
Interruptin C, a Radioprotective Agent, Derived from Cyclosorus terminans Protect Normal Breast MCF-10A and Human Keratinocyte HaCaT Cells against Radiation-Induced Damage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
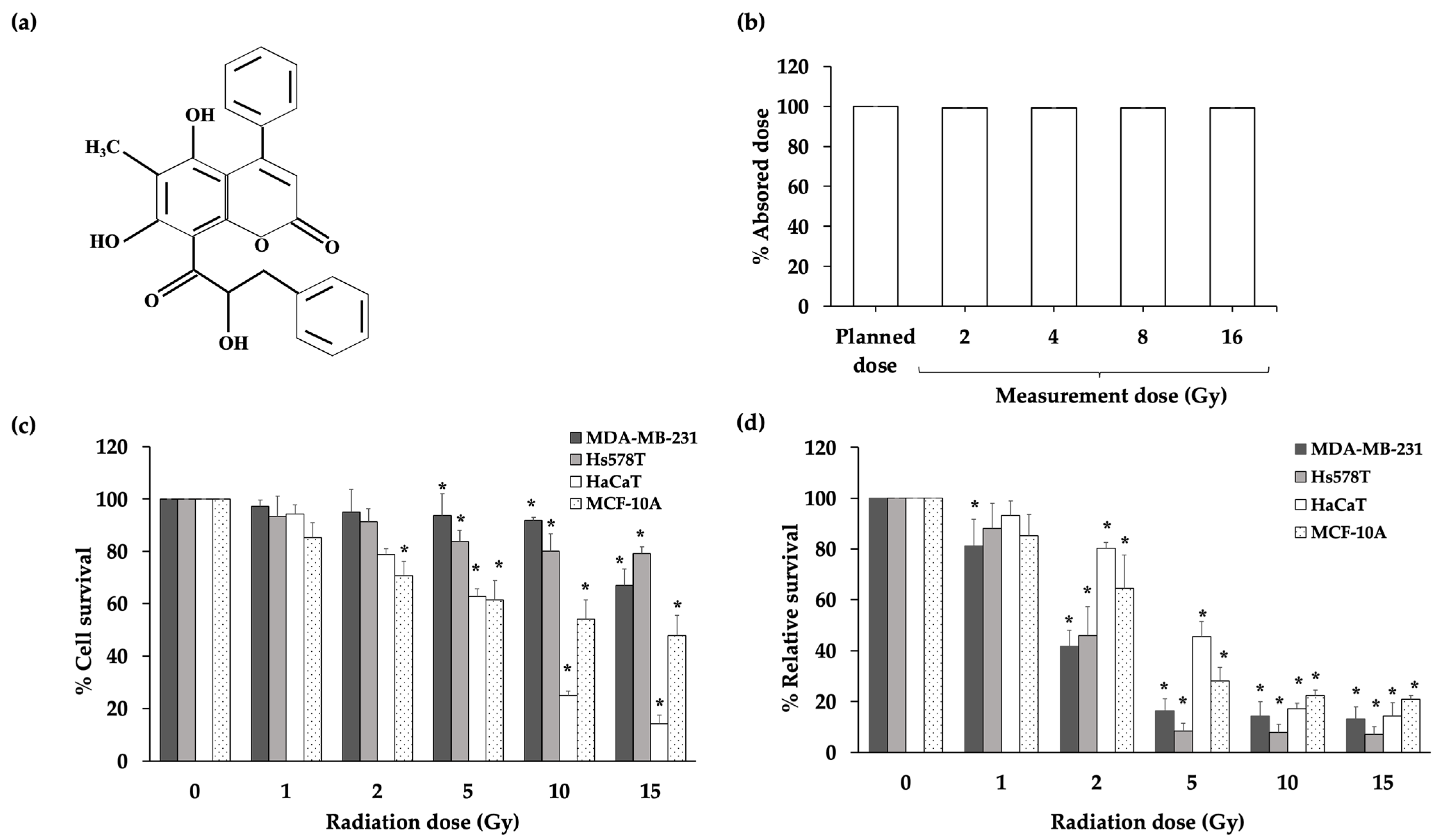
2. Results
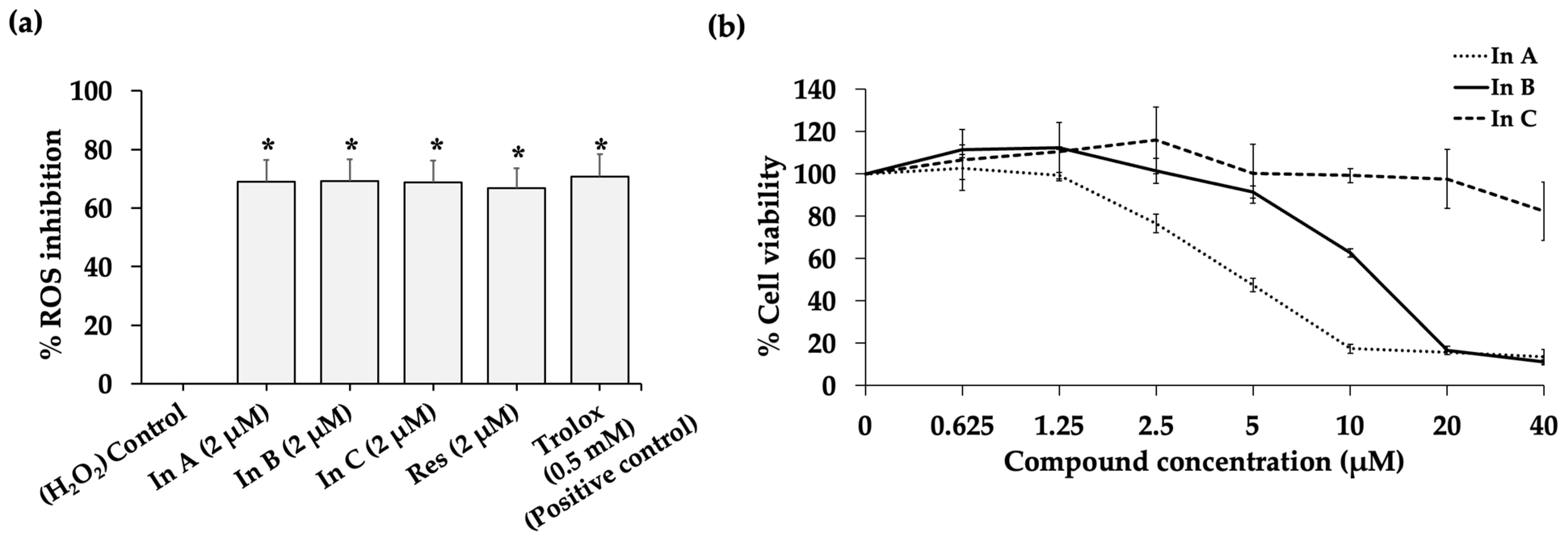
2.1. Interruptin C Showed Free Radical Scavenging Activity and Low Toxicity
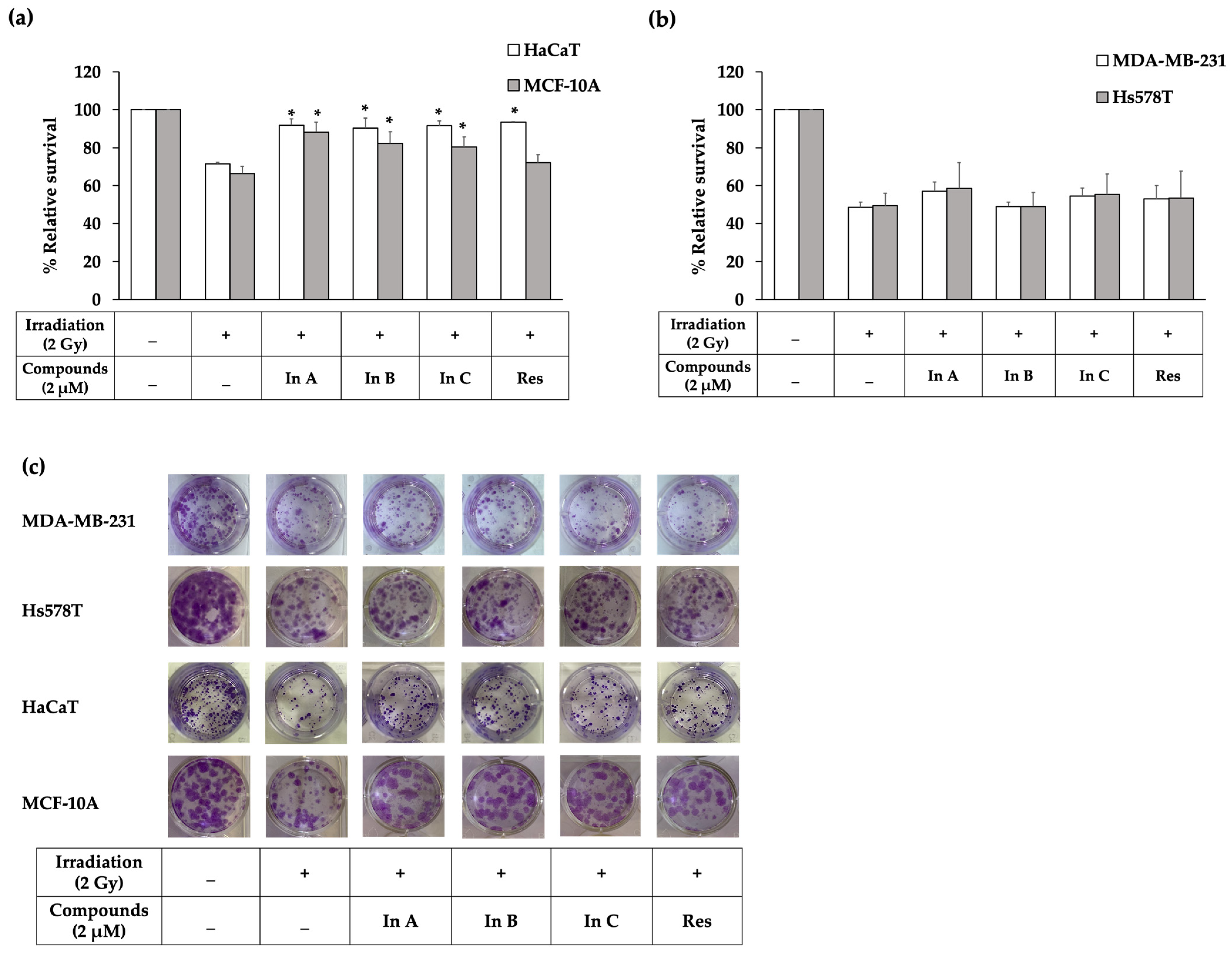
2.2. Interruptins Increased Survival of Normal Cells and Did Not Affect Proliferation of Cancer Cells
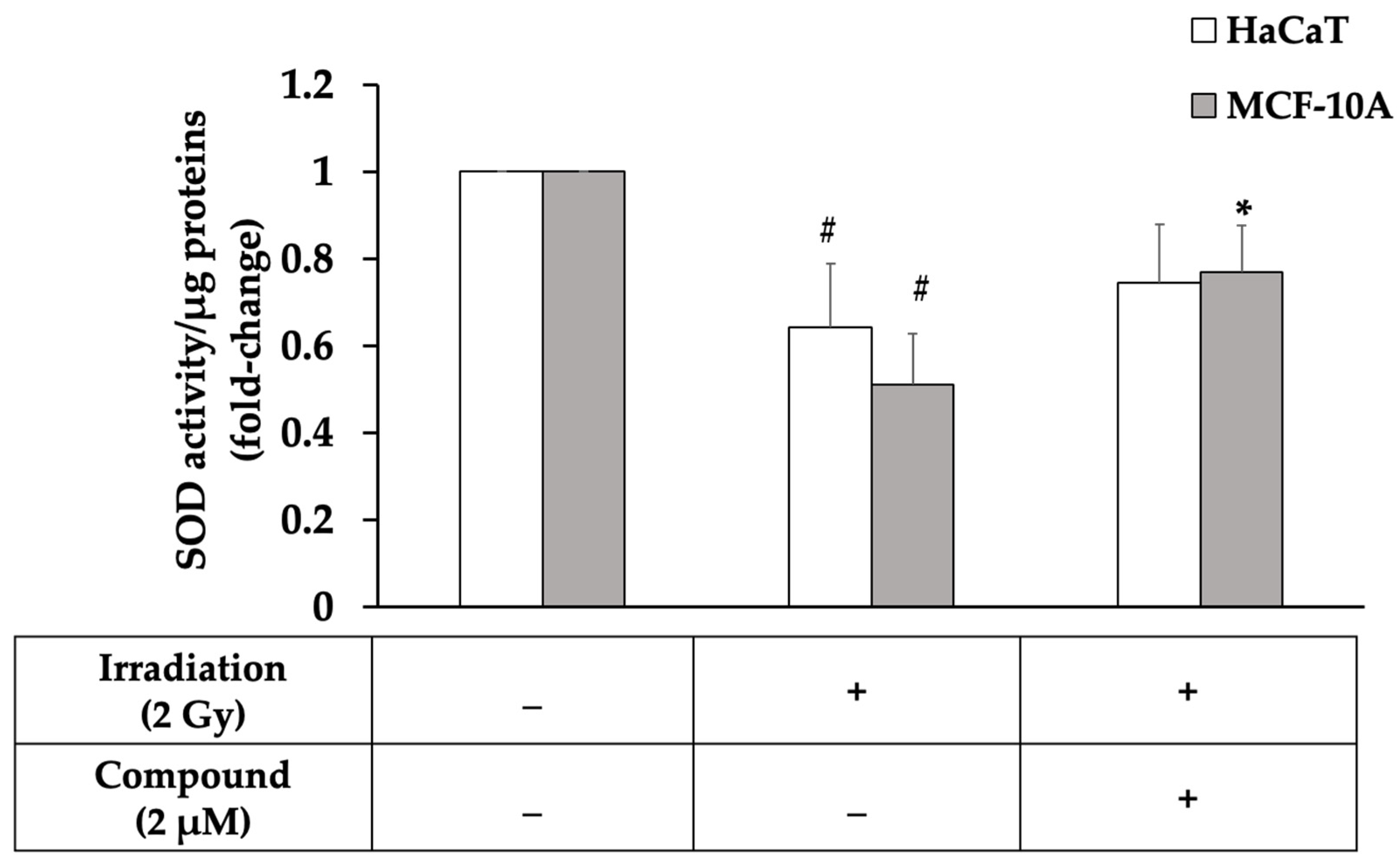
2.3. Oxidative Stress from Radiation Exposure Was Reduced by Pretreatment with Interruptin C
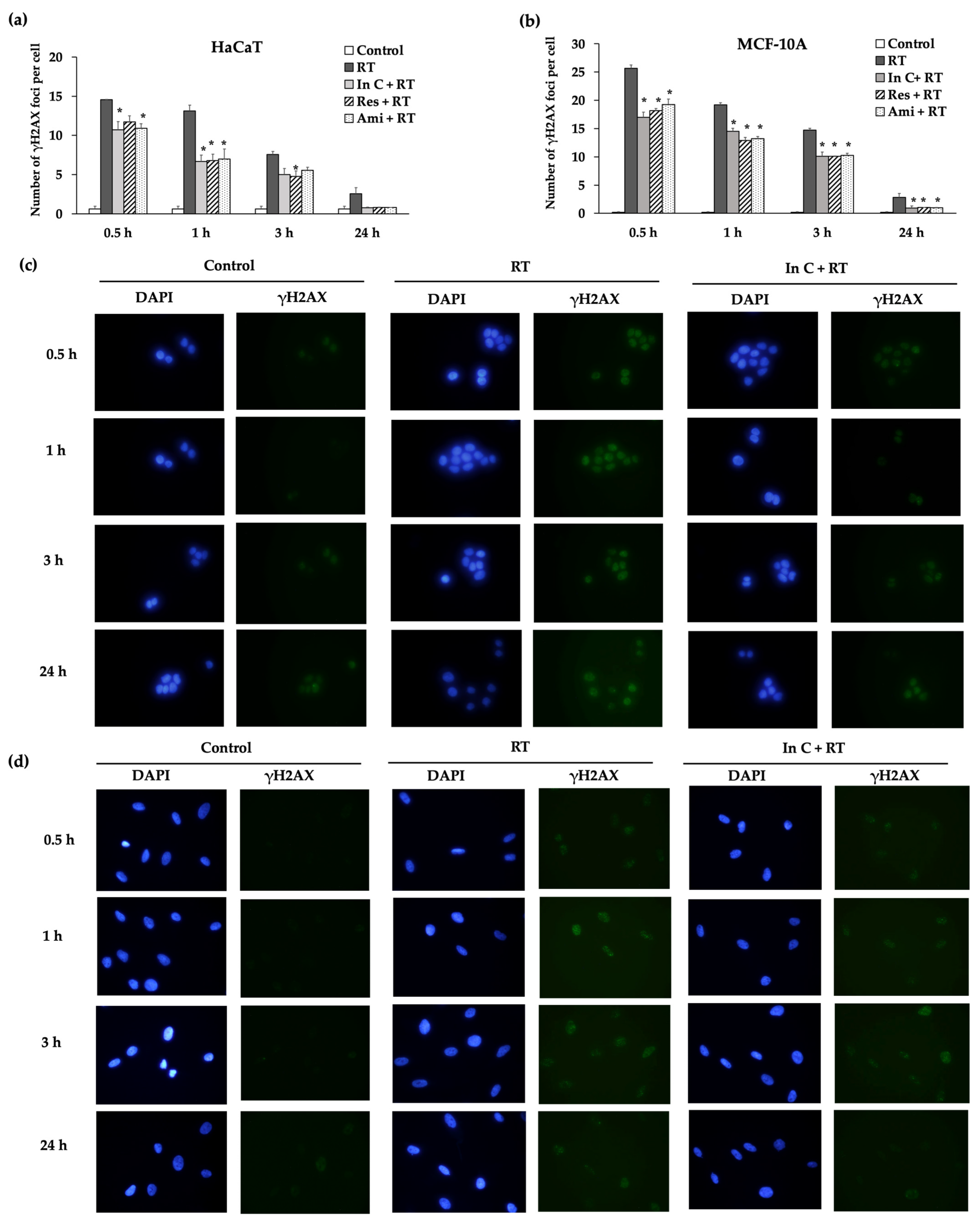
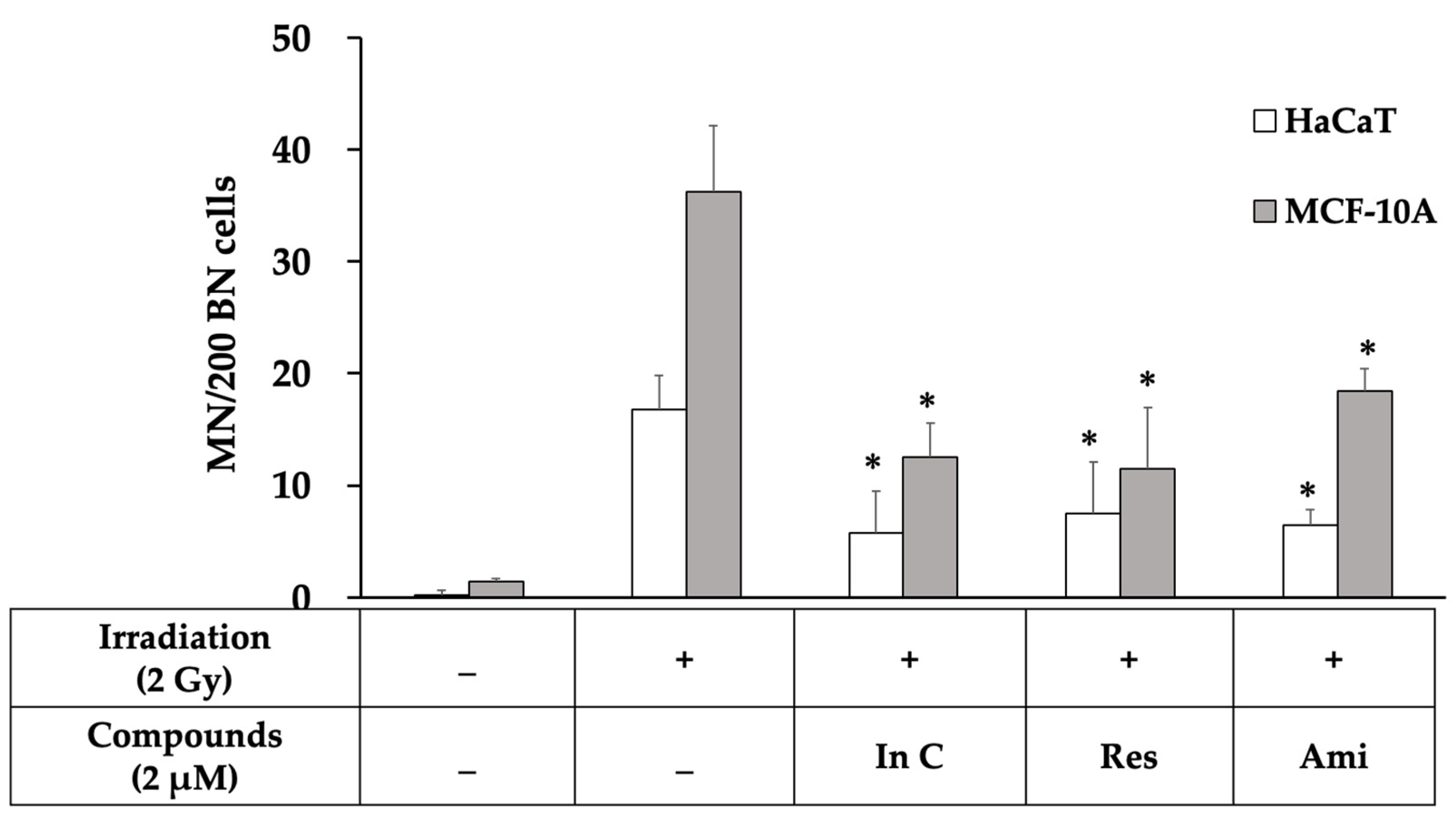
2.4. Interruptin C Decreased DNA Damage after Irradiation
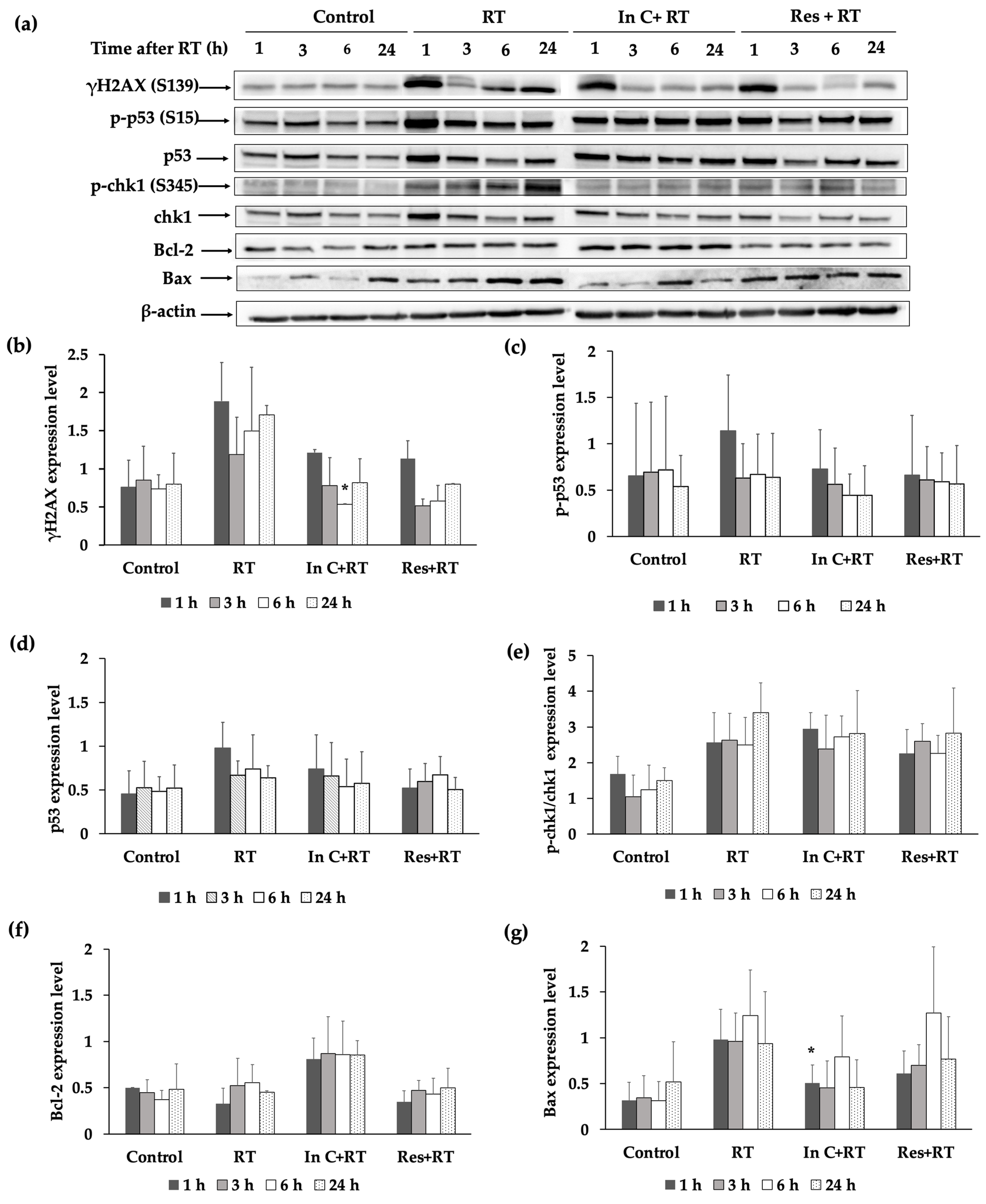
2.5. Effect of Interruptin C on DNA Damage Response in Irradiated Normal Cells
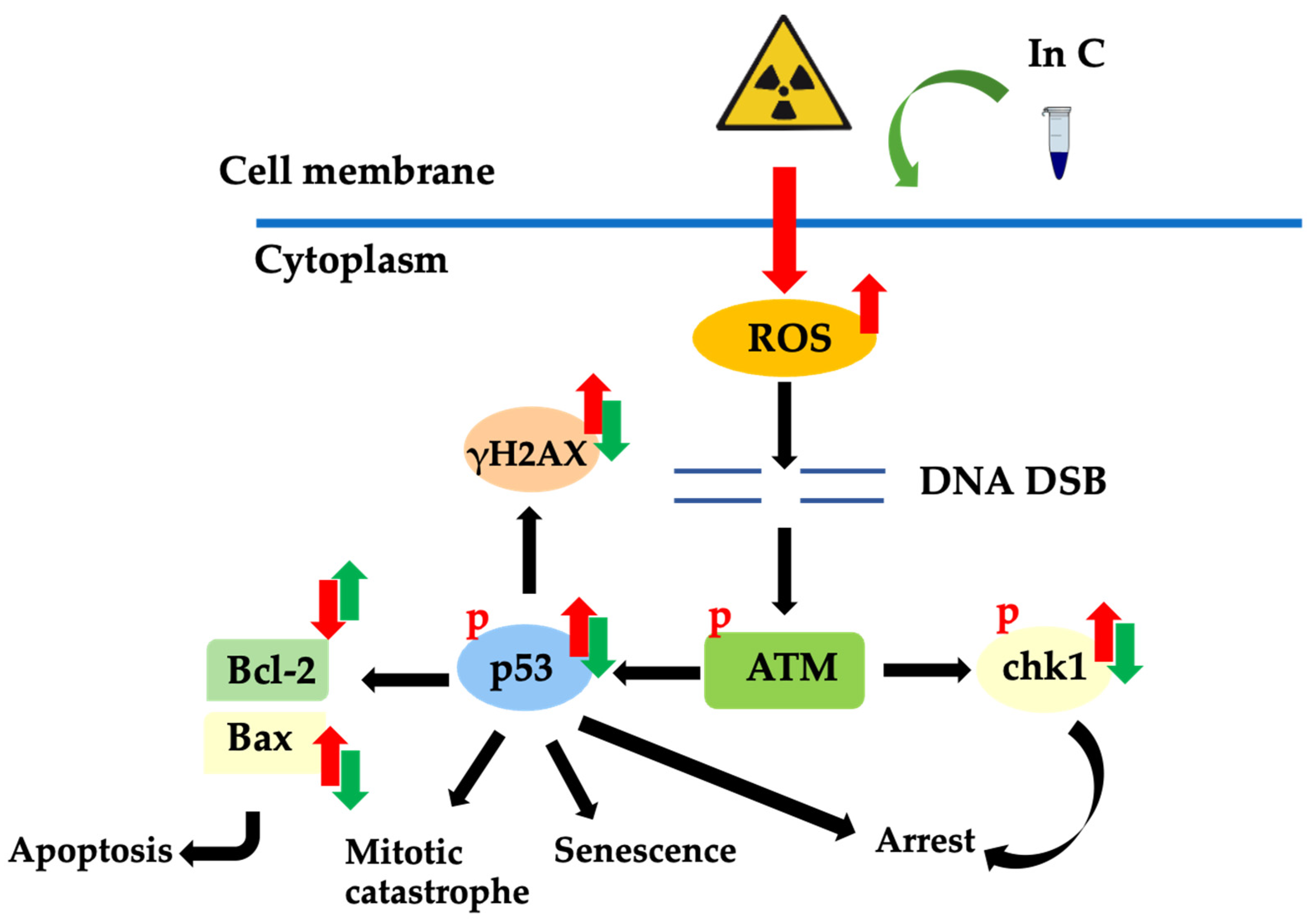
3. Discussion
4. Materials and Methods
4.1. Compound Preparation
4.2. Cell Culture and X-ray Irradiation
4.3. DCFH-DA Assay
4.4. MTT Assay
4.5. Clonogenic Cell Survival Assay
4.6. SOD Activity Assay
4.7. γH2AX Assay
4.8. MN Formation Assay
4.9. Western Blot Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Podgorsak, E.B. Radiation Oncology Physics: A Handbook for Teachers and Students; International Atomic Energy Agency: Vienna, Austria, 2005. [Google Scholar]
- Wild, C.P.; Weiderpass, E.; Stewart, B.W. World Cancer Report: Cancer Research for Cancer Prevention; International Agency for Research on Cancer: Lyon, France, 2020. [Google Scholar]
- Greenberger, J.S. Radioprotection. In Vivo 2009, 23, 323–336. [Google Scholar] [PubMed]
- Nair, C.K.; Parida, D.K.; Nomura, T. Radioprotectors in radiotherapy. J. Radiat Res. 2001, 42, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Maurya, D.K.; Devasagayam, T.P. Role of radioprotectors in the inhibition of DNA damage and modulation of DNA repair after exposure to gamma-radiation. In Selected Topics in DNA Repair; Chen, C., Ed.; InTech: Shanghai, China, 2011; pp. 483–496. [Google Scholar] [CrossRef] [Green Version]
- Raviraj, J.; Bokkasam, V.; Kumar, V.; Reddy, U.; Suman, V. Radiosensitizers, radioprotectors, and radiation mitigators. Indian J. Dent. Res. 2014, 25, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Grochová, D.; Šmardová, J. The antimutagenic and cytoprotective effects of amifostine: The role of p53. J. Appl Biomed. 2007, 5, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Kouvarisa, J.R.; Koulouliasb, V.E.; Vlahosa, L.J. Amifostine: The first selective-target and broad-spectrum radioprotector. Oncologist 2007, 12, 738–747. [Google Scholar] [CrossRef] [Green Version]
- Putora, P.M.; Schmuecking, M.; Aebersold, D.; Plasswilm, L. Compensability index for compensation radiotherapy after treatment interruptions. Radiat Oncol. 2012, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Fischer, N.; Seo, E.J.; Efferth, T. Review: Prevention from radiation damage by natural products. Phytomedicine 2017, 47, 192–200. [Google Scholar] [CrossRef]
- Maurya, D.K.; Devasagayam, T.; Nair, C. Some novel approaches for radioprotection and the beneficial effect of natural products. Indian J. Exp. Biol. 2006, 44, 93–114. [Google Scholar]
- Al-Amiery, A.A.; Saour, K.Y.; A-Duhaidahawi, D.L.; Al-Majedy, Y.K.; Kadhum, A.A.; Mohamad, A.B. Comparative molecular modelling studies of coumarin derivatives as potential antioxidant agents. Free Radic Res. 2017, 7, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Plants of the World Online. Available online: https://powo.science.kew.org/taxon/urn:Lsid:ipni.org:names:17462260-1 (accessed on 29 April 2022).
- Winter, W.P.; Amoroso, V.B. Plant. Resources of South-East Asia. Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 56–58. [Google Scholar]
- Lin, Y.X.; Li, Z.Y.; Iwatsuki, K.; Smith, A.R. Thelypteridaceae. In Flora of China; Pteridophytes; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2013; Volume 2–3, p. 376. [Google Scholar]
- Quadri-Spinelli, T.; Heilmann, J.; Rali, T.; Sticher, O. Bioactive coumarin derivatives from the fern Cyclosorus interruptus. Planta Med. 2000, 66, 728–733. [Google Scholar] [CrossRef]
- Kaewsuwan, S.; Yuenyongsawad, S.; Plubrukarn, A.; Kaewchoothong, A.; Raksawong, A.; Puttarak, P.; Apirug, C. Bioactive interruptin A and B from Cyclosorus terminans: Antibacterial, anticancer, stem cell proliferation and ROS scavenging activities. Songklanakarin J. Sci Technol. 2015, 37, 309–317. [Google Scholar]
- Smith, T.A.; Kirkpatrick, D.R.; Smith, S.; Smith, T.K.; Pearson, T.; Kailasam, A.; Herrmann, K.Z.; Schubert, J.; Agrawal, D.K. Radioprotective agents to prevent cellular damage due to ionizing radiation. J. Transl. Med. 2017, 5, 232. [Google Scholar] [CrossRef] [PubMed]
- Mirzayans, R.; Andrais, B.; Murray, D. Impact of premature senescence on radiosensitivity measured by high throughput cell-based assays. Int. J. Mol. Sci. 2017, 18, 1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.I.; Oberley, L.W.; Li, Y. A simple method for clinical assay of superoxide dismutase. Clin. Chem. 1988, 34, 497–500. [Google Scholar] [CrossRef]
- Candas, D.; Li, J.J. MnSOD in oxidative stress response-potential regulation via mitochondrial protein influx. Antioxid. Redox. Signal. 2014, 20, 1599–1617. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.P.; Roe, J.H.; Chock, P.B.; Yim, M.B. Transcriptional activation of the human manganese superoxide dismutase gene mediated by tetradecanoylphorbol acetate. J. Biol. Chem. 1999, 274, 37455–37460. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Wu, W.; Fu, W.; Hu, Y. The effects of phototherapy and melanocytes on keratinocytes. Exp. Ther. Med. 2018, 15, 3459–3466. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.; Moon, H.W.; Oh, Y.; Kim, K.; Kim, D.D.; Lim, C.J. Defensive Properties of Ginsenoside Re against UV-B-Induced Oxidative Stress through Up-Regulating Glutathione and Superoxide Dismutase in HaCaT Keratinocytes. Iran J. Pharm. Sci. 2018, 17, 249–260. [Google Scholar] [PubMed]
- Becuwe, P.; Ennen, M.; Klotz, R.; Barbieux, C.; Grandemange, S. Manganese superoxide dismutase in breast cancer: From molecular mechanisms of gene regulation to biological and clinical significance. Free Radic. Biol. Med. 2014, 77, 139–151. [Google Scholar] [CrossRef]
- Papa, L.; Hahn, M.; Marsh, E.L.; Evans, B.S.; Germain, D. SOD2 to SOD1 Switch in Breast Cancer. J. Biol. Chem. 2014, 289, 5412–5416. [Google Scholar] [CrossRef] [Green Version]
- Banasiak, D.; Barnetson, A.R.; Odell, R.A. Comparison between the Clonogenic MTT and SRB assays for determining radiosensitivity in a panel of human bladder cancer cell lines and a ureteral cell line. Radiat. Oncol. Investig. 1999, 7, 7–85. [Google Scholar] [CrossRef]
- Lomax, M.E.; Folkes, L.K.; O’Neill, P. Biological consequences of radiation-induced DNA damage: Relevance to radiotherapy. Clin. Oncol. (R Coll. Radiol.). 2013, 25, 578–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tommasino, F.; Friedrich, T.; Jakob, B.; Meyer, B.; Durante, M.; Scholz, M. Induction and processing of the radiation-induced gamma-H2AX ssignal and its link to the underlying pattern of DSB: A combined experimental and modelling study. PLoS ONE. 2015, 10, e0129416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. GammaH2AX: A sensitive molecular marker of DNA damage and repair. Lukemia 2010, 24, 679–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariotti, L.G.; Pirovano, G.; Savage, K.I.; Ghita, M.; Ottolenghi, A.; Prise, K.M.; Schettino, G. Use of the γ-H2AX assay to investigate DNA repair dynamics following multiple radiation exposures. PLoS ONE. 2013, 8, e79541. [Google Scholar] [CrossRef] [Green Version]
- Hall, E.J.; Giaccia, A.J. Physics and Chemistry of Radiation Absorption. In Radiobiology for the Radiologist, 7th ed.; Wolters Kluwer/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012; pp. 3–11. [Google Scholar]
- Wouters, B.G.; Begg, A.C. Irradiation-induced damage and the DNA damage response. In Basic Clinical Radiobiology, 4th ed.; Joiner, M., Kogel, A., Eds.; Hodder Arnold: London, UK, 2009; pp. 11–26. [Google Scholar]
- Baskar, R.; Dai, J.; Wenlong, N.; Yeo, R.; Yeoh, K.W. Biological response of cancer cells to radiation treatment. Front. Mol. Biosci. 2014, 1, 24. [Google Scholar] [CrossRef] [Green Version]
- Parshad, R.; Sanford, K.K.; Jones, G.M. Chromatid damage after G2 phase x-irradiation of cells from cancer-prone individuals implicates deficiency in DNA repair. Proc. Natl. Acad. Sci. USA 1983, 80, 5612–5616. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, M.; Mozdarani, H.; Mueller, W.U. Radiosensitivity and repair kinetics of gamma-irradiated leukocytes from sporadic prostate cancer patients and healthy individuals assessed by alkaline comet assay. Iran. Biomed. J. 2010, 14, 67–75. [Google Scholar]
- Anders, C.K.; Abramson, V.; Tan, T.; Dent, R. The Evolution of Triple-Negative Breast Cancer: From Biology to Novel Therapeutics. Am. Soc. Clin. Oncol. Educ. Book 2016, 35, 34–42. [Google Scholar] [CrossRef]
- Sharma, P. Biology and Management of Patients With Triple-Negative Breast Cancer. Oncologist 2016, 21, 1050–1062. [Google Scholar] [CrossRef] [Green Version]
- Hall, E.J.; Giaccia, A.J. Clinical Response of Normal Tissues. In Radiobiology for the Radiologist, 7th ed.; Wolters Kluwer/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012; pp. 327–355. [Google Scholar]
- Moding, E.J.; Kastan, M.B.; Kirsch, D.G. Strategies for optimizing the response of cancer and normal tissues to radiation. Nat. Rev. Drug Discov. 2013, 12, 526–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, P.; Hartmann, L.; Wenz, F.; Herskind, C. Cellular Pathways in Response to Ionizing Radiation and Their Targetability for Tumor Radiosensitization. Int. J. Mol. Sci. 2016, 17, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paglin, S.; Delohery, T.; Erlandson, R.; Yahalom, J. Radiation-induced micronuclei formation in human breast cancer cells: Dependence on serum and cell cycle distribution. Biochem. Biophys. Res. Commun. 1997, 237, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Cheki, M.; Shirazi, A.; Mahmoudzadeh, A.; Bazzaz, J.T.; Hosseinimehr, S.J. The radioprotective effect of metformin against cytotoxicity and genotoxicity induced by ionizing radiation in cultured human blood lymphocytes. Mutat Res. Genet. Toxicol. Environ. Mutagen. 2016, 809, 24–32. [Google Scholar] [CrossRef]
- Li, J.; Feng, L.; Xing, Y.; Wang, Y.; Du, L.; Xu, C.; Cao, Y.; Wang, Q.; Fan, S.; Liu, Q.; et al. Radioprotective and antioxidant effect of resveratrol in hippocampus by activating Sirt1. Int. J. Mol. Sci. 2014, 15, 5928–5939. [Google Scholar] [CrossRef] [Green Version]
- Jeong, M.H.; Yang, K.M.; Jeong, D.H.; Lee, C.G.; Oh, S.J.; Jeong, S.K.; Lee, K.W.; Jo, Y.R.; Jo, W.S. Protective activity of a novel resveratrol analogue, HS-1793, against DNA damage in 137Cs-irradiated CHO-K1 cells. J. Radiat. Res. 2014, 55, 464–475. [Google Scholar] [CrossRef] [Green Version]
- Özgen, S.C.; Dökmeci, D.; Akpolat, M.; Karadağ, C.H.; Gündüz, Ö.; Erbaş, H.; Benian, Ö.; Uzal, C.; Turan, F.N. The protective effect of curcumin on ionizing radiation-induced cataractogenesis in rats. Balkan Med. J. 2012, 29, 358–363. [Google Scholar] [CrossRef]
- Kumar, A.; Choudhary, S.; Adhikari, J.S.; Chaudhury, N.K. Sesamol Ameliorates Radiation Induced DNA Damage in Hematopoietic System of Whole Body γ-Irradiated Mice. Environ. Mol. Mutagen. 2018, 59, 79–90. [Google Scholar] [CrossRef]
- Song, L.; Ma, L.; Cong, F.; Shen, X.; Jing, P.; Ying, X.; Zhou, H.; Jiang, J.; Fu, Y.; Yan, H. Radioprotective effects of genistein on HL-7702 cells via the inhibition of apoptosis and DNA damage. Cancer Lett. 2015, 366, 100–111. [Google Scholar] [CrossRef]
- Aranda, A.; Sequedo, L.; Tolosa, L.; Quintas, G.; Burello, E.; Castell, J.V.; Gombau, L. Dichloro-dihydro-fluorescein diacetate (DCFH-DA) assay: A quantitative method for oxidative stress assessment of nanoparticle-treated cells. Toxicol. Vitro. 2013, 27, 954–963. [Google Scholar] [CrossRef]
- Buch, K.; Peters, T.; Nawroth, T.; Sanger, M.; Schmidberger, H.; Langguth, P. Determination of cell survival after irradiation via clonogenic assay versus multiple MTT Assay—A comparative study. Radiat. Oncol. 2012, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munshi, A.; Hobbs, M.; Meyn, R.E. Clonogenic cell survival assay. Methods Mol. Med. 2005, 110, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Khoram, N.M.; Bigdeli, B.; Nikoofar, A.; Goliaei, B. Caffeic Acid Phenethyl Ester Increases Radiosensitivity of Estrogen Receptor-Positive and -Negative Breast Cancer Cells by Prolonging Radiation-Induced DNA Damage. J. Breast Cancer 2016, 19, 18–25. [Google Scholar] [CrossRef]
- Khongkow, P.; Gomes, A.; Gong, C.; Man, E.P.; Tsang, J.W.-H.; Zhao, F.; Monteiro, L.J.; Coombes, R.C.; Medema, R.H.; Khoo, U.S.; et al. Paclitaxel targets FOXM1 to regulate KIF20A in mitotic catastrophe and breast cancer paclitaxel resistance. Oncogene 2016, 35, 990–1002. [Google Scholar] [CrossRef] [Green Version]
- Khongkow, P.; Middleton, A.W.; Wong, J.P.; Kandola, A.K.; Kongsema, M.; Moraes, G. Erratum to: In Vitro Methods for Studying the Mechanisms of Resistance to DNA-Damaging Therapeutic Drugs. In Cancer Drug Resistance. Methods in Molecular Biology; Rueff, J., Rodrigues, A., Eds.; Humana Press: New York, NY, USA, 2016; Volume 1395. [Google Scholar] [CrossRef] [Green Version]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Weydert, C.J.; Cullen, J.J. Measurements of the antioxidant enzyme activities of superoxide dismutase, catalase, and glutathione peroxidase in cultured cells and tissue. Nat. Protoc. 2010, 5, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Mah, L.; Vasireddy, R.S.; Tang, M.M.; Georgiadis, G.T.; El-Osta, A.; Karagiannis, T.C. Quantification of γH2AX Foci in Response to Ionising Radiation. JoVE 2010, 38, e1957. [Google Scholar] [CrossRef] [Green Version]
- Khongkow, P.; Karunarathna, U.; Khongkow, M.; Gong, C.; Gomes, A.R.; Yagüe, E.; Monteiro, L.J.; Kongsema, M.; Zona, S.; Man, E.P.; et al. FOXM1 targets NBS1 to regulate DNA damage-induced senescence and epirubicin resistance. Oncogene 2014, 33, 4144–4156. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.M.; Kang, C.M.; Cho, E.S.; Kang, S.M.; Lee, S.B.; Um, H.D. Ionizing radiation-induced micronucleus formation is mediated by reactive oxygen species that are produced in a manner dependent on mitochondria, Nox1, and JNK. Oncol. Rep. 2007, 17, 1183–1188. [Google Scholar] [CrossRef] [Green Version]
- Kamran, M.Z.; Ranjan, A.; Kaur, N.; Sur, S.; Tandon, V. Radioprotective Agents: Strategies and translational advances. Med. Res. Rev. 2016, 36, 461–493. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chumsuwan, N.; Khongkow, P.; Kaewsuwan, S.; Kanokwiroon, K. Interruptin C, a Radioprotective Agent, Derived from Cyclosorus terminans Protect Normal Breast MCF-10A and Human Keratinocyte HaCaT Cells against Radiation-Induced Damage. Molecules 2022, 27, 3298. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103298
Chumsuwan N, Khongkow P, Kaewsuwan S, Kanokwiroon K. Interruptin C, a Radioprotective Agent, Derived from Cyclosorus terminans Protect Normal Breast MCF-10A and Human Keratinocyte HaCaT Cells against Radiation-Induced Damage. Molecules. 2022; 27(10):3298. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103298
Chicago/Turabian StyleChumsuwan, Nipha, Pasarat Khongkow, Sireewan Kaewsuwan, and Kanyanatt Kanokwiroon. 2022. "Interruptin C, a Radioprotective Agent, Derived from Cyclosorus terminans Protect Normal Breast MCF-10A and Human Keratinocyte HaCaT Cells against Radiation-Induced Damage" Molecules 27, no. 10: 3298. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103298