
Cloning and Functional Characterization of Two Germacrene A Oxidases Isolated from Xanthium sibiricum

 , ,
, ,
Abstract
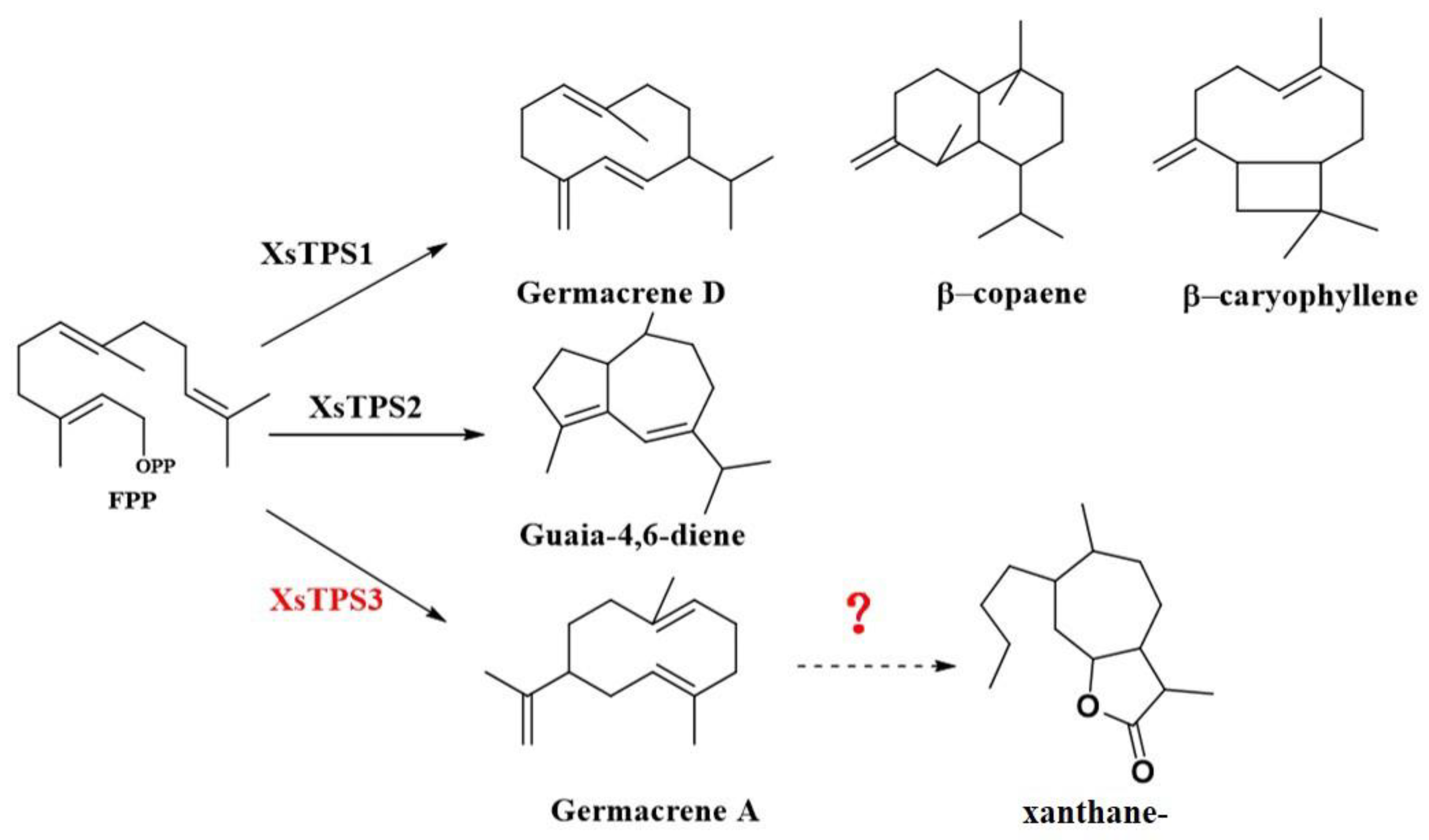
:1. Introduction
2. Results
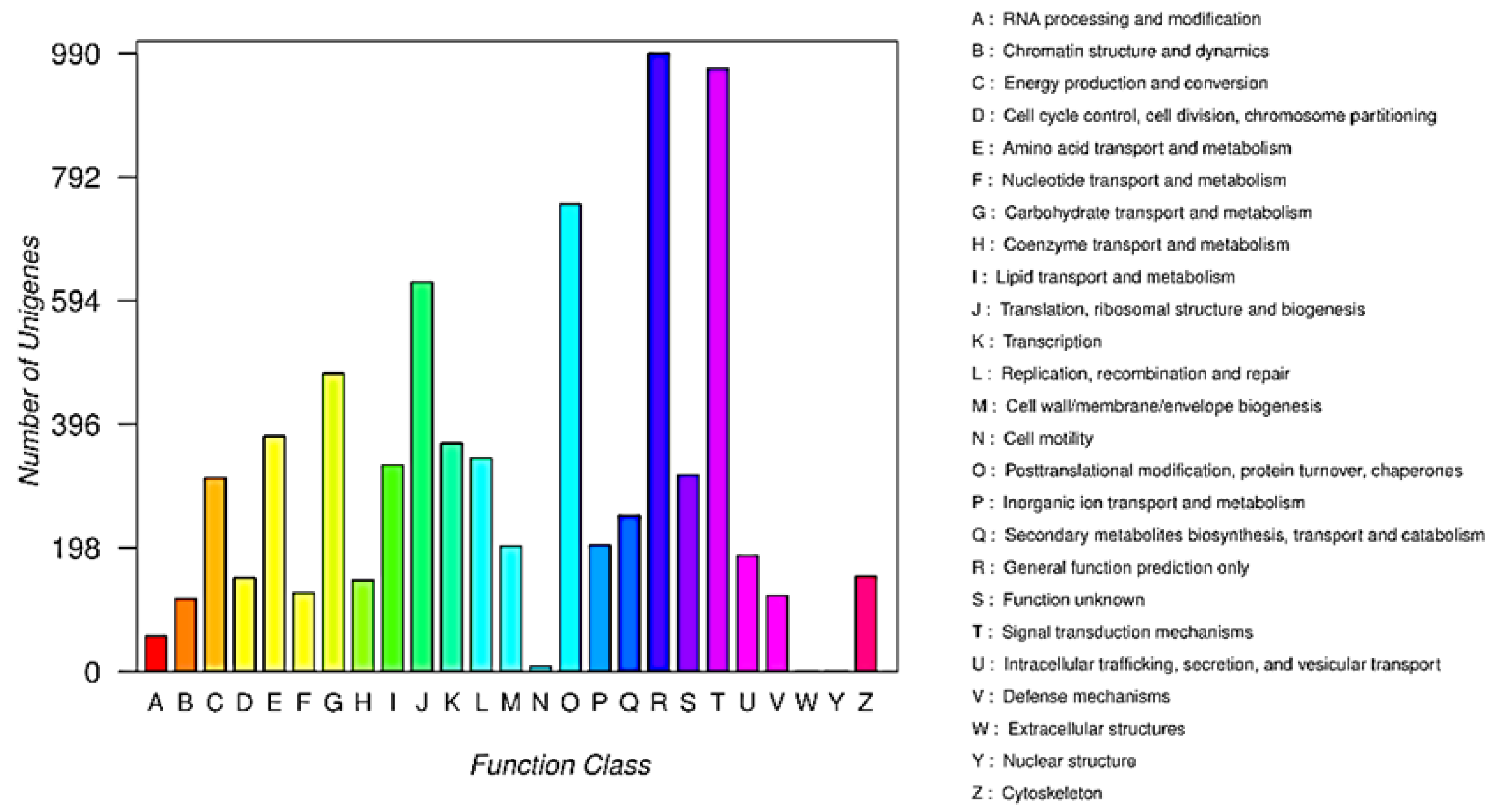
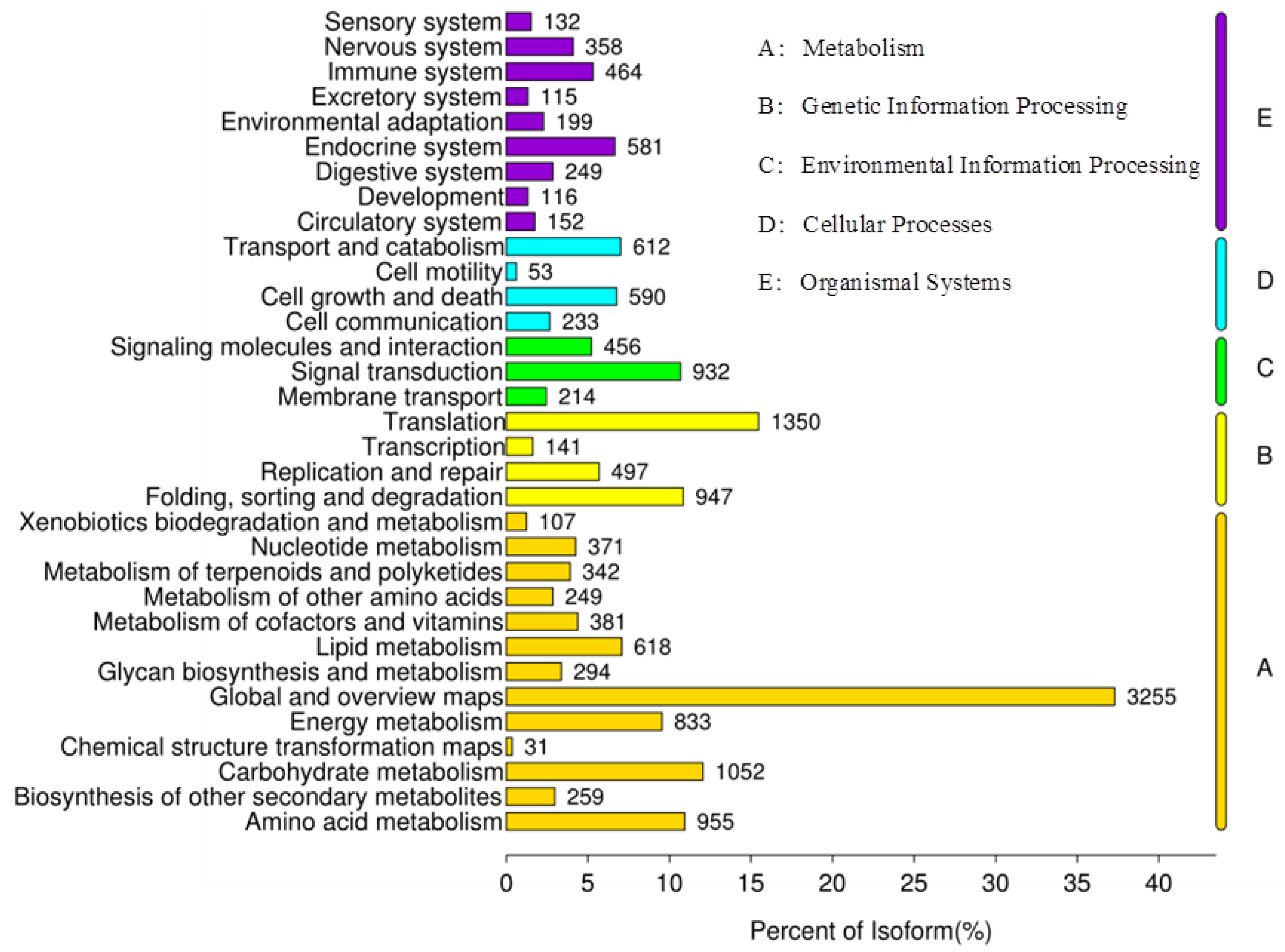
2.1. Establishment of a Transcriptome Library and Gene Annotation of X. sibiricum
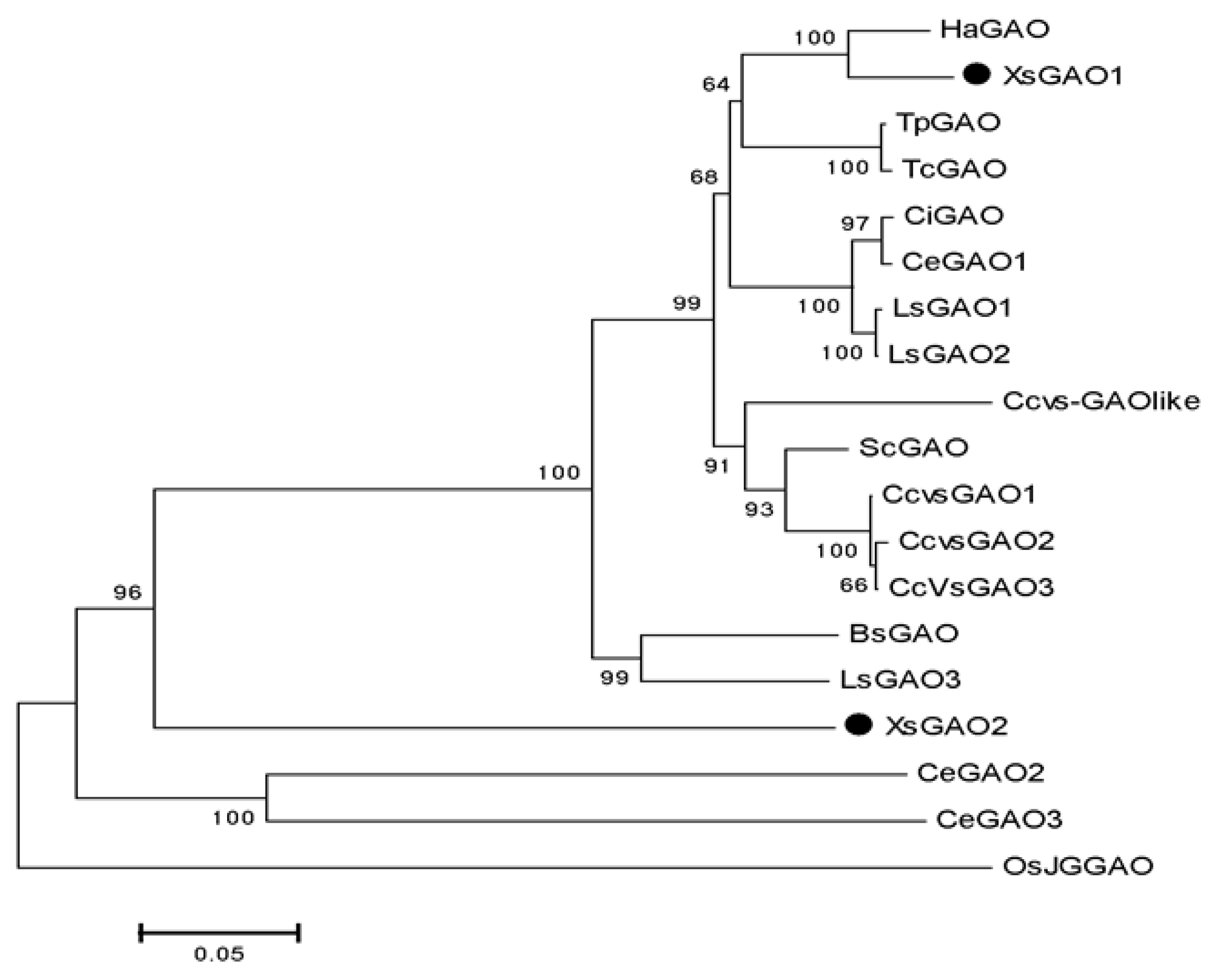
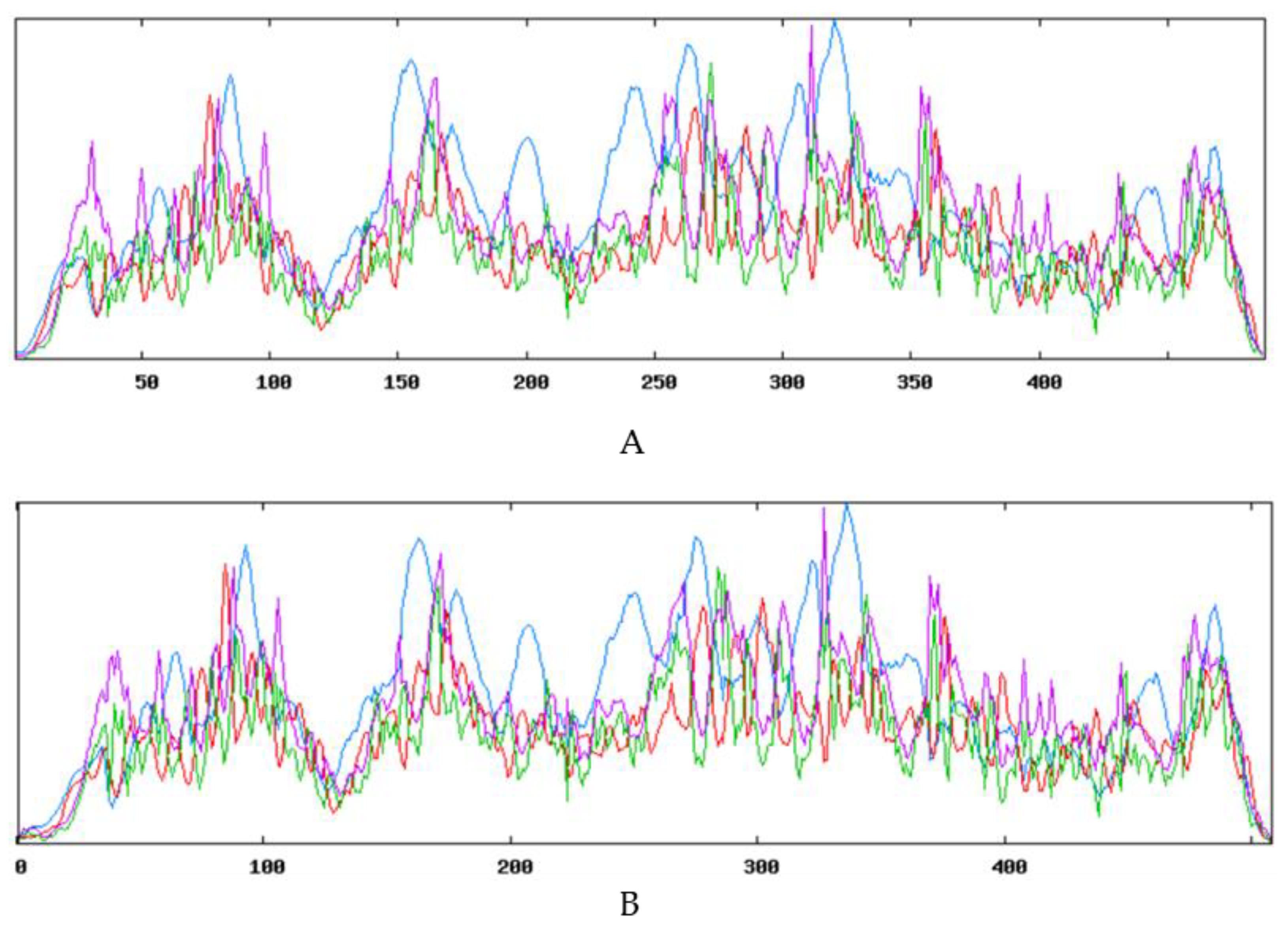
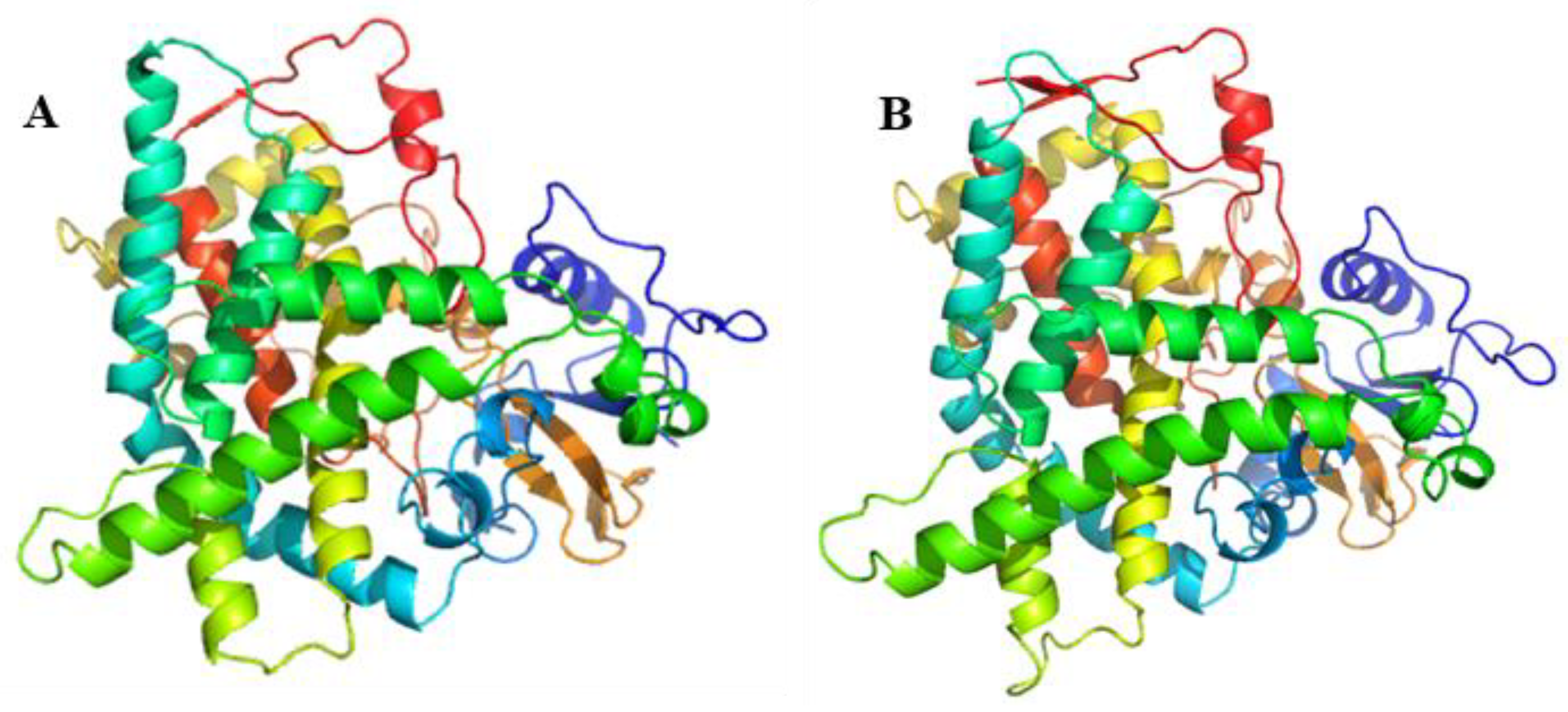
2.2. Cloning and Bioinformatics Analysis of XsGAO Genes
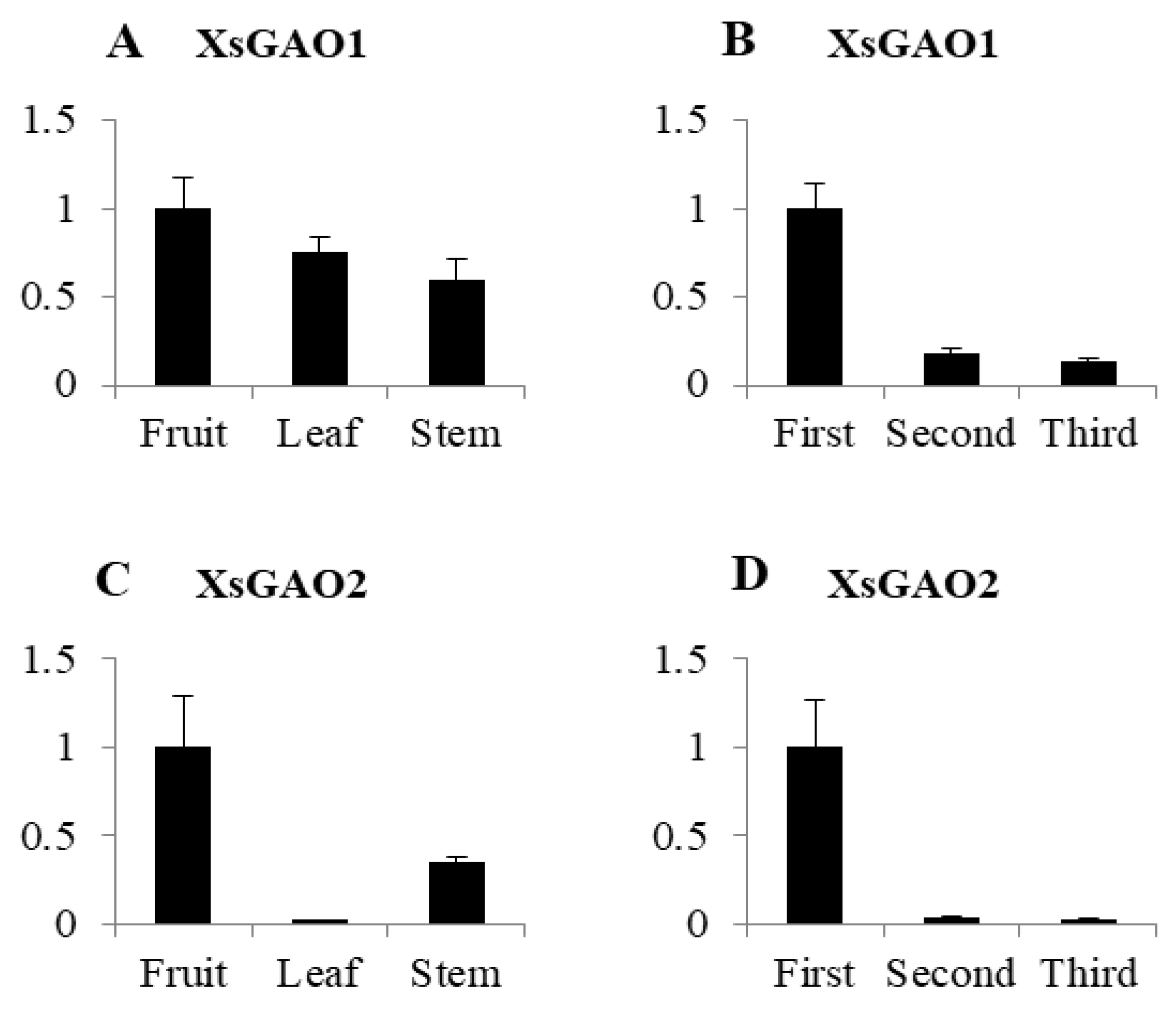
2.3. XsGAO1 and XsGAO2 Expression Patterns in X. sibiricum
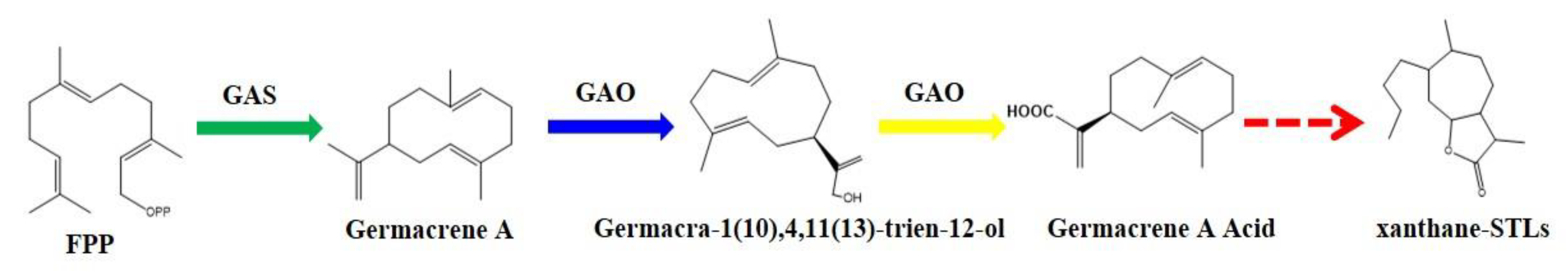
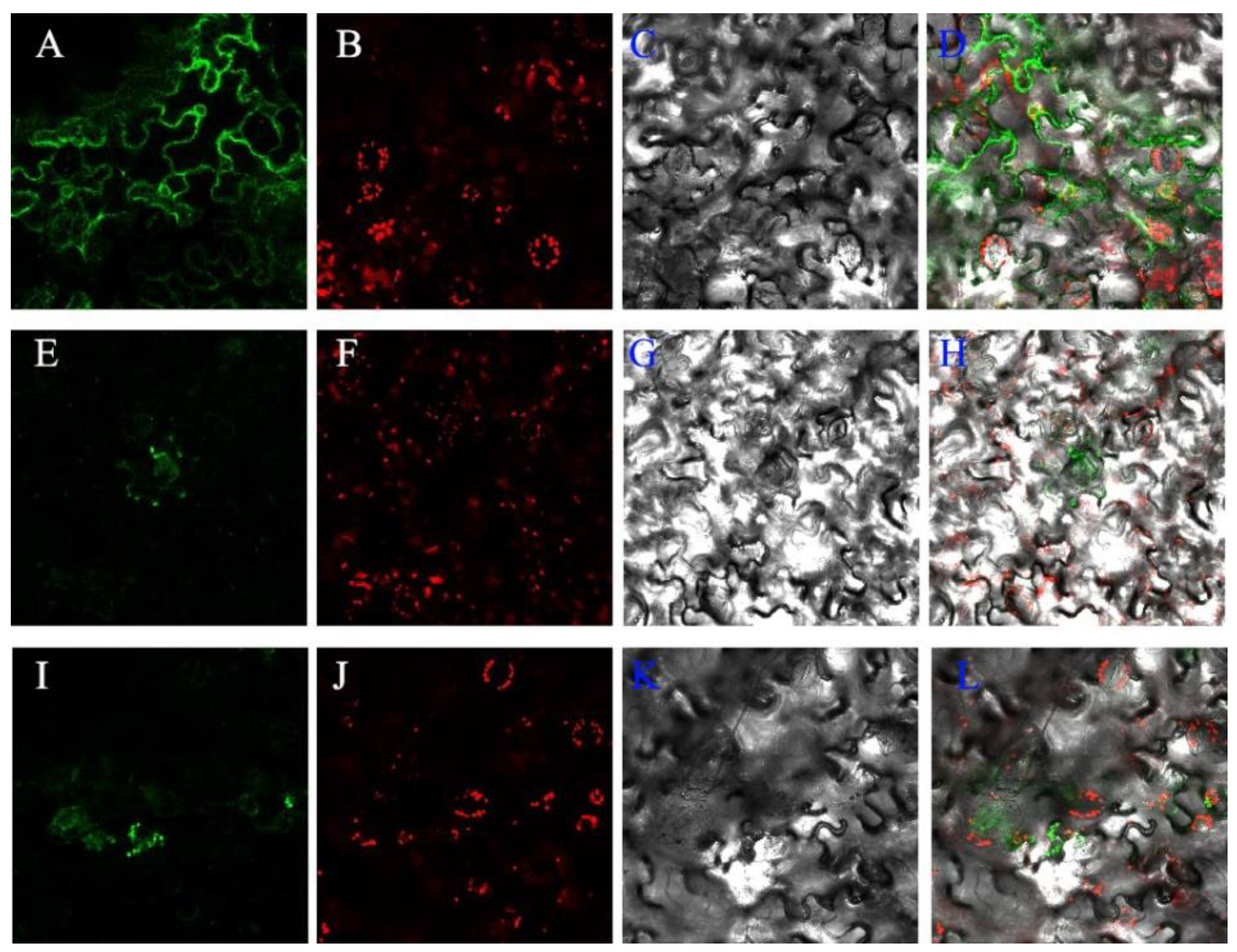
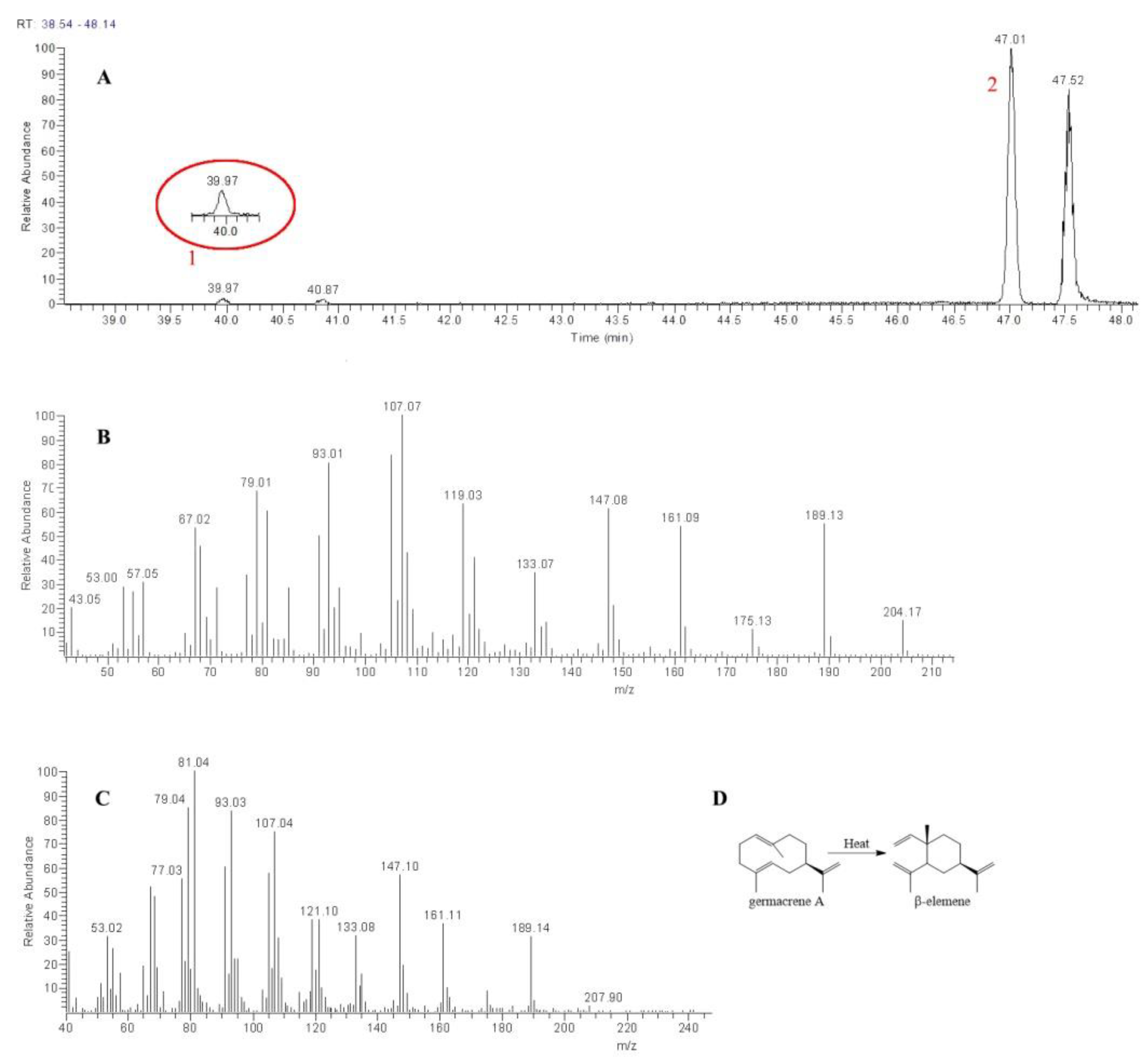
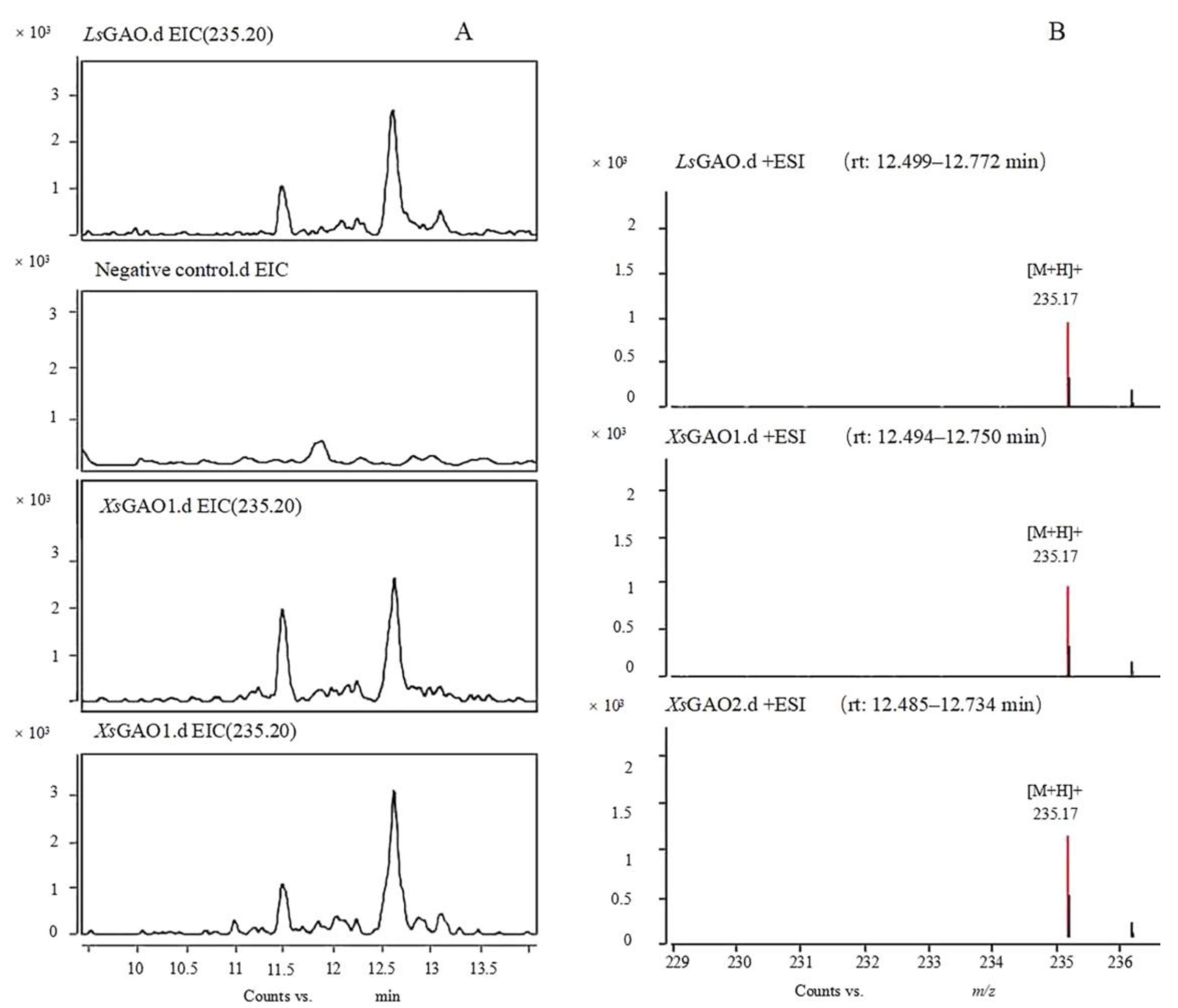
2.4. Functional Study of XsGAO1 and XsGAO2
3. Discussion
4. Materials and Methods
4.1. Establishment of a Transcriptome Library and Gene Annotation
4.2. Cloning and Bioinformatics Analysis of XsGAO1 and XsGAO2
4.3. Examination of the Expression Patterns of the XsGAO1 and XsGAO2 Genes
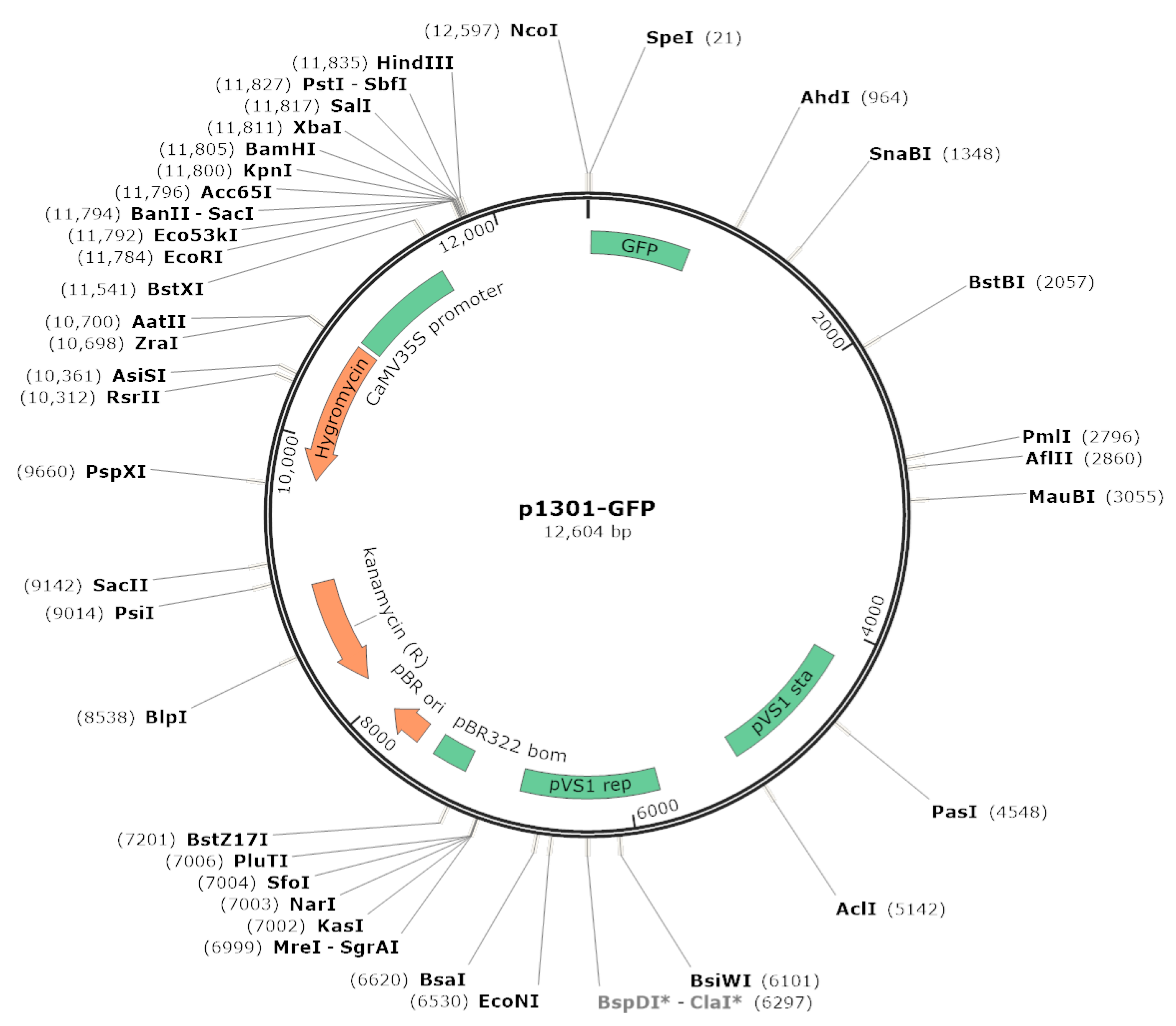
4.4. Functional Study of XsGAO1 and XsGAO2 Genes in Yeast
4.5. GC-MS and LC-MS Analyses
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence |
|---|---|
| XsGAO1F | ATGGAAGTCTCCCTCACCACTTC |
| XsGAO1R | TTAAAAACTTGGTACCAATATCAACCC |
| XsGAO2F | ATGGAACTCCATTTTCCCAC |
| XsGAO2R | TCCAAATATCACTATCCTTCG |
| M13F | CAGGAAACAGCTATGAC |
| M13R | GTAAAACGACGGCCAGT |
| Primer | Primer Sequence |
|---|---|
| QXsGAO1F | CTAATAAGGTGTCCGAGAG |
| QXsGAO1R | GGCAGGTCTTGAATATCT |
| QXsGAO2F | TCTCAACCATAGTAATCTCA |
| QXsGAO2R | CGATGTCTGTGTAATTGTAT |
| ActinF | TACTACAACGGCAGAACGGGAAA |
| ActinR | TCATAGACGGCTGGAACAAAACC |
| GFP-XsGAO1F | acgggggactcttgaccatggATGGAAGTCTCCCTCACCACTTC |
| GFP-XsGAO1R | gcccttgctcaccatactagtAAAACTTGGTACCAATATCAACCCA |
| GFP-XsGAO2F | acgggggactcttgaccatggATGGAACTCCATTTTCCCACC |
| GFP-XsGAO2R | gcccttgctcaccatactagtCATTGTGTTGTAAGGTGTTGGGA |
| LsGASF | ATGGCAGCAGTTGACACTAATG |
| LsGASR | TTACATGGATACAGAACCAAC |
| LsGAOF | ATGGAGCTTTCAATAACCACC |
| LsGAOR | CTAAAAACTCGGTACGAGTAACAAC |
| pYeDP60-XsGAO1F | acacactaaattaccggatccATGGAAGTCTCCCTCACCACTTC |
| pYeDP60-XsGAO1R | gggagatcccccgcggaattcTTAAAAACTTGGTACCAATATCAACCC |
| pYeDP60-XsGAO2F | acacactaaattaccggatccATGGAACTCCATTTTCCCACC |
| pYeDP60-XsGAO2R | gggagatcccccgcggaattcTCACATTGTGTTGTAAGGTGTTGG |
| pYeDP60-LsGAOF | acacactaaattaccggatccATGGAGCTTTCAATAACCACCTCC |
| pYeDP60-LsGAOR | gggagatcccccgcggaattcCTAAAAACTCGGTACGAGTAACAACTC |
| Name | Species | Gene Bank Accession No. |
|---|---|---|
| CcVsGAO1 | Cynara cardunculus var. scolymus | AIA09035.1 |
| CcVsGAO2 | XP_024977750.1 | |
| CcVsGAO3 | AIA09037.1 | |
| CcVs-GAOlike | XP_024977969.1 | |
| TpGAO | Tanacetum parthenium | AHN62855.1 |
| LsGAO1 | Lactuca sativa | XP_023734551.1 |
| LsGAO2 | ADF32078.1 | |
| LsGAO3 | AIX97103.1 | |
| BsGAO | Barnadesia spinosa | ADF43083.1 |
| HaGAO | Helianthus annuus | ADF43082.1 |
| ScGAO | Saussurea costus | ADF43081.1 |
| CiGAO | Cichorium intybus | ADF43080.1 |
| TcGAO | Tanacetum cinerariifolium | AGO03789.1 |
| CeGAO1 | Cichorium endivia | AZI95573.1 |
| CeGAO2 | AZI95575.1 | |
| CeGAO3 | AZI95574.1 | |
| OsJGGAO | Oryza sativa Japonica Group | XP_015624875.1 |









References
- Guo, L.M.; Lv, J.L.; Zhang, L.B. Research progress on anti-inflammatory mechanism of natural sesquiterpenoids. China J. Chin. Mater. Med. 2018, 43, 3989–3999. [Google Scholar]
- Xu, D.; Liu, D.; Zhang, C.H.; Zhang, R.F.; Wang, Q.L.; Zou, K.; Wang, H. Antitumor constituents from roots of Ligularia veitchiana. J. Cent. China Norm. Univ. (Nat. Sci.) 2021, 55, 72–75. [Google Scholar]
- Yan, Z.P.; Xu, T.T.; An, Z.T.; Hu, Y.; Chen, W.Z.; Ma, J.X.; Shao, C.L.; Zhu, F.S. Costunolide induces mitochondria-mediated apoptosis in human gastric adenocarcinoma BGC-823 cells. BMC Complementary Altern. Med. 2019, 19, 151–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, W.; Wang, X.; Li, X.; Li, J.; Li, X. Advances in broad-spectrum antitumor effect of four sesquiterpenoids. Chin. J. Biochem. Pharm. 2016, 36, 5–8. [Google Scholar]
- Jiang, Y.Q.; Dong, Y.J.; Zhou, F.J.; Chen, J.P.; Zhou, Y.T.; Tian, C.W.; Chen, C.Q. Research progress on artemisinin and its derviatives. Chin. Tradit. Herb. Drugs 2022, 53, 599–609. [Google Scholar]
- Ramírez-Erosa, I.; Huang, Y.G.; Hickie, R.A.; Sutherland, R.G.; Barl, B. Xanthatin and xanthinosin from the burs of Xanthium strumarium L. as potential anticancer agents. Can. J. Physiol. Phamacol. 2007, 85, 1160–1172. [Google Scholar] [CrossRef]
- Zhang, L.; Ruan, J.S.; Yan, L.G.; Li, W.D.; Wu, Y.; Tao, L.; Zhang, F.; Zheng, S.Z.; Wang, A.Y.; Lu, Y. Xanthatin Induces Cell Cycle Arrest at G2/M Checkpoint and Apoptosis via Disrupting NF-κB Pathway in A549 Non-Small-Cell Lung Cancer Cells. Molecules 2012, 17, 3736–3750. [Google Scholar] [CrossRef] [Green Version]
- Aranjani, J.M.; Manuel, A.; Rao, C.M.; Udupa, N.; Rao, J.V.; Joy, A.M.; Gandhi, P.; Radhakrishnan, E.K. Preliminary evaluation of in vitro cytotoxicity and in vivo antitumor activity of Xanthium strumarium in transplantable tumors in mice. Am. J. Chin. Med. 2013, 41, 145–162. [Google Scholar] [CrossRef]
- Wang, L.J.; Fang, X.; Yang, C.Q.; Li, J.X.; Chen, X.Y. Biosynthesis and regulation of secondary terpenoid metabolism in plants. Scienta Sinica Vitae. 2013, 43, 1030–1046. [Google Scholar]
- Gou, J.B.; Hao, F.H.; Huang, C.Y.; Kwon, M.; Chen, F.F.; Li, C.F.; Liu, C.Y.; Ro, D.; Tang, H.R.; Zhang, Y.S. Discovery of a non-stereoselective cytochrome P450 catalyzing either 8α- or 8β-hydroxylation of germacrene A acid from the Chinese medicinal plant, Inula hupehensis. Plant J. 2018, 93, 92–106. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Kashkooli, A.B.; Manzano, D.; Pateraki, I.; Richard, L.; Kolkman, P.; Lucas, L.F.; Guallar, V.; Vos, R.C.; Franssen, M.C.R.; et al. Kauniolide synthase is a P450 with unusual hydroxylation and cyclization-elimination activity. Nat. Commun. 2018, 9, 4657. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.; Hemmerlin, A.; Bach, T.J.; Chye, M.L. The potential of the mevalonate pathway for enhanced isoprenoid production. Biotechnol. Adv. 2016, 34, 697–713. [Google Scholar] [CrossRef] [PubMed]
- Zong, Z.; Cheng, L.; Chen, Z.J.; Wang, L.; Wang, C.; Qi, Y.G.; Liu, A.J. Research progress on biosynthesis of terpenoids for food. China Brew. 2018, 37, 22–27. [Google Scholar]
- Shi, M.; Wang, Y.; Zhou, W.; Hua, Q.; Kai, G.Y. Research progress in terms of the biosynthesis and regulation of terpenoids from medicinal plants. Sci. Sin. Vitae 2018, 48, 352–364. [Google Scholar]
- Xiao, L.; Chen, R.B.; Wu, Y.; Zhang, L. Research progress on effect of AP2/ERF transcription factors in regulating secondary metabolite biosynthesis. China J. Chin. Mater. Med. 2020, 45, 5412–5420. [Google Scholar]
- Little, D.B.; Croteau, R.B. Alteration of product formation by directed mutagenesis and truncation of the multiple-product sesquiterpene synthases δ-selinene synthase and γ-humulene synthase. Arch. Biochem. Biophys. 2002, 402, 120–135. [Google Scholar] [CrossRef]
- Yoshikuni, Y.; Martin, V.J.; Ferrin, T.E.; Keasling, J.D. Engineering cotton (+)-δ-cadinene synthase to an altered function: Germacrene D-4-ol synthase. Chem. Biol. 2006, 13, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Frey, M.; Schmauder, K.; Paterki, I.; Spring, O. Biosynthesis of Eupatolide-A metabolic route for sesquiterpene lactone formation involving the P450 enzyme CYP71DD6. ACS Chem. Biol. 2018, 13, 1536–1543. [Google Scholar] [CrossRef]
- Wallaart, T.E.; Bouwmeester, H.J.; Hille, J.; Poppinga, L.; Maijers, N.C. Amorpha-4,11-diene synthase: Cloning and functional expression of a key enzyme in the biosynthetic pathway of the novel antimalarial drug artemisinin. Planta 2001, 212, 460–465. [Google Scholar] [CrossRef] [Green Version]
- Kraker, J.M.; Franssen, M.C.; Joerink, M.; Groot, A.D.; Bouwmeester, H.J. Biosynthesis of costunolide, dihydrocostunolide, and leucodin. Demonstration of cytochrome P450-catalyzed formation of the lactone ring present in sesquiterpene lactones of chicory. Plant Physiol. 2002, 129, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Manzano, D.; Tanic, N.; Pesic, M.; Bankovic, J.; Pateraki, I.; Ricard, L.; Ferrer, A.; Vos, R.D.; Krol, S.V.D.; et al. Elucidation and in planta reconstitution of the parthenolide biosynthetic pathway. Metab. Eng. 2014, 23, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Chen, F.F.; Li, Z.Q.; Zhang, Y.S. Identification and functional characterization of sesquiterpene synthases from Xanthium strumarium. Plant Cell Physiol. 2016, 57, 630–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Lin, F.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.T.; Gopfert, J.C.; Ikezawa, N.; MacNevin, G.; Kathiresan, M.; Gonrad, J.; Spring, O.; Ro, D.K. Biochemical conservation and evolution of germacrene a oxidase in Asteraceae. J. Biol. Chem. 2010, 285, 16588–16598. [Google Scholar] [CrossRef] [Green Version]
- Hui, F.; Liu, X.Y.; Li, Z.Y.; Liu, F.S.; Yang, S.H. Application of transcriptome sequencing in study of medicinal plants. Chin. Tradit. Herb. Drugs 2019, 50, 6149–6155. [Google Scholar]
- Li, Y.J. Cloning and Functional Analysis of Genes Involved in the Biosynthesis Sesquiterpene Lactones in Xanthium strumarium L. Ph.D. Thesis, Chinese Academy of Sciences, Beijing, China, 2017. [Google Scholar]
- Chen, F.F.; Hao, F.H.; Li, C.F.; Gou, J.B.; Lu, D.Y.; Gong, F.J.; Tang, H.R.; Zhang, Y.S. Identifying three ecological chemotypes of Xanthium strumarium glandular trichomes using a combined NMR and LC-MS method. PLoS ONE 2013, 8, 10. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Kwon, M.; Kim, S.U.; Fischer, C.; Ro, D.K. Catalytic plasticity of germacrene A oxidase underlies sesquiterpene lactone diversification. Plant Physiol. 2019, 181, 945–960. [Google Scholar] [CrossRef] [Green Version]
- Hamberger, B.; Bak, S. Plant P450s as versatile drivers for evolution of species-specific chemical diversity. Philos. Trans. R. Soc. B 2013, 368, 20120426. [Google Scholar] [CrossRef] [Green Version]
- Kraker, J.W.D.; Franssen, M.C.R.; Groot, A.D.; Shibata, T.; Bouwmeester, H.J. Germacrenes from fresh costus roots. Phytochemistry 2011, 58, 481–487. [Google Scholar] [CrossRef]
- Falara, V.; Akhtar, T.A.; Nguyen, T.T.; Spyropoulou, E.A.; Bleeker, P.M.; Schauvinhold, I.; Matsuba, Y.; Bonini, M.E.; Schilmiller, A.L.; Last, A.L.; et al. The tomato terpene synthase gene family. Plant Physiology. 2011, 157, 770–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappers, I.F.; Aharoni, A.; Herpen, T.W.; Luckerhoff, L.L.; Dicke, M. Genetic engineering of terpenoid metabolism attracts bodyguards to Arabidopsis. Science 2005, 309, 2070–2072. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.H.; Mansfield, J.W.; Lewis, M.J.; Beale, M.H. Cloning and expression of sesquiterpene synthase genes from lettuce (Lactuca satica L.). Phytochemistry 2002, 60, 255–261. [Google Scholar] [CrossRef]
- Liu, Q.; Majdi, M.; Cankar, K.; Goedbloed, M.; Charnikhova, T.; Verstappen, F.W.A.; Vos, R.C.H.D.; Beekwilder, J.; Krol, S.V.D.; Bouwmeester, H.J. Reconstitution of the costunolide biosynthetic pathway in yeast and Nicotiana benthamiana. PLoS ONE 2011, 6, e23255. [Google Scholar] [CrossRef]
- Zheng, L.F.; Chen, F.F.; Li, Y.J.; Li, C.F. Identification of a unique germacrene A oxidase from Xanthium strumarium. Phyton-Int. J. Exp. Bot. 2022, 91, 989–998. [Google Scholar] [CrossRef]
- Swinnen, G.; Jacobs, T.; Pauwels, L.; Goossens, A. CRISPR-Cas-Mediated Gene Knockout in Tomato. Methods Mol. Biol. 2020, 2083, 321–341. [Google Scholar]
- Li, B.; Cui, G.; Shen, G.; Zhan, Z.; Huang, L.; Chen, J.; Qi, X. Targeted mutagenesis in the medicinal plant Salvia miltiorrhiza. Sci. Rep. 2017, 7, 43320. [Google Scholar] [CrossRef] [Green Version]
- Shi, T.L.; Zhang, L.; Cheng, Q.Y.; Yu, J.S.; Liu, J.; Shen, Y.J. Xanthatin induces apoptosis by activating endoplasmic reticulum stress in hepatoma cells. Eur. J. Pharmacol. 2019, 843, 1–11. [Google Scholar] [CrossRef]
- Wang, L.; Wang, J.; Li, F.; Liu, X.; Chen, B.; Tang, Y.X.; Wang, M.K. Cytotoxic sesquiterpene lactones from aerial parts of Xanthium sibiricum. Planta Med. 2013, 79, 661–665. [Google Scholar] [CrossRef] [Green Version]
- Yao, S.B.; Wang, W.Z.; Li, Y.J.; Wang, Y.S.; Li, Y.J.; Gao, L.P.; Xia, T. The gene cloning and expression analysis of C4H in tea plant (Camellia sinensis). J. Tea Sci. 2015, 1, 35–44. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; Mcwilliam, H.; Valentin, F.; Wallace, I.; Wilm, A.; Lppez, R. Clustal W and clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.N.; Wang, Q.Y.; Deng, J.; Ming, S.F.; Gong, L.; Yu, K.; Xu, J.Z.; Wu, N. Cloning and analysis of two HMGR genes (AIHMGR) in Atractylodes lancea. Genom. Appl. Biol. 2020, 4, 1732–1740. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, D.-M.; Zhang, Q.; Xin, L.-K.; Wang, G.-K.; Liu, C.-B.; Qin, M.-J. Cloning and Functional Characterization of Two Germacrene A Oxidases Isolated from Xanthium sibiricum. Molecules 2022, 27, 3322. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103322
Xie D-M, Zhang Q, Xin L-K, Wang G-K, Liu C-B, Qin M-J. Cloning and Functional Characterization of Two Germacrene A Oxidases Isolated from Xanthium sibiricum. Molecules. 2022; 27(10):3322. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103322
Chicago/Turabian StyleXie, Dong-Mei, Qiang Zhang, Ling-Kai Xin, Guo-Kai Wang, Cong-Bin Liu, and Min-Jian Qin. 2022. "Cloning and Functional Characterization of Two Germacrene A Oxidases Isolated from Xanthium sibiricum" Molecules 27, no. 10: 3322. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103322