
Three Diverse Granule Preparation Methods for Proteomic Analysis of Mature Rice (Oryza sativa L.) Starch Grain

 ,
,  and
and
Abstract
:1. Introduction
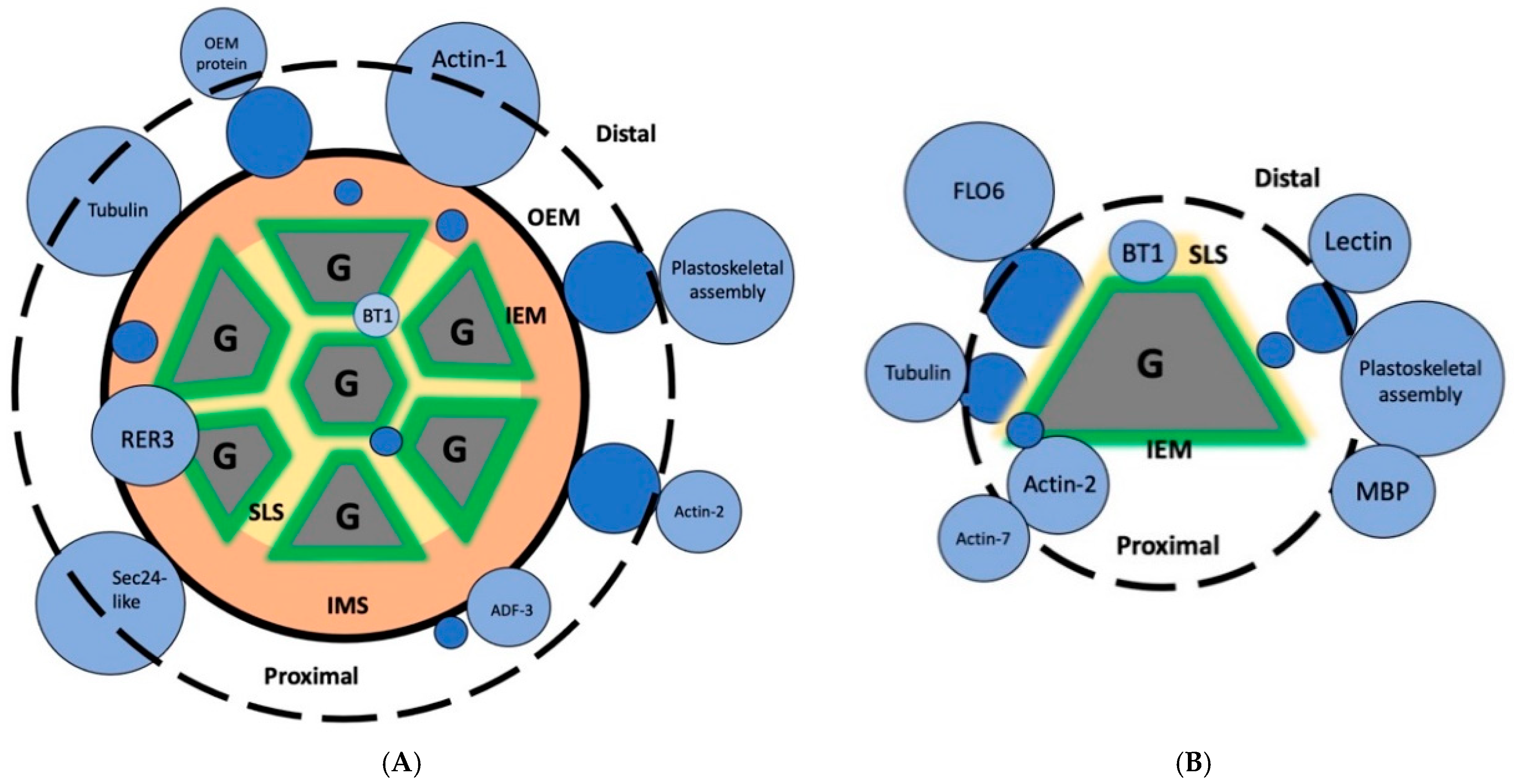
2. Results
3. Discussion
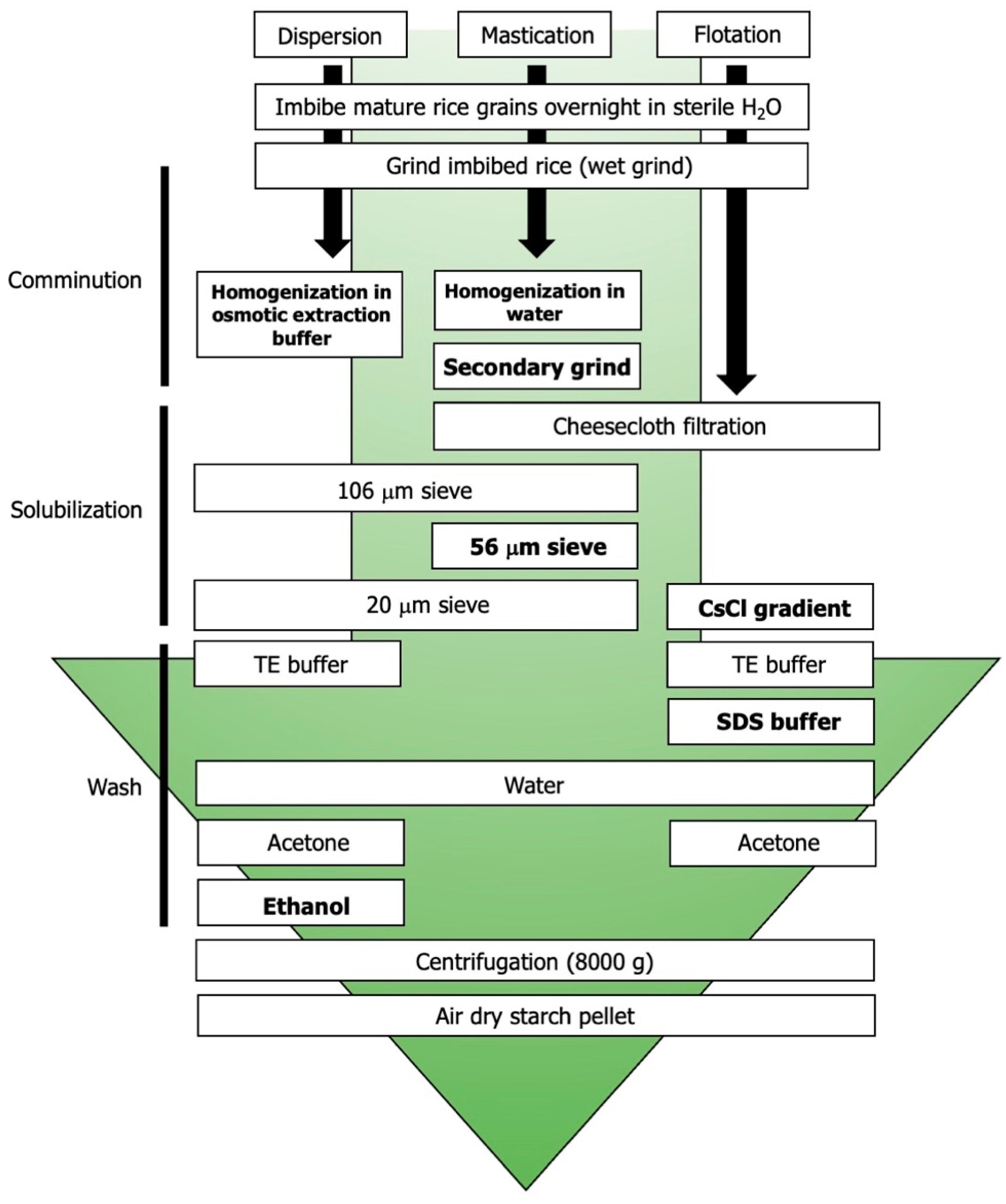
- Dispersion-based disruption of the compound granule using osmotic buffer;
- Microsieving;
- Flotation-based disruption using a cesium-chloride gradient.
- This analysis used mature rice kernels, and so limits analysis on SGAPs involved in grain architecture during development (such as plastid division proteins), which will no longer be present in the mature endosperm. A time-course analysis of the SGAP proteome during rice kernel development must be performed to obtain a dynamic model. Observing the development of a simple SG, such as in maize, would provide side-by-side proteomic comparisons and could reveal novel candidates for compound SG architecture development. Whether these types of organization differ in starch mobilization rate is unknown and should also be investigated.
- Each method of preparation disrupts SGs into individual granules (Figure 2), suggesting that the internal or core SLS proteins are as exposed to the protein extraction methods as the proteins in the distal proteome. A fine-tuned, gentler approach would involve the preparation of intact SGs so that one can distinguish between the outer and inner proteome of the rice starch grain.
4. Materials and Methods
4.1. Plant Material
4.2. Starch Granule Preparation
- Dispersion method [51]: rice kernels (5 g) were ground via mortar and pestle for five minutes prior to the addition of 10 mL starch extraction buffer (50 mM Tris-HCl, pH 7; 10% glycerol; 10 mM EDTA; 1.25 mM DTT). The sample was subjected to vacuum sieve filtration through a 106 μm sieve and the resulting filtrate was centrifuged (4600× g for 15 min at 4 °C). The supernatant was discarded, and the pellet was resuspended with 5 mL starch extraction buffer. The dispersion was subjected to vacuum sieve filtration through a 20 μm sieve. The filtrate was washed with starch extraction buffer followed by cold 95% ethanol and acetone. Centrifuging was performed between each wash (8000× g, 10 min, 4 °C). Pellets were air-dried under laminar flow for 48 h.
- Microsieve method [52]: rice kernels (5 g) were manually ground via mortar and pestle for five minutes. Then, 10 mL of sterile double-distilled water was added before continuing the grinding process for an additional five minutes. This slurry was filtered through five layers of cheesecloth and reground for two minutes with mortar and pestle. The resulting dispersion was transferred to a vacuum sieve and filtered through 106 μm, 53 μm, and 20 μm sieves (Gilson Company, Inc., Lewis Center, OH, USA) in series. The filtrate was centrifuged twice (4600× g for 15 min at 4 °C) and the supernatant was discarded. Pellets were air-dried under laminar flow for 48 h.
- Flotation method [53]: rice kernels (5 g) were manually ground via mortar and pestle for five minutes. Then, 10 mL of sterile double-distilled water was added before continuing the grinding process for an additional five minutes. The dispersion was filtered through five layers of cheesecloth and centrifuged (4600× g for 15 min at 4 °C). The supernatant was discarded, and the pellet was resuspended in 1 mL of sterile double-distilled water overlaid with 80% w/v cesium chloride. The solution was centrifuged (4600× g for five minutes at 4 °C) and the supernatant discarded. The pellet was washed with a wash buffer (62.5 mM Tris-HCl, pH 6.8; 10 mM EDTA; 4% SDS), sterile double-distilled water, and acetone. Centrifuging was performed between each wash (8000× g at 10 min for 4 °C). Pellets were air-dried under laminar flow for 48 h.
4.3. Peptide Preparation
4.4. Scanning Electron Microscopy (SEM)
4.5. Nano-HPLC-MS/MS Analyses of Peptides
4.6. Peptide Identification
4.7. Bioinformatics Analysis
4.8. Public Database Repository
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Pal, S.K.; Liput, M.; Piques, M.; Ishihara, H.; Obata, T.; Martins, M.C.; Sulpice, R.; van Dongen, J.T.; Fernie, A.R.; Yadav, U.P.; et al. Diurnal changes of polysome loading track sucrose content in the rosette of wild-type arabidopsis and the starchless pgm mutant. Plant Physiol. 2013, 162, 1246–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichert, E.T. The Differentiation and Specificity of Starches in Relation to Genera, Species, etc: Stereochemistry Applied to Protoplasmic Processes and Products, and as a Strictly Scientific Basis for the Classification of Plants and Animals; Carnegie Institution of Washington: Washington, DC, USA, 1913. [Google Scholar]
- Matsushima, R.; Maekawa, M.; Fujita, N.; Sakamoto, W. A rapid, direct observation method to isolate mutants with defects in starch grain morphology in rice. Plant Cell Physiol. 2010, 51, 728–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, M.-S.; Kawagoe, Y. Amyloplast division progresses simultaneously at multiple sites in the endosperm of rice. Plant Cell Physiol. 2009, 50, 1617–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyosawa, Y.; Kawagoe, Y.; Matsushima, R.; Crofts, N.; Ogawa, M.; Fukuda, M.; Kumamaru, T.; Okazaki, Y.; Kusano, M.; Saito, K. Deficiency of starch synthase IIIa and IVb alters starch granule morphology from polyhedral to spherical in rice endosperm. Plant Physiol. 2016, 170, 1255–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, M.-S.; Kawagoe, Y. Septum formation in amyloplasts produces compound granules in the rice endosperm and is regulated by plastid division proteins. Plant Cell Physiol. 2010, 51, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Yi, N.; Kim, Y.S.; Jeong, M.-H.; Bang, S.-W.; Choi, Y.D.; Kim, J.-K. Analysis of five novel putative constitutive gene promoters in transgenic rice plants. J. Exp. Bot. 2010, 61, 2459–2467. [Google Scholar] [CrossRef] [Green Version]
- Cakir, B.; Shiraishi, S.; Tuncel, A.; Matsusaka, H.; Satoh, R.; Singh, S.; Crofts, N.; Hosaka, Y.; Fujita, N.; Hwang, S.-K. Analysis of the rice ADP-glucose transporter (OsBT1) indicates the presence of regulatory processes in the amyloplast stroma that control ADP-glucose flux into starch. Plant Physiol. 2016, 170, 1271–1283. [Google Scholar]
- Li, S.; Wei, X.; Ren, Y.; Qiu, J.; Jiao, G.; Guo, X.; Tang, S.; Wan, J.; Hu, P. OsBT1 encodes an ADP-glucose transporter involved in starch synthesis and compound granule formation in rice endosperm. Sci. Rep. 2017, 7, 40124. [Google Scholar] [CrossRef] [Green Version]
- Benmoussa, M.; Hamaker, B.R.; Huang, C.P.; Sherman, D.M.; Weil, C.F.; BeMiller, J.N. Elucidation of maize endosperm starch granule channel proteins and evidence for plastoskeletal structures in maize endosperm amyloplasts. J. Cereal Sci. 2010, 52, 22–29. [Google Scholar] [CrossRef]
- Yoshida, Y.; Miyagishima, S.-y.; Kuroiwa, H.; Kuroiwa, T. The plastid-dividing machinery: Formation, constriction and fission. Curr. Opin. Plant Biol. 2012, 15, 714–721. [Google Scholar] [CrossRef]
- Maple, J.; Vojta, L.; Soll, J.; Møller, S.G. ARC3 is a stromal Z-ring accessory protein essential for plastid division. EMBO Rep. 2007, 8, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, P.M. Starch granule-associated proteins and polypeptides: A review. Starch-Stärke 2001, 53, 475–503. [Google Scholar] [CrossRef]
- Hamaker, B.; Griffin, V.; Moldenhauer, K. Potential influence of a starch granule-associated protein on cooked rice stickiness. J. Food Sci. 1991, 56, 1327–1329. [Google Scholar] [CrossRef]
- Koziol, A.G.; Marquez, B.K.; Huebsch, M.P.; Smith, J.C.; Altosaar, I. Commercially produced rice and maize starches contain nonhost proteins, as shown by mass spectrometry. Cereal Chem. 2012, 89, 262–264. [Google Scholar] [CrossRef]
- Koziol, A.G.; Marquez, B.K.; Huebsch, M.P.; Smith, J.C.; Altosaar, I. The starch granule associated proteomes of commercially purified starch reference materials from rice and maize. J. Proteom. 2012, 75, 993–1003. [Google Scholar] [CrossRef]
- Wall, M.; Wheeler, H.; Huebsch, M.; Smith, J.; Figeys, D.; Altosaar, I. The tryptophan-rich domain of puroindoline is directly associated with the starch granule surface as judged by tryptic shaving and mass spectrometry. J. Cereal Sci. 2010, 52, 115–120. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Kirchberger, S.; Leroch, M.; Huynen, M.A.; Wahl, M.; Neuhaus, H.E.; Tjaden, J. Molecular and biochemical analysis of the plastidic ADP-glucose transporter (ZmBT1) from Zea mays. J. Biol. Chem. 2007, 282, 22481–22491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staiger, C.J.; Gibbon, B.C.; Kovar, D.R.; Zonia, L.E. Profilin and actin-depolymerizing factor: Modulators of actin organization in plants. Trends Plant Sci. 1997, 2, 275–281. [Google Scholar] [CrossRef]
- Wu, C.H.; Arighi, C.N.; Ross, K.E. Protein Bioinformatics: From Protein Modifications and Networks to Proteomics (Methods in Molecular Biology); Springer Nature: Berlin, Germany, 2017. [Google Scholar]
- Cheng, J.; Romantsov, T.; Engel, K.; Doxey, A.C.; Rose, D.R.; Neufeld, J.D.; Charles, T.C. Functional metagenomics reveals novel β-galactosidases not predictable from gene sequences. PLoS ONE 2017, 12, e0172545. [Google Scholar]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [Green Version]
- Seung, D.; Soyk, S.; Coiro, M.; Maier, B.A.; Eicke, S.; Zeeman, S.C. PROTEIN TARGETING TO STARCH is required for localising GRANULE-BOUND STARCH SYNTHASE to starch granules and for normal amylose synthesis in Arabidopsis. PLoS Biol. 2015, 13, e1002080. [Google Scholar] [CrossRef] [Green Version]
- Boraston, A.B.; Bolam, D.N.; Gilbert, H.J.; Davies, G.J. Carbohydrate-binding modules: Fine-tuning polysaccharide recognition. Biochem. J. 2004, 382, 769–781. [Google Scholar] [CrossRef]
- Keeratijarut, A.; Lohnoo, T.; Rujirawat, T.; Yingyong, W.; Kalambaheti, T.; Miller, S.; Phuntumart, V.; Krajaejun, T. The immunoreactive exo-1, 3-β-glucanase from the pathogenic oomycete Pythium insidiosum is temperature regulated and exhibits glycoside hydrolase activity. PLoS ONE 2015, 10, e0135239. [Google Scholar] [CrossRef]
- Simpson, C.; Thomas, C.; Findlay, K.; Bayer, E.; Maule, A.J. An Arabidopsis GPI-anchor plasmodesmal neck protein with callose binding activity and potential to regulate cell-to-cell trafficking. Plant Cell 2009, 21, 581–594. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, D.; Mukhopadhyay, C. Binding diversity of the two binding sites of ricin B lectin. Biopolym. Orig. Res. Biomol. 2006, 83, 83–94. [Google Scholar] [CrossRef]
- Rutishauser, U.; Sachs, L. Cell-to-cell binding induced by different lectins. J. Cell Biol. 1975, 65, 247–257. [Google Scholar] [CrossRef]
- Champagne, E. Rice starch composition and characteristics. Cereal Foods World 1996, 41, 833–838. [Google Scholar]
- Boukebbous, K.; Moresco, J.J.; Yates, J.R., III; Altosaar, I. Protein Extraction Efficiency of Soft and Hard Seeds Using the Precellys Lysing Kits. Department of Chemical Physiology, The Scripps Research Institute: La Jolla, CA, USA, 24 September 2014. [Google Scholar]
- Lee, B.-h.; Henderson, D.A.; Zhu, J.-K. The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell 2005, 17, 3155–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, D.K. The 14-3-3 proteins: Integrators of diverse signaling cues that impact cell fate and cancer development. Trends Cell Biol. 2009, 19, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeli, K.; Altosaar, I. Compartmentalization of seed reserve proteins: Proposed discriminatory factors. FEBS Lett. 1984, 178, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Osborne, T. The Proteins of the Wheat Kernel; The Carnegie Institution of Washington: Washington, DC, USA, 1907; p. 107. [Google Scholar]
- Robert, L.S.; Nozzolillo, C.; Altosaar, I. Homology between rice glutelin and oat 12 S globulin. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 1985, 829, 19–26. [Google Scholar] [CrossRef]
- Robert, L.S.; Nozzolillo, C.; Cudjoe, A.; Altosaar, I. Total solubilization of groat proteins in high protein oat (Avena sativa L. cv. Hinoat): Evidence that glutelins are a minor component. Can. Inst. Food Sci. Technol. J. 1983, 16, 196–200. [Google Scholar] [CrossRef]
- Kurata, N.; Nonomura, K.I.; Harushima, Y. Rice genome organization: The centromere and genome interactions. Ann. Bot. 2002, 90, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Niu, L.; Ding, H.; Hao, R.; Liu, H.; Wu, X.; Hu, X.; Wang, W. A rapid and universal method for isolating starch granules in plant tissues. Plant Cell Environ. 2019, 42, 3355–3371. [Google Scholar] [CrossRef]
- Lou, H.-Y.; Zhao, W.; Li, X.; Duan, L.; Powers, A.; Akamatsu, M.; Santoro, F.; McGuire, A.F.; Cui, Y.; Drubin, D.G. Membrane curvature underlies actin reorganization in response to nanoscale surface topography. Proc. Natl. Acad. Sci. USA 2019, 116, 23143–23151. [Google Scholar] [CrossRef]
- Lundquist, P.K.; Rosar, C.; Bräutigam, A.; Weber, A.P. Plastid signals and the bundle sheath: Mesophyll development in reticulate mutants. Mol. Plant 2014, 7, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Pérez, J.M.; Esteve-Bruna, D.; González-Bayón, R.; Kangasjärvi, S.; Caldana, C.; Hannah, M.A.; Willmitzer, L.; Ponce, M.R.; Micol, J.L. Functional redundancy and divergence within the Arabidopsis RETICULATA-RELATED gene family. Plant Physiol. 2013, 162, 589–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Bayon, R.; Kinsman, E.A.; Quesada, V.; Vera, A.; Robles, P.; Ponce, M.R.; Pyke, K.A.; Micol, J.L. Mutations in the RETICULATA gene dramatically alter internal architecture but have little effect on overall organ shape in Arabidopsis leaves. J. Exp. Bot. 2006, 57, 3019–3031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anstis, P.; Northcote, D. Development of chloroplasts from amyloplasts in potato tuber discs. New Phytol. 1973, 72, 449–463. [Google Scholar] [CrossRef]
- Billault-Chaumartin, I.; Martin, S.G. Capping protein insulates Arp2/3-assembled actin patches from formins. Curr. Biol. 2019, 29, 3165–3176.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, L.-R.; Park, C.H.; Venu, R.; Gough, J.; Wang, G.-L. Classification, expression pattern, and E3 ligase activity assay of rice U-box-containing proteins. Mol. Plant 2008, 1, 800–815. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Zhang, X.; Yang, X.; Li, G.; Tang, S.; Wang, S.; Ding, Y.; Liu, Z. Proteomic analysis of proteins related to rice grain chalkiness using iTRAQ and a novel comparison system based on a notched-belly mutant with white-belly. BMC Plant Biol. 2014, 14, 163. [Google Scholar] [CrossRef] [Green Version]
- Xing, S.; Meng, X.; Zhou, L.; Mujahid, H.; Zhao, C.; Zhang, Y.; Wang, C.; Peng, Z. Proteome profile of starch granules purified from rice (Oryza sativa) endosperm. PLoS ONE 2016, 11, e0168467. [Google Scholar]
- Man, J.; Lin, L.; Wang, Z.; Wang, Y.; Liu, Q.; Wei, C. Different structures of heterogeneous starch granules from high-amylose rice. J. Agric. Food Chem. 2014, 62, 11254–11263. [Google Scholar] [CrossRef]
- Peng, M.; Gao, M.; Abdel-Aal, E.S.; Hucl, P.; Chibbar, R. Separation and characterization of A-and B-type starch granules in wheat endosperm. Cereal Chem. 1999, 76, 375–379. [Google Scholar] [CrossRef]
- Burgos-Canul, Y.Y.; Canto-Canché, B.; Berezovski, M.V.; Mironov, G.; Loyola-Vargas, V.M.; de Rosa, A.P.B.; Tzec-Simá, M.; Brito-Argáez, L.; Carrillo-Pech, M.; Grijalva-Arango, R. The cell wall proteome from two strains of Pseudocercospora fijiensis with differences in virulence. World J. Microbiol. Biotechnol. 2019, 35, 105. [Google Scholar] [CrossRef]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. PEAKS DB: De novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteom. 2012, 11, M111.010587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimmy, K.; Eng, A.; Yates, J., III. An approach to correlate tandem mass spectral data of peptides with amino acid sequences in a protein database. Am. Soc. Mass Spectr. 1994, 5, 979–989. [Google Scholar]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; Hewapathirana, S.; García-Seisdedos, D.; Kamatchinathan, S.; Kundu, D.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A Hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Starch Preparation Method | Protein Extraction Method | # of Peptides (# Uncharacterized) |
|---|---|---|
| Dispersion | Trypsin | 1575(663) |
| Isopropanol | 559(212) | |
| Microsieving | Trypsin | 123(39) |
| Isopropanol | 154(45) | |
| Flotation | Trypsin | 24(8) |
| Isopropanol | 49(10) |
| UniProt ID | Description |
|---|---|
| Carbohydrate Synthesis and Metabolism | |
| Q0JKM8 | Aspartic proteinase oryzasin-1-like |
| Q852L2 | Cupincin |
| B7EVB8 | Glucose-1-phosphate adenylyltransferase |
| P15280 | Glucose-1-phosphate adenylyltransferase small subunit 2 * |
| Q65XK0 | Ketol-acid reductoisomerase |
| Q75M03 | Putative H+-pyrophosphatase |
| Q6AVA8 | Pyruvate, phosphate dikinase 1, chloroplastic * |
| Q6ZBH2 | Sorbitol dehydrogenase * |
| B3VDJ4 | Starch branching enzyme |
| Q43009 | Sucrose synthase 3 |
| Q93X08 | UTP-glucose-1-phosphate uridylyltransferase |
| Miscellaneous | |
| Q6ZKC0 | 14-3-3-like protein GF14-C |
| Q6Z782 | Brittle1 (BT1) |
| Q0DEP9 | Early nodulin-93 |
| A0A0P0WFP9 | Fatty acid export 2, chloroplastic |
| Q6ZHP6 | Outer envelope membrane protein 7 |
| Q9FWV6 | Probable aquaporin TIP3-1 |
| Q10Q18 | Transport protein Sec61 alpha subunit isoform 2 |
| Q6Z0Z9 | V-type proton ATPase proteolipid subunit |
| Seed Reserve | |
| Q8GVK5 | 13 kDa prolamin |
| Q75GX9 | 63 kDa globulin-like protein |
| Q6K7K6 | Glutelin |
| A1YQG5 | Glutelin |
| Q6ESW6 | Glutelin |
| P07728 | Glutelin type-A 1 |
| P07730 | Glutelin type-A 2 |
| Q09151 | Glutelin type-A 3 |
| P14323 | Glutelin type-B 1 |
| Q02897 | Glutelin type-B 2 |
| Q8S0E1 | Patatin |
| Q65XA1 | Putative legumin |
| Q0DS36 | Vicilin-like seed storage protein |
| Stress Response | |
| Q8H920 | AWPM-19-like family protein |
| Q6Z7B0 | Heat shock 70 kDa protein BIP1 |
| Q75LL0 | Putative stress-related protein |
| Transcriptional/Translational Machinery | |
| O64937 | Elongation factor 1-alpha |
| Q6L500 | Probable histone H2A.4 |
| Q2QS71 | Probable histone H2A.7 |
| UniProt ID | Description | Score | Coverage (%) a | # Peptides b |
|---|---|---|---|---|
| Actin | ||||
| Q10DV7 | Actin-1 | 85.957 | 51.1936 | 15(3) |
| Q75HX0 | Actin-1 | 79.968 | 44.9468 | 12(9) |
| A3C6D7 | Actin-2 | 77.034 | 50.9284 | 14(3) |
| Q67G20 | Actin-7 | 71.587 | 46.6843 | 13(1) |
| Q10DV7 | Actin-1 | 27.620 | 38.1963 | 9(1) |
| Q75HX0 | Actin-1 | 20.401 | 22.3404 | 6(4) |
| P0C540 | Actin-7 | 14.061 | 21.0106 | 6(1) |
| Q9AY76 | Actin-depolymerizing factor 2 | 13.516 | 22.0690 | 2(2) |
| Q84TB6 | Actin-depolymerizing factor 3 | 6.3583 | 34.6667 | 3(3) |
| Q84TB3 | Actin-depolymerizing factor 4 | 4.7321 | 11.5108 | 1(1) |
| Q0DLA3 | Actin-depolymerizing factor 7 | 1.5514 | 8.63309 | 1(1) |
| Q84TB6 | Actin-depolymerizing factor 3 | 1.5320 | 9.33333 | 1(1) |
| Lectin | ||||
| Q10M12 | Ricin B-like lectin R40C1-domain containing | 42.362 | 49.1379 | 12(10) |
| Membrane-Associated | ||||
| Q75GB3 | Outer membrane protein | 11.295 | 6.26896 | 4(4) |
| Structural | ||||
| Q0D3Z9 | Transport protein SEC31 homolog B | 48.342 | 14.6406 | 11(11) |
| Q7EYR6 | Prohibitin-2 | 32.952 | 24.9135 | 5(2) |
| Q654U5 | Phragmoplastin | 19.075 | 7.24479 | 4(4) |
| Q9AWU6 | WD-repeat containing protein 1 | 16.142 | 10.1639 | 4(4) |
| Q5N7E8 | Microtubule binding motor protein | 9.2205 | 13.3080 | 3(3) |
| Q2QX21 | Myotonica WD repeat-containing protein | 5.8419 | 4.08526 | 1(1) |
| Q6ZIG6 | Myosin heavy-chain related protein | 2.0988 | 1.96592 | 1(1) |
| Q5NBL8 | Klaroid, isoform A-related | 1.6375 | 4.61539 | 1(1) |
| Reticulata Related-Like | ||||
| Q5JK51 | Reticulata-related 4-like | 1.4744 | 2.57069 | 1(1) |
| Q5VQR0 | Reticulata-related 3-like | 20.840 | 20.6896 | 4(4) |
| Starch granule-binding | ||||
| Q6YXZ6 | Glucan endo-1,3-beta-glucosidase 6 | 5.8419 | 4.08526 | 1(1) |
| Q10F03 | FLOURY6 | 2.0380 | 1.51229 | 1(1) |
| Tegument | ||||
| Q0JF82 | Transport protein Sec24-like | 19.287 | 7.75946 | 5(5) |
| Q0JF82 | Transport protein Sec24-like | 5.5103 | 4.26770 | 2(2) |
| Q5JML5 | Altered inheritance of mitochondria protein 3-like | 2.6716 | 2.86195 | 1(1) |
| Tubulin and Tubulin-Like | ||||
| P46265 | Tubulin beta-5 chain-like | 71.042 | 32.4385 | 12(2) |
| A3ANA0 | Tubulin beta-7 chain | 70.215 | 32.6577 | 12(2) |
| P45960 | Tubulin beta-4 chain-like | 66.176 | 30.4251 | 11(2) |
| Q75GI3 | Tubulin alpha-1 chain | 60.193 | 40.3548 | 11(7) |
| A3AL37 | Tubulin beta chain | 56.060 | 24.8918 | 9(1) |
| P37832 | Tubulin beta-7 chain | 42.446 | 30.8559 | 12(12) |
| Q0PVB0 | Tubulin alpha-1 chain-like | 37.643 | 29.7778 | 8(4) |
| Q53M52 | Tubulin alpha-2 chain-like | 18.417 | 13.5255 | 4(2) |
| Q10PW2 | Tubulin alpha chain, putative | 15.823 | 22.4944 | 6(4) |
| P28752 | Tubulin alpha-1 chain | 15.433 | 13.5556 | 4(2) |
| Q10PW2 | Tubulin alpha chain, putative | 3.8827 | 3.11804 | 1(1) |
| UniProt ID | Description | Score | Coverage (%) a | # Peptides b |
|---|---|---|---|---|
| Actin | ||||
| Q10DV7 | Actin-1 | 27.619 | 38.1962 | 1(1) |
| Q10AZ4 | Actin-3 | 1.2836 | 6.10079 | 1(1) |
| Tubulin | ||||
| Q53M52 | Tubulin alpha-2 chain | 1.5231 | 3.32594 | 1(1) |
| P45960 | Tubulin beta-4 chain | 1.5136 | 8.50111 | 2(2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Provost, Z.; Hansen, E.O.; Lynds, M.V.; Flinn, B.S.; Minic, Z.; Berezovski, M.V.; Altosaar, I. Three Diverse Granule Preparation Methods for Proteomic Analysis of Mature Rice (Oryza sativa L.) Starch Grain. Molecules 2022, 27, 3307. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103307
Provost Z, Hansen EO, Lynds MV, Flinn BS, Minic Z, Berezovski MV, Altosaar I. Three Diverse Granule Preparation Methods for Proteomic Analysis of Mature Rice (Oryza sativa L.) Starch Grain. Molecules. 2022; 27(10):3307. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103307
Chicago/Turabian StyleProvost, Zachary, Ella Olivia Hansen, Morgan Viola Lynds, Barry S. Flinn, Zoran Minic, Maxim V. Berezovski, and Illimar Altosaar. 2022. "Three Diverse Granule Preparation Methods for Proteomic Analysis of Mature Rice (Oryza sativa L.) Starch Grain" Molecules 27, no. 10: 3307. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103307