
The Effect of Standardised Leaf Extracts of Gaultheria procumbens on Multiple Oxidants, Inflammation-Related Enzymes, and Pro-Oxidant and Pro-Inflammatory Functions of Human Neutrophils

 , ,
, ,
Abstract
:1. Introduction
2. Results
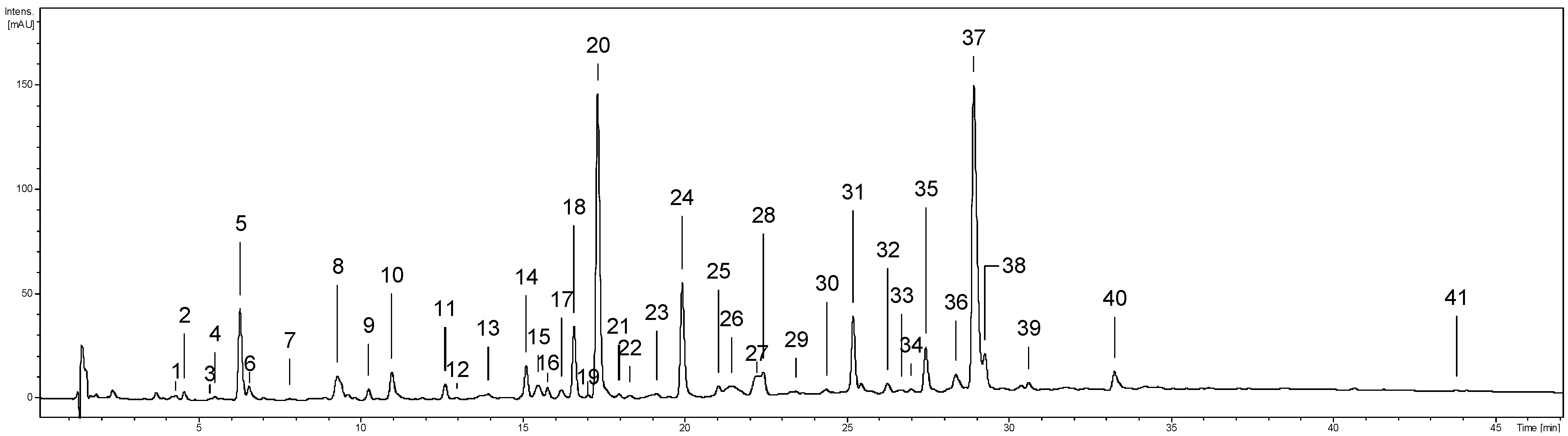
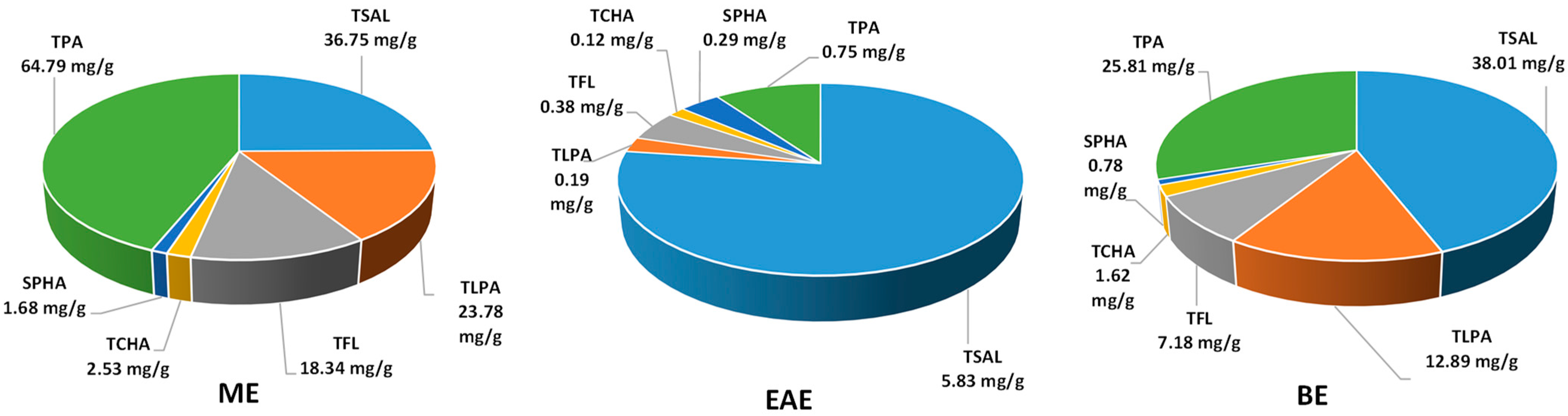
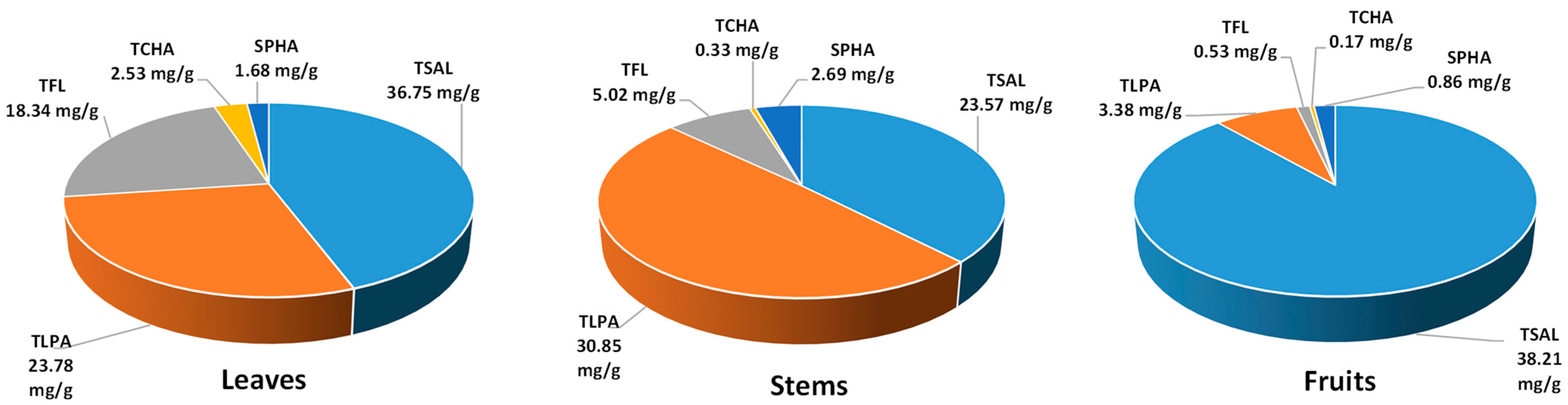
2.1. Phytochemical Standardisation of the Leaf Extracts
2.2. Antioxidant and Anti-Inflammatory Activity in Non-Cellular In Vitro Models
2.3. Influence on Pro-Oxidant and Pro-Inflammatory Functions of Human Neutrophils Ex Vivo
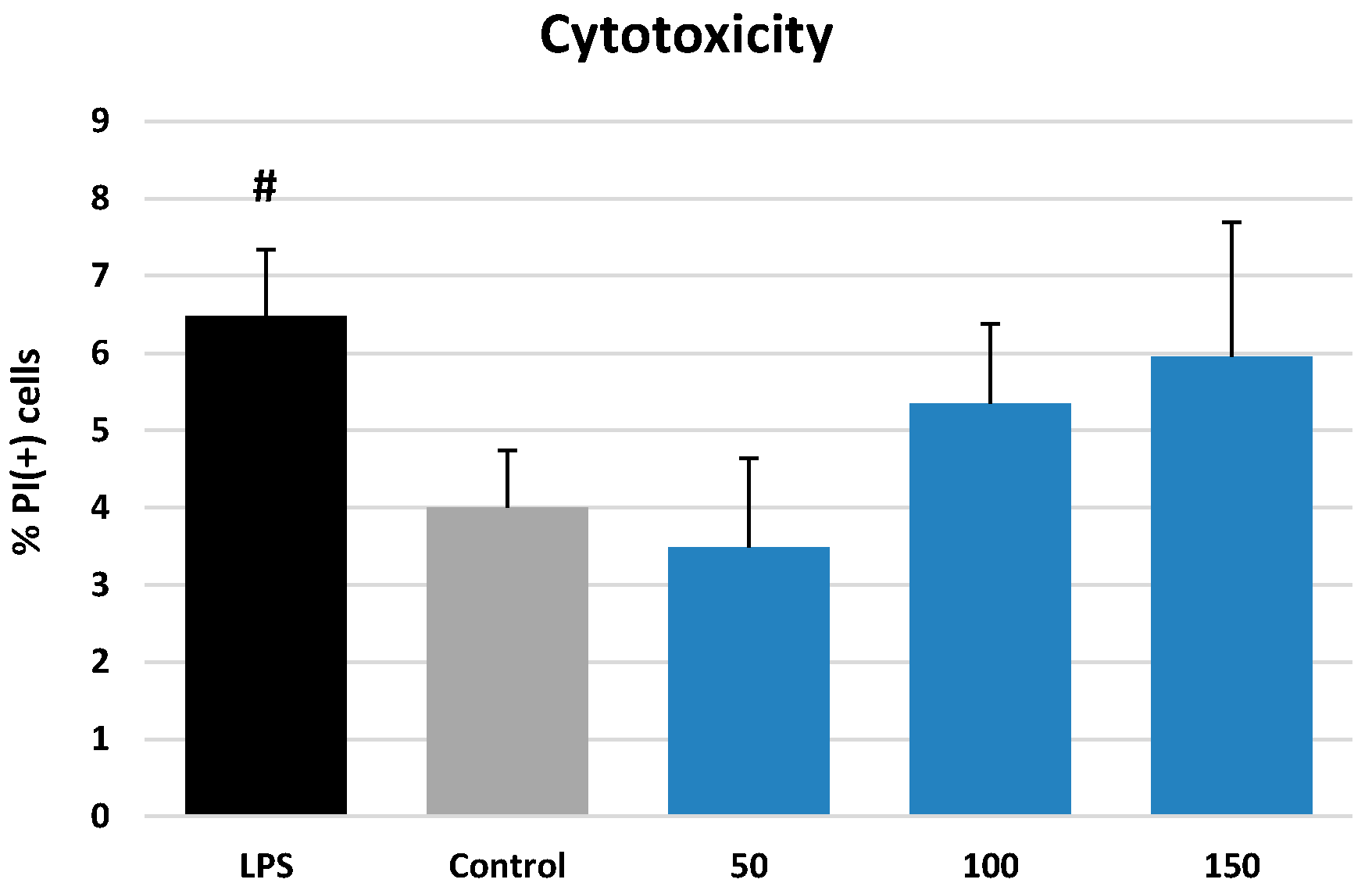
2.3.1. Effects on Neutrophil Viability
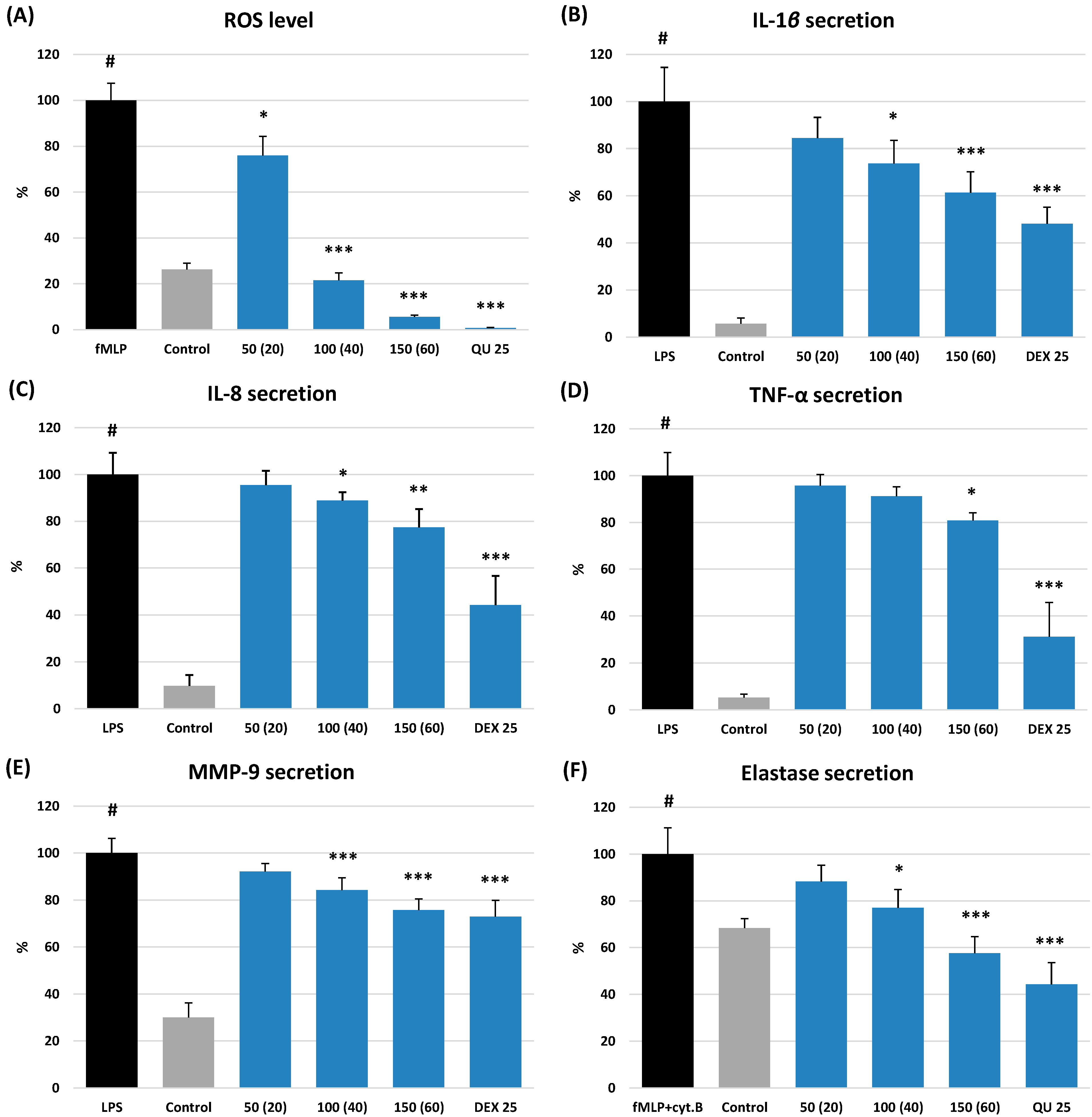
2.3.2. Effects on ROS Production
2.3.3. Effects on the Release of Pro-Inflammatory Cytokines and Enzymes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction
4.2. Qualitative LC–MS/MS Analysis and Quantitative Phytochemical Profiling
4.3. Biological Activity Tests
4.3.1. Non-Cellular In Vitro Models
4.3.2. Cellular Model of Human Neutrophils Ex Vivo
4.4. Statistical and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| BE | n-butanol extract |
| COX-2 | cyclooxygenase 2 |
| CYE | cyanidin chloride equivalents |
| DEX | dexamethasone |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| dw | dry weight |
| EAE | ethyl acetate extract |
| ELA-2 | neutrophils elastase 2 |
| fMLP | N-formyl-l-methionyl-l-leucyl-l-phenylalanine |
| FRAP | ferric reducing antioxidant power |
| GAE | gallic acid equivalents |
| H2O2 | hydrogen peroxide |
| HYAL | hyaluronidase |
| IND | indomethacin |
| IL | interleukin |
| LOX | lipoxygenase |
| LPS | bacterial lipopolysaccharide |
| ME | methanol–water (75:25, v/v) extract |
| MMP-9 | matrix metalloproteinase 9 |
| O2•− | superoxide anion |
| •OH | hydroxyl radical |
| QU | quercetin |
| ROS | reactive oxygen species |
| SAAVNA | N-succinyl-alanine-alanine-valine p-nitroanilide |
| SPHA | total content of simple hydroxybenzoic and hydroxycinnamic acids (HPLC) |
| TBARS | thiobarbituric acid-reactive substances |
| TCHA | total content of chlorogenic acid isomers (HPLC) |
| TFL | total content of flavonoids (HPLC) |
| TLPA | total content of low-molecular-weight flavanols and proanthocyanidins (HPLC) |
| TNF-α | tumour necrosis factor α |
| TPA | total proanthocyanidin content (n-butanol/HCl assay) |
| TPC | total phenolic content (Folin-Ciocalteu assay) |
| TPH | total phenolic content (HPLC) |
| TPHA | total content of phenolic acids (HPLC) |
| TSAL | total content of salicylates (HPLC) |
| TX | (±)-6-hydroxy-2,2,7,8-tetramethylchroman-2-carboxylic acid (Trolox®) |
References
- Middleton, D.J. Infrageneric classification of the genus Gaultheria L. (Ericaceae). Bot. J. Linn. Soc. 1991, 106, 229–258. [Google Scholar] [CrossRef]
- Middleton, D.J. A systematic survey of leaf and stem anatomical characters in the genus Gaultheria and related genera (Ericaceae). Bot. J. Linn. Soc. 1993, 113, 199–215. [Google Scholar] [CrossRef]
- Middleton, D.J. A chemotaxonomic survey of flavonoids and simple phenols in the leaves of Gaultheria L. and related genera (Ericaceae). Bot. J. Linn. Soc. 1992, 110, 313–324. [Google Scholar] [CrossRef]
- Saleem, A.; Harris, C.S.; Asim, M.; Cuerrier, A.; Martineau, L.; Haddad, P.S.; Arnason, J.T. A RP-HPLC-DAD-APCI/MSD method for the characterisation of medicinal Ericaceae used by the Eeyou Istchee Cree First Nations. Phytochem. Anal. 2010, 21, 328–339. [Google Scholar] [CrossRef]
- Cuchet, A.; Jame, P.; Anchisi, A.; Schiets, F.; Oberlin, C.; Lefèvre, J.C.; Carénini, E.; Casabianca, H. Authentication of the naturalness of wintergreen (Gaultheria genus) essential oils by gas chromatography, isotope ratio mass spectrometry and radiocarbon assessment. Ind. Crops Prod. 2019, 142, 111873. [Google Scholar] [CrossRef]
- Magiera, A.; Sienkiewicz, M.; Olszewska, M.A.; Kicel, A.; Michel, P. Chemical profile and antibacterial activity of essential oils from leaves and fruits of Gaultheria procumbens L. cultivated in Poland. Acta Pol. Pharm. Drug Res. 2019, 76, 93–102. [Google Scholar] [CrossRef]
- Liu, W.R.; Qiao, W.L.; Liu, Z.Z.; Wang, X.H.; Jiang, R.; Li, S.Y.; Shi, R.B.; She, G.M. Gaultheria: Phytochemical and pharmacological characteristics. Molecules 2013, 18, 12071–12108. [Google Scholar] [CrossRef]
- Michel, P.; Granica, S.; Magiera, A.; Rosińska, K.; Jurek, M.; Poraj, Ł.; Olszewska, M.A. Salicylate and procyanidin-rich stem extracts of Gaultheria procumbens L. inhibit pro-inflammatory enzymes and suppress pro-inflammatory and pro-oxidant functions of human neutrophils ex vivo. Int. J. Mol. Sci. 2019, 20, 1753. [Google Scholar] [CrossRef] [Green Version]
- Michel, P.; Granica, S.; Rosińska, K.; Rojek, J.; Poraj, Ł.; Olszewska, M.A. Biological and chemical insight into Gaultheria procumbens fruits: A rich source of anti-inflammatory and antioxidant salicylate glycosides and procyanidins for food and functional application. Food Funct. 2020, 11, 7532–7544. [Google Scholar] [CrossRef]
- Koch, W. Dietary polyphenols—important non-nutrients in the prevention of chronic noncommunicable diseases. A systematic review. Nutrients 2019, 11, 1039. [Google Scholar] [CrossRef] [Green Version]
- Carmona, F.; Pereira, A.M.S. Herbal medicines: Old and new concepts, truths and misunderstandings. Rev. Bras. Farmacogn. 2013, 23, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Garg, V.; Dhar, V.J.; Sharma, A.; Dutt, R. Facts about standardization of herbal medicine: A review. J. Chin. Integr. Med. 2012, 10, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Gu, R.; Kennelly, E.J.; Long, C. Gaultheria ethnobotany and bioactivity: Blueberry relatives with anti-inflammatory, antioxidant, and anticancer constituents. Curr. Med. Chem. 2017, 25, 5168–5176. [Google Scholar] [CrossRef] [PubMed]
- Olszewska, M.A.; Owczarek, A.; Magiera, A.; Granica, S.; Michel, P. Screening for the active anti-inflammatory and antioxidant polyphenols of Gaultheria procumbens and their application for standardisation: From identification, through cellular studies to quantitative determination. Int. J. Mol. Sci. 2021, 22, 11532. [Google Scholar] [CrossRef] [PubMed]
- Michel, P.; Dobrowolska, A.; Kicel, A.; Owczarek, A.; Bazylko, A.; Granica, S.; Piwowarski, J.P.; Olszewska, M.A. Polyphenolic profile, antioxidant and anti-inflammatory activity of eastern teaberry (Gaultheria procumbens L.) leaf extracts. Molecules 2014, 19, 20498–20520. [Google Scholar] [CrossRef] [Green Version]
- Michel, P.; Owczarek, A.; Kosno, M.; Gontarek, D.; Matczak, M.; Olszewska, M.A. Variation in polyphenolic profile and in vitro antioxidant activity of eastern teaberry (Gaultheria procumbens L.) leaves following foliar development. Phytochem. Lett. 2017, 20, 356–364. [Google Scholar] [CrossRef]
- Katanić, J.; Pferschy-Wenzig, E.M.; Mihailović, V.; Boroja, T.; Pan, S.P.; Nikles, S.; Kretschmer, N.; Rosić, G.; Selaković, D.; Joksimović, J.; et al. Phytochemical analysis and anti-inflammatory effects of Filipendula vulgaris Moench extracts. Food Chem. Toxicol. 2018, 122, 151–162. [Google Scholar] [CrossRef]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.G.; Lightfoot, D.A. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef]
- Pandey, A.; Tripathi, S.; Pandey, C.A. Concept of standardization, extraction and pre phytochemical screening strategies for herbal drug. J. Pharmacogn. Phytochem. 2014, 115, 115–119. [Google Scholar]
- Voicu, S.N.; Hermenean, A.; Olah, N.K.; Badulescu, L.; Stan, M.S.; Dinischiotu, A. Designing new mother tinctures for increased antioxidant efficiency on inflammatory intestinal diseases. Farmacia 2019, 67, 290–295. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; He, X.L.; Ding, Y.; Du, G.H. Gaultherin, a natural salicylate derivative from Gaultheria yunnanensis: Towards a better non-steroidal anti-inflammatory drug. Eur. J. Pharmacol. 2006, 530, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Cong, F.; Joshi, K.R.; Devkota, H.P.; Watanabe, T.; Yahara, S. Dhasingreoside: New flavonoid from the stems and leaves of Gaultheria fragrantissima. Nat. Prod. Res. 2015, 29, 1442–1448. [Google Scholar] [CrossRef]
- Zhang, B.; Li, J.B.; Zhang, D.M.; Ding, Y.; Du, G.H. Analgesic and anti-inflammatory activities of a fraction rich in gaultherin isolated from Gaultheria yunnanensis (Franch.) Rehder. Biol. Pharm. Bull. 2007, 30, 465–469. [Google Scholar] [CrossRef] [Green Version]
- She, G.M.; Li, D.C.; Zhang, Y.; Guo, Z.Q.; Lv, H.N.; She, D.M. Chemical constituents of aerial parts of Dianbaizhu (Gaultheria leucocarpa var.yunanensis or var.crenulata). J. Beijing Univ. Tradit. Chin. Med. 2010, 33, 62–63. [Google Scholar]
- Liu, Z.; Jiang, R.; Xie, M.; Xu, G.; Liu, W.; Wang, X.; Lin, H.; Lu, J.; She, G. A rapid new approach for the quality evaluation of the folk medicine Dianbaizhu based on chemometrics. Chem. Pharm. Bull. 2014, 62, 1083–1091. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.L.; Liu, Z.Z.; Xie, M.; Zhang, X.; Yang, Y.; Yan, C.; Qiao, W.L.; Liu, W.R.; Jiang, R.; Wang, X.H.; et al. Salicylic acid derivatives and other components from Gaultheria trichoclada. Chem. Nat. Compd. 2016, 52, 301–303. [Google Scholar] [CrossRef]
- Christensen, L.P.; Christensen, K.B. The role of direct and indirect polyphenolic antioxidants in protection against oxidative stress. In Polyphenols in Human Health and Disease, 1st ed.; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 289–309. [Google Scholar]
- Kesavan, P.; Banerjee, A.; Banerjee, A.; Murugesan, R.; Marotta, F.; Pathak, S. An overview of dietary polyphenols and their therapeutic effects. In Polyphenols: Mechanisms of Action in Human Health and Disease, 2nd ed.; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 221–235. [Google Scholar]
- Biswas, S.K. Does the interdependence between oxidative stress and inflammation explain the antioxidant paradox? Oxid. Med. Cell. Longev. 2016, 2016, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and oxidative stress in human diseases: From molecular mechanisms to novel treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipinski, B. Hydroxyl radical and its scavengers in health and disease. Oxid. Med. Cell. Longev. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, H.J.; Traa, A.; Van Raamsdonk, J.M. Beneficial and detrimental effects of reactive oxygen species on lifespan: A comprehensive review of comparative and experimental studies. Front. Cell Dev. Biol. 2021, 9, 181. [Google Scholar] [CrossRef] [PubMed]
- Girish, K.; Kemparaju, K.; Nagaraju, S.; Vishwanath, B. Hyaluronidase inhibitors: A biological and therapeutic perspective. Curr. Med. Chem. 2009, 16, 2261–2288. [Google Scholar] [CrossRef] [PubMed]
- Mashima, R.; Okuyama, T. The role of lipoxygenases in pathophysiology; new insights and future perspectives. Redox Biol. 2015, 6, 297–310. [Google Scholar] [CrossRef] [Green Version]
- Simon, L.S. Role and regulation of cyclooxygenase-2 during inflammation. Am. J. Med. 1999, 106, 37S–42S. [Google Scholar] [CrossRef]
- Sieniawska, E. Activities of tannins—From in vitro studies to clinical trials. Nat. Prod. Commun. 2015, 10, 1877–1884. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.R.; Robb, C.T.; Perretti, M.; Rossi, A.G. The role of neutrophils in inflammation resolution. Semin. Immunol. 2016, 28, 137–145. [Google Scholar] [CrossRef]
- Henriksen, P.A. The potential of neutrophil elastase inhibitors as anti-inflammatory therapies. Curr. Opin. Hematol. 2014, 21, 23–28. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive oxygen species in metabolic and inflammatory signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- Garin, A.; Proudfoot, A.E.I. Chemokines as targets for therapy. Exp. Cell Res. 2011, 317, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood 2011, 117, 3720–3732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; Morikawa, S.; Masumoto, J. The role of interleukin-1 in general pathology. Inflamm. Regen. 2019, 39, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polańska, B. Neutrophil elastase as an inflammatory mediator. Cent. J. Immunol. 2002, 27, 160–168. [Google Scholar]
- Zhang, D.; Liu, R.; Sun, L.; Huang, C.; Wang, C.; Zhang, D.M.; Zhang, T.T.; Du, G.H. Anti-inflammatory activity of methyl salicylate glycosides isolated from Gaultheria yunnanensis (Franch.) Rehder. Molecules 2011, 16, 3875–3884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutinho, M.A.S.; Muzitano, M.F.; Cruz, E.A.; Bergonzi, M.C.; Kaiser, C.R.; Tinoco, L.W.; Bilia, A.R.; Vincieri, F.F.; Rossi-Bergmann, B.; Costa, S.S. Flowers from Kalanchoe pinnata are a rich source of T cell-suppressive flavonoids. Nat. Prod. Commun. 2012, 7, 175–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.L.; Wu, H.C.; Hwang, T.L.; Lin, C.H.; Cheng, Y.H.; Wang, C.C.; Kan, H.L.; Kuo, Y.H.; Chen, I.S.; Chang, H.S.; et al. Anti-inflammatory and antibacterial activity constituents from the stem of Cinnamomum validinerve. Molecules 2020, 25, 3382. [Google Scholar] [CrossRef]
- Chen, L.; Yang, Y.; Yuan, P.; Yang, Y.; Chen, K.; Jia, Q.; Li, Y. Immunosuppressive effects of A-type procyanidin oligomers from Cinnamomum tamala. Evid. Based Complement. Altern. Med. 2014, 2014, 365258. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Shi, H.; Wang, W.; Ke, Z.; Xu, P.; Zhong, Z.; Li, X.; Wang, S. Antithrombotic effect of grape seed proanthocyanidins extract in a rat model of deep vein thrombosis. J. Vasc. Surg. 2011, 53, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Sun, L.; Liu, R.; Zhang, D.; Lan, X.; Huang, C.; Xin, W.; Wang, C.; Zhang, D.; Du, G. A novel naturally occurring salicylic acid analogue acts as an anti-inflammatory agent by inhibiting nuclear factor-kappaB activity in RAW264.7 macrophages. Mol. Pharm. 2012, 9, 671–677. [Google Scholar] [CrossRef]
- Xin, W.; Huang, C.; Zhang, X.; Zhang, G.; Ma, X.; Sun, L.; Wang, C.; Zhang, D.; Zhang, T.; Du, G. Evaluation of the new anti-inflammatory compound ethyl salicylate 2-O-β-d-glucoside and its possible mechanism of action. Int. Immunopharmacol. 2013, 15, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W.; Jung, S.Y.; Kim, G.D.; Lee, S.Y.; Shin, H.S. Miquelianin inhibits allergic responses in mice by suppressing CD4+ T cell proliferation. Antioxidants 2021, 10, 1120. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Beak, S.Y.; Choi, I.; Sung, J.S. Quercetin and its metabolites protect hepatocytes against ethanol-induced oxidative stress by activation of Nrf2 and AP-1. Food Sci. Biotechnol. 2018, 27, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, Y.; Zheng, Y.; Zhao, J.; Yu, H.; Zhu, J. Relationship between neuroprotective effects and structure of procyanidins. Molecules 2022, 27, 2308. [Google Scholar] [CrossRef]
- Olszewska, M.A.; Michel, P. Activity-guided isolation and identification of free radical-scavenging components from various leaf extracts of Sorbus aria (L.) Crantz. Nat. Prod. Res. 2012, 26, 243–254. [Google Scholar] [CrossRef]
- Matczak, M.; Marchelak, A.; Michel, P.; Owczarek, A.; Piszczan, A.; Kolodziejczyk-Czepas, J.; Nowak, P.; Olszewska, M.A. Sorbus domestica L. leaf extracts as functional products: Phytochemical profiling, cellular safety, pro-inflammatory enzymes inhibition and protective effects against oxidative stress in vitro. J. Funct. Foods 2018, 40, 207–218. [Google Scholar] [CrossRef]
- Marchelak, A.; Owczarek, A.; Rutkowska, M.; Michel, P.; Kolodziejczyk-Czepas, J.; Nowak, P.; Olszewska, M.A. New insights into antioxidant activity of Prunus spinosa flowers: Extracts, model polyphenols and their phenolic metabolites in plasma towards multiple in vivo-relevant oxidants. Phytochem. Lett. 2019, 30, 288–295. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound/Fraction | Methanol–Water (ME) | Ethyl Acetate (EAE) | n-Butanol (BE) |
|---|---|---|---|
| Extraction yield | 371.57 ± 15.57 C | 20.23 ± 1.01 A | 295.56 ± 11.78 B |
| Phenolic fractions: | |||
| TPC | 302.35 ± 1.01 C | 168.12 ± 0.69 A | 174.93 ± 1.64 B |
| TPH | 223.54 ± 3.02 B | 336.72 ± 4.04 C | 204.63 ± 3.66 A |
| TSAL | 98.89 ± 0.47 A | 288.13 ± 3.91 C | 128.61 ± 3.06 B |
| TPA | 174.38 ± 2.35 C | 36.93 ± 0.35 A | 87.32 ± 1.37 B |
| TLPA | 63.99 ± 2.01 C | 9.67 ± 0.16 A | 43.64 ± 1.14 B |
| TFL | 49.35 ± 0.69 C | 18.59 ± 0.27 A | 24.28 ± 0.19 B |
| TPHA | 11.31 ± 0.13 B | 20.33 ± 0.12 C | 8.10 ± 0.12 A |
| TCHA | 6.80 ± 0.09 C | 5.88 ± 0.08 B | 5.47 ± 0.08 A |
| SPHA | 4.51 ± 0.07 B | 14.45 ± 0.16 C | 2.63 ± 0.06 A |
| Primary compounds: | |||
| Neochlorogenic acid (5) | 4.24 ± 0.08 B | 2.89 ± 0.03 A | 2.91 ± 0.09 A |
| Chlorogenic acid (10) | 1.68 ± 0.04 B | 2.99 ± 0.04 C | 1.31 ± 0.05 A |
| Cryptochlorogenic acid (12) | 0.88 ± 0.04 A | n.d. | 1.25 ± 0.02 B |
| Methyl salicylate triglycoside (11) | 0.49 ± 0.01 A | n.d. | 0.79 ± 0.04 B |
| Gaultherin (20) | 98.41 ± 0.31 A | 288.13 ± 3.91 C | 127.81 ± 2.81 B |
| (-)-Epicatechin (18) | 9.07 ± 0.29 C | 7.91 ± 0.13 B | 0.54 ± 0.02 A |
| Procyanidin B2 (14) | 13.15 ± 0.37 B | n.d. | 6.29 ± 0.08 A |
| Cinnamtanin B-1 (24) | 18.61 ± 0.28 C | 0.77 ± 0.03 A | 12.89 ± 0.58 B |
| Wintergreenoside A (31) | 7.26 ± 0.24 B | n.d. | 1.24 ± 0.04 A |
| Hyperoside (35) | 5.01 ± 0.16 A | 9.46 ± 0.37 C | 5.33 ± 0.26 B |
| Isoquercitrin (36) | 1.16 ± 0.05 B | 2.97 ± 0.09 C | 1.10 ± 0.04 A |
| Miquelianin (37) | 32.14 ± 0.69 C | 1.55 ± 0.06 A | 6.72 ± 0.13 B |
| Guaijaverin (39) | 1.46 ± 0.02 A | 4.17 ± 0.19 C | 1.73 ± 0.07 B |
| Quercetin (41) | 0.22 ± 0.01 A | 0.43 ± 0.02 B | 7.07 ± 0.17 C |
| Analyte | DPPH | FRAP | TBARS | O2•− | •OH | H2O2 |
|---|---|---|---|---|---|---|
| SC50 (µg/mL) a | mmol Fe2+/g b | IC50 (µg/mL) c | SC50 (µg/mL) a | SC50 (µg/mL) a | SC50 (µg/mL) a | |
| ME | 6.77 ± 0.28 C | 6.36 ± 0.14 C | 8.46 ± 0.19 C | 26.33 ± 0.88 B | 152.04 ± 4.28 B | 44.41 ± 1.96 C |
| EAE | 14.17 ± 0.29 E | 3.82 ± 0.13 A | 14.71 ± 2.06 E | 39.30 ± 1.55 C | 480.77 ± 13.01 E | 83.32 ± 2.98 D |
| BE | 8.33 ± 0.09 D | 4.41 ± 0.18 B | 10.68 ± 0.46 D | 62.36 ± 2.31 D | 236.51 ± 8.64 D | 43.25 ± 1.61 C |
| QU | 1.52 ± 0.03 A | 49.04 ± 0.59 F | 1.69 ± 0.05 A | 7.35 ± 0.19 A | 41.07 ± 3.89 A | 6.96 ± 0.42 A |
| TX | 4.23 ± 0.05 B | 12.56 ± 0.23 D | 4.58 ± 0.33 B | 142.15 ± 2.19 E | 172.26 ± 3.01 C | 15.76 ± 0.31 B |
| Analyte | HYAL | LOX | COX-2 |
|---|---|---|---|
| IC50 (µg/mL) a | IC50 (µg/mL) a | IC50 (μg/mL) a | |
| ME | 18.66 ± 0.86 C | 351.55 ± 13.76 D | 711.08 ± 25.55 D |
| EAE | 34.57 ± 1.82 D | 626.25 ± 11.19 E | 1416.93 ± 50.85 F |
| BE | 14.63 ± 0.66 B | 267.04 ± 12.98 C | 970.64 ± 38.53 E |
| IND | 12.68 ± 1.79 A | 91.89 ± 3.95 A | 184.32 ± 8.56 A |
| DEX | 14.07 ± 1.25 B | 120.16 ± 4.86 B | 511.23 ± 14.58 C |
| QU | 31.78 ± 1.52 D | 88.35 ± 7.37 A | 469.46 ± 14.52 B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michel, P.; Granica, S.; Rosińska, K.; Glige, M.; Rojek, J.; Poraj, Ł.; Olszewska, M.A. The Effect of Standardised Leaf Extracts of Gaultheria procumbens on Multiple Oxidants, Inflammation-Related Enzymes, and Pro-Oxidant and Pro-Inflammatory Functions of Human Neutrophils. Molecules 2022, 27, 3357. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103357
Michel P, Granica S, Rosińska K, Glige M, Rojek J, Poraj Ł, Olszewska MA. The Effect of Standardised Leaf Extracts of Gaultheria procumbens on Multiple Oxidants, Inflammation-Related Enzymes, and Pro-Oxidant and Pro-Inflammatory Functions of Human Neutrophils. Molecules. 2022; 27(10):3357. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103357
Chicago/Turabian StyleMichel, Piotr, Sebastian Granica, Karolina Rosińska, Małgorzata Glige, Jarosław Rojek, Łukasz Poraj, and Monika Anna Olszewska. 2022. "The Effect of Standardised Leaf Extracts of Gaultheria procumbens on Multiple Oxidants, Inflammation-Related Enzymes, and Pro-Oxidant and Pro-Inflammatory Functions of Human Neutrophils" Molecules 27, no. 10: 3357. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103357