
α-Glucosidase Inhibitory Activity of Fermented Okara Broth Started with the Strain Bacillus amyloliquefaciens SY07

{kind=link}
{kind=link}
{kind=link}
{kind=link}
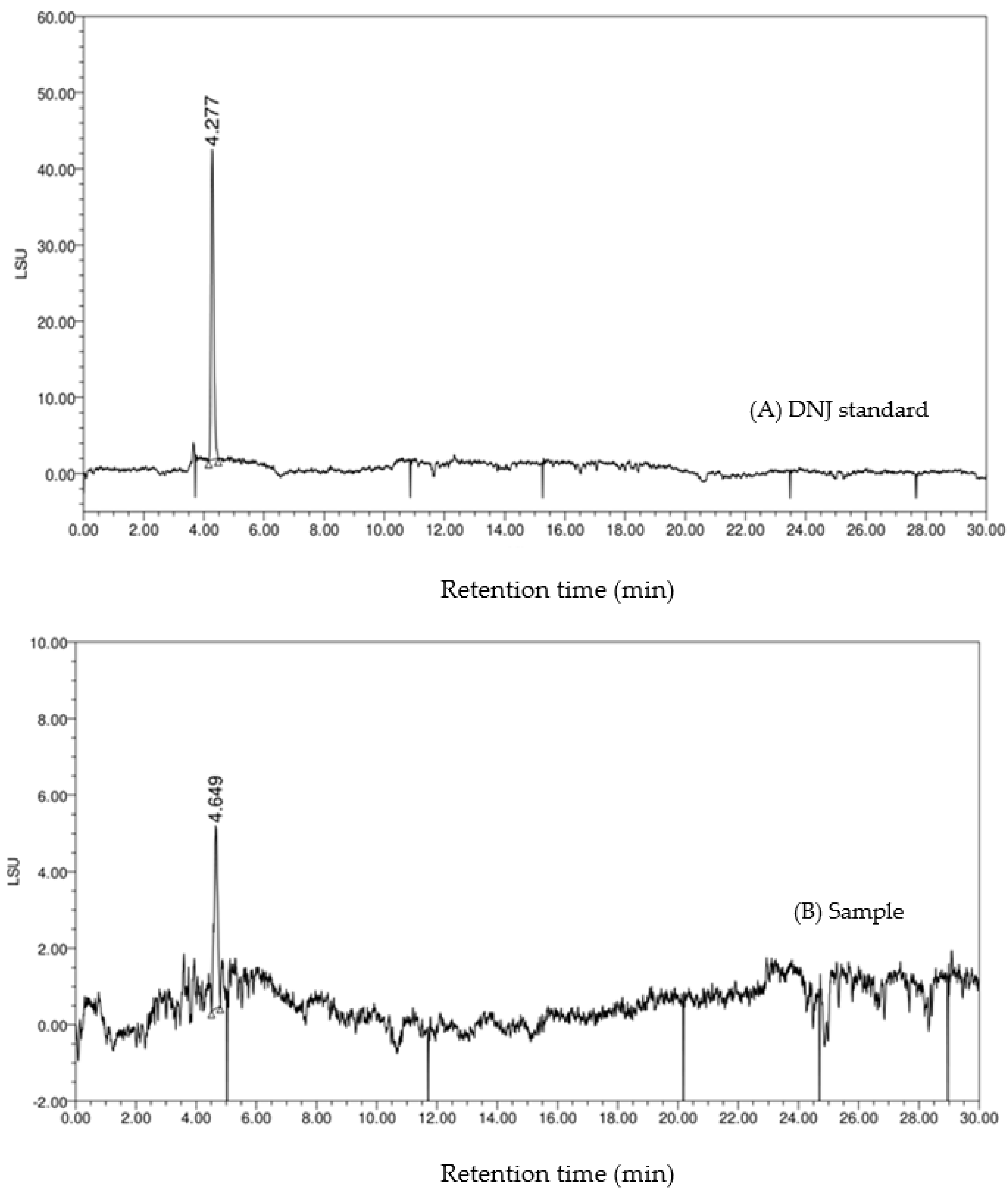{kind=link}
{kind=link}
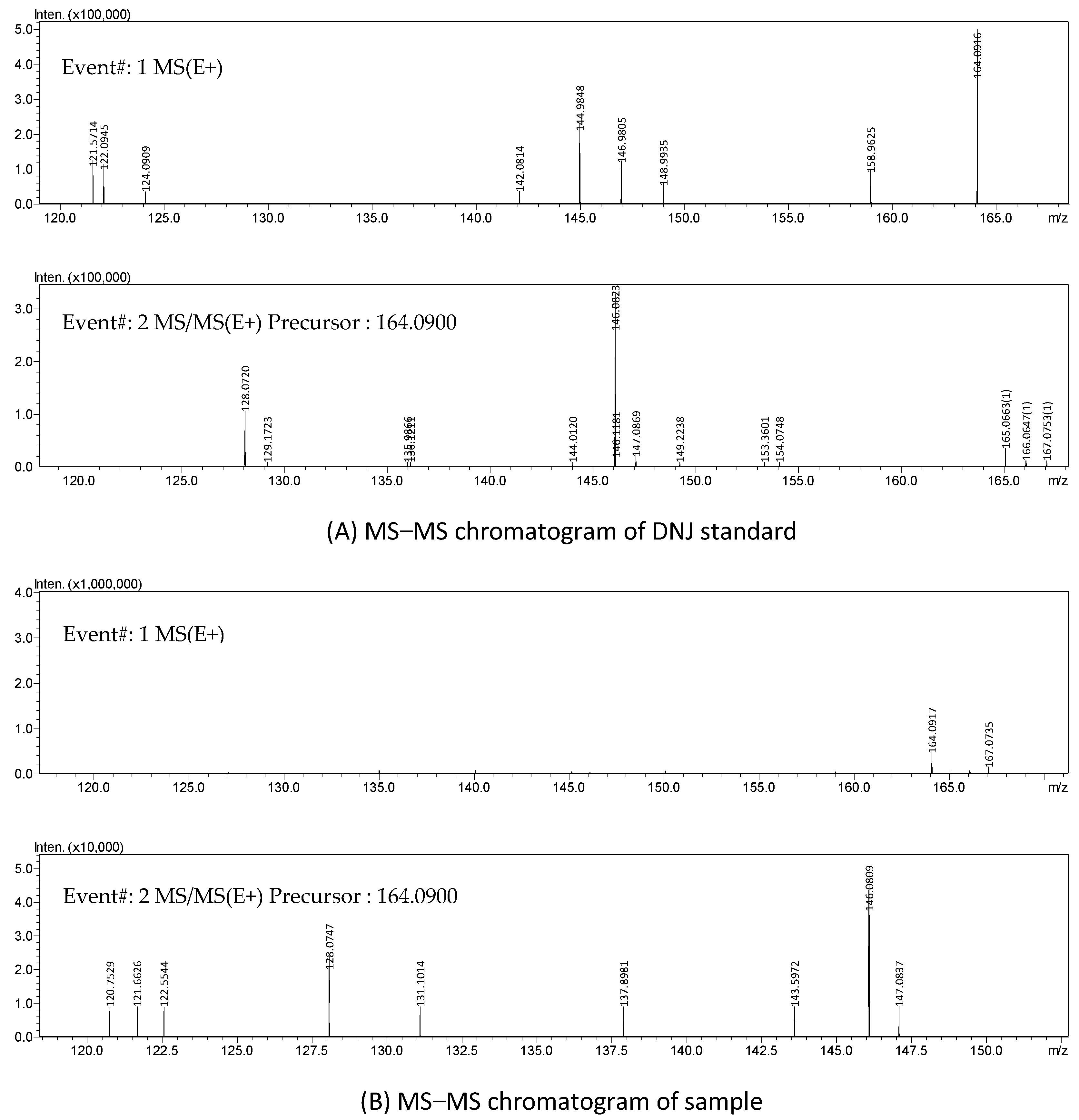{kind=link}
Abstract
:1. Introduction
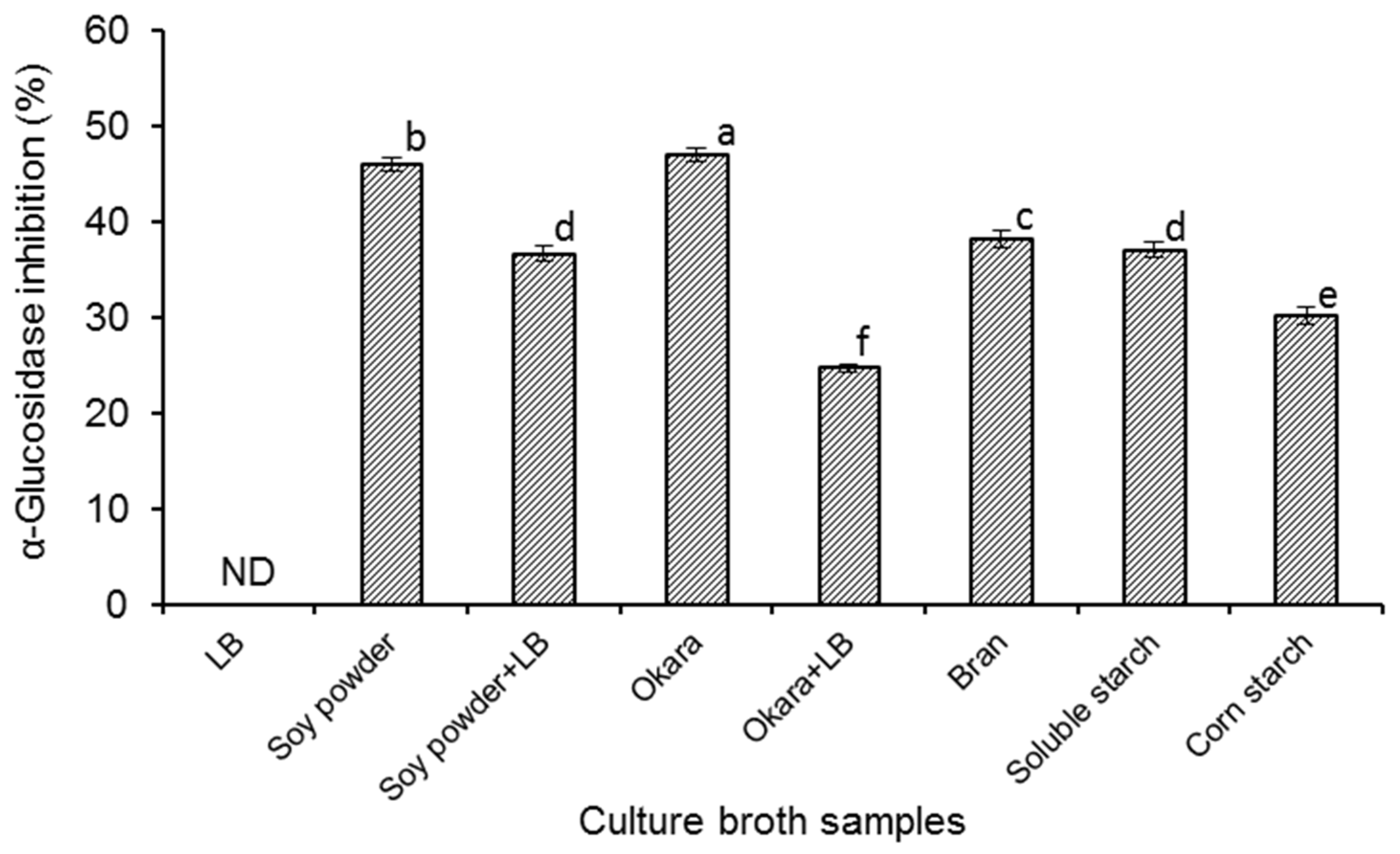
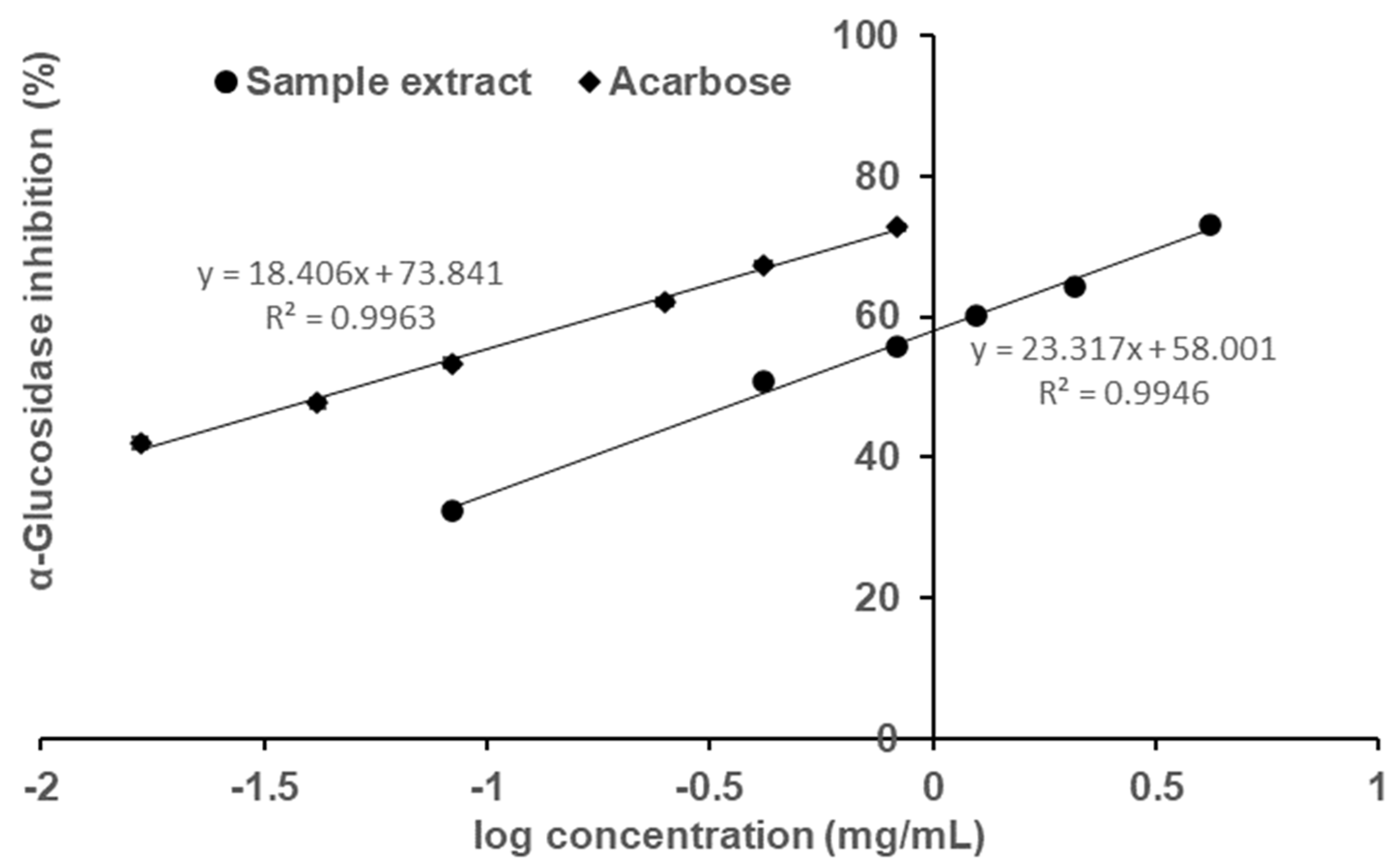
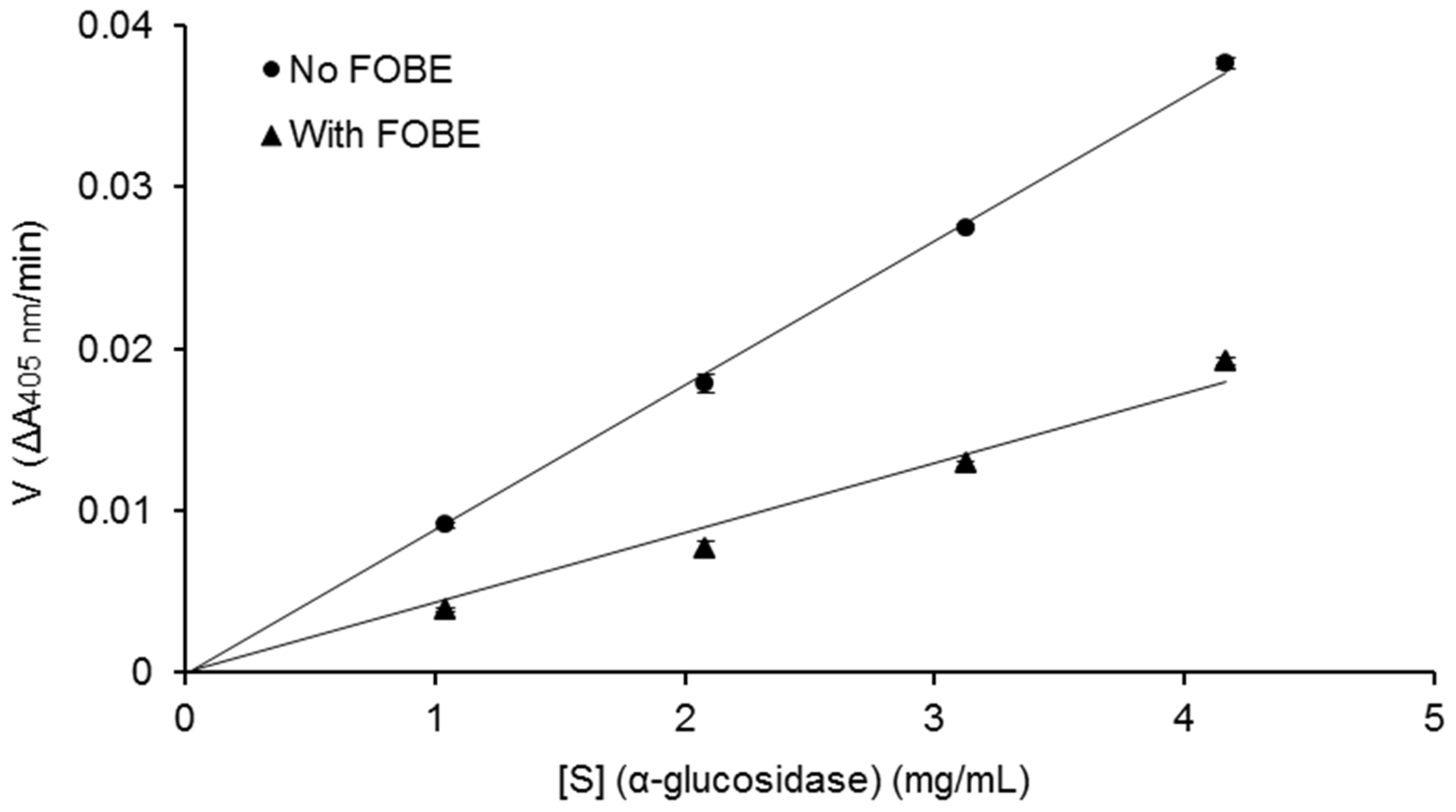
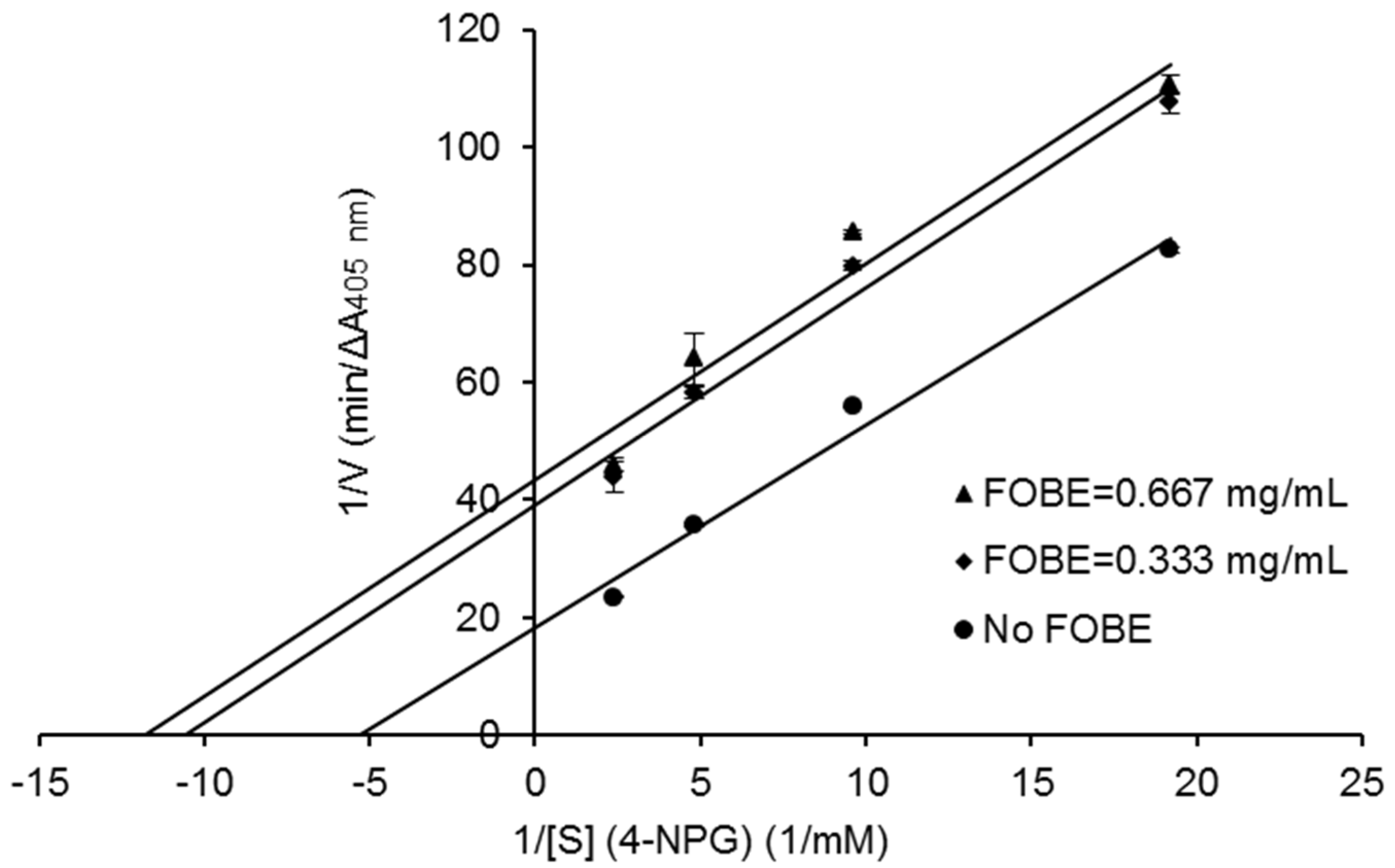
2. Results and Discussion
3. Materials and Methods
3.1. Microorganism and Materials
3.2. Preparation of Starter Culture
3.3. Production of α-Glucosidase Inhibitors in Different Growth Media
3.4. Preparation of Sample Extracts
3.5. Assay for α-Glucosidase Inhibitory Activity
3.6. Separation and Purification
3.7. High-Performance Liquid Chromatography (HPLC) and Mass-Mass Spectrometer (MS-MS) Analysis
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Ni, M.; Hu, X.; Gong, D.; Zhang, G. Inhibitory mechanism of vitexin on α-glucosidase and its synergy with acarbose. Food Hydrocoll. 2020, 105, 105824. [Google Scholar] [CrossRef]
- Feng, J.; Yang, X.W.; Wang, R.F. Bio-assay guided isolation and identification of α-glucosidase inhibitors from the leaves of Aquilaria sinensis. Phytochemistry 2011, 72, 242–247. [Google Scholar] [CrossRef]
- Liu, C.; Xiang, W.; Yu, Y.; Shi, Z.; Huang, X.; Xu, L. Comparative analysis of 1-deoxynojirimycin contribution degree to α-glucosidase inhibitory activity and physiological distribution in Morus alba L. Ind. Crops Prod. 2015, 70, 309–315. [Google Scholar] [CrossRef]
- Rosas-Ramírez, D.; Escandón-Rivera, S.; Pereda-Miranda, R. Morning glory resin glycosides as α-glucosidase inhibitors: In Vitro and in silico analysis. Phytochemistry 2018, 148, 39–47. [Google Scholar] [CrossRef]
- Liu, Z.X.; Liu, C.T.; Liu, Q.B.; Ren, J.; Li, L.Z.; Huang, X.X.; Wang, Z.Z.; Song, S.J. Iridoid glycosides from the flower buds of Lonicera japonica and their nitric oxide production and α-glucosidase inhibitory activities. J. Funct. Foods 2015, 18, 512–519. [Google Scholar] [CrossRef]
- Choi, M.; Kang, Y.R.; Zu, H.D.; Lim, I.S.; Jung, S.K.; Chang, Y.H. Effects of time on phenolics and in vitro bioactivity in autoclave extraction of Graviola (Annona muricata) leaf. Biotechnol. Bioprocess Eng. 2020, 25, 9–15. [Google Scholar] [CrossRef]
- Ifie, I.; Ifie, B.E.; Ibitoye, D.O.; Marshall, L.J.; Williamson, G. Seasonal variation in Hibiscus sabdariffa (Roselle) calyx phytochemical profile, soluble solids and α-glucosidase inhibition. Food Chem. 2018, 261, 164–168. [Google Scholar] [CrossRef]
- Chelladurai, G.R.M.; Chinnachamy, C. Alpha amylase and alpha glucosidase inhibitory effects of aqueous stem extract of Salacia oblonga and its GC-MS analysis. Braz. J. Pharm. Sci. 2018, 54, 1–10. [Google Scholar] [CrossRef]
- Sun, S.; Kadouh, H.C.; Zhu, W.; Zhou, K. Bioactivity-guided isolation and purification of α-glucosidase inhibitor, 6-O-D-glycosides, from Tinta Cão grape pomace. J. Funct. Foods 2016, 23, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T.; Inouye, S.; Tsuruoka, T.; Koaze, Y.; Niida, T. “Nojirimycin” as a potent inhibitor of glucosidase. Agric. Biol. Chem. 1970, 34, 966–968. [Google Scholar] [CrossRef]
- Schmidt, D.D.; Frommer, W.; Junge, B.; Müller, L.; Wingender, W.; Truscheit, E.; Schäfer, D. α-Glucosidase inhibitors—New complex oligosaccharides of microbial origin. Naturwissenschaften 1977, 64, 535–536. [Google Scholar] [CrossRef] [PubMed]
- Kanieda, Y.; Asano, N.; Teranishi, M.; Matsui, K. New cyclitols, degradation of validamycin A by Flavobacterium saccharophilum. J. Antibiot. 1980, 33, 1573–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.P.; Yamaki, K.; Yoshihashi, T.; Ohnishi Kameyama, M.; Li, X.T.; Cheng, Y.Q.; Mori, Y.; Li, L.-T. Purification and identification of 1-deoxynojirimycin (DNJ) in Okara fermented by Bacillus subtilis B2 from Chinese traditional food (Meitaoza). J. Agric. Food Chem. 2010, 58, 4097–4103. [Google Scholar] [CrossRef] [PubMed]
- Onose, S.; Ikeda, R.; Nakagawa, K.; Kimura, T.; Yamagishi, K.; Higuchi, O.; Miyazawa, T. Production of the α-glycosidase inhibitor 1-deoxynojirimycin from Bacillus species. Food Chem. 2013, 138, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Shin, H.H.; Kim, H.R.; Nam, Y.D.; Seo, D.H.; Seo, M.J. Culture optimization strategy for 1-Deoxynojirimycin-producing Bacillus methylotrophicus K26 isolated from Korean fermented soybean paste, Doenjang. Biotechnol. Bioprocess Eng. 2018, 23, 424–431. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Wang, S.L. New novel α-glucosidase inhibitors produced by microbial conversion. Process Biochem. 2018, 65, 228–232. [Google Scholar] [CrossRef]
- Fang, Y.; Li, F.; Chen, M.; Li, Z.; Wang, C. Optimization of liquid fermentation conditions for the production of α-glucosidase inhibitors by Bacillus amyloliquefaciens SY07. China Condiment 2016, 41, 10–14. (In Chinese) [Google Scholar]
- Muganga, L.; Liu, X.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Screening for lactic acid bacteria based on antihyperglycaemic and probiotic potential and application in synbiotic set yoghurt. J. Funct. Foods 2015, 16, 125–136. [Google Scholar] [CrossRef]
- Zhu, Y.P.; Yin, L.J.; Cheng, Y.Q.; Yamaki, K.; Mori, Y.; Su, Y.C.; Li, L.T. Effects of sources of carbon and nitrogen on production of α-glucosidase inhibitor by a newly isolated strain of Bacillus subtilis B2. Food Chem. 2008, 109, 737–742. [Google Scholar] [CrossRef]
- Peng, Y.; Huang, Q.; Zhang, R.H.; Zhang, Y.Z. Purification and characterization of a fibrinolytic enzyme produced by Bacillus amyloliquefaciens DC-4 screened from douchi, a traditional Chinese soybean food. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 134, 45–52. [Google Scholar] [CrossRef]
- Kaewklom, S.; Lumlert, S.; Kraikul, W.; Aunpad, R. Control of Listeria monocytogenes on sliced bologna sausage using a novel bacteriocin, amysin, produced by Bacillus amyloliquefaciens isolated from Thai shrimp paste (Kapi). Food Control 2013, 32, 552–557. [Google Scholar] [CrossRef]
- Cui, J.; Gu, X.; Wang, F.; Ouyang, J.; Wang, J. Purification and structural characterization of an α-glucosidase inhibitory polysaccharide from apricot (Armeniaca sibirica L. Lam.) pulp. Carbohydr. Polym. 2015, 121, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, K.; Onose, S.; Takasu, S.; Ito, J.; Ikeda, R.; Kimura, T.; Nakagawa, K.; Miyazawa, T. Lactose increases the production of 1-deoxynojirimycin in Bacillus amyloliquefaciens. Food Sci. Technol. Res. 2017, 23, 349–353. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Kim, S.U.; Lee, H.S.; Kim, I.; Ahn, M.Y.; Ryu, K.S. Determination of 1-deoxynojirimycin in Morus alba L. leaves by derivatization with 9-fluorenylmethyl chloroformate followed by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 2003, 1002, 93–99. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Bian, W.; Fang, Y.; Du, P.; Liu, X.; Zhao, X.; Li, F. α-Glucosidase Inhibitory Activity of Fermented Okara Broth Started with the Strain Bacillus amyloliquefaciens SY07. Molecules 2022, 27, 1127. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27031127
Gao Y, Bian W, Fang Y, Du P, Liu X, Zhao X, Li F. α-Glucosidase Inhibitory Activity of Fermented Okara Broth Started with the Strain Bacillus amyloliquefaciens SY07. Molecules. 2022; 27(3):1127. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27031127
Chicago/Turabian StyleGao, Yifang, Wenhui Bian, Yuanyuan Fang, Peng Du, Xueting Liu, Xueru Zhao, and Fengjuan Li. 2022. "α-Glucosidase Inhibitory Activity of Fermented Okara Broth Started with the Strain Bacillus amyloliquefaciens SY07" Molecules 27, no. 3: 1127. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27031127