
Excretion, Metabolism, and Tissue Distribution of Gelsemium elegans (Gardn. & Champ.) Benth in Pigs


Abstract
:1. Introduction
2. Results
2.1. Characterization of Compounds in Plasma
2.2. Characterization of Compounds in Urine
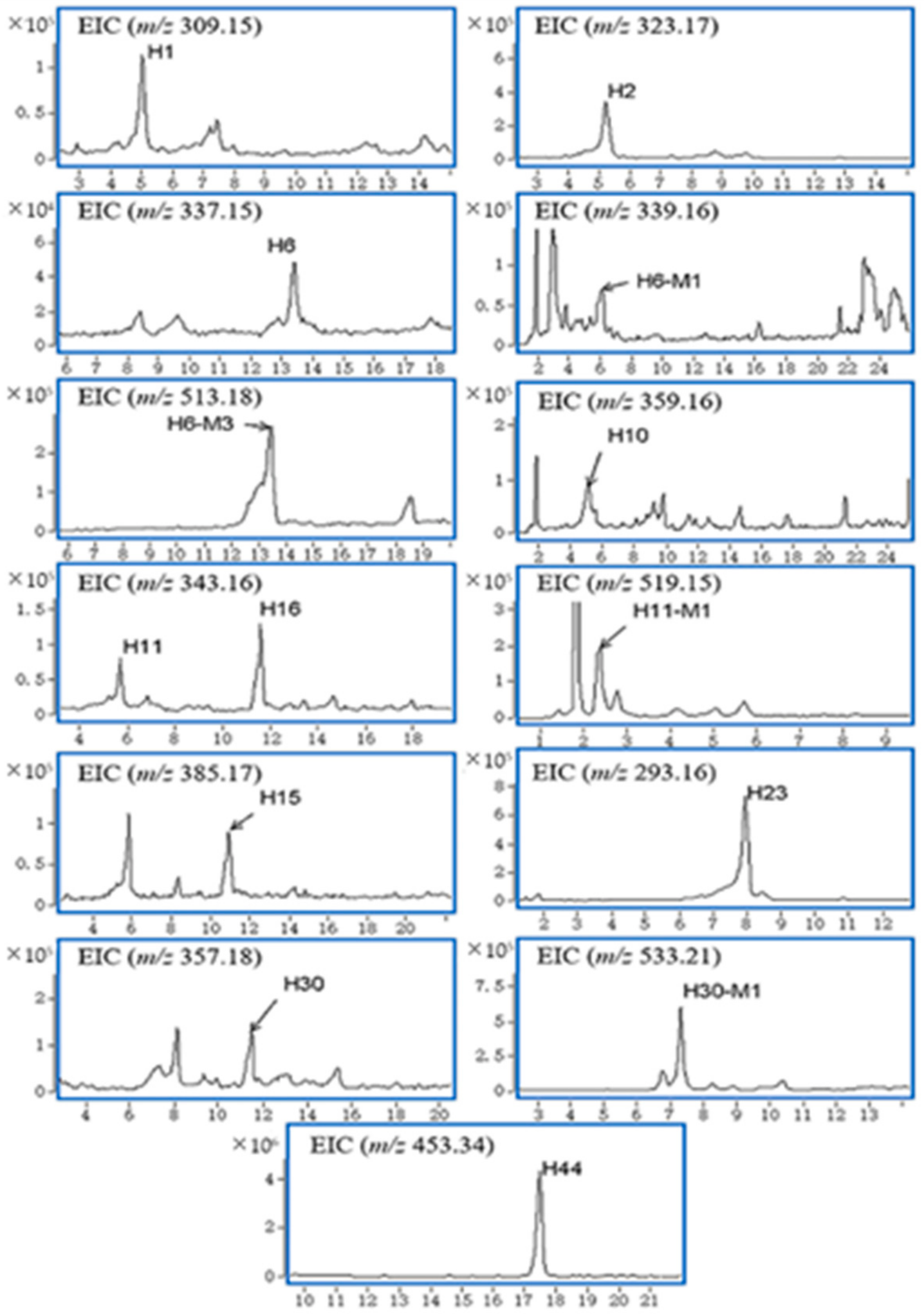
2.2.1. Gelsemine-Type Alkaloids (H6) and Its Metabolic Products
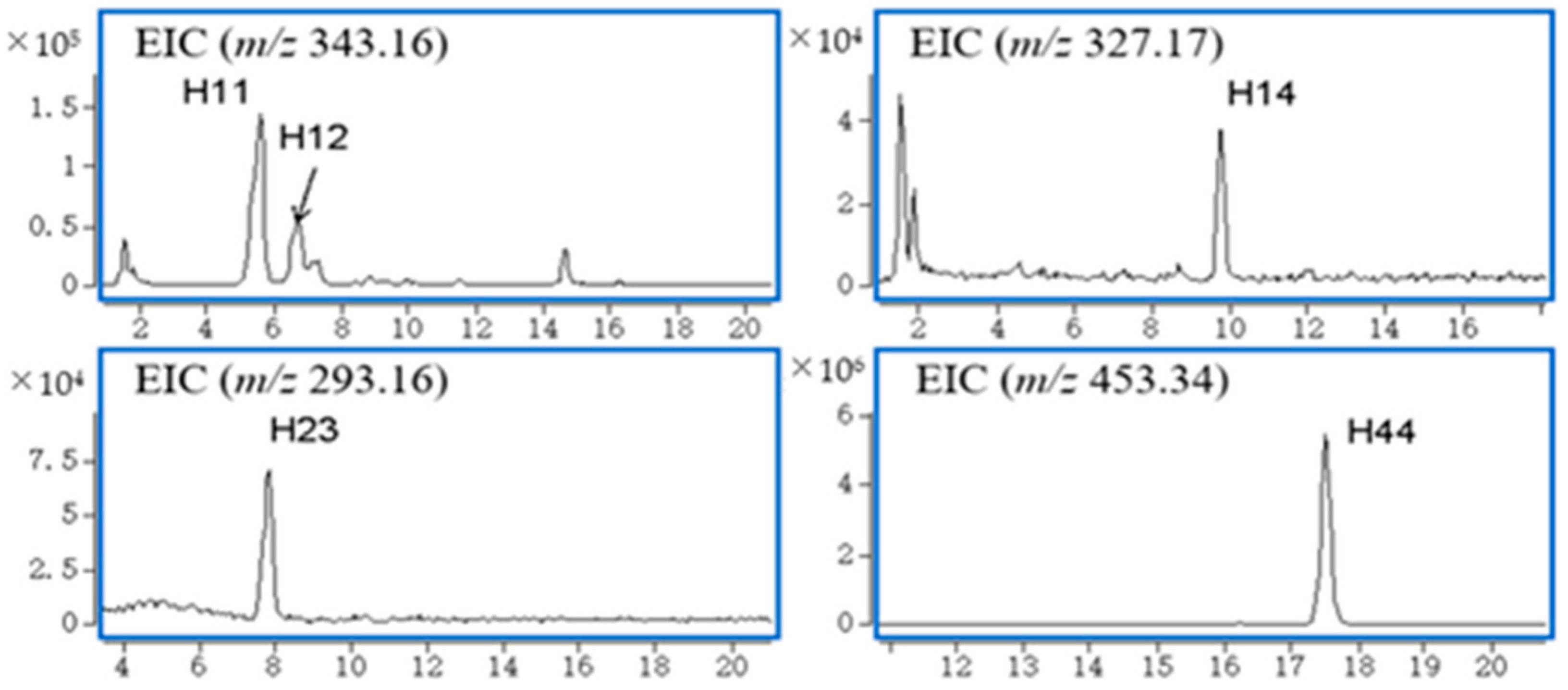
2.2.2. Gelsedine-Type Alkaloids (H10, H11, H15, H16, H18) and Their Metabolic Products
2.2.3. Sarpagine-Type Alkaloids (H23)
2.2.4. Humantenine-Type Alkaloids (H30) and Its Metabolic Products
2.2.5. Non-Alkaloids (H37, H39, H44)
2.3. Characterization of Compounds in Bile
2.4. Compounds Identification in Feces
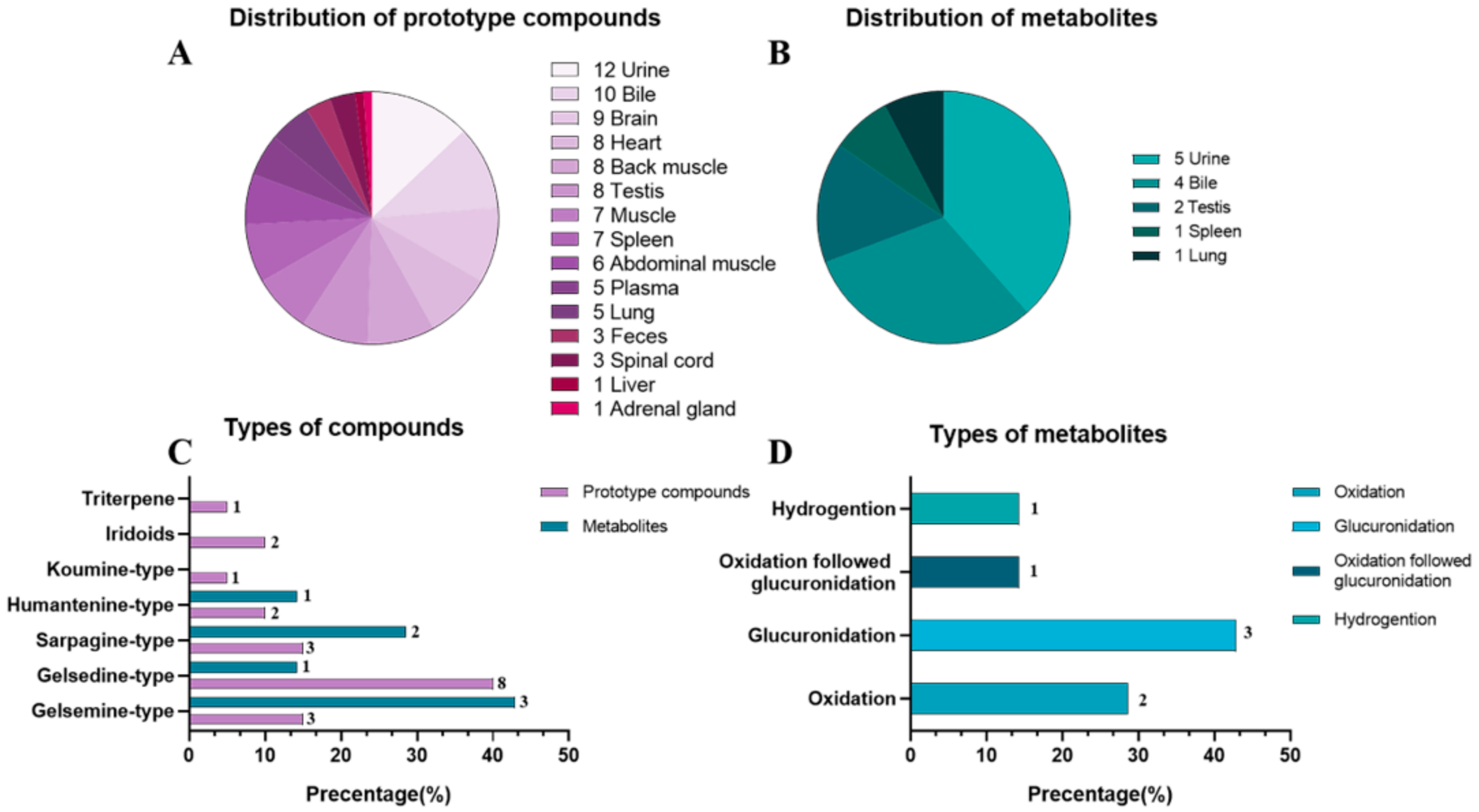
2.5. Characterization of Compounds in Tissue
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plants Material
4.3. Animal Experiments
4.3.1. Plasma Samples
4.3.2. Urine Samples
4.3.3. Feces Samples
4.3.4. Bile Samples
4.3.5. Tissue Samples
4.3.6. HPLC/QqTOF-MS Analysis Conditions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Jin, G.L.; Su, Y.P.; Liu, M.; Xu, Y.; Yang, J.; Liao, K.J.; Yu, C.X. Medicinal plants of the genus Gelsemium (Gelsemiaceae, Gentianales)—A review of their phytochemistry, pharmacology, toxicology and traditional use. J. Ethnopharmacol. 2014, 152, 33–52. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.K.; Yang, L.; Liao, S.G.; Cao, P.; Wu, B.; Hu, H.B.; Guo, J.; Zhang, P. Koumine, Humantenine, and Yohimbane Alkaloids from Gelsemium elegans. J. Nat. Prod. 2015, 78, 1511–1517. [Google Scholar] [CrossRef] [PubMed]
- Paris, A.; Schmidlin, S.; Mouret, S.; Hodaj, E.; Marijnen, P.; Boujedaini, N.; Polosan, M.; Cracowski, J.L. Effect of Gelsemium 5CH and 15CH on anticipatory anxiety: A phase III, single-centre, randomized, placebo-controlled study. Fundam. Clin. Pharmacol. 2012, 26, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Pascarella, J.B. Mechanisms of prezygotic reproductive isolation between two sympatric species, Gelsemium rankinji and G. sempervirens (gelsemiaceae), in the southeastern united states. Am. J. Bot. 2007, 94, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.J.; Han, X.F. Mechanism, clinical characteristics and management of Gelsemium elegans poisoning. Advers. Drug React. J. 2006, 8, 202–204. [Google Scholar]
- Wang, J.; Sun, Z.L. Effect and mechanism of feeding hook-kiss dry powder on growth performance of piglets. Chin. J. Tradit. Vet. Sci. 2019, 2, 3–4. [Google Scholar]
- Liu, M.; Huang, H.H.; Yang, J.; Su, Y.P.; Lin, H.W.; Lin, L.Q.; Liao, W.J.; Yu, C.X. The active alkaloids of Gelsemium elegans Benth. are potent anxiolytics. Psychopharmacology 2013, 225, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Xiao, S.; Yang, K.; Ling, L.; Sun, Z.L.; Liu, Z.-Y. Comprehensive identification and structural characterization of target components from Gelsemium elegans by high-performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry based on accurate mass databases combined with MS/MS spectra. J. Mass Spectrom. 2017, 52, 378–396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, S.Y.; Wang, G.Y.; Li, N.P.; Chen, M.F.; Gu, J.H.; Zhang, D.M.; Wang, L.; Ye, W.C. Five new koumine-type alkaloids from the roots of Gelsemium elegans. Fitoterapia 2017, 118, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Zuo, M.T.; Liu, Z.Y. The Metabolism and Disposition of Koumine, Gelsemine and Humantenmine from Gelsemium. Curr. Drug Metab. 2019, 20, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.J.; Chen, M.H.; Hu, Y.X.; Lan, Y.; Gan, L.L.; You, G.Q.; Yue, M.; Wang, H.M.; Xia, B.J.; Zhao, J.; et al. CYP3A4/5 mediates the metabolic detoxification of humantenmine, a highly toxic alkaloid from Gelsemium elegans Benth. J. Appl. Toxicol. 2019, 39, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Long, X.M.; Cao, J.J.; Li, Y.J.; Wu, Y.; Bai, X.; Sun, Z.L.; Liu, Z.Y. An analytical strategy to explore the multicomponent pharmacokinetics of herbal medicine independently of standards: Application in Gelsemium elegans extracts. J. Pharm. Biomed. Anal. 2019, 176, 112833. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sun, Q.; Zhao, N.; Wen, Y.Q.; Song, Y.; Meng, F.H. Ultra-Liquid Chromatography Tandem Mass Spectrometry (UPLC-MS/MS)-Based Pharmacokinetics and Tissue Distribution Study of Koumine and the Detoxification Mechanism of Glycyrrhiza uralensis Fisch on Gelsemium elegans Benth. Molecules 2018, 23, 1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, M.T.; Wang, Z.Y.; Yang, K.; Li, Y.J.; Huang, C.Y.; Liu, Y.C.; Yu, H.; Zhao, X.J.; Liu, Z.Y. Characterization of absorbed and produced constituents in goat plasma urine and faeces from the herbal medicine Gelsemium elegans by using high-performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. J. Ethnopharmacol. 2020, 252, 112617. [Google Scholar] [CrossRef] [PubMed]
- Yang, K. Study on Pharmacokinetics and Elimination Rules of Gelsemium Alkaloids in Pigs; Hunan Agricultural University: Changsha, China, 2019. [Google Scholar]
- Yang, K.; Long, X.M.; Liu, Y.C.; Chen, F.H.; Liu, X.F.; Sun, Z.L.; Liu, Z.Y. Development and in-house validation of a sensitive LC–MS/MS method for simultaneous quantification of gelsemine, koumine and humantenmine in porcine plasma. J. Chromatogr. B 2018, 1076, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Huang, Y.J.; Liu, Y.C.; Sun, Z.L.; Liu, Z.Y. In vitro metabolism of koumine in pig. Chin. J. Vet. Sci. 2018, 38, 1568–1572. [Google Scholar] [CrossRef]
- Yang, K.; Huang, Y.J.; Xiao, S.; Liu, Y.C.; Sun, Z.L.; Liu, Y.S.; Tang, Q.; Liu, Z.Y. Identification of gelsemine metabolites in rat liver S9 by high-performance liquid chromatography/quadrupole-time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2017, 32, 19–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.Y.; Yang, K.; Cao, J.J.; Wang, Z.Y.; Wu, Y.; Sun, Z.L.; Liu, Z.Y. Integration of metabolomics and transcriptomics to comprehensively evaluate the metabolic effects of Gelsemium elegans on pigs. Animals 2021, 11, 1192. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
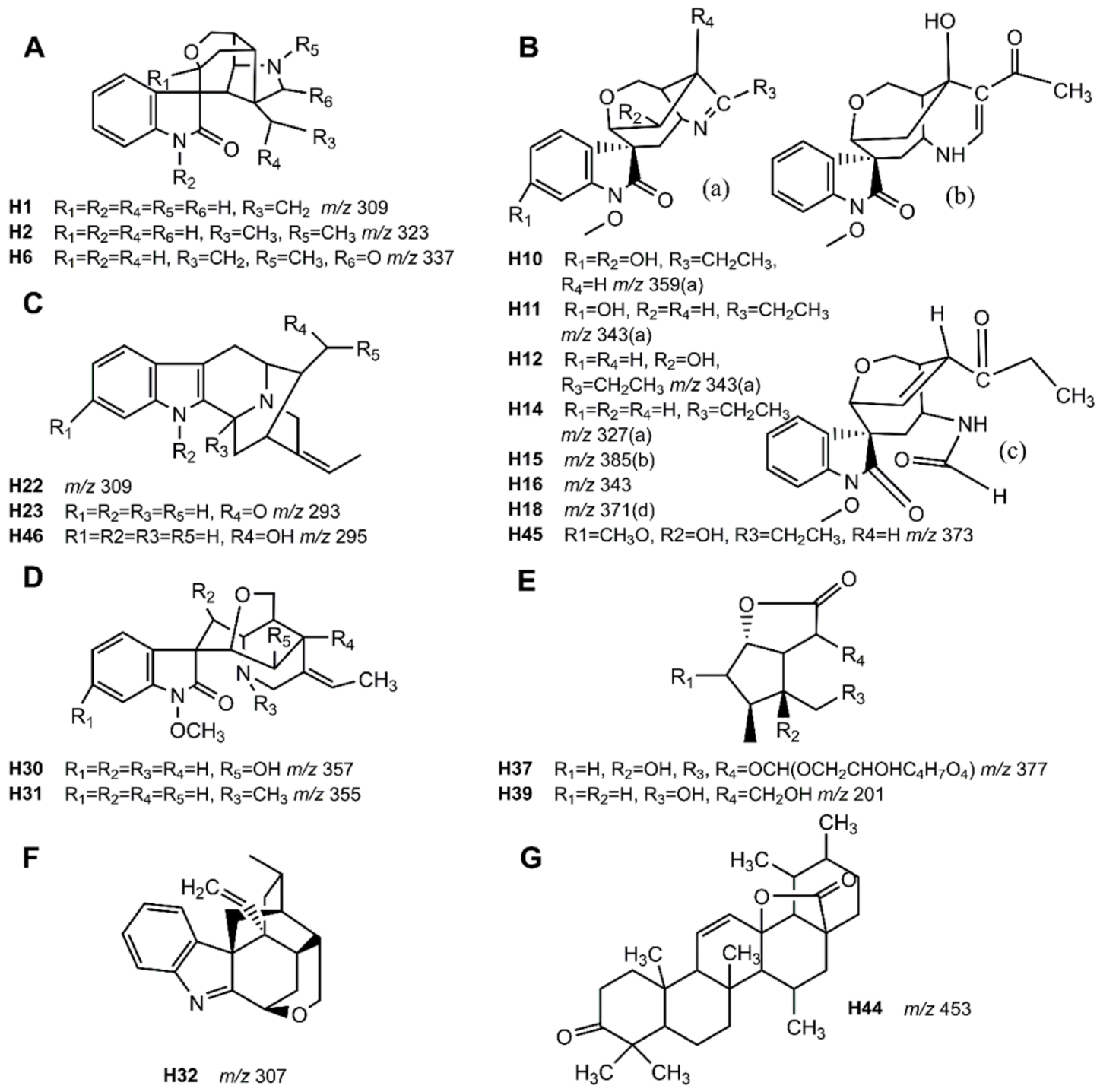
| Compound | RT (min) | Molecular Formula | [M + H]+ (m/z) | Error (ppm) | Fragment Ions (m/z) | Compound Class | Name | Proposed Metabolism | Source |
|---|---|---|---|---|---|---|---|---|---|
| H1 | 5.0 | C19H20N2O2 | 309.1579 | 6.02 | 236.1023, 191.0609, 91.0533, 56.0469 | Alkaloid, Gelsemine-type | Unknown | B | |
| H2 | 5.2 | C20H22N2O2 | 323.1750 | 1.26 | 236.1076, 115.0551, 77.0367, 70.0641 | Alkaloid, Gelsemine-type | Gelsemine | B | |
| H6 | 13.4 | C20H20N2O3 | 337.1565 | −5.45 | 236.1052, 91.0501 | Alkaloid, Gelsemine-type | 21-Oxogelsemine | B, U | |
| H6-M1 | 6.0 | C20H22N2O3 | 339.1667 | 10.7 | 206.0973, 77.0389 | +2H | B | ||
| H6-M2 | 9.8 | C20H20N2O4 | 353.1500 | −1.18 | 218.0893, 180.0933, 91.0536, 77.0355 | +O | U | ||
| H6-M3 | 13.4 | C26H28N2O9 | 513.1845 | 4.41 | 337.1425, 91.0490, 77.0335 | +GlcA | B, U | ||
| H10 | 5.4 | C19H22N2O5 | 359.1604 | −0.7 | 311.11170, 80.0467, 68.0465 | Alkaloid, Gelsedine-type | 11,14-Dihydroxygelsenicine | (B), U | |
| H11 | 5.6 | C19H22N2O4 | 343.1634 | 5.36 | 312.1356, 281.1184, 108.0689, 96.0817, 80.0452 | Alkaloid, Gelsedine-type | 11-Hydroxygelsenicine | B, U, P | |
| H11-M1 | 2.4 | C25H30N2O10 | 519.1978 | −0.92 | 312.1504, 281.1258, 108.0839, 80.0476 | +GlcA | B, U | ||
| H12 | 6.6 | C19H22N2O4 | 343.1640 | 3.61 | 312.1428, 80.0491 | Alkaloid, Gelsedine-type | 14-Hydroxygelsenicine | P | |
| H14 | 9.7 | C19H22N2O3 | 327.1703 | 0.06 | 265.1240, 108.0703, 95.0869 | Alkaloid, Gelsedine-type | Gelsenicine | P | |
| H15 | 10.9 | C21H24N2O5 | 385.1758 | 0 | 354.1537, 309.1507, 180.0876, 134.0581 | Alkaloid, Gelsedine-type | 15-Hydroxyhumantenoxenine | B, U | |
| H16 | 11.6 | C19H22N2O4 | 343.1631 | 6.24 | 238.0790, 128.0493, 118.0639 | Alkaloid, Gelsedine-type | Hydroxyl of gelsenicine | B, U | |
| H18 | 11.4 | C20H22N2O5 | 371.1571 | 8.24 | 340.1368, 295.1354, 120.0412, 91.0508 | Alkaloid, Gelsedine-type | Gelsemolenine B | U | |
| H22 | 7.6 | C19H20N2O2 | 309.1551 | 11.53 | 194.0904, 167.0698, 115.0490 | Alkaloid, Sarpagine-type | Oxokoumidine | U | |
| H23 | 7.9 | C19H20N2O | 293.1650 | −0.55 | 218.0904, 194.0917, 167.0721, 115.0486, 91.0548 | Alkaloid, Sarpagine-type | Dehydrokoumidine | B, U, F, P | |
| H23-M1 | 6.3 | C25H28N2O8 | 485.1920 | −0.33 | 309.1506, 194.0918, 167.0697, 154.0607, 115.0543 | +O + GlcA | U | ||
| H23-M2 | 6.5 | C19H20N2O2 | 309.1586 | 3.75 | 291.1457, 234.1227, 194.0946, 193.0890 | +O | Only detected in tissue samples | ||
| H30 | 11.5 | C20H24N2O4 | 357.1804 | 1.36 | 311.1188, 178.1097, 108.0794, 77.0377 | Alkaloid, Humantenine-type | 14-Hydroxyrankinidine | B, U, F | |
| H30-M1 | 7.3 | C26H32N2O10 | 533.2136 | −1.18 | 326.1654, 311.1506, 164.1064, 148.0401, | +GlcA | B, U | ||
| H31 | 13.6 | C21H26N2O3 | 355.2006 | 2.88 | 309.1609, 122.0968 | Alkaloid, Humantenine-type | Humantennine | Only detected in tissue samples | |
| H32 | 7.2 | C20H22N2O | 307.1786 | 6.17 | 277.1684, 220.1106, 238.1197, 130.0643, 70.0643 | Alkaloid, Koumine-type | Koumine | Only detected in tissue samples | |
| H37 | 4.7 | C16H24O10 | 377.1422 | 5.38 | 165.0797, 119.0854, 105.0678, 93.0710 | Iridoids | 9-Hydroxysemperoside | U | |
| H39 | 4.6 | C10H16O4 | 201.1119 | 1.18 | 119.0859, 103.0541, 91.0538, 77.0391 | Iridoids | Gelsemiol | U | |
| H44 | 17.5 | C30H44O3 | 453.3410 | −10.78 | 209.1656, 114.0907, 96.0787, 69.0680 | Triterpene | 3-keto-urs-11-en-13β(28)-olide | B, U, P, F | |
| H45 | 17.7 | C20H24N2O5 | 373.1758 | −2.15 | 260.0920, 214.0844, 130.1252 | Alkaloid, Gelsedine-type | GS-2(11-Methoxy-14-Hydroxygelsenicine) | Only detected in tissue samples | |
| H46 | 8.3 | C19H22N2O | 295.1801 | 1.33 | 277.1667, 156.0797, 144.0781, 138.0900, 108.0778 | Alkaloid, Sarpagine-type | Koumidine | Only detected in tissue samples |
| Tissues | Gelsemine (H2) | 11-Hydroxygelsenicine (H11) | Gelsenicine (H14) | Dehydrokoumidine (H23) | 14-Hydroxyrankinidine (H30) | Humantennine (H31) | Koumine (H32) | 3-keto-urs-11-en-13β (28)-olide (H44) | GS-2(11-Methoxy-14-Hydroxygelsenicine) (H45) | Koumidine (H46) | H23-M2 | H30-M1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Brain | 8 × 106 | 2.9 × 105 | - | 0.9 × 106 | 2.1 × 105 | 2 × 105 | 4 × 105 | 2.9 × 107 | 2.2 × 105 | 3.1 × 105 | - | - |
| Spinal cord | 3 ×105 | - | - | 1 × 106 | - | - | - | - | 2.1 × 105 | - | - | - |
| Heart | 8.2 × 105 | 8 × 105 | 2.9 × 105 | 2.9 × 106 | 6.1 × 105 | 1.1 × 105 | 2.6 × 105 | 3.9 × 107 | - | - | - | - |
| Liver | - | - | - | 2.9 × 105 | - | - | - | - | - | - | - | - |
| Lung | 2.0 × 106 | - | 1.9 × 105 | 3 × 107 | 2.9 × 107 | - | - | - | - | 5.9 × 106 | 3.1 × 105 | - |
| Spleen | 2.5 × 106 | 5.8 × 105 | 4.9 ×105 | 6 × 106 | 1.5 × 106 | - | 3.2 × 105 | - | - | 1 × 105 | 7 × 104 | - |
| Adrenal gland | - | - | - | 0.7 × 106 | - | - | - | - | - | - | - | - |
| Testis | 6.1 × 105 | 1.3 × 106 | 2 × 105 | 2 × 106 | 4.2 × 105 | 1.0 × 105 | 3.9 × 105 | 3 × 107 | - | - | 5.6 × 104 | 3.7 × 105 |
| Thigh muscle | 2.0 × 106 | 1.4 × 106 | 3 × 105 | 4.4 × 106 | 0.75 × 106 | 0.9 × 105 | 0.5 × 105 | - | - | - | - | - |
| Abdominal muscle | 1.25 × 106 | 1 × 106 | 4 × 105 | 2.1 × 106 | 3.9 × 105 | - | 6 × 105 | - | - | - | - | - |
| Back muscle | 7.9 × 105 | 5 × 105 | 1.8 × 105 | 1.9 × 106 | 3.1 × 105 | 8 × 104 | 2.5 × 105 | 4.1 × 107 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Wang, Z.-Y.; Zuo, M.-T.; Yang, K.; Sun, Z.-L.; Wu, Y.; Liu, Z.-Y. Excretion, Metabolism, and Tissue Distribution of Gelsemium elegans (Gardn. & Champ.) Benth in Pigs. Molecules 2022, 27, 2605. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27082605
Ma X, Wang Z-Y, Zuo M-T, Yang K, Sun Z-L, Wu Y, Liu Z-Y. Excretion, Metabolism, and Tissue Distribution of Gelsemium elegans (Gardn. & Champ.) Benth in Pigs. Molecules. 2022; 27(8):2605. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27082605
Chicago/Turabian StyleMa, Xiao, Zi-Yuan Wang, Meng-Ting Zuo, Kun Yang, Zhi-Liang Sun, Yong Wu, and Zhao-Ying Liu. 2022. "Excretion, Metabolism, and Tissue Distribution of Gelsemium elegans (Gardn. & Champ.) Benth in Pigs" Molecules 27, no. 8: 2605. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27082605