
Upgrading the Nutritional Value of PKC Using a Bacillus subtilis Derived Monocomponent β-Mannanase


Abstract
:1. Introduction
2. Results
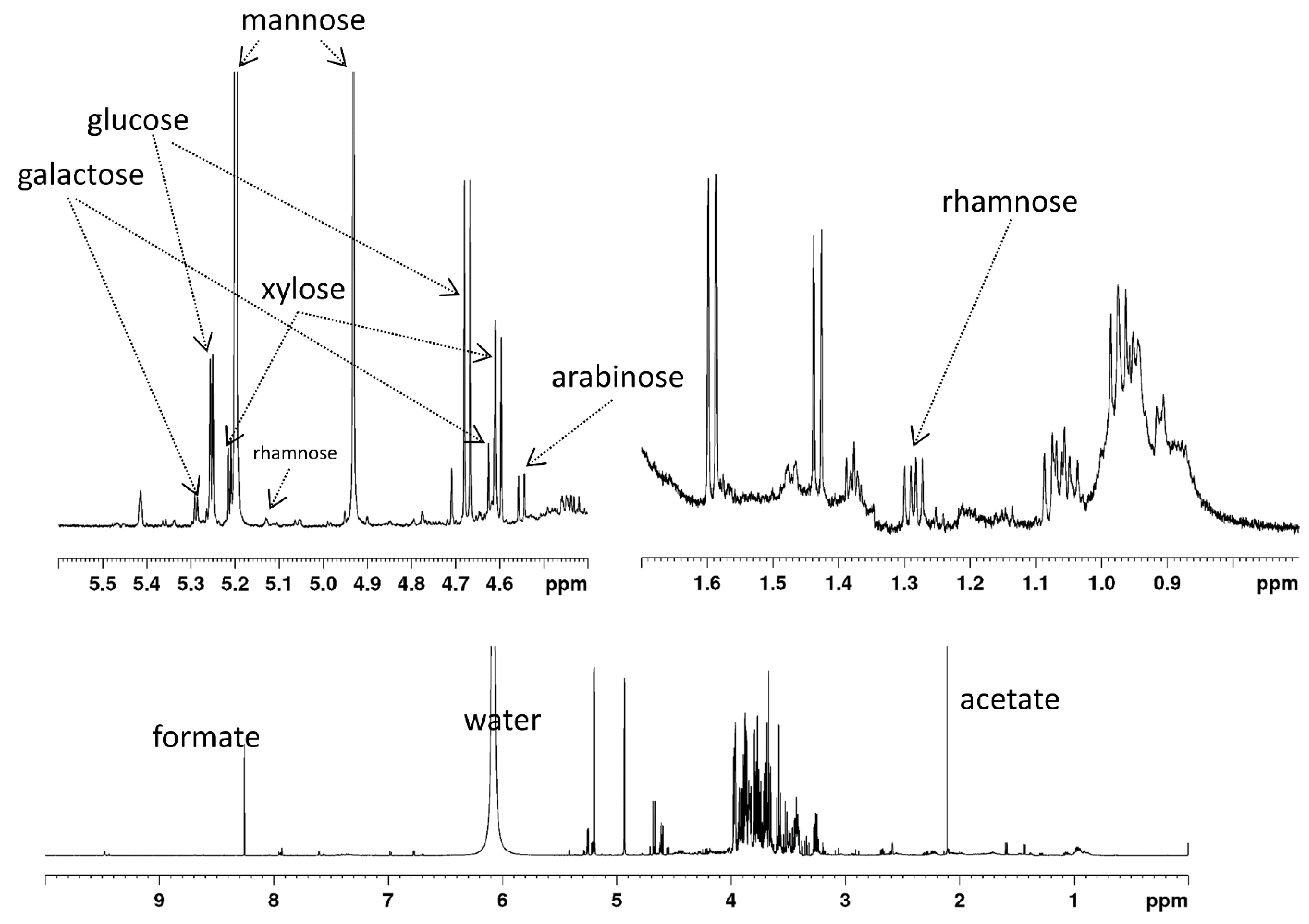
2.1. Analysis of PKC Used
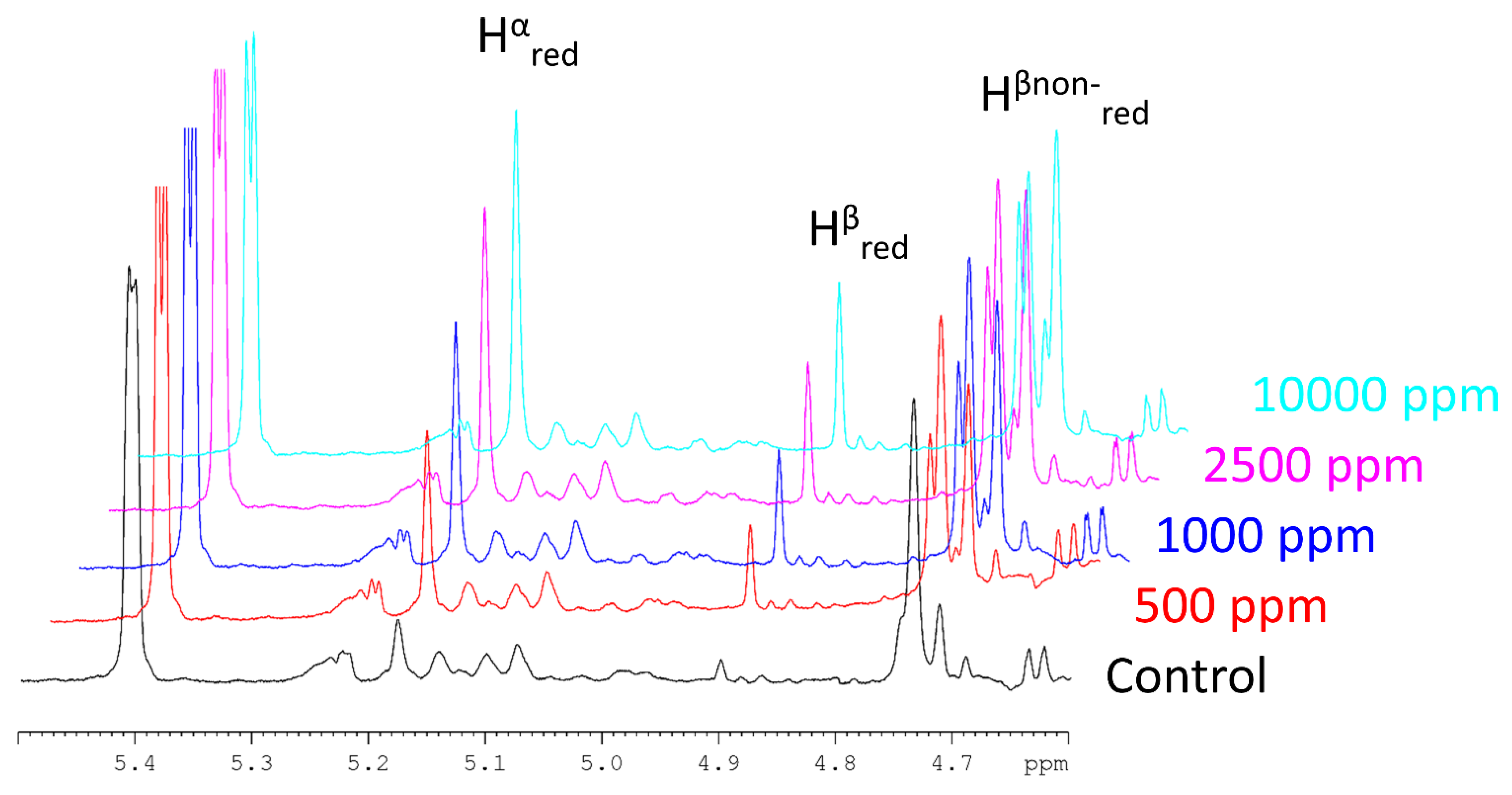
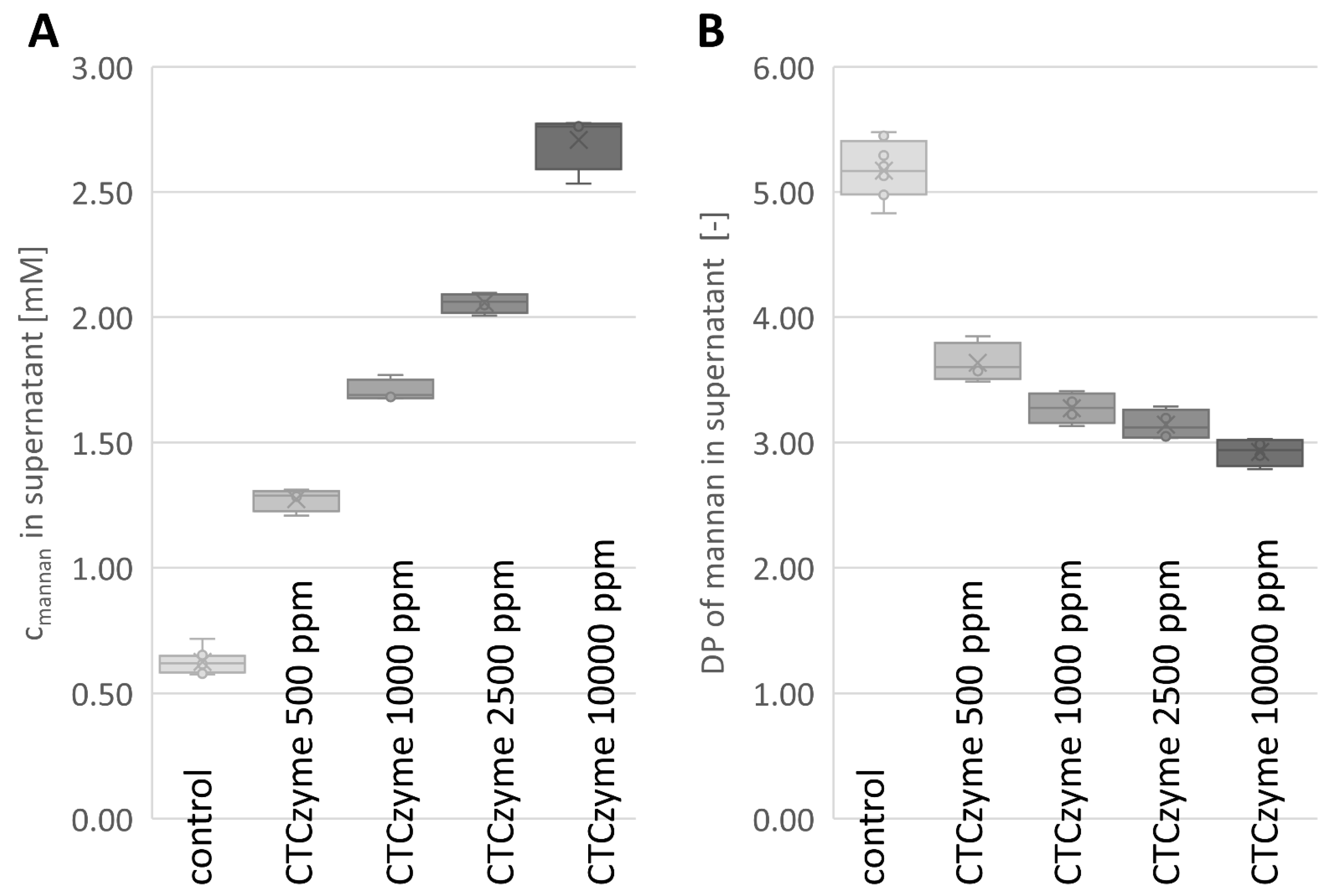
2.2. Enzymatic Digestion
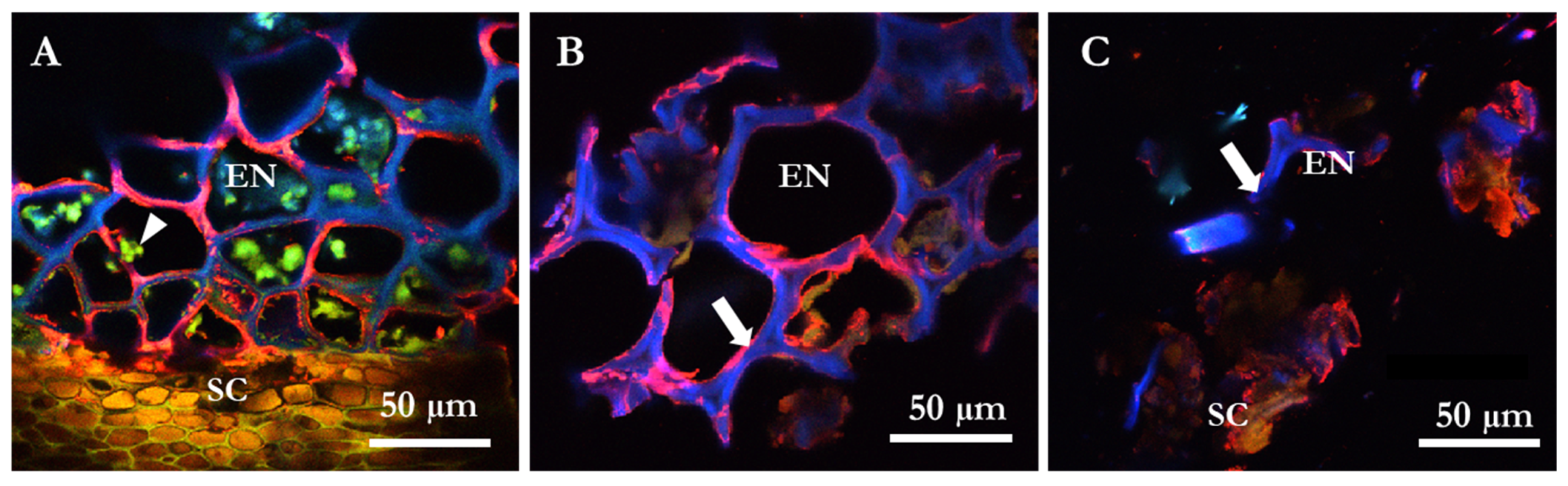
2.3. Microscopy
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.2.1. Determination of Moisture
4.2.2. Determination of Ether Extracts
4.2.3. Determination of Crude Protein
4.2.4. Determination of Crude Fiber
4.3. Enzyme Product
4.4. Enzyme Treatment of PKC
4.5. Colorimetric Assay of Reducing Sugar Content (DNS-Assay)
4.6. Analysis of Mannose Oligosaccharides
4.7. Immunolocalization and Histology with Confocal Microscopy
4.8. Total Hydrolysis of PKC, Calculation of Degree of Enzymatic Conversion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Food Outlook—Biannual Report on Global Food Markets; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Heuzé, V.; Tran, G.; Sauvant, D.; Noblet, J.; Renaudeau, D.; Bastianelli, D.; Lebas, F. Palm kernel meal. In Feedipedia, a Programme by INRAE, CIRAD, AFZ and FAO; INRA: Paris, France; CIRAD: Montpellier, France; AFZ: Paris, France; FAO: Rome, Italy, 2016; Available online: https://www.feedipedia.org/node/43 (accessed on 25 August 2021).
- Azizi, M.N.; Loh, T.C.; Foo, H.L.; Chung, E.L.T. Is palm kernel cake a suitable alternative feed ingredient for poultry? Animals 2021, 11, 338. [Google Scholar] [CrossRef]
- Sharmila, A.; Alimon, A.; Azhar, K.; Noor, H. Improving nutritional values of palm kernel cake (PKC) as poultry feeds: A review. Malays. J. Anim. Sci. 2014, 17, 1–18. [Google Scholar]
- Choct, M.; Annison, G. Anti-nutritive activity of wheat pentosans in broiler diets. Br. Poult. Sci. 1990, 31, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Choct, M.; Hughes, R.J.; Bedford, M.R. Effects of a xylanase on individual bird variation, starch digestion throughout the intestine, and ileal and caecal volatile fatty acid production in chickens fed wheat. Br. Poult. Sci. 1999, 40, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Józefiak, D.; Rutkowski, A.; Martin, S. Carbohydrate fermentation in the avian ceca: A review. Anim. Feed. Sci. Technol. 2004, 113, 1–15. [Google Scholar] [CrossRef]
- Navidshad, B.; Liang, J.B.; Jahromi, M.F.; Akhlaghi, A.; Abdullah, N. Effects of enzymatic treatment and shell content of palm kernel expeller meal on performance, nutrient digestibility, and ileal bacterial population in broiler chickens. J. Appl. Poult. Res. 2016, 25, 474–482. [Google Scholar] [CrossRef]
- Düsterhöft, E.M.; Voragen, A.G.J.; Engels, F.M. Non-starch polysaccharides from sunflower (Helianthus annuus) meal and palm kernel (Elaeis guineensis) meal-preparation of cell wall material and extraction of polysaccharide fractions. J. Sci. Food Agric. 1991, 55, 411–422. [Google Scholar] [CrossRef]
- Düsterhöft, E.M.; Posthumus, M.A.; Voragen, A.G.J. Non-starch polysaccharides from sunflower (Helianthus annuus) meal and palm-kernel (Elaeis guineensis) meal—Investigation of the structure of major polysaccharides. J. Sci. Food Agric. 1992, 59, 151–160. [Google Scholar] [CrossRef]
- Van der Wielen, P.W.J.J.; Biesterveld, S.; Notermans, S.; Hofstra, H.; Urlings, B.A.P.; van Knapen, F. Role of Volatile Fatty Acids in Development of the Cecal Microflora in Broiler Chickens during Growth. Appl. Environ. Microbiol. 2000, 66, 2536–2540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shashidhara, R.G.; Devegowda, G. Effect of dietary mannan oligosaccharide on broiler breeder production traits and immunity. Poult. Sci. 2003, 82, 1319–1325. [Google Scholar] [CrossRef]
- Rezaei, S.; Faseleh Jahromi, M.; Liang, J.B.; Zulkifli, I.; Farjam, A.S.; Laudadio, V.; Tufarelli, V. Effect of oligosaccharides extract from palm kernel expeller on growth performance, gut microbiota and immune response in broiler chickens. Poult. Sci. 2015, 94, 2414–2420. [Google Scholar] [CrossRef] [PubMed]
- Hesselman, K.; Åman, P. The effect of β-glucanase on the utilization of starch and nitrogen by broiler chickens fed on barley of low- or high-viscosity. Anim. Feed. Sci. Technol. 1986, 15, 83–93. [Google Scholar] [CrossRef]
- Pettersson, D.; Aman, P. Enzyme supplementation of a poultry diet containing rye and wheat. Br. J. Nutr. 1989, 62, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizvi, S. Feed Enzymes Market by Type (Phytase, Carbohydrase, and Protease), Livestock (Poultry, Swine, Ruminants, and Aquatic Animals), Source (Microorganism, Plant, and Animal), Form (Dry and Liquid), and Region—Global Forecast to 2025. 2020. Available online: https://www.researchandmarkets.com/reports/5129255/feed-enzymes-market-by-type-phytase?utm_source=GNOM&utm_medium=PressRelease&utm_code=qj4fhb&utm_campaign=1428743+-+Global+Feed+Enzymes+Industry+Report+2020-2025%3a+Dominated+by+BASF%2c+DowDuPont%2c+Koninklijke+DSM%2c+Kemin+Industries%2c+and+Cargill+Incorporated&utm_exec=joca220prd (accessed on 1 September 2021).
- Onwudike, O.C. Palm kernel meal as a feed for poultry. 2. Diets containing palm kernel meal for starter and grower pullets. Anim. Feed Sci. Technol. 1986, 16, 187–194. [Google Scholar] [CrossRef]
- Chong, C.H.; Zulkifli, I.; Blair, R. Effects of dietary inclusion of palm kernel cake and palm oil, and enzyme supplementation on performance of laying hens. Asian Australas. J. Anim. Sci. 2008, 21, 1053–1058. [Google Scholar] [CrossRef]
- Matos, N.; Polanco, R.; De Jesus, C.; Vasquez, R. Effects of palm kernel cake on daily gain and carcass yield of broiler chicks. In Proceedings of the Caribbean Food Crops Society > 44th Annual Meeting, Miami, FL, USA, 13–17 July 2008. [Google Scholar]
- Soltan, M.A. Growth performance, immune response and carcass traits of broiler chicks fed on graded levels of palm kernel cake without or with enzyme supplementation. Livest. Res. Rural Dev. 2009, 21, 37. [Google Scholar]
- Emeka, V.E.; Julius, M.O. Nutritional evaluation of palm kernel meal types: 1. Proximate composition and metabolizable energy values. Afr. J. Biotechnol. 2007, 6, 2484–2486. [Google Scholar] [CrossRef] [Green Version]
- Cerveró, J.M.; Skovgaard, P.A.; Felby, C.; Sørensen, H.R.; Jørgensen, H. Enzymatic hydrolysis and fermentation of palm kernel press cake for production of bioethanol. Enzym. Microb. Technol. 2010, 46, 177–184. [Google Scholar] [CrossRef]
- Breuil, C.; Saddler, J.N. Comparison of the 3,5-dinitrosalicylic acid and Nelson-Somogyi methods of assaying for reducing sugars and determining cellulase activity. Enzym. Microb. Technol. 1985, 7, 327–332. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Arnling Bååth, J.; Martínez-Abad, A.; Berglund, J.; Larsbrink, J.; Vilaplana, F.; Olsson, L. Mannanase hydrolysis of spruce galactoglucomannan focusing on the influence of acetylation on enzymatic mannan degradation. Biotechnol. Biofuels 2018, 11, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bello, B.; Mustafa, S.; Tan, J.S.; Ibrahim, T.A.T.; Tam, Y.J.; Ariff, A.B.; Manap, M.Y.; Abbasiliasi, S. Evaluation of the effect of soluble polysaccharides of palm kernel cake as a potential prebiotic on the growth of probiotics. 3 Biotech 2018, 8, 346. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Liu, H.J.; Shi, Y.Q.; Yan, Q.J.; You, X.; Jiang, Z.Q. Preparation, characterization, and prebiotic activity of manno-oligosaccharides produced from cassia gum by a glycoside hydrolase family 134 β-mannanase. Food Chem. 2020, 309, 125709. [Google Scholar] [CrossRef] [PubMed]
- Kalidas, N.R.; Saminathan, M.; Ismail, I.S.; Abas, F.; Maity, P.; Islam, S.S.; Manshoor, N.; Shaari, K. Structural characterization and evaluation of prebiotic activity of oil palm kernel cake mannanoligosaccharides. Food Chem. 2017, 234, 348–355. [Google Scholar] [CrossRef]
- Jana, U.K.; Kango, N. Characteristics and bioactive properties of mannooligosaccharides derived from agro-waste mannans. Int. J. Biol. Macromol. 2020, 149, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Liepman, A.H.; Nairn, C.J.; Willats, W.G.T.; Sørensen, I.; Roberts, A.W.; Keegstra, K. Functional genomic analysis supports conservation of function among cellulose synthase-like a gene family members and suggests diverse roles of mannans in plants. Plant Physiol. 2007, 143, 1881–1893. [Google Scholar] [CrossRef] [Green Version]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- Schröder, R.; Atkinson, R.G.; Redgwell, R.J. Re-interpreting the role of endo-β-mannanases as mannan endotransglycosylase/hydrolases in the plant cell wall. Ann. Bot. 2009, 104, 197–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daud, M.J.; Jarvis, M.C. Mannan of oil palm kernel. Phytochemistry 1992, 31, 463–464. [Google Scholar] [CrossRef]
- Pedersen, N.R.; Ravn, J.L.; Pettersson, D. A multienzyme NSP product solubilises and degrades NSP structures in canola and mediates protein solubilisation and degradation in vitro. Anim. Feed Sci. Technol. 2017, 234, 244–252. [Google Scholar] [CrossRef]
- Ravn, J.L.; Martens, H.J.; Pettersson, D.; Pedersen, N.R. Enzymatic solubilisation and degradation of soybean fibre demonstrated by viscosity, fibre analysis and microscopy. J. Agric. Sci. 2015, 7, 1. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Pineiro, M.; Asp, N.G.; Reid, G.; Macfarlane, S.; Morelli, L.; Brunser, O.; Tuohy, K. FAO Technical meeting on prebiotics. J. Clin. Gastroenterol. 2008, 42 (Suppl. S3), S156–S159. [Google Scholar] [CrossRef] [PubMed]
- Oyofo, B.A.; DeLoach, J.R.; Corrier, D.E.; Norman, J.O.; Ziprin, R.L.; Mollenhauer, H.H. Prevention of Salmonella typhimurium colonization of broilers with D-mannose. Poult. Sci. 1989, 68, 1357–1360. [Google Scholar] [CrossRef] [PubMed]
- Spring, P.; Wenk, C.; Dawson, K.A.; Newman, K.E. The effects of dietary mannanoligosaccharides on cecal parameters and the concentrations of enteric bacteria in the ceca of salmonella-challenged broiler chicks. Poult. Sci. 2000, 79, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Agunos, A.; Ibuki, M.; Yokomizo, F.; Mine, Y. Effect of dietary β 1–4 mannobiose in the prevention of Salmonella enteritidis infection in broilers. Br. Poult. Sci. 2007, 48, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, N.; Chu, D.C.; Akachi, S.; Juneja, L.R. Preventive effect of partially hydrolyzed guar gum on infection of Salmonella enteritidis in young and laying hens. Poult. Sci. 2000, 79, 689–697. [Google Scholar] [CrossRef]
- Jahromi, M.F.; Shokryazdan, P.; Idrus, Z.; Ebrahimi, R.; Bashokouh, F.; Liang, J.B. Modulation of immune function in rats using oligosaccharides extracted from palm kernel cake. Biomed. Res. Int. 2017, 2017, 2576921. [Google Scholar] [CrossRef]
- Ofek, I.; Mirelman, D.; Sharon, N. Adherence of Escherichia coli to human mucosal cells mediated by mannose receptors. Nature 1977, 265, 623–625. [Google Scholar] [CrossRef]
- Zang, H.; Xie, S.; Zhu, B.; Yang, X.; Gu, C.; Hu, B.; Gao, T.; Chen, Y.; Gao, X. Mannan oligosaccharides trigger multiple defence responses in rice and tobacco as a novel danger-associated molecular pattern. Mol. Plant Pathol. 2019, 20, 1067–1079. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Tao, Y.; Lai, C.; Huang, C.; Zhou, Y.; Yong, Q. Effects of mannanoligosaccharide supplementation on the growth performance, immunity, and oxidative status of partridge shank chickens. Animals 2019, 9, 817. [Google Scholar] [CrossRef] [Green Version]
- Mussini, F.J.; Coto, C.A.; Goodgame, S.D.; Lu, C.; Karimi, A.J.; Lee, J.H.; Waldroup, P.W. Effect of ß-mannanase on broiler performance and dry matter output using corn soybean meal based diets. Int. J. Poult. Sci. 2011, 10, 778–781. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.S.; Kim, M.C.; Martinez-Pitargue, F.; Choi, H.S.; Kil, D.Y. Dietary β-mannanase decreases cloacal temperature of broiler chickens under hot conditions without affecting growth performance. Rev. Colomb. Ciencias Pecu. 2019, 32, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Ryu, M.H.; Hosseindoust, A.; Kim, J.S.; Choi, Y.H.; Lee, S.H.; Kim, M.J.; Lee, J.H.; Chae, B.J. β-mannanase derived from Bacillus Subtilis WL-7 improves the performance of commercial laying hens fed low or high mannan-based diets. J. Poult. Sci. 2017, 54, 212–217. [Google Scholar] [CrossRef] [PubMed]
- White, D.; Adhikari, R.; Wang, J.; Chen, C.; Lee, J.H.; Kim, W.K. Effects of dietary protein, energy and β-mannanase on laying performance, egg quality, and ileal amino acid digestibility in laying hens. Poult. Sci. 2021, 100, 101312. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods. Available online: http://www.aoacofficialmethod.org (accessed on 30 September 2021).
- Thiex, N.J.; Anderson, S.; Gildemeister, B.; Adcock, W.; Boedigheimer, J.; Bogren, E.; Coffin, R.; Conway, K.; DeBaker, A.; Frankenius, E.; et al. Crude fat, hexanes extraction, in feed, cereal grain, and forage (Randall/Soxtec/Submersion method): Collaborative Study. J. AOAC Int. 2003, 86, 899–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, R.A. Generic combustion method for determination of crude protein in feeds: Collaborative study. J. AOAC Int. 1989, 72, 770–774. [Google Scholar] [CrossRef]
- McCleary, B.V.; McGeough, P. A Comparison of polysaccharide substrates and reducing sugar methods for the measurement of endo-1,4-β-Xylanase. Appl. Biochem. Biotechnol. 2015, 177, 1152–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wider, G.; Dreier, L. Measuring protein concentrations by NMR spectroscopy. J. Am. Chem. Soc. 2006, 128, 2571–2576. [Google Scholar] [CrossRef]
- De Souza, A.C.; Rietkerk, T.; Selin, C.G.M.; Lankhorst, P.P. A robust and universal NMR method for the compositional analysis of polysaccharides. Carbohydr. Polym. 2013, 95, 657–663. [Google Scholar] [CrossRef]
- Wimmer, R.; Wider, G. Real-time imaging of the spatial distribution of rf-heating in NMR samples during broadband decoupling. J. Magn. Reson. 2007, 187, 184–192. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monosaccharide | % (w/w) of Total Carbohydrate b,c | |
|---|---|---|
| mannose | 11.53 ± 0.59% | 65.2 ± 3.31% |
| glucose | 4.25 ± 0.36% | 24.0 ± 2.01% |
| galactose | 0.69 ± 0.10% | 3.9 ± 0.54% |
| xylose | 0.84 ± 0.06% | 4.8 ± 0.32% |
| arabinose | 0.30 ± 0.02% | 1.7 ± 0.11% |
| rhamnose | 0.08 ± 0.03% | 0.5 ± 0.18% |
| Treatment | Reducing Sugars b,c A540 [a.u.] | CMOS c,i in Supernatant [mM] | [-] |
|---|---|---|---|
| Control | 0.37 0.03 | 0.630.04 d | 5.170.22 |
| B. subtilis mannanase 1× a | 0.44 0.05 | 1.27 0.04 e | 3.630.13 |
| B. subtilis mannanase 2× a | 0.52 0.02 | 0.04 f | 3.270.10 |
| B. subtilis mannanase 5× a | 0.60 0.03 | 2.06 0.03 g | 3.140.1 |
| B. subtilis mannanase 20× a | ± 0.02 | 2.71 0.10 h | 2.920.09 |
| Treatment | [g/mol] b | x (% mannan solubilized) e,f | ||
|---|---|---|---|---|
| Control | 856.28 | 6.38 ± 0.27 | 0.0 | 0.0% |
| B. subtilis mannanase 500 ppm a | 606.59 | 9.27 ± 0.28 | 2.89 ± 0.43 | 1.82 ± 0.27% |
| B. subtilis mannanase 1000 ppm a | 540.11 | 11.06 ± 0.24 | 4.68 ± 0.27 | 2.95 ± 0.17% |
| B. subtilis mannanase 2500 ppm a | 509.30 | 12.57 ± 0.21 | 6.20 ± 0.15 | 3.89 ± 0.09% |
| B. subtilis mannanase 10,000 ppm a | 486.60 | 15.81 ± 0.59 | 9.44 ± 0.79 | 5.91 ± 0.50% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez-Osorio, L.-M.; Nielsen, J.U.; Martens, H.J.; Wimmer, R. Upgrading the Nutritional Value of PKC Using a Bacillus subtilis Derived Monocomponent β-Mannanase. Molecules 2022, 27, 563. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27020563
Gomez-Osorio L-M, Nielsen JU, Martens HJ, Wimmer R. Upgrading the Nutritional Value of PKC Using a Bacillus subtilis Derived Monocomponent β-Mannanase. Molecules. 2022; 27(2):563. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27020563
Chicago/Turabian StyleGomez-Osorio, Luis-Miguel, Janni Ulnits Nielsen, Helle Jakobe Martens, and Reinhard Wimmer. 2022. "Upgrading the Nutritional Value of PKC Using a Bacillus subtilis Derived Monocomponent β-Mannanase" Molecules 27, no. 2: 563. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27020563