
The Root Extract of Peucedanum praeruptorum Dunn Exerts Anticancer Effects in Human Non-Small-Cell Lung Cancer Cells with Different EGFR Mutation Statuses by Suppressing MET Activity


Abstract
:1. Introduction
2. Results
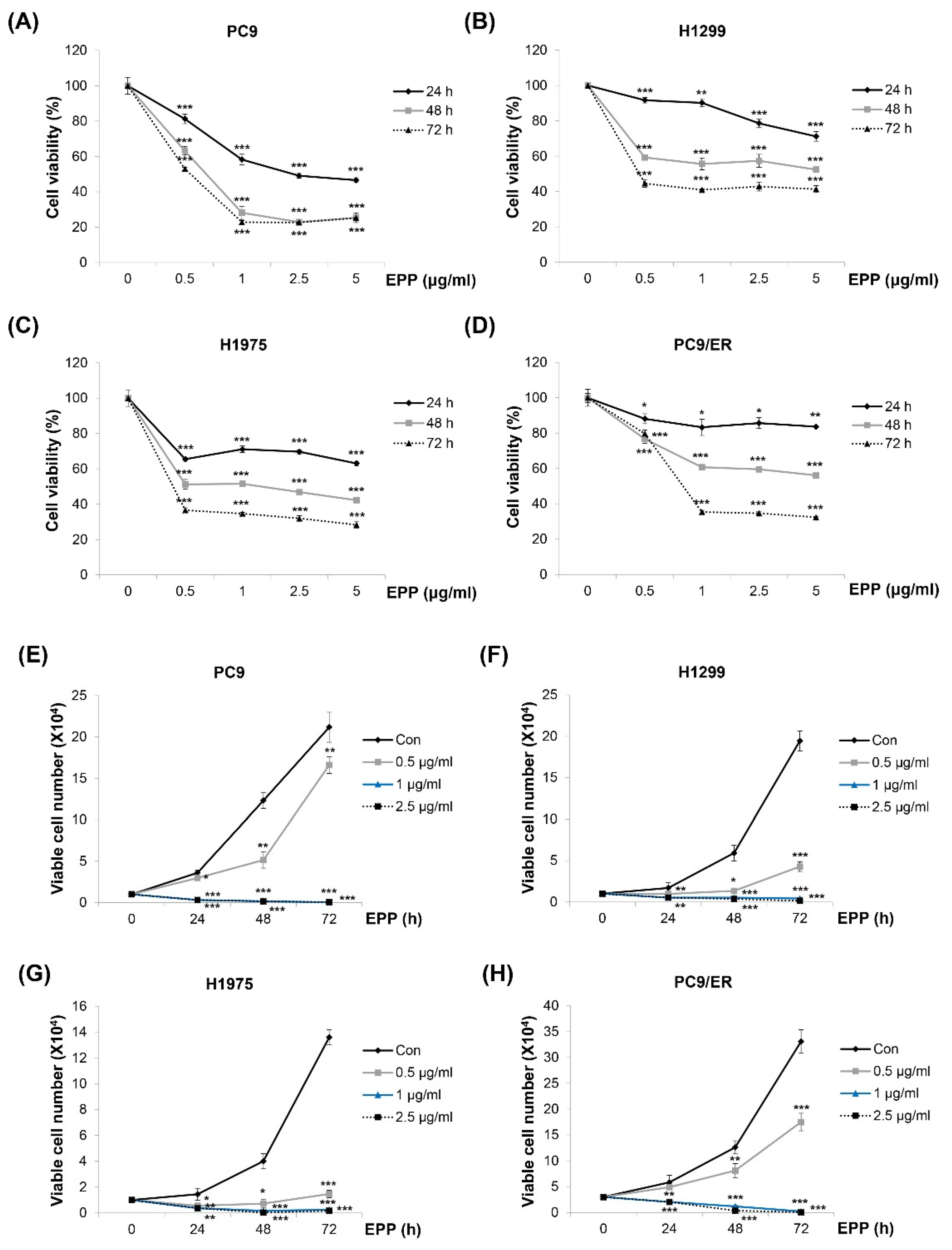
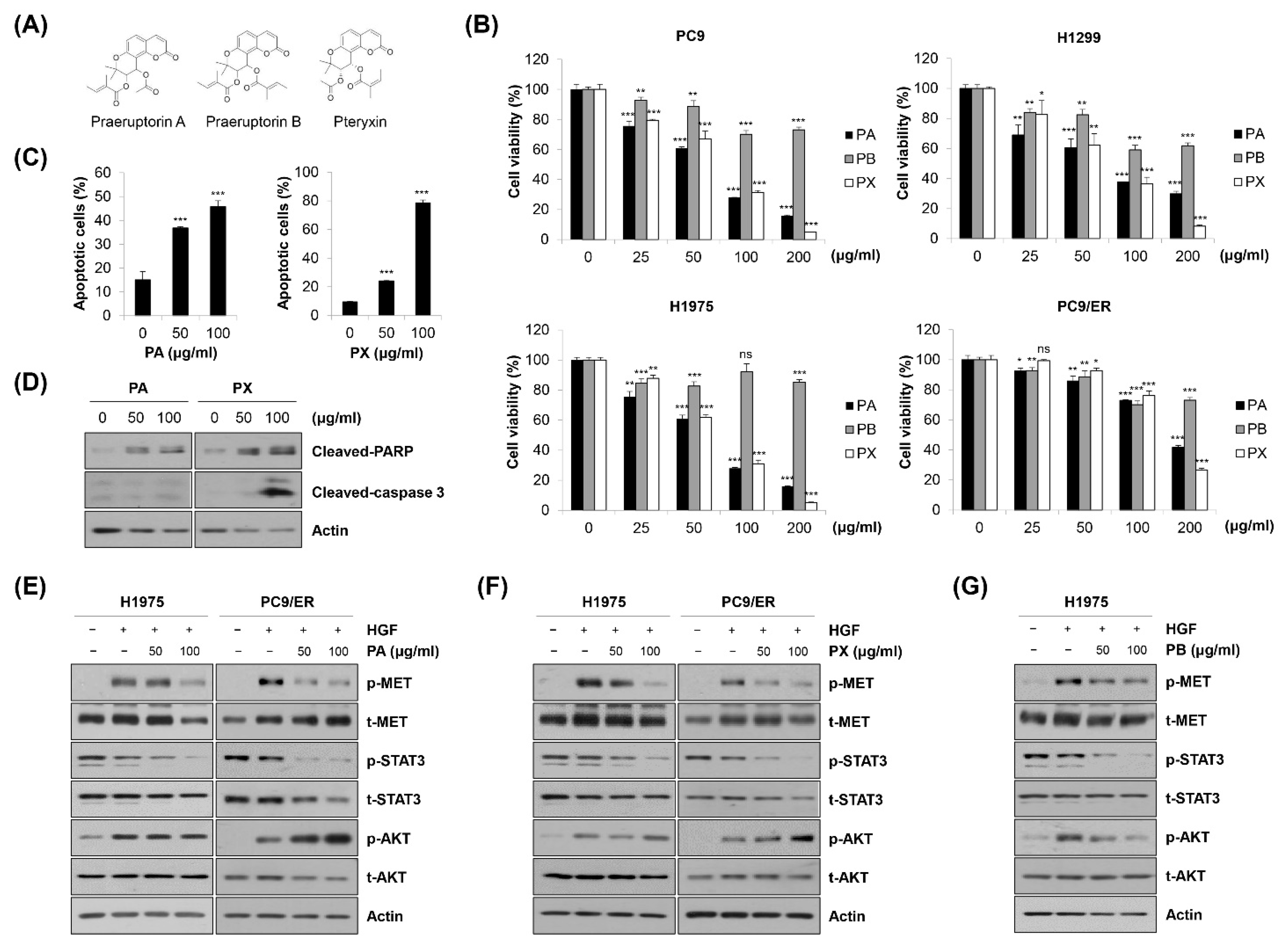
2.1. Suppression of Cell Growth by EPP in NSCLC Cells
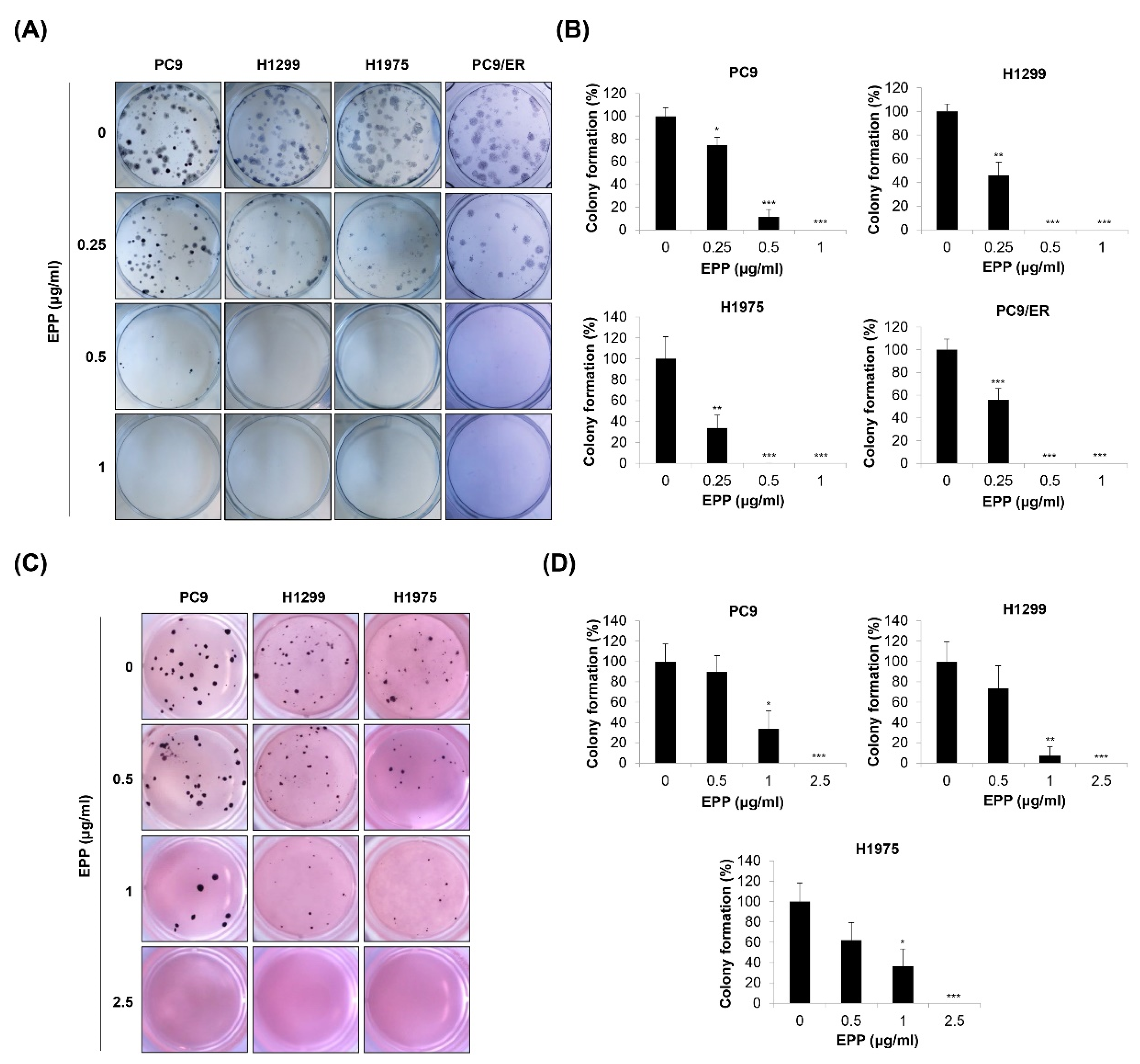
2.2. Inhibition of Colony Formation by EPP in NSCLC Cells
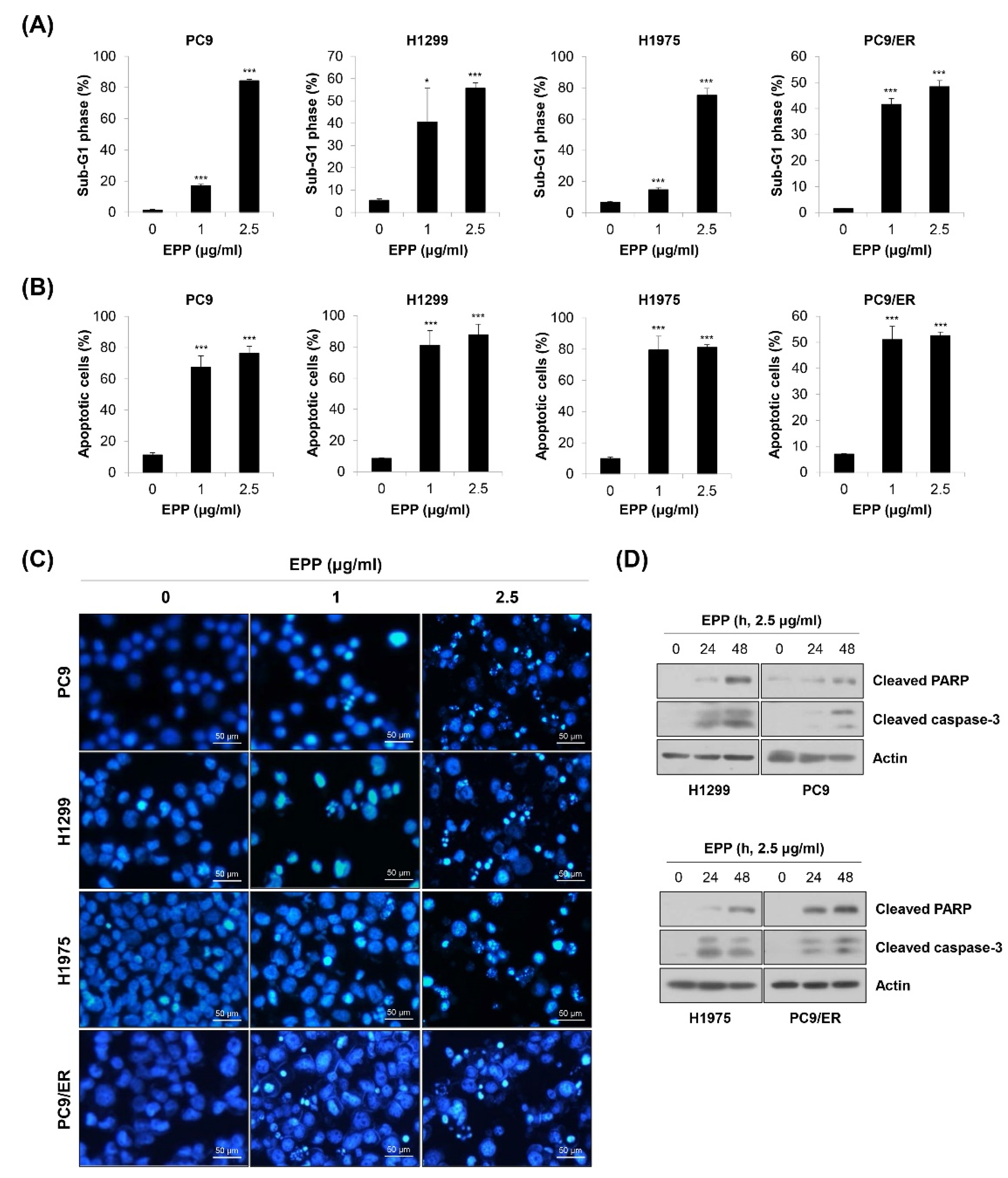
2.3. Induction of Apoptosis by EPP in NSCLC Cells
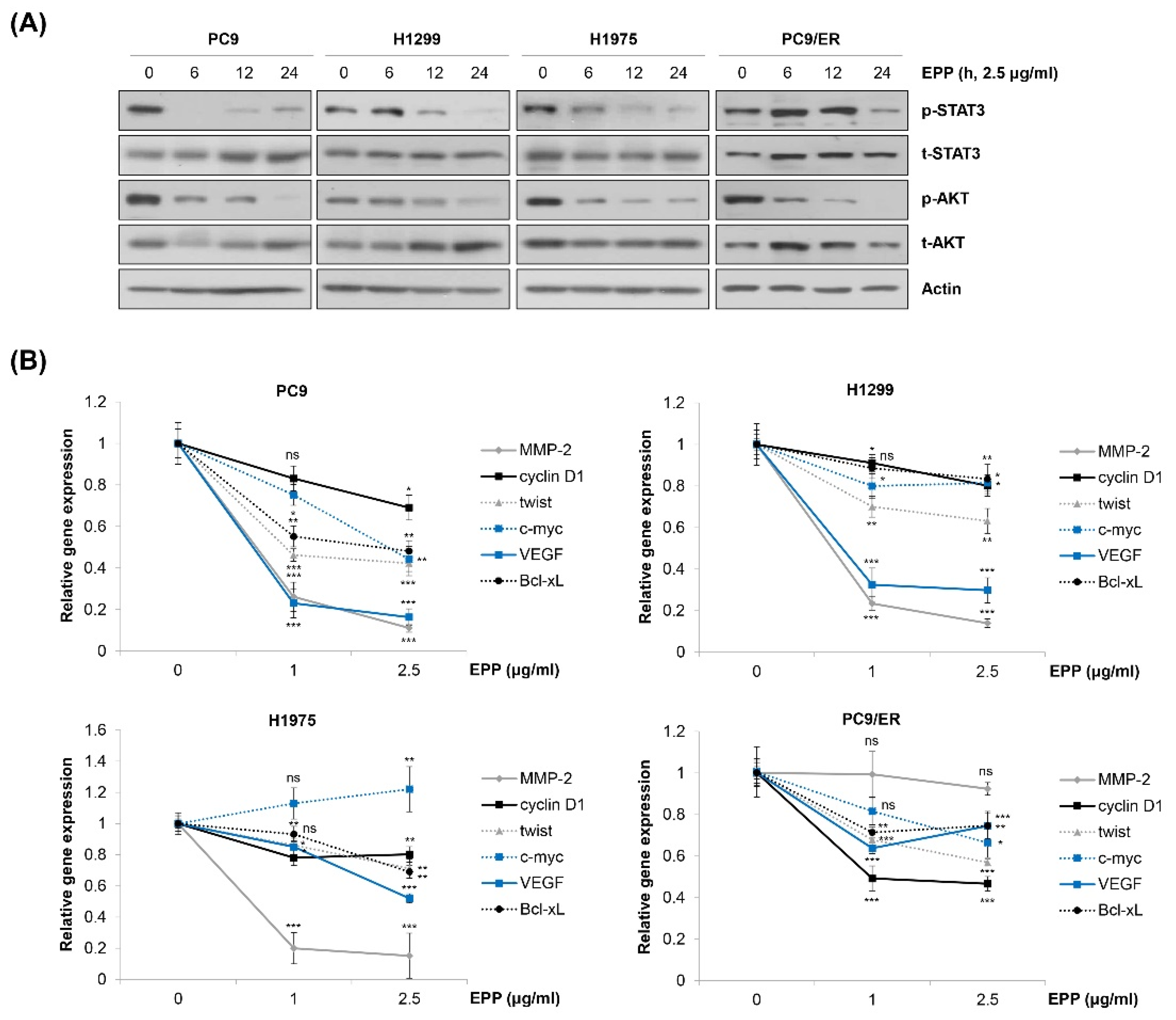
2.4. Inactivation of STAT3 and AKT by EPP in NSCLC Cells
2.5. Blockage of the MET Signaling Pathway by EPP in NSCLC Cells
2.6. Identification of Specific Compounds Contributing to the Anticancer Effect of EPP
3. Discussion
4. Materials and Methods
4.1. Preparation of EPP
4.2. HPLC-MS Analysis
4.3. Cell Culture
4.4. MTT Assay
4.5. Trypan Blue Exclusion Assay
4.6. Colony Formation Assay
4.7. Flow Cytometry
4.8. DAPI Staining
4.9. Western Blot Analysis
4.10. Reverse Transcription–Quantitative Polymerase Chain Reaction (RT-qPCR)
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Yuan, J.Q.; Wang, K.F.; Fu, X.H.; Han, X.R.; Threapleton, D.; Yang, Z.Y.; Mao, C.; Tang, J.L. The prevalence of EGFR mutation in patients with non-small cell lung cancer: A systematic review and meta-analysis. Oncotarget 2016, 7, 78985–78993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, U.; Sharma, S.V.; Settleman, J. High-Throughput lung cancer cell line screening for genotype-correlated sensitivity to an EGFR kinase inhibitor. Methods Enzymol. 2008, 438, 331–341. [Google Scholar]
- Scaltriti, M.; Baselga, J. The epidermal growth factor receptor pathway: A model for targeted therapy. Clin. Cancer Res. 2006, 12, 5268–5272. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Wu, Y.L.; Chen, G.; Feng, J.; Liu, X.Q.; Wang, C.; Zhang, S.; Wang, J.; Zhou, S.; Ren, S.; et al. Erlotinib versus chem-otherapy as first-line treatment for patients with advanced EGFR mutation-positive non-small-cell lung cancer (OPTIMAL, CTONG-0802): A multicentre, open-label, randomised, phase 3 study. Lancet Oncol. 2011, 12, 735–742. [Google Scholar] [CrossRef]
- Rosell, R.; Carcereny, E.; Gervais, R.; Vergnenegre, A.; Massuti, B.; Felip, E.; Palmero, R.; Garcia-Gomez, R.; Pallares, C.; Sanchez, J.M.; et al. Erlotinib versus standard chemotherapy as first-line treatment for European patients with advanced EGFR mutation-positive non-small-cell lung cancer (EURTAC): A multicentre, open-label, randomised phase 3 trial. Lancet Oncol. 2012, 13, 239–246. [Google Scholar] [CrossRef]
- Maemondo, M.; Inoue, A.; Kobayashi, K.; Sugawara, S.; Oizumi, S.; Isobe, H.; Gemma, A.; Harada, M.; Yoshizawa, H.; Ki-noshita, I.; et al. Gefitinib or chemotherapy for non-small-cell lung cancer with mutated EGFR. N. Engl. J. Med. 2010, 362, 2380–2388. [Google Scholar] [CrossRef] [Green Version]
- Mitsudomi, T.; Morita, S.; Yatabe, Y.; Negoro, S.; Okamoto, I.; Tsurutani, J.; Seto, T.; Satouchi, M.; Tada, H.; Hirashima, T.; et al. Gefitinib versus cisplatin plus docetaxel in patients with non-small-cell lung cancer harbouring mutations of the epi-dermal growth factor receptor (WJTOG3405): An open label, randomised phase 3 trial. Lancet Oncol. 2010, 11, 121–128. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Zhong, W.Z.; Zhang, X.C.; Su, J.; Yang, X.N.; Chan, Z.H.; Yang, J.J.; Zhou, Q.; Yan, H.H.; An, S.-J.; et al. Egfr mutation heterogeneity and the mixed response to EGFR tyrosine kinase inhibitors of lung adenocarcinomas. Oncologist 2012, 17, 978–985. [Google Scholar] [CrossRef] [Green Version]
- Passaro, A.; Malapelle, U.; Del, R.M.; Attili, I.; Russo, A.; Guerini, R.E.; Fumagalli, C.; Pisapia, P.; Pepe, F.; De, L.C.; et al. Understanding EGFR heterogeneity in lung cancer. ESMO Open 2020, 5, e000919. [Google Scholar] [CrossRef]
- Taniguchi, K.; Okami, J.; Kodama, K.; Higashiyama, M.; Kato, K. Intratumor heterogeneity of epidermal growth factor receptor mutations in lung cancer and its correlation to the response to gefitinib. Cancer Sci. 2008, 99, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Yatabe, Y.; Matsuo, K.; Mitsudomi, T. Heterogeneous distribution of EGFR mutations is extremely rare in lung adenocarcinoma. J. Clin. Oncol. 2011, 29, 2972–2977. [Google Scholar] [CrossRef]
- Yu, H.A.; Arcila, M.E.; Rekhtman, N.; Sima, C.S.; Zakowski, M.F.; Pao, W.; Kris, M.G.; Miller, V.A.; Ladanyi, M.; Riely, G.J. Analysis of Tumor Specimens at the Time of Acquired Resistance to EGFR-TKI Therapy in 155 Patients with EGFR-Mutant Lung Cancers. Clin. Cancer Res. 2013, 19, 2240–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.-H.; Lu, J.-J. Osimertinib resistance in non-small cell lung cancer: Mechanisms and therapeutic strategies. Cancer Lett. 2018, 420, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Chong, C.R.; Jänne, A.P. The quest to overcome resistance to EGFR-targeted therapies in cancer. Nat. Med. 2013, 19, 1389–1400. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Jing, W.; Yan, R.; Wang, Y. Research progress of the studies on the roots of Peucedanum praeruptorum dunn (Peucedani radix). Pak. J. Pharm. Sci. 2015, 28, 71–81. [Google Scholar]
- Lee, J.; Lee, Y.J.; Kim, J.; Bang, O.S. Pyranocoumarins from root extracts of Peucedanum praeruptorum Dunn with multidrug resistance reversal and anti-inflammatory activities. Molecules 2015, 12, 20967–20978. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.Y.; Wu, F.H.; Wang, J.S.; Kang, Y.L. Attenuation of airway hyperreactivity and T helper cell type 2 responses by coumarins from Peucedanum praeruptorum Dunn in a murine model of allergic airway inflammation. J. Ethnopharmacol. 2012, 141, 314–321. [Google Scholar] [CrossRef]
- Ji, Y.; Rao, M.R. Effects of Peucedanum praeruptorum extractum on blood pressure, left ventricular hypertrophy and hemodynamic changes in left ventricular hypertrophied hypertensive rats. Zhongguo Zhongxiyi Jiehe Zazhi 1996, 6, 676–678. [Google Scholar]
- Lee, A.R.; Chun, J.M.; Lee, A.Y.; Kim, H.S.; Gu, G.J.; Kwon, B.I. Reduced allergic lung inflammation by root extracts from two species of Peucedanum through inhibition of Th2 cell activation. J. Ethnopharmacol. 2017, 196, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.H.; Wang, M.Z.; Liu, J.Q. Chemical constituents of the volatile oil from the roots of Peucedanum praeruptorum and its antibacterial activities. Redai Yaredai Zhiwu Xuebao 2002, 4, 366–370. [Google Scholar]
- Liu, R.; Feng, L.; Sun, A.; Kong, L. Preparative isolation and purification of coumarins from Peucedanum praeruptorum Dunn by high-speed counter-current chromatography. J. Chromatogr. A 2004, 1057, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Sos, M.L.; Koker, M.; Weir, B.A.; Heynck, S.; Rabinovsky, R.; Zander, T.; Seeger, J.M.; Weiss, J.; Fischer, F.; Frommolt, P.; et al. PTEN loss contributes to erlotinib resistance in EGFR-mutant lung cancer by activation of Akt and EGFR. Cancer Res. 2009, 69, 3256–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulkifli, A.A.; Tan, F.H.; Putoczki, T.L.; Stylli, S.S.; Luwor, R.B. STAT3 signaling mediates tumour resistance to EGFR targeted therapeutics. Mol. Cell Endocrinol. 2017, 451, 15–23. [Google Scholar] [CrossRef]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Bode, A.M.; Dong, Z.; Lee, M.H. AKT as a Therapeutic Target for Cancer. Cancer Res. 2019, 79, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Trovato, M.; Torre, M.L.; Ragonese, M.; Simone, A.; Scarfì, R.; Barresi, V.; Giuffrè, G.; Benvenga, S.; Angileri, F.F.; Tuccari, G.; et al. HGF/c-met system targeting PI3K/AKT and STAT3/phosphorylated-STAT3 pathways in pituitary adenomas: An immunohistochemical characterization in view of targeted therapies. Endocrine 2013, 44, 735–743. [Google Scholar] [CrossRef]
- Liang, T.; Yue, W.; Li, Q. Chemopreventive effects of Peucedanum praeruptorum DUNN and its major constituents on SGC7901 gastric cancer cells. Molecules 2010, 15, 8060–8071. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Yang, S.; Wang, K.; Sun, S.Y. MET inhibitors for targeted therapy of EGFR TKI-resistant lung cancer. J. Hematol. Oncol. 2019, 12, 63. [Google Scholar] [CrossRef]
- Mo, H.N.; Liu, P. Targeting MET in cancer therapy. Chronic Dis. Transl. Med. 2017, 3, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, F.; Giovannetti, E.; Saso, L.; Firuzi, O. HGF/MET pathway aberrations as diagnostic, prognostic, and predictive biomarkers in human cancers. Crit. Rev. Clin. Lab. Sci. 2019, 56, 533–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, W.S.; Chinn, D.C.; Lara, P.N., Jr.; Gandara, D.R.; Mack, P.C. Effects of AKT inhibition on HGF-mediated erlotinib resistance in non-small cell lung cancer cell lines. J. Cancer Res. Clin. Oncol. 2015, 141, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.Y.; Lee, C.H.; Chiou, H.L.; Lin, C.L.; Chen, P.N.; Lin, M.T.; Hsieh, Y.H.; Chou, M.C. Praeruptorin-B Inhibits 12-O-Tetradecanoylphorbol-13-Acetate-Induced Cell Invasion by Targeting AKT/NF-kappaB via Matrix Metalloproteinase-2/-9 Expression in Human Cervical Cancer Cells. Cell. Physiol. Biochem. 2019, 52, 1255–1266. [Google Scholar] [PubMed]
- Park, H.J.; Park, S.H.; Choi, Y.H.; Chi, G.Y. The Root Extract of Scutellaria baicalensis Induces Apoptosis in EGFR TKI-Resistant Human Lung Cancer Cells by Inactivation of STAT3. Int. J. Mol. Sci. 2021, 22, 5181. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | RT 1 | MS 2 [M + H]+ (m/z) |
|---|---|---|
| PA 3 | 25.95 | 404.0 |
| EPP 4 peak | 25.94 | 404.1 |
| Name | Primer Sequence (5′→3′) | AT 1 (°C) |
|---|---|---|
| MMP-2 | Forward: CGC ATC TGG GGC TTT AAA CAT Reverse: CCA TTA GCG CCT CCA TCG TA | 60 |
| Cyclin D1 | Forward: CCT GTC CTA CTA CCG CCT CA Reverse: TCC TCC TCT TCC TCC TCC TC | 55 |
| Twist | Forward: TGT CCG CGT CCC ACT AGC Reverse: TGT CCA TTT TCT CCT TCT CTG GA | 55 |
| c-myc | Forward: CTT CTC TCC GTC CTC GGA TTC T Reverse: GAA GGT GAT CCA GAC TCT GAC CTT | 55 |
| VEGF | Forward: AGG AGG AGG GCA GAA TCA TCA Reverse: CTC GAT TGG ATG GCA GTA GCT | 55 |
| Bcl-xL | Forward: GTT CCC TTT CCT TCC ATC C Reverse: TAG CCA GTC CAG AGG TGA G | 55 |
| Actin | Forward: ACT ACC TCA TGA AGA TC Reverse: GAT CCA CAT CTG CTG GAA | 55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.-J.; Jeong, J.-H.; Park, S.-H. The Root Extract of Peucedanum praeruptorum Dunn Exerts Anticancer Effects in Human Non-Small-Cell Lung Cancer Cells with Different EGFR Mutation Statuses by Suppressing MET Activity. Molecules 2022, 27, 2360. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27072360
Park H-J, Jeong J-H, Park S-H. The Root Extract of Peucedanum praeruptorum Dunn Exerts Anticancer Effects in Human Non-Small-Cell Lung Cancer Cells with Different EGFR Mutation Statuses by Suppressing MET Activity. Molecules. 2022; 27(7):2360. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27072360
Chicago/Turabian StylePark, Hyun-Ji, Jae-Hoon Jeong, and Shin-Hyung Park. 2022. "The Root Extract of Peucedanum praeruptorum Dunn Exerts Anticancer Effects in Human Non-Small-Cell Lung Cancer Cells with Different EGFR Mutation Statuses by Suppressing MET Activity" Molecules 27, no. 7: 2360. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27072360