
Antimicrobial Activity of Dihydroisocoumarin Isolated from Wadi Lajab Sediment-Derived Fungus Penicillium chrysogenum: In Vitro and In Silico Study


 ,
,
Abstract
:1. Introduction
2. Results and Discussion
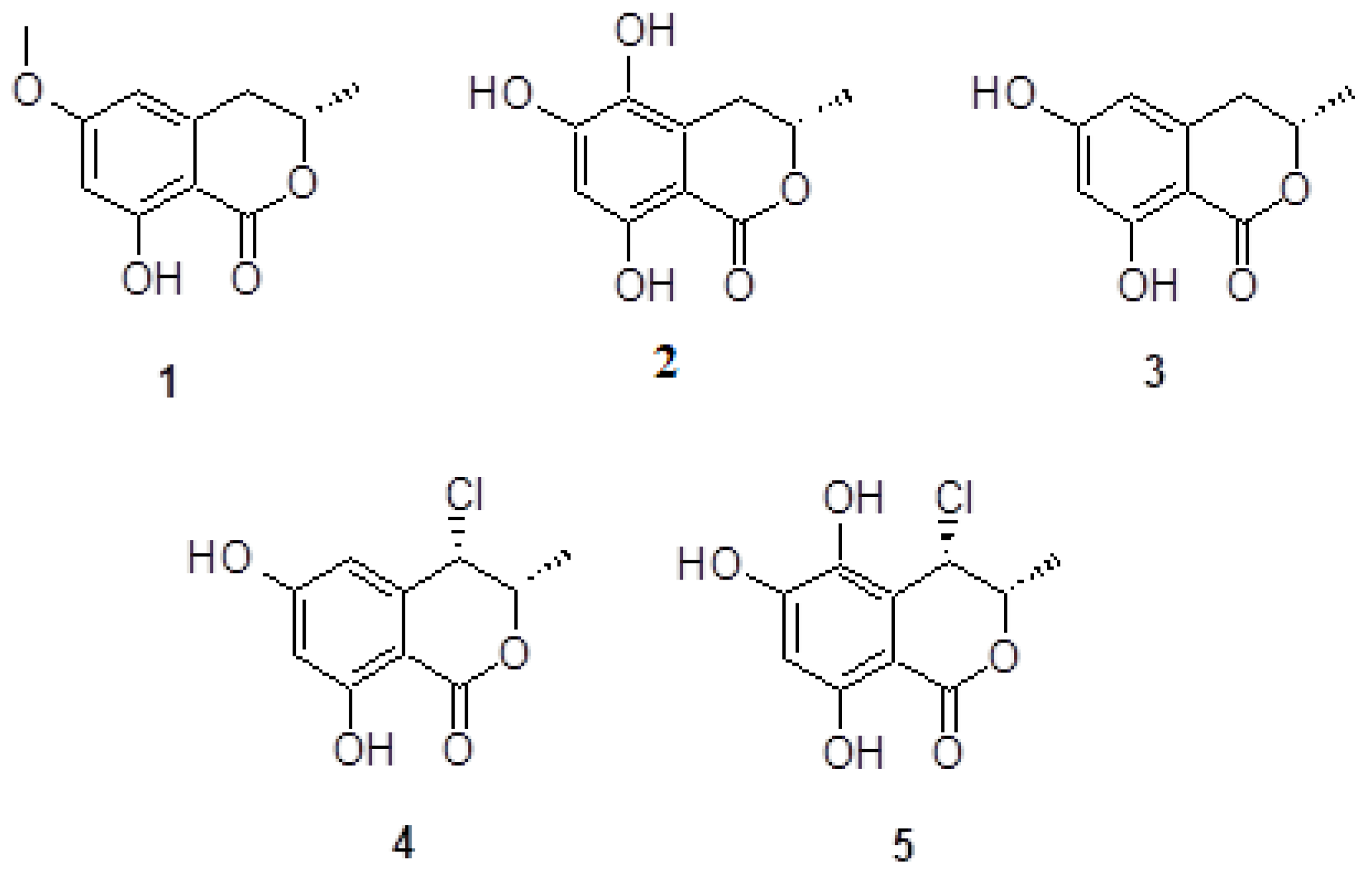
2.1. Chemistry
2.2. Biology
Bioactivity of Tested Compounds against the Pathogenic Bacteria
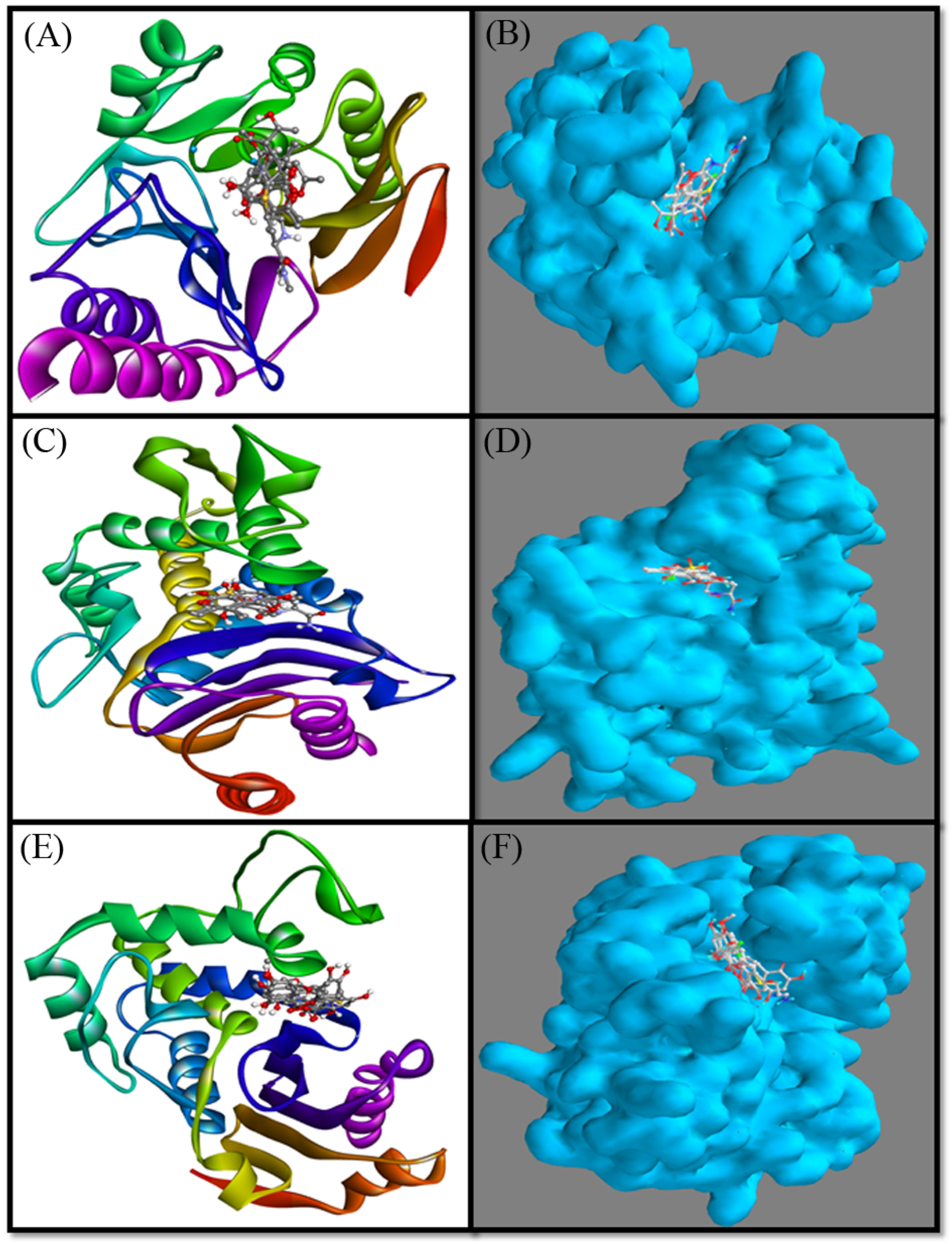
2.3. Molecular Modelling
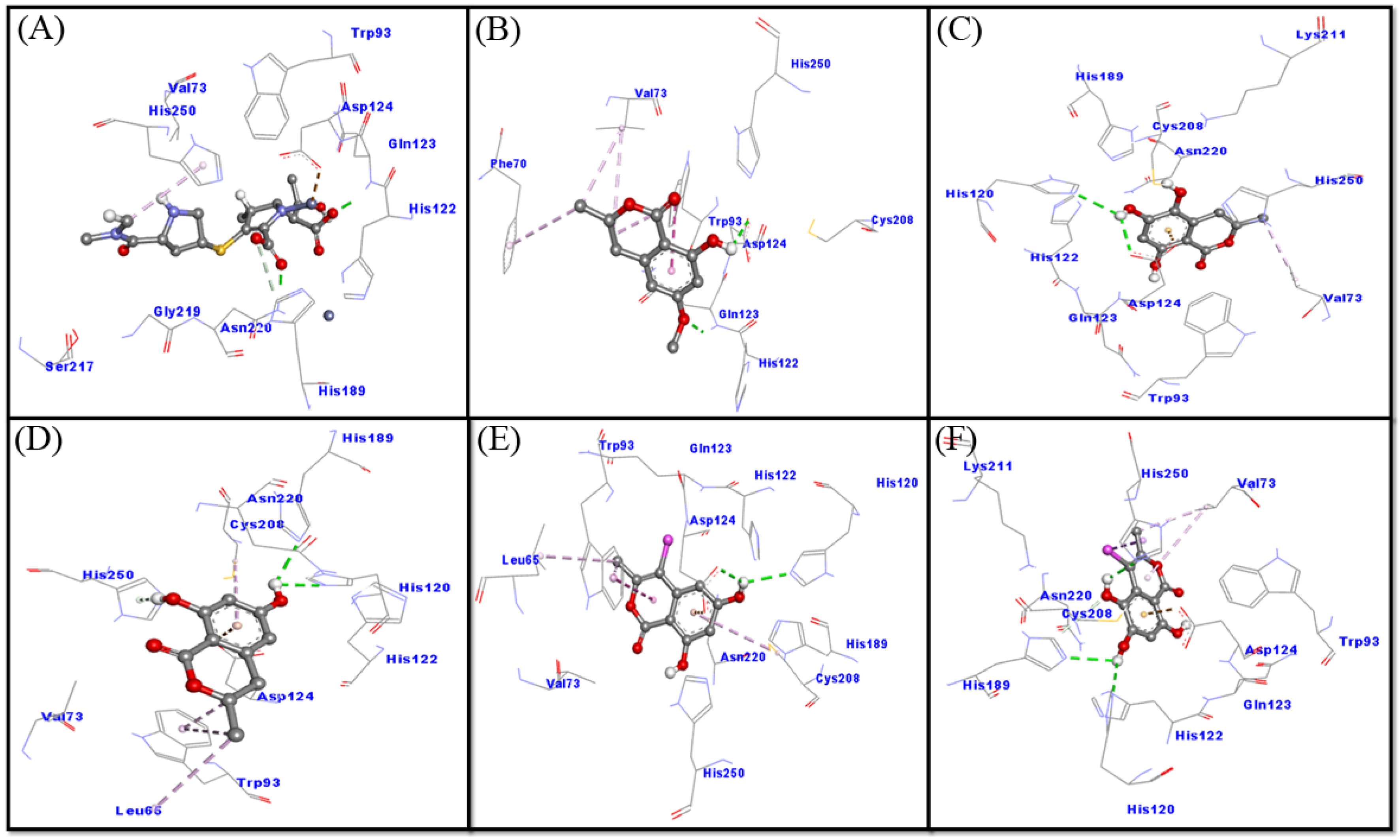
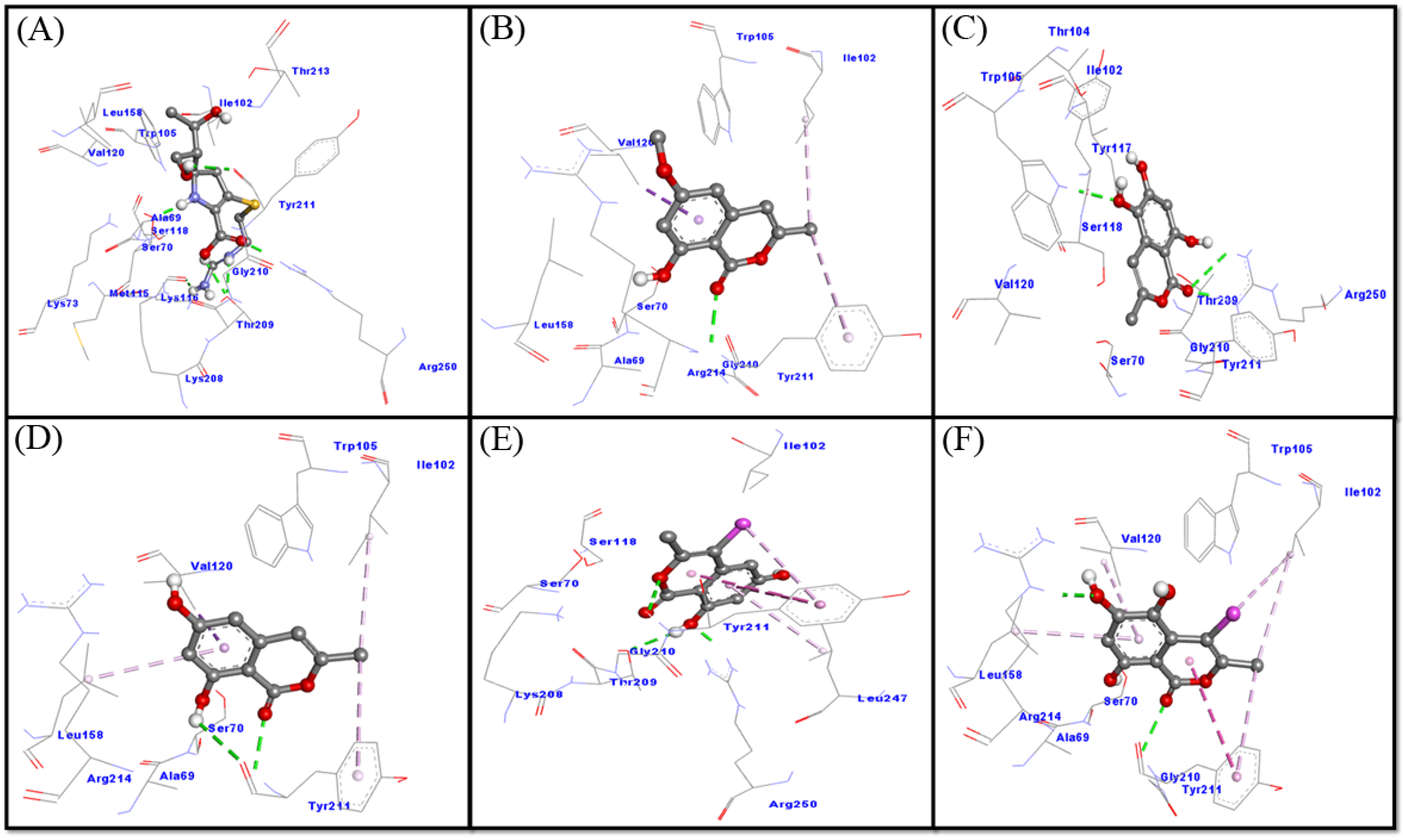
2.3.1. Interaction between NDM-1 and Selected Ligands
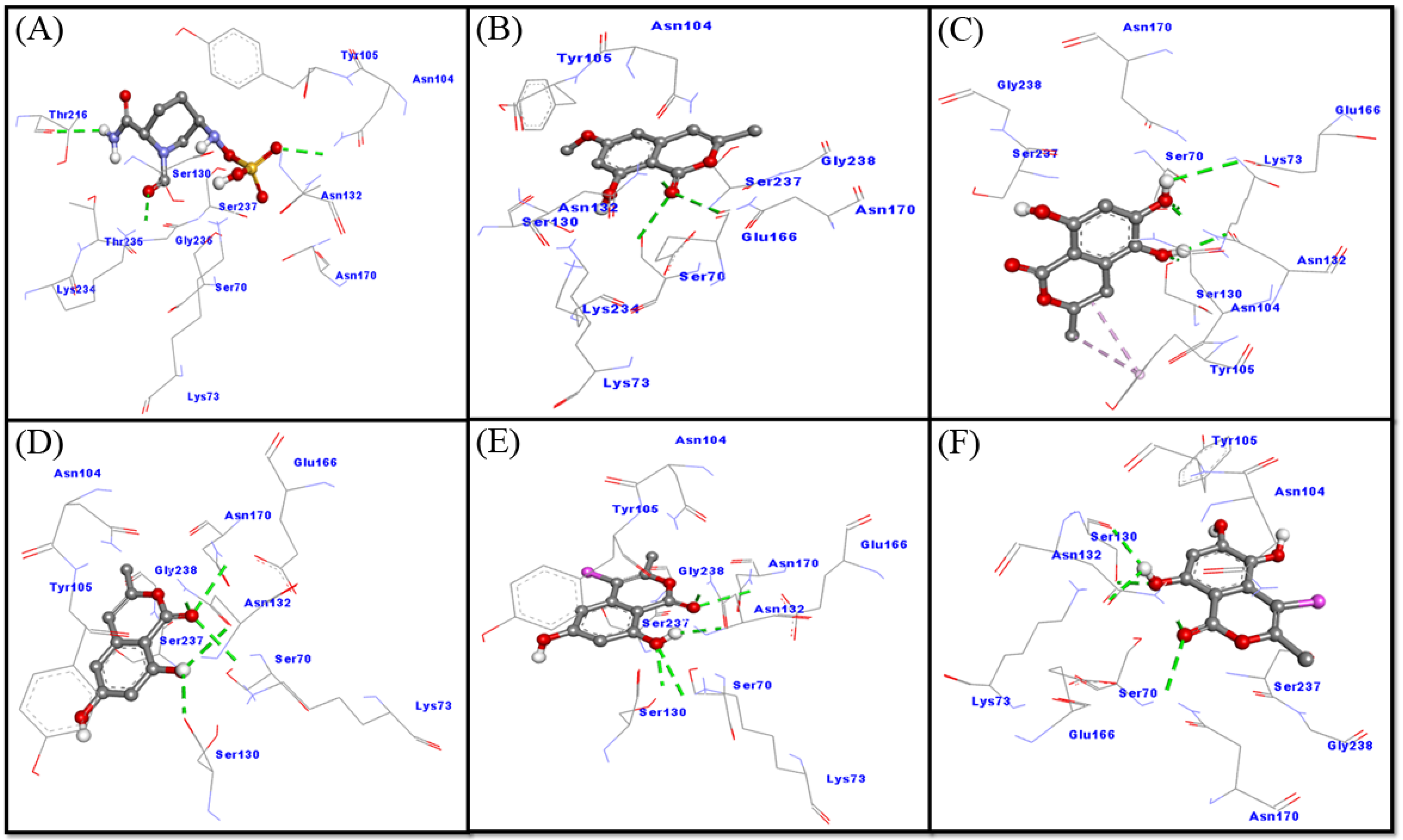
2.3.2. Interaction between CTX-M-15 and Selected Ligands
2.3.3. Interaction between OXA-48 and Selected Ligands
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Fungal Strain Materials
3.3. Fermentation, Extraction and Isolation
3.4. Antibacterial Assay
3.4.1. Agar Diffusion Method
3.4.2. Minimum Inhibitory Concentration
3.5. Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Tang, S.S.; Apisarnthanarak, A.; Hsu, L.Y. Mechanisms of β-Lactam Antimicrobial Resistance and Epidemiology of Major Community- and Healthcare-Associated Multidrug-Resistant Bacteria. Adv. Drug Deliv. Rev. 2014, 78, 3–13. [Google Scholar] [CrossRef]
- King, D.T.; King, A.M.; Lal, S.M.; Wright, G.D.; Strynadka, N.C.J. Molecular Mechanism of Avibactam-Mediated β-Lactamase Inhibition. ACS Infect. Dis. 2015, 1, 175–184. [Google Scholar] [CrossRef]
- Hawkey, P.M. Multidrug-Resistant Gram-Negative Bacteria: A Product of Globalization. J. Hosp. Infect. 2015, 89, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Gelband, H.; Miller-Petrie, M.; Pant, S.; Gandra, S.; Levinson, J.; Barter, D.; White, A.; Laxminarayan, R. State of the World’s Antibiotics; Center for Disease Dynamics Economics & Policy (CDDEP): Geneva, Switzerland, 2015. [Google Scholar]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Pfeifer, Y.; Cullik, A.; Witte, W. Resistance to Cephalosporins and Carbapenems in Gram-Negative Bacterial Pathogens. Int. J. Med. Microbiol. 2010, 300, 371–379. [Google Scholar] [CrossRef]
- Johnson, A.P.; Woodford, N. Global Spread of Antibiotic Resistance: The Example of New Delhi Metallo-β-Lactamase (NDM)-Mediated Carbapenem Resistance. J. Med. Microbiol. 2013, 62, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a New Antibiotic Resistance Mechanism in India, Pakistan, and the UK: A Molecular, Biological, and Epidemiological Study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Y.; Walsh, T.R.; Liu, D.; Shen, Z.; Zhang, R.; Yin, W.; Yao, H.; Li, J.; Shen, J. Plasmid-Mediated Novel BlaNDM-17 Gene Encoding a Carbapenemase with Enhanced Activity in a Sequence Type 48 Escherichia coli Strain. Antimicrob. Agents Chemother. 2017, 61, e02233-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalde, S.S.; Choi, H.K. Recent Advances in the Development of β-Lactamase Inhibitors. J. Microbiol. 2020, 58, 633–647. [Google Scholar] [CrossRef]
- Poirel, L.; Carbonnelle, E.; Bernabeu, S.; Gutmann, L.; Rotimi, V.; Nordmann, P. Importation of OXA-48-Producing Klebsiella Pneumoniae from Kuwait. J. Antimicrob. Chemother. 2012, 67, 2051–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, B.A.; Amyes, S.G.B. OXA β-Lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [Green Version]
- Khaldi, N.; Seifuddin, F.T.; Turner, G.; Haft, D.; Nierman, W.C.; Wolfe, K.H.; Fedorova, N.D. SMURF: Genomic Mapping of Fungal Secondary Metabolite Clusters. Fungal Genet. Biol. 2010, 47, 736–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, K.; Medema, M.H.; Kottmann, R.; Lee, S.Y.; Weber, T. The AntiSMASH Database, a Comprehensive Database of Microbial Secondary Metabolite Biosynthetic Gene Clusters. Nucleic Acids Res. 2017, 45, D555–D559. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, J.C.; Grijseels, S.; Prigent, S.; Ji, B.; Dainat, J.; Nielsen, K.F.; Frisvad, J.C.; Workman, M.; Nielsen, J. Global Analysis of Biosynthetic Gene Clusters Reveals Vast Potential of Secondary Metabolite Production in Penicillium Species. Nat. Microbiol. 2017, 2, 17044. [Google Scholar] [CrossRef] [PubMed]
- Gombert, A.K.; Veiga, T.; Puig-Martinez, M.; Lamboo, F.; Nijland, J.G.; Driessen, A.J.M.; Pronk, J.T.; Daran, J.M. Functional Characterization of the Oxaloacetase Encoding Gene and Elimination of Oxalate Formation in the β-Lactam Producer Penicillium chrysogenum. Fungal Genet. Biol. 2011, 48, 831–839. [Google Scholar] [CrossRef]
- Houbraken, J.; Frisvad, J.C.; Samson, R.A. Fleming’s Penicillin Producing Strain Is Not Penicillium chrysogenum but P. rubens. IMA Fungus 2011, 2, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Müller, R.; Wohlleben, W.; et al. AntiSMASH 3.0—A Comprehensive Resource for the Genome Mining of Biosynthetic Gene Clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, A.; Qasim, M. Total Synthesis of Cytotoxic Metabolite (±)-Desmethyldiaportinol from Ampelomyces sp. Nat. Prod. Res. 2014, 28, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Chatare, V.; Pal, M. Isocoumarin and Its Derivatives: An Overview on Their Synthesis and Applications. COC Curr. Org. Chem. 2011, 15, 782–800. [Google Scholar] [CrossRef]
- Wu, H.; Wang, Y.-C.; Shatskiy, A.; Li, Q.-Y.; Liu, J.-Q.; Kärkäs, M.D.; Wang, X.-S. Modular Synthesis of 3-Substituted Isocoumarins via Silver-Catalyzed Aerobic Oxidation/ 6-Endo Heterocyclization of Ortho -Alkynylbenzaldehydes. Org. Biomol. Chem. 2021, 19, 6657–6664. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Regasini, L.O.; Silva, G.H.; Pfenning, L.H.; Young, M.C.M.; Berlinck, R.G.S.; Bolzani, V.S.; Araujo, A.R. Dihydroisocoumarins Produced by Xylaria sp. and Penicillium sp., Endophytic Fungi Associated with Piper Aduncum and Alibertia Macrophylla. Phytochem. Lett. 2011, 4, 93–96. [Google Scholar] [CrossRef]
- Kuramata, M.; Fujioka, S.; Shimada, A.; Kawano, T.; Kimura, Y. Citrinolactones A, B and C, and Sclerotinin C, Plant Growth Regulators from Penicillium Citrinum. Biosci. Biotechnol. Biochem. 2007, 71, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.M.; Silva, G.H.; Regasini, L.O.; Zanardi, L.M.; Evangelista, A.H.; Young, M.C.M.; Bolzani, V.S.; Araujo, A.R. Bioactive Metabolites Produced by Penicillium sp.1 and sp.2, Two Endophytes Associated with Alibertia Macrophylla (Rubiaceae). Z. Nat. C 2009, 64, 824–830. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.-X.; Huang, C.Y.; Yan, Z.Y.; Chen, T.; Hong, K.; Long, Y.-H. New Furo[3,2-h]Isochroman from the Mangrove Endophytic Fungus Aspergillus Sp. 085242. Chin. J. Nat. Med. 2020, 18, 855–859. [Google Scholar] [CrossRef]
- Kruse, J.; Kummer, V.; Shivas, R.G.; Thines, M. The First Smut Fungus, Thecaphora Anthemidis Sp. Nov. (Glomosporiaceae), Described from Anthemis (Asteraceae). MycoKeys 2018, 41, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orfali, R.; Perveen, S.; Al-Taweel, A.; Ahmed, A.F.; Majrashi, N.; Alluhay, K.; Khan, A.; Luciano, P.; Taglialatela-Scafati, O. Penipyranicins A–C: Antibacterial Methylpyran Polyketides from a Hydrothermal Spring Sediment Penicillium sp. J. Nat. Prod. 2020, 83, 3591–3597. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.H.; Shehabeldine, A.M.; Abdelaziz, A.M.; Amin, B.H.; Sharaf, M.H. Antifungal Activity of Endophytic Aspergillus Terreus Extract Against Some Fungi Causing Mucormycosis: Ultrastructural Study. Appl Biochem Biotechnol 2022, 2, 1–15. [Google Scholar] [CrossRef]
- Valgas, C.; de Souza, S.M.; Smânia, E.F.A.; Smânia Jr., A. Screening Methods to Determine Antibacterial Activity 687 of Natural Products. Braz. J. Microbiol. 2007, 38, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing Natural Products as Potential Anti-Biofilm Agents. Chin Med. 2019, 14, 11. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Li, H.; Qi, J.; Chen, C.; Lu, Y.; Wang, Y. Genome Mining of Secondary Metabolites from a Marine-Derived Aspergillus Terreus B12. Arch Microbiol. 2021, 203, 5621–5633. [Google Scholar] [CrossRef] [PubMed]
- Krohn, K.; Flörke, U.; Rao, M.S.; Steingröver, K.; Aust, H.-J.; Draeger, S.; Schulz, B. Metabolites from Fungi 15. New Isocoumarins from an Endophytic Fungus Isolated from the Canadian Thistle Cirsium arvense. Nat. Prod. Lett. 2001, 15, 353–361. [Google Scholar] [CrossRef]
- Abdel-Hameed, M.; Bertrand, R.L.; Piercey-Normore, M.D.; Sorensen, J.L. Identification of 6-Hydroxymellein Synthase and Accessory Genes in the Lichen Cladonia uncialis. J. Nat. Prod. 2016, 79, 1645–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Elimat, T.; Figueroa, M.; Raja, H.A.; Alnabulsi, S.; Oberlies, N.H. Coumarins, Dihydroisocoumarins, a Dibenzo-α-Pyrone, a Meroterpenoid, and a Merodrimane from Talaromyces Amestolkiae. Tetrahedron Lett. 2021, 72, 153067. [Google Scholar] [CrossRef] [PubMed]
- Krohn, K.; Bahramsari, R.; Flörke, U.; Ludewig, K.; Kliche-Spory, C.; Michel, A.; Aust, H.-J.; Draeger, S.; Schulz, B.; Antus, S. Dihydroisocoumarins from Fungi: Isolation, Structure Elucidation, Circular Dichroism and Biological Activity. Phytochemistry 1997, 45, 313–320. [Google Scholar] [CrossRef]
- Hou, A.; Dickschat, J.S. The Biosynthetic Gene Cluster for Sestermobaraenes—Discovery of a Geranylfarnesyl Diphosphate Synthase and a Multiproduct Sesterterpene Synthase from Streptomyces mobaraensis. Angew. Chem. Int. 2020, 59, 19961–19965. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Z.; Liu, H.; Pan, Y.; Li, J.; Liu, L.; She, Z. Dichloroisocoumarins with Potential Anti-Inflammatory Activity from the Mangrove Endophytic Fungus Ascomycota sp. CYSK-4. Mar. Drugs 2018, 16, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orfali, R.; Perveen, S. Secondary Metabolites from the Aspergillus sp. in the Rhizosphere Soil of Phoenix dactylifera (Palm Tree). BMC Chem. 2019, 13, 103. [Google Scholar] [CrossRef] [PubMed]
- Orfali, R.; Perveen, S.; Khan, M.F.; Ahmed, A.F.; Wadaan, M.A.; Al-Taweel, A.M.; Alqahtani, A.S.; Nasr, F.A.; Tabassum, S.; Luciano, P.; et al. Antiproliferative Illudalane Sesquiterpenes from the Marine Sediment Ascomycete Aspergillus oryzae. Mar. Drugs 2021, 19, 333. [Google Scholar] [CrossRef]
- Orfali, R.; Perveen, S.; Khan, M.F.; Ahmed, A.F.; Tabassum, S.; Luciano, P.; Chianese, G.; Taglialatela-Scafati, O. Asporychalasin, a Bioactive Cytochalasan with an Unprecedented 6/6/11 Skeleton from the Red Sea Sediment Aspergillus oryzae. Phytochemistry 2021, 192, 112952. [Google Scholar] [CrossRef]
- Masalha, M.; Rayan, M.; Adawi, A.; Abdallah, Z.; Rayan, A. Capturing Antibacterial Natural Products with In�silico Techniques. Mol Med Report. 2018, 18, 763–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Yousef, H.M.; Ahmed, A.F.; Al-Shabib, N.A.; Laeeq, S.; Khan, R.A.; Rehman, M.T.; Alsalme, A.; Al-Ajmi, M.F.; Khan, M.S.; Husain, F.M. Onion Peel Ethylacetate Fraction and Its Derived Constituent Quercetin 4′-O-β-D Glucopyranoside Attenuates Quorum Sensing Regulated Virulence and Biofilm Formation. Front. Microbiol. 2017, 8, 1675. [Google Scholar] [CrossRef] [Green Version]
- Ebrahim, W.; El-Neketi, M.; Lewald, L.-I.; Orfali, R.S.; Lin, W.; Rehberg, N.; Kalscheuer, R.; Daletos, G.; Proksch, P. Metabolites from the Fungal Endophyte Aspergillus Austroafricanus in Axenic Culture and in Fungal–Bacterial Mixed Cultures. J. Nat. Prod. 2016, 79, 914–922. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, M.T.; Shamsi, H.; Khan, A.U. Insight into the binding mechanism of imipenem to human serum albumin by spectroscopic and computational approaches. Mol. Pharmaceut. 2014, 11, 1785–1797. [Google Scholar] [CrossRef]
- Rabbani, N.; Tabrez, S.; ul Islam, B.; Rehman, M.T.; Alsenaidy, A.M.; AlAjmi, M.F.; Khan, R.A.; Alsenaidy, M.A.; Khan, M.S. Characterization of Colchicine Binding with Normal and Glycated Albumin: In Vitro and Molecular Docking Analysis. J. Biomol. Struct. Dyn. 2018, 36, 3453–3462. [Google Scholar] [CrossRef] [PubMed]
- King, D.T.; Worrall, L.J.; Gruninger, R.; Strynadka, N.C.J. New Delhi Metallo-β-Lactamase: Structural Insights into β-Lactam Recognition and Inhibition. J. Am. Chem. Soc. 2012, 134, 11362–11365. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | S. aureus | B. licheniformis | E. xiangfangensis | E. fergusonii | P. aeruginosa |
|---|---|---|---|---|---|
| 1 | 10.4 | 12.2 | 21.6 | 18.4 | 14.8 |
| 2 | 14.5 | 16.3 | >25 | >25 | >25 |
| 3 | >25 | >25 | >25 | >25 | >25 |
| 4 | 1.00 | 0.8 | 3.5 | 4.7 | 5.3 |
| 5 | 3.8 | 4.3 | 5.2 | 7.5 | 9.7 |
| Amikacin | 0.5 | 0.2 | 0.3 | 0.4 | 0.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orfali, R.; Perveen, S.; AlAjmI, M.F.; Ghaffar, S.; Rehman, M.T.; AlanzI, A.R.; Gamea, S.B.; Essa Khwayri, M. Antimicrobial Activity of Dihydroisocoumarin Isolated from Wadi Lajab Sediment-Derived Fungus Penicillium chrysogenum: In Vitro and In Silico Study. Molecules 2022, 27, 3630. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27113630
Orfali R, Perveen S, AlAjmI MF, Ghaffar S, Rehman MT, AlanzI AR, Gamea SB, Essa Khwayri M. Antimicrobial Activity of Dihydroisocoumarin Isolated from Wadi Lajab Sediment-Derived Fungus Penicillium chrysogenum: In Vitro and In Silico Study. Molecules. 2022; 27(11):3630. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27113630
Chicago/Turabian StyleOrfali, Raha, Shagufta Perveen, Mohamed Fahad AlAjmI, Safina Ghaffar, Md Tabish Rehman, Abdullah R. AlanzI, Saja Bane Gamea, and Mona Essa Khwayri. 2022. "Antimicrobial Activity of Dihydroisocoumarin Isolated from Wadi Lajab Sediment-Derived Fungus Penicillium chrysogenum: In Vitro and In Silico Study" Molecules 27, no. 11: 3630. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27113630