Restores antioxidant enzymes’ levels | |||
| Modulates serotonin levels | |||
| Downregulates inflammation mediators | |||
| Inhibits acetylcholinesterase activity | [180] | ||
| Mitigates oxidative damage | |||
| Upregulates BDNF, Sonic Hedgehog (Shh) signaling, NKX2.2, and PAX6 | [181] | ||
| Coumaric acid | Improves behavioral hopelessness | [182] | |
| Lessens inflammation-associated alterations | |||
| Enhances neurotrophic activity | |||
| Ferulic acid | Abrogates depression-like behaviors | [183] | |
| Downregulates pro-inflammatory cytokines | |||
| Downregulates factors associated with inflammation and apoptosis | [184] | ||
| Mitigates depression |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iftikhar, A.; Nausheen, R.; Muzaffar, H.; Naeem, M.A.; Farooq, M.; Khurshid, M.; Almatroudi, A.; Alrumaihi, F.; Allemailem, K.S.; Anwar, H. Potential Therapeutic Benefits of Honey in Neurological Disorders: The Role of Polyphenols. Molecules 2022, 27, 3297. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103297
Iftikhar A, Nausheen R, Muzaffar H, Naeem MA, Farooq M, Khurshid M, Almatroudi A, Alrumaihi F, Allemailem KS, Anwar H. Potential Therapeutic Benefits of Honey in Neurological Disorders: The Role of Polyphenols. Molecules. 2022; 27(10):3297. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103297
Chicago/Turabian StyleIftikhar, Arslan, Rimsha Nausheen, Humaira Muzaffar, Muhammad Ahsan Naeem, Muhammad Farooq, Mohsin Khurshid, Ahmad Almatroudi, Faris Alrumaihi, Khaled S. Allemailem, and Haseeb Anwar. 2022. "Potential Therapeutic Benefits of Honey in Neurological Disorders: The Role of Polyphenols" Molecules 27, no. 10: 3297. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103297