
Computational Strategy for Minimizing Mycotoxins in Cereal Crops: Assessment of the Biological Activity of Compounds Resulting from Virtual Screening

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
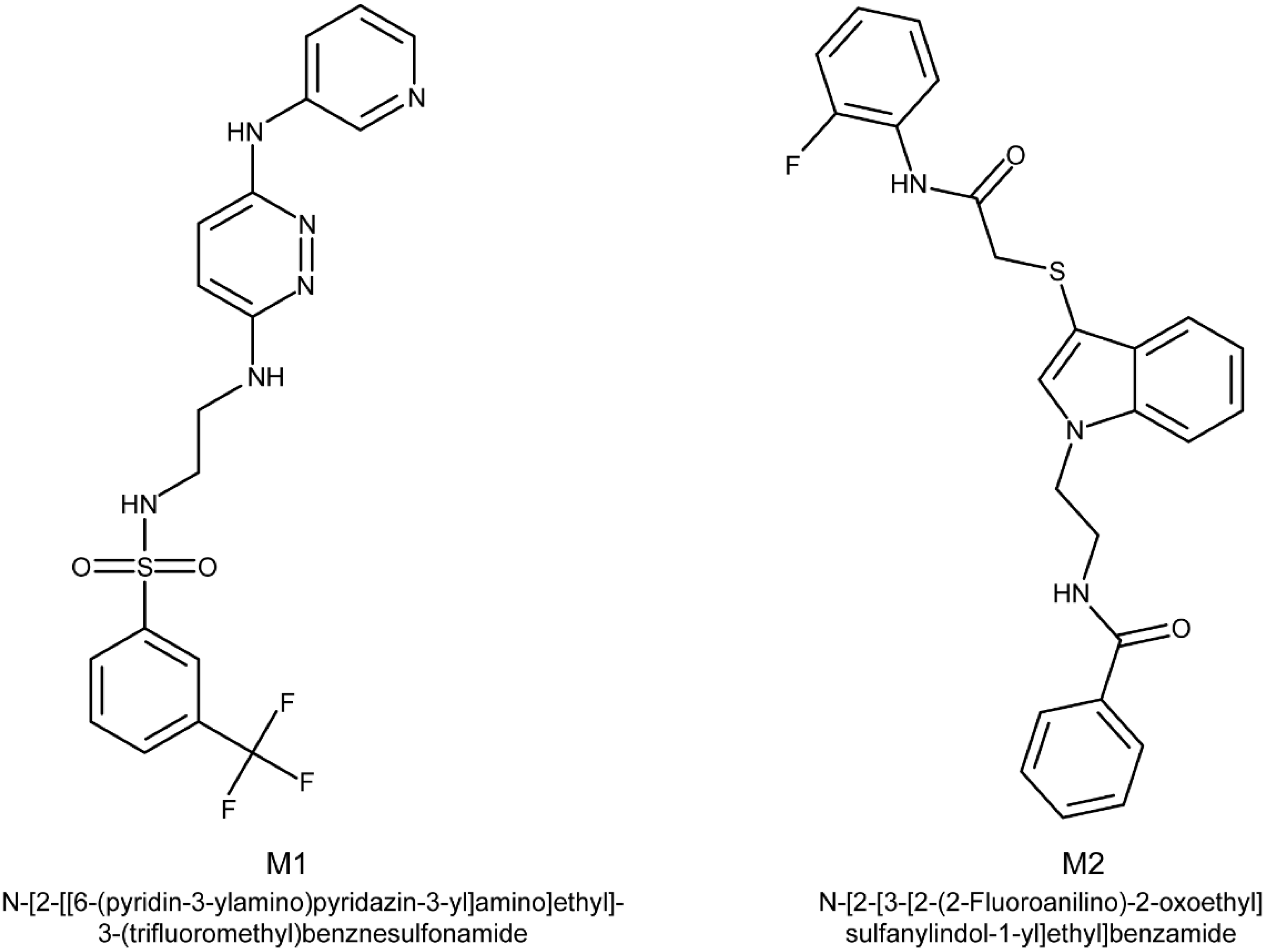
2.1. Impact of M1 and M2 on the Fungal Growth and Mycotoxin Accumulation by F. graminearum CBS 185.32
2.2. Effect of M1 on the Mycotoxin Production by Eight Fusarium Strains
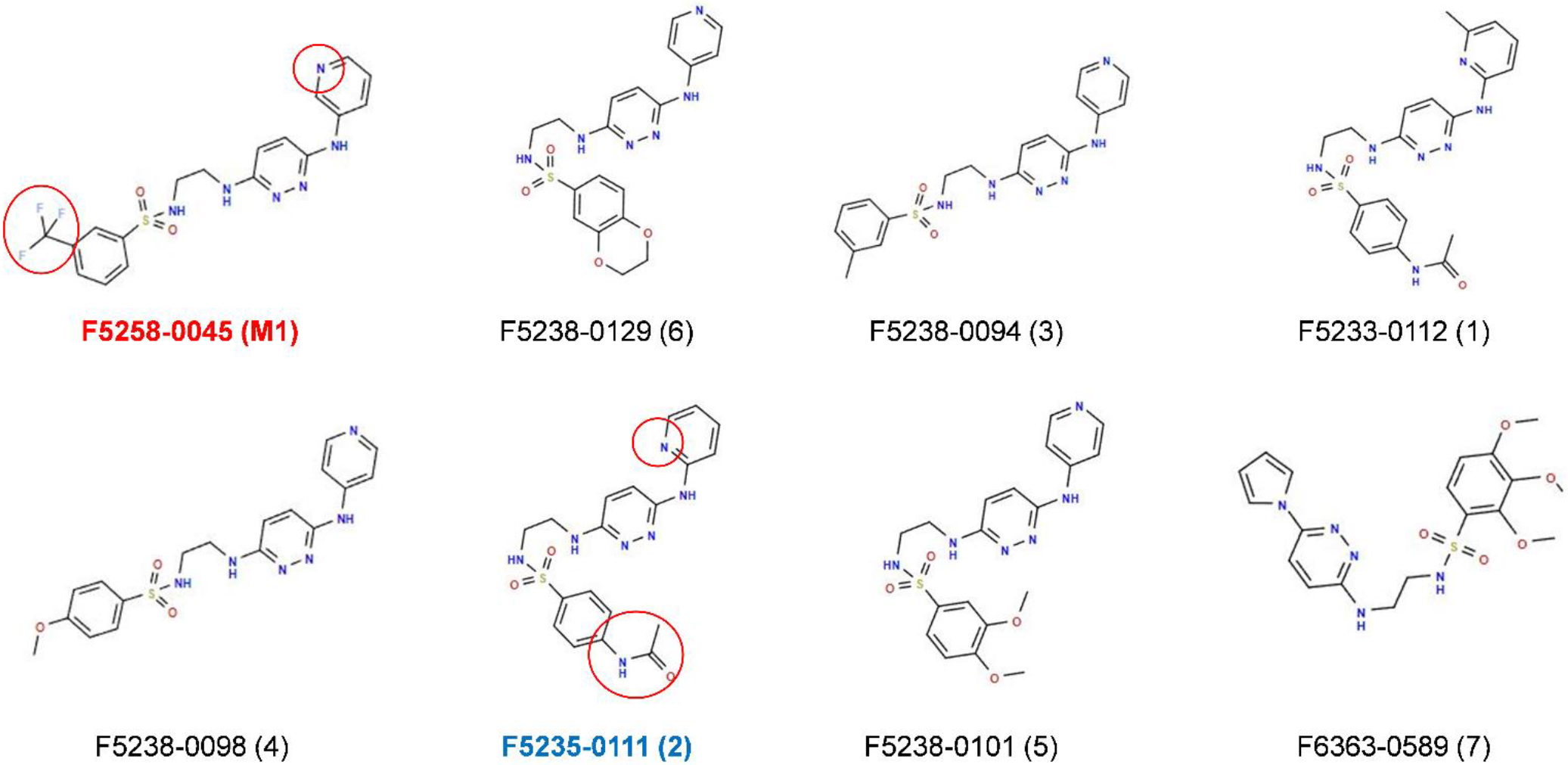
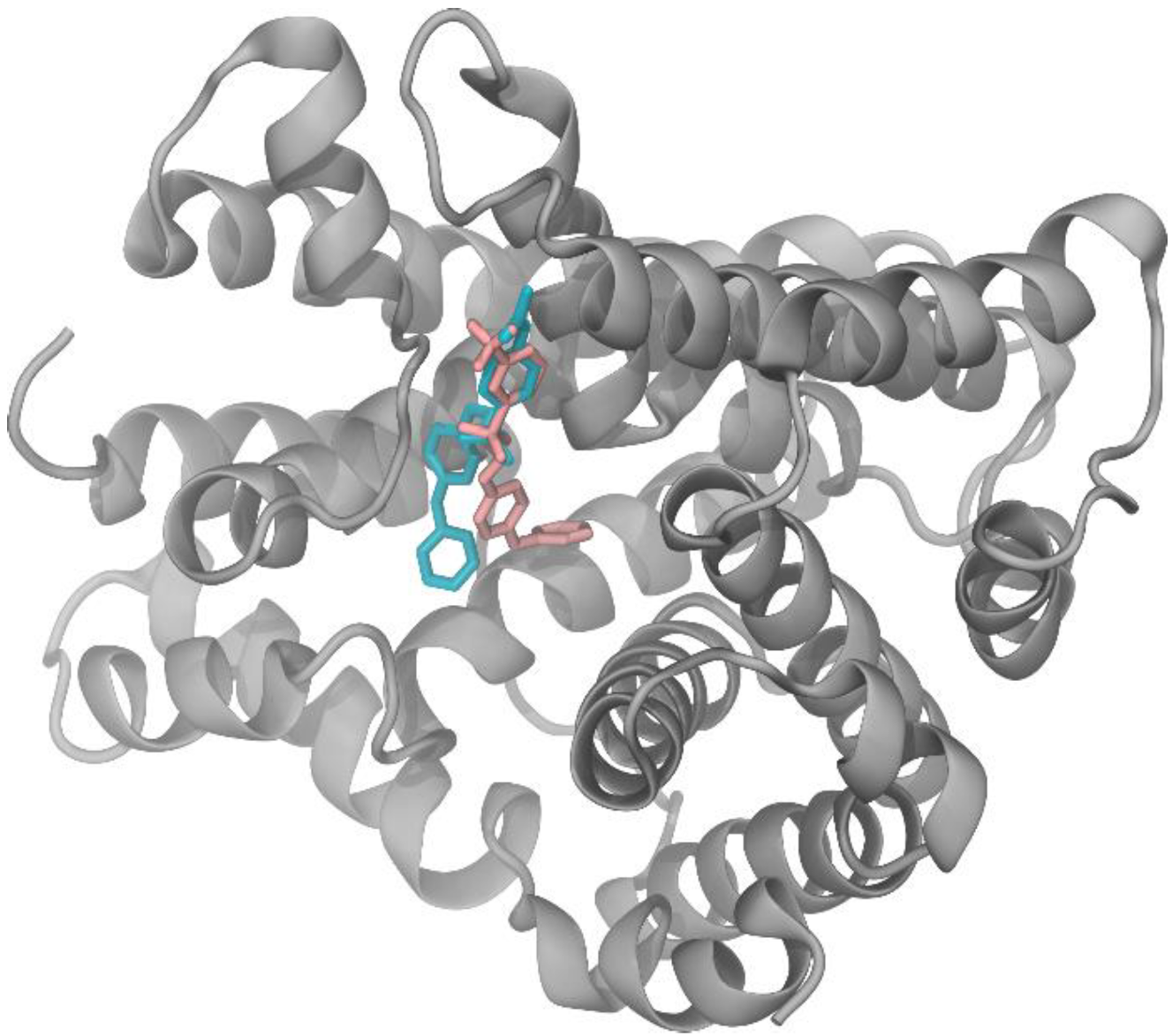
2.3. Search for M1 Analogs
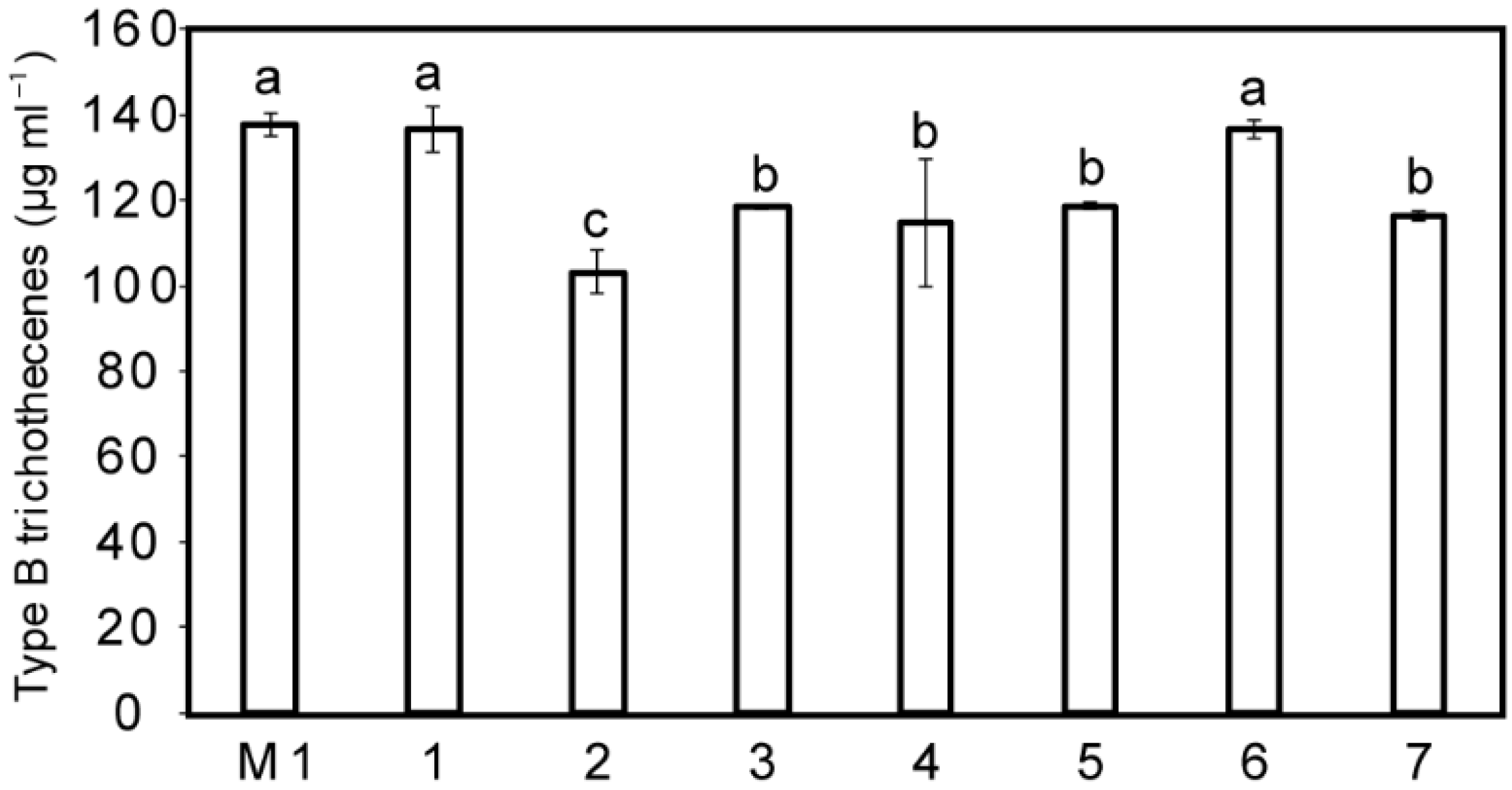
2.4. Impact of the Seven Additional Compounds on the Mycotoxin Production by F. graminearum Strain CBS 185.32
3. Discussion
4. Materials and Methods
4.1. Fusarium Strains
4.2. Medium and Culture Conditions
4.3. Extraction and TCTB Quantification
4.4. Expression of Results and Statistical Analyses
4.5. Molecular Docking Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Duba, A.; Goriewa-Duba, K.; Wachowska, U. A review of the interactions between wheat and wheat pathogens: Zymoseptoria Tritici, Fusarium Spp. and Parastagonospora Nodorum. Int. J. Mol. Sci. 2018, 19, 1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancova, K.; Hajslova, J.; Kostelanska, M.; Kohoutkova, J.; Nedelnik, J.; Moravcova, H.; Vanova, M. Fate of trichothecene mycotoxins during the processing: Milling and baking. Food Addit. Contam. Part A 2008, 25, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Magan, N. Mycotoxin contamination of food in Europe: Early detection and prevention strategies. Mycopathologia 2006, 162, 245. [Google Scholar] [CrossRef] [PubMed]
- Magan, N.; Aldred, D. Post-harvest control strategies: Minimizing mycotoxins in the food chain. Int. J. Food Microbiol. 2007, 119, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergamini, E.; Catellani, D.; Dall’asta, C.; Galaverna, G.; Dossena, A.; Marchelli, R.; Suman, M. Fate of Fusarium mycotoxins in the cereal product supply cChain: The deoxynivalenol (DON) case within industrial bread-making technology. Food Addit. Contam. Part A 2010, 27, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef]
- Rocha, O.; Ansari, K.; Doohan, F.M. Effects of trichothecene mycotoxins on eukaryotic cells: A review. Food Addit. Contam. 2005, 22, 369–378. [Google Scholar] [CrossRef]
- Hou, S.; Ma, J.; Cheng, Y.; Wang, H.; Sun, J.; Yan, Y. The toxicity mechanisms of DON to humans and animals and potential biological treatment strategies. Crit. Rev. Food Sci. Nutr. 2021, 14, 1–23. [Google Scholar] [CrossRef]
- Hazel, C.M.; Patel, S. Influence of processing on trichothecene levels. Toxicol. Lett. 2004, 153, 51–59. [Google Scholar] [CrossRef]
- Paul, P.A.; Salgado, J.D.; Bergstrom, G.; Bradley, C.A.; Byamukama, E.; Byrne, A.M.; Chapara, V.; Cummings, J.A.; Chilvers, M.I.; Dill-Macky, R.; et al. Integrated effects of genetic resistance and prothioconazole + tebuconazole application timing on Fusarium Head Blight in wheat. Plant Dis. 2019, 103, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, G.; Jeon, J. Computer-aided drug discovery in plant pathology. Plant Pathol. J. 2017, 33, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.F.; Bresso, E.; Togawa, R.C.; Urban, M.; Antoniw, J.; Maigret, B.; Hammond-Kosack, K. Searching for novel targets to control wheat head blight disease―I-protein identification, 3D modeling and virtual screening. Adv. Microbiol. 2016, 6, 811–830. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Esquivel, B.D.; White, T.C. Overexpression or deletion of ergosterol biosynthesis genes alters doubling time, response to stress agents, and drug susceptibility in Saccharomyces Cerevisiae. mBio 2018, 9, e01291-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willett, P. Similarity methods in chemoinformatics. Annu. Rev. Inf. Sci. Technol. 2009, 43, 1–117. [Google Scholar] [CrossRef] [Green Version]
- Pani, G.; Scherm, B.; Azara, E.; Balmas, V.; Jahanshiri, Z.; Carta, P.; Fabbri, D.; Dettori, M.A.; Fadda, A.; Dessi, A.; et al. Natural and natural-like phenolic inhibitors of type B trichothecene in vitro production by the wheat (Triticum Sp.) pathogen Fusarium culmorum. J. Agric. Food Chem. 2014, 62, 4969–4978. [Google Scholar] [CrossRef]
- Tetko, I.V.; Tanchuk, V.Y. Application of associative neural networks for prediction of lipophilicity in ALOGPS 2.1 Program. J. Chem. Inf. Comput. Sci. 2002, 42, 1136–1145. [Google Scholar] [CrossRef]
- Banerjee, P.; Eckert, A.O.; Schrey, A.K.; Preissner, R. ProTox-II: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef] [Green Version]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein-ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand–protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Audenaert, K.; Vanheule, A.; Höfte, M.; Haesaert, G. Deoxynivalenol: A major player in the multifaceted response of Fusarium to its environment. Toxins 2014, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, G.H.; Desjardins, A.E.; Plattner, R.D. Deoxynivalenol-nonproducing Fusarium graminearum causes initial infection, but does not cause disease spread in wheat spikes. Mycopathologia 2002, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Obst, A.; Lepschy, J.; Beck, R.; Bauer, G.; Bechtel, A. The risk of toxins by Fusarium graminearum in wheat—Interactions between weather and agronomic factors. Mycotoxin Res. 2000, 16, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Ponts, N. Mycotoxins are a component of Fusarium graminearum stress-response system. Front. Microbiol. 2015, 6, 1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talas, F.; Würschum, T.; Reif, J.C.; Parzies, H.K.; Miedaner, T. Association of single nucleotide polymorphic sites in candidate genes with aggressiveness and deoxynivalenol production in Fusarium graminearum causing wheat head blight. BMC Genet. 2012, 13, 14. [Google Scholar] [CrossRef] [Green Version]
- Vanheule, A.; De Boevre, M.; Moretti, A.; Scauflaire, J.; Munaut, F.; De Saeger, S.; Bekaert, B.; Haesaert, G.; Waalwijk, C.; Van der Lee, T.; et al. Genetic divergence and chemotype diversity in the Fusarium head blight pathogen Fusarium poae. Toxins 2017, 9, 255. [Google Scholar] [CrossRef]
- Li, M.; Goncearenco, A.; Panchenko, A.R. Annotating mutational effects on proteins and protein interactions: Designing novel and revisiting existing protocols. In Proteomics: Methods in Molecular Biology; Comai, L., Katz, J., Mallick, P., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1550, pp. 235–260. [Google Scholar] [CrossRef] [Green Version]
- Boutigny, A.-L.; Barreau, C.; Atanasova-Penichon, V.; Verdal-Bonnin, M.-N.; Pinson-Gadais, L.; Richard-Forget, F. Ferulic Acid, an efficient inhibitor of type B trichothecene biosynthesis and Tri gene expression in Fusarium liquid cultures. Mycol. Res. 2009, 113, 746–753. [Google Scholar] [CrossRef]
- Atanasova-Penichon, V.; Legoahec, L.; Bernillon, S.; Deborde, C.; Maucourt, M.; Verdal-Bonnin, M.-N.; Pinson-Gadais, L.; Ponts, N.; Moing, A.; Richard-Forget, F. Mycotoxin biosynthesis and central metabolism are two interlinked pathways in Fusarium graminearum, as demonstrated by the extensive metabolic changes induced by caffeic acid exposure. Appl. Environ. Microbiol. 2018, 84, e01705-17. [Google Scholar] [CrossRef] [Green Version]
- Gautier, C.; Pinson-Gadais, L.; Verdal-Bonnin, M.-N.; Ducos, C.; Tremblay, J.; Chéreau, S.; Atanasova, V.; Richard-Forget, F. Investigating the efficiency of hydroxycinnamic acids to inhibit the production of enniatins by Fusarium avenaceum and modulate the expression of enniatins biosynthetic genes. Toxins 2020, 12, 735. [Google Scholar] [CrossRef]
- Rynkiewicz, M.J.; Cane, D.E.; Christianson, D.W. Structure of trichodiene synthase from Fusarium sporotrichioides provides mechanistic inferences on the terpene cyclization cascade. Proc. Natl. Acad. Sci. USA 2001, 98, 13543–13548. [Google Scholar] [CrossRef] [Green Version]
- Vedula, L.S.; Rynkiewicz, M.J.; Pyun, H.-J.; Coates, R.M.; Cane, D.E.; Christianson, D.W. Molecular recognition of the substrate diphosphate group governs product diversity in trichodiene synthase mutants. Biochemistry 2005, 44, 6153–6163. [Google Scholar] [CrossRef] [PubMed]
- Gaber, R.F.; Copple, D.M.; Kennedy, B.K.; Vidal, M.; Bard, M. The yeast gene ERG6 is required for normal membrane function but is not essential for biosynthesis of the cell-cycle-sparking sterol. Mol. Cell. Biol. 1989, 9, 3447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konecna, A.; Toth Hervay, N.; Valachovic, M.; Gbelska, Y. ERG6 gene deletion modifies Kluyveromyces lactis susceptibility to various growth inhibitors. Yeast 2016, 33, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Leal, P.C.; Mascarello, A.; Derita, M.; Zuljan, F.; Nunes, R.J.; Zacchino, S.; Yunes, R.A. Relation between lipophilicity of alkyl gallates and antifungal activity against yeasts and filamentous fungi. Bioorg. Med. Chem. Lett. 2009, 19, 1793–1796. [Google Scholar] [CrossRef]
- Nicholson, P.; Simpson, D.R.; Weston, G.; Rezanoor, H.N.; Lees, A.K.; Parry, D.W.; Joyce, D. Detection and quantification Of Fusarium culmorum and Fusarium graminearum in cereals using PCR assays. Physiol. Mol. Plant Pathol. 1998, 53, 17–37. [Google Scholar] [CrossRef]
- Bakan, B.; Pinson, L.; Cahagnier, B.; Melcion, D.; Semon, E.; Richard-Molard, D. Toxigenic potential of Fusarium culmorum strains isolated from French wheat. Food Addit. Contam. 2001, 18, 998–1003. [Google Scholar] [CrossRef]
- Montibus, M.; Vitrac, X.; Coma, V.; Loron, A.; Pinson-Gadais, L.; Ferrer, N.; Verdal-Bonnin, M.-N.; Gabaston, J.; Waffo-Téguo, P.; Richard-Forget, F.; et al. Screening of wood/forest and vine by-products as sources of new drugs for sustainable strategies to control Fusarium graminearum and the production of mycotoxins. Molecules 2021, 26, 405. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Chemotype | Source | Sum of Type B Trichothecenes (µg g−1 of Dry Biomass) | |
|---|---|---|---|---|
| Control | M1 | |||
| F. graminearum | ||||
| CBS 185.32 | DON/15-ADON | CBS Collection, The Netherlands | 38,379.2 ± 1059.8 | 24,391 ± 1314.2 * |
| PH-1 | DON/15-ADON | The Fungal Genetics Stock Center, USA | 76.8 ± 13.3 | <LOQ * |
| F. culmorum | ||||
| Fc 130 | NIV/FX | INRAE, MycSA, France | 157.8 ± 7.6 | 107.3± 8.3 * |
| Fc 337 | NIV/FX | INRAE, MycSA, France | 166.1 ± 18.5 | 126.2 ± 5.5 * |
| Fc 319 | NIV/FX | INRAE, MycSA, France | 7617.3 ± 1026.6 | 6565.7 ± 483.6 |
| MCf 21 | DON/3-ADON | University of Sassari, Italy [16] | 185.5 ± 68.6 | 148.4 ± 16.8 |
| Fc 124 | DON/3-ADON | INRAE, MycSA, France | 304.9 ± 29.3 | 148.7 ± 24.2 * |
| Fc 305 | DON/3-ADON | INRAE, MycSA, France | 5994.1 ± 39.1 | 6465.7 ± 547.7 |
| Compound | Label | Chemical Formula | GOLD Score | GOLD Rank | Estimated Lipophilicity (LogP) |
|---|---|---|---|---|---|
| F5258-0045 | M1 | C18H17F3N6O2S | 80.5 | 1 | 4.30 |
| F5238-0129 | 6 | C19H20N6O4S | 74.1 | 3 | 3.13 |
| F5238-0094 | 3 | C18H20N6O2S | 73.9 | 4 | 3.39 |
| F5233-0112 | 1 | C20H23N7O3S | 73.3 | 7 | 2.72 |
| F5238-0098 | 4 | C18H20N6O3S | 71.9 | 9 | 3.16 |
| F5235-0111 | 2 | C19H21N7O3S | 69.8 | 19 | 2.22 |
| F5238-0101 | 5 | C19H22N6O4S | 68.4 | 27 | 2.86 |
| F6363-0589 | 7 | C19H23N5O5S | 67.4 | 34 | 1.95 |
| Residue N° | Type of Interaction | M1 | Analog 2 |
|---|---|---|---|
| Glu164 | polar | + | + |
| Leu181 | hydrophobic | + | + |
| Arg182 | H-bond | − | + |
| Asn185 | H-bond | + | + |
| Asn225 | H-bond | + | + |
| Ser229 | H-bond | − | + |
| Asp239 | H-bond polar | + − | − + |
| Gln240 | H-bond | + | + |
| Ile241 | H-bond hydrophobic | + + | + + |
| Tyr295 | H-bond | + | − |
| Trp298 | π stacking | + | − |
| Tyr305 | H-bond π stacking | + − | − + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atanasova, V.; Bresso, E.; Maigret, B.; Martins, N.F.; Richard-Forget, F. Computational Strategy for Minimizing Mycotoxins in Cereal Crops: Assessment of the Biological Activity of Compounds Resulting from Virtual Screening. Molecules 2022, 27, 2582. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27082582
Atanasova V, Bresso E, Maigret B, Martins NF, Richard-Forget F. Computational Strategy for Minimizing Mycotoxins in Cereal Crops: Assessment of the Biological Activity of Compounds Resulting from Virtual Screening. Molecules. 2022; 27(8):2582. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27082582
Chicago/Turabian StyleAtanasova, Vessela, Emmanuel Bresso, Bernard Maigret, Natalia Florencio Martins, and Florence Richard-Forget. 2022. "Computational Strategy for Minimizing Mycotoxins in Cereal Crops: Assessment of the Biological Activity of Compounds Resulting from Virtual Screening" Molecules 27, no. 8: 2582. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27082582