
Adaptive Laboratory Evolution of Halomonas bluephagenesis Enhances Acetate Tolerance and Utilization to Produce Poly(3-hydroxybutyrate)


Abstract
:1. Introduction
2. Results and Discussion
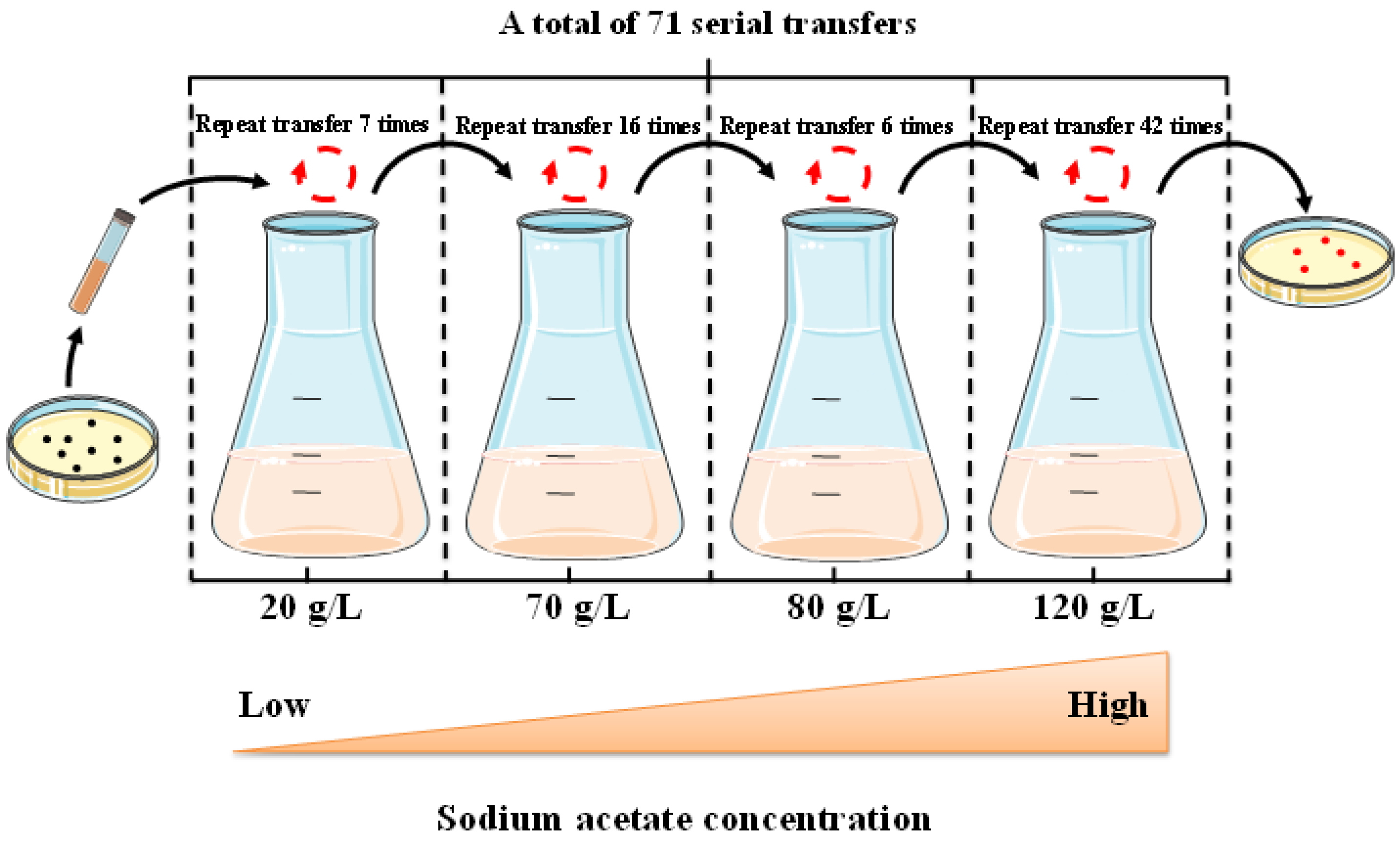
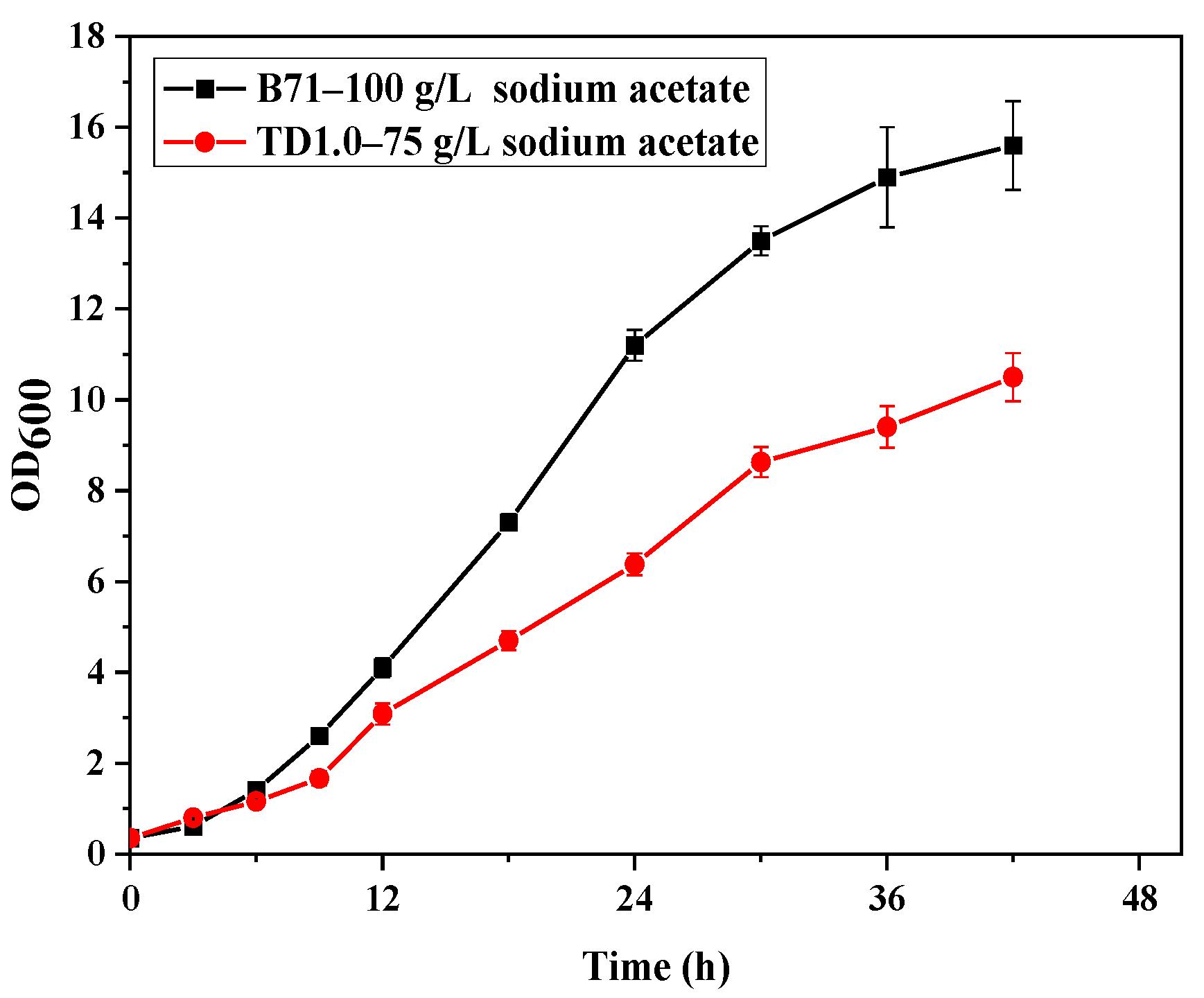
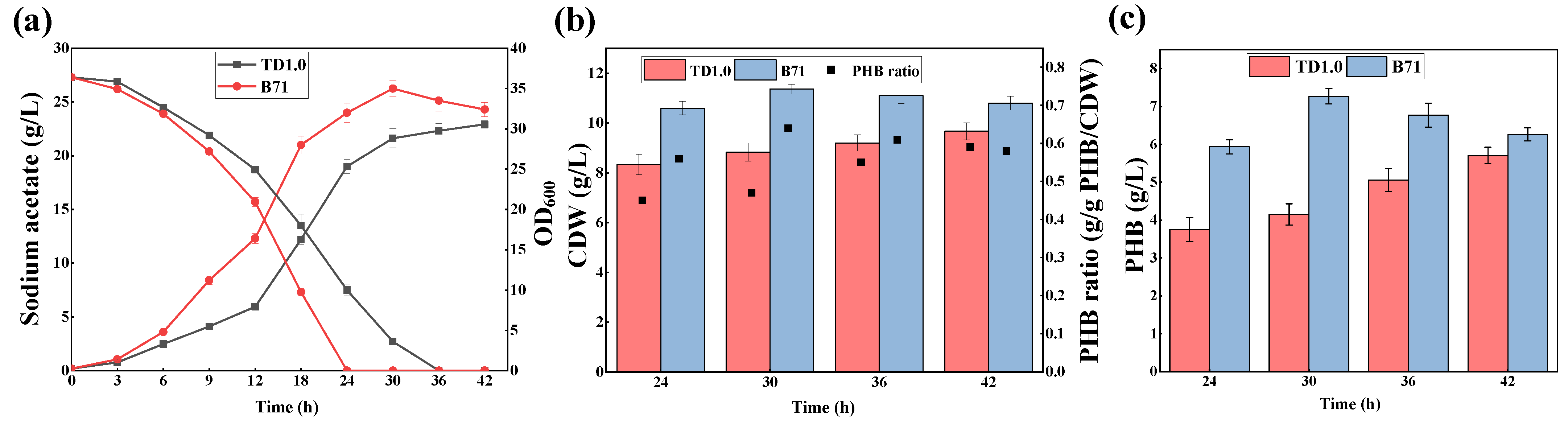
2.1. Adaptive Laboratory Evolution of H. Bluephagenesis in the Presence of Acetate
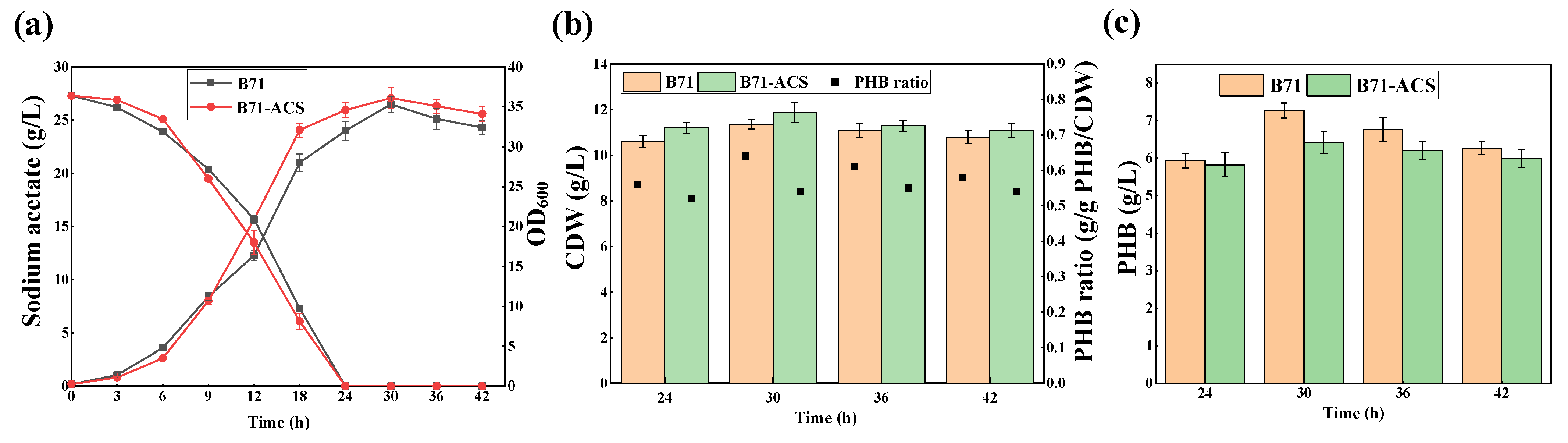
2.2. Overexpression of Acetyl-CoA Synthase in B71
2.3. Whole-Genome Sequence of the Evolved Strain B71
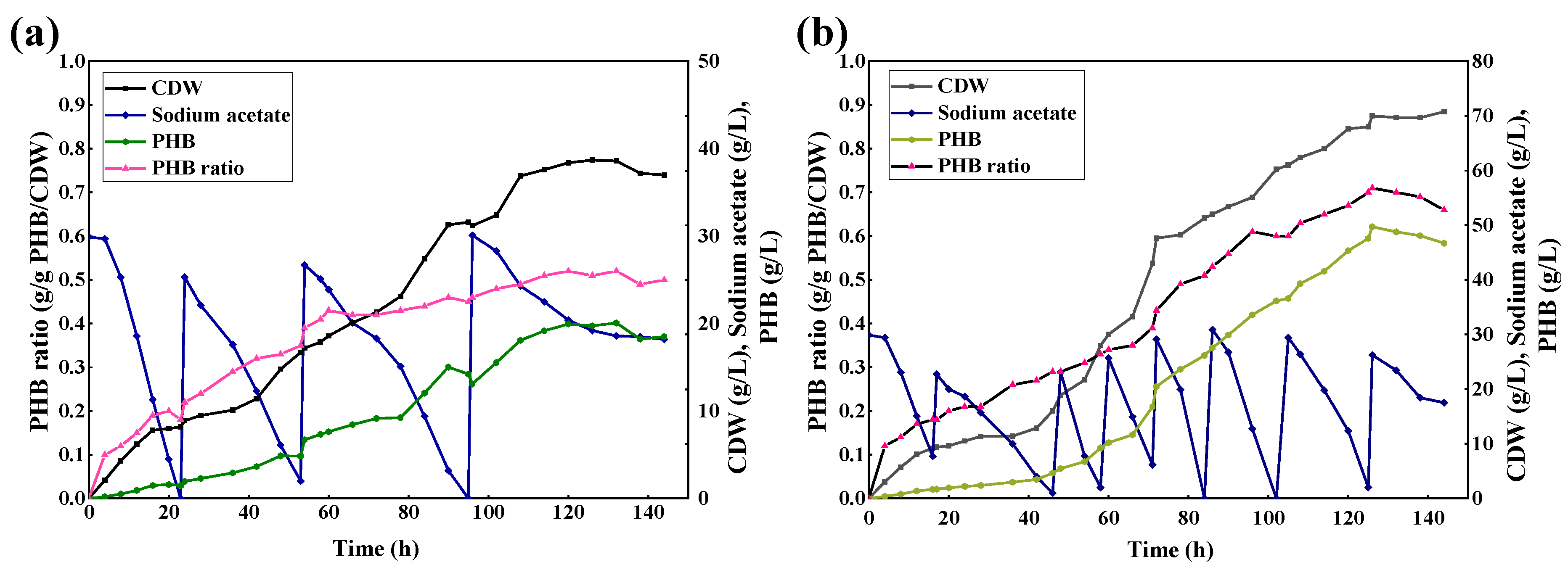
2.4. Fed-Batch for Evolved Strain B71
3. Materials and Methods
3.1. Strains and Growth Conditions
3.2. ALE Procedure for H. Bluephagenesis
3.3. Plasmids Construction and Conjugation
3.4. Shake-Flask and Fed-Batch Fermentation Studies
3.5. Analytical Methods
3.6. Whole-Genome Sequencing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Kiefer, D.; Merkel, M.; Lilge, L.; Henkel, M.; Hausmann, R. From acetate to bio-based products: Underexploited potential for industrial biotechnology. Trends Biotechnol. 2020, 39, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.G.; Lee, J.; Noh, M.; Jung, G. Rediscovering acetate metabolism: Its potential sources and utilization for biobased transformation into value-added chemicals. J. Agric. Food Chem. 2018, 66, 3998–4006. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lama, S.; Agrawal, D.; Kumar, V.; Park, S. Acetate as a potential feedstock for the production of value-added chemicals: Metabolism and applications. Biotechnol. Adv. 2021, 49, 107736. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Yang, H.; Fang, G.; Zhang, X.; Wu, H.; Li, Z.; Ye, Q. Central pathway engineering for enhanced succinate biosynthesis from acetate in Escherichia coli. Biotechnol. Bioeng. 2018, 115, 943–954. [Google Scholar] [CrossRef]
- Henningsen, B.; Hon, S.; Covalla, S.; Sonu, C.; Argyros, D.; Barrett, T.; Wiswall, E.; Froehlich, A.; Zelle, R. Increasing anaerobic acetate consumption and ethanol yields in Saccharomyces cerevisiae with NADPH-specific alcohol dehydrogenase. Appl. Environ. Microbiol. 2015, 81, 8108–8117. [Google Scholar] [CrossRef] [Green Version]
- Gerstmeir, R.; Wendisch, V.F.; Schnicke, S.; Ruan, H.; Farwick, M.; Reinscheid, D.; Eikmanns, B. Acetate metabolism and its regulation in Corynebacterium glutamicum. J. Biotechnol. 2003, 104, 99–122. [Google Scholar] [CrossRef]
- Christophe, G.; Deo, J.; Kumar, V.; Nouaille, R.; Fontanille, P.; Larroche, C. Production of oils from acetic acid by the oleaginous yeast Cryptococcus curvatus. Appl. Biochem. Biotechnol. 2011, 167, 1270–1279. [Google Scholar] [CrossRef]
- Canganella, F.; Kuk, S.U.; Morgan, H.; Wiegel, J. Clostridium thermobutyricum: Growth studies and stimulation of butyrate formation by acetate supplementation. Microbiol. Res. 2002, 157, 149–156. [Google Scholar] [CrossRef]
- Noh, M.; Lim, H.G.; Woo, S.; Song, J.; Jung, G. Production of itaconic acid from acetate by engineering acid-tolerant Escherichia coli W. Biotechnol. Bioeng. 2017, 115, 729–738. [Google Scholar] [CrossRef]
- Leone, S.; Sannino, F.; Tutino, M.; Parrilli, E.; Picone, D. Acetate: Friend or foe? Efficient production of a sweet protein in Escherichia coli BL21 using acetate as a carbon source. Microb. Cell Fact. 2015, 14, 106. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.; Dai, W.; Mao, Y.; Cui, Z.; Zhang, Z.; Wang, Z.; Ma, H.; Chen, T. Enhanced 3-hydroxypropionic acid production from acetate via the malonyl-CoA pathway in Corynebacterium glutamicum. Front. Bioeng. Biotechnol. 2021, 9, 808258. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, T.; Salazar, M.; Weng, L.; Palsson, B.; Feist, A. The emergence of adaptive laboratory evolution as an efficient tool for biological discovery and industrial biotechnology. Metab. Eng. 2019, 56, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Seong, W.; Han, G.H.; Lim, H.; Baek, J.; Kim, S.J.; Kim, D.; Kim, S.; Lee, H.; Kim, H.; Lee, S.G.; et al. Adaptive laboratory evolution of Escherichia coli lacking cellular byproduct formation for enhanced acetate utilization through compensatory ATP consumption. Metab. Eng. 2020, 62, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sandoval, M.T.; Huerta-Beristain, G.; Trujillo-Martinez, B.; Bustos, P.; González, V.; Bolivar, F.; Gosset, G.; Martinez, A. Laboratory metabolic evolution improves acetate tolerance and growth on acetate of ethanologenic Escherichia coli under non-aerated conditions in glucose-mineral medium. Appl. Microbiol. Biotechnol. 2012, 96, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Salas Navarrete, P.C.; de Oca Miranda, A.I.M.; Martinez, A.; Caspeta, L. Evolutionary and reverse engineering to increase Saccharomyces cerevisiae tolerance to acetic acid, acidic pH, and high temperature. Appl. Microbiol. Biotechnol. 2022, 106, 383–399. [Google Scholar] [CrossRef]
- Eiteman, M.; Altman, E. Overcoming acetate in Escherichia coli recombinant protein fermentation. Trends Biotechnol. 2006, 24, 530–536. [Google Scholar] [CrossRef]
- Sirohi, R.; Prakash Pandey, J.; Kumar Gaur, V.; Gnansounou, E.; Sindhu, R. Critical overview of biomass feedstocks as sustainable substrates for the production of polyhydroxybutyrate (PHB). Bioresour. Technol. 2020, 311, 123536. [Google Scholar] [CrossRef]
- Yu, L.P.; Yan, X.; Zhang, X.; Chen, X.B.; Wu, Q.; Jiang, X.R.; Chen, G.Q. Biosynthesis of functional polyhydroxyalkanoates by engineered Halomonas bluephagenesis. Metab. Eng. 2020, 59, 119–130. [Google Scholar] [CrossRef]
- Zhang, J.; Jin, B.; Hong, K.; Lv, Y.; Wang, Z.; Chen, T. Cell catalysis of citrate to itaconate by engineered Halomonas bluephagenesis. ACS Synth. Biol. 2021, 10, 3017–3027. [Google Scholar] [CrossRef]
- Zhao, C.; Zheng, T.; Feng, Y.; Wang, X.; Zhang, L.; Hu, Q.; Chen, J.; Wu, F.; Chen, G.Q. Engineered Halomonas spp. for production of l-Lysine and cadaverine. Bioresour. Technol. 2022, 349, 126865. [Google Scholar] [CrossRef]
- Ye, J.W.; Chen, G.Q. Halomonas as a chassis. Essays Biochem. 2021, 65, 393–403. [Google Scholar] [PubMed]
- Zhang, J.; Zhang, X.; Mao, Y.; Jin, B.; Guo, Y.; Wang, Z.; Chen, T. Substrate profiling and tolerance testing of Halomonas TD01 suggest its potential application in sustainable manufacturing of chemicals. J. Biotechnol. 2020, 316, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lasko, D.R.; Zamboni, N.; Sauer, U. Bacterial response to acetate challenge: A comparison of tolerance among species. Appl. Microbiol. Biotechnol. 2000, 54, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lin, Y.; Wu, Q.; Wang, Y.; Chen, G.Q. Synthetic biology and genome-editing tools for improving PHA metabolic engineering. Trends Biotechnol. 2019, 38, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Hsieh, C.W.; Chang, Y.S.; Wung, B.S. Effect of acetic acid on ethanol production by Zymomonas mobilis mutant strains through continuous adaptation. BMC Biotechnol. 2017, 17, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutscha, R.; Pflügl, S. Microbial upgrading of acetate into value-added products—Examining microbial diversity, bioenergetic constraints and metabolic engineering approaches. Int. J. Mol. Sci. 2020, 21, 8777. [Google Scholar] [CrossRef]
- Conrad, T.M.; Lewis, N.E.; Palsson, B.O. Microbial laboratory evolution in the era of genome-scale science. Mol. Syst. Biol. 2011, 7, 509. [Google Scholar] [CrossRef]
- Ma, H.; Zhao, Y.; Huang, W.; Zhang, L.; Wu, F.; Ye, J.; Chen, G.Q. Rational flux-tuning of Halomonas bluephagenesis for co-production of bioplastic PHB and ectoine. Nat. Commun. 2020, 11, 3313. [Google Scholar] [CrossRef]
- Matsumoto, K.; Tanaka, Y.; Watanabe, T.; Motohashi, R.; Ikeda, K.; Tobitani, K.; Yao, M.; Tanaka, I.; Taguchi, S. Directed evolution and structural analysis of NADPH-dependent acetoacetyl coenzyme A (acetoacetyl-CoA) reductase from Ralstonia eutropha reveals two mutations responsible for enhanced kinetics. Appl. Environ. Microbiol. 2013, 79, 6134–6139. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.; Mitaku, S. Mechanisms of secondary structure breakers in soluble proteins. Biophysics 2005, 1, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Veno, J.; Rahman, R.; Masomian, M.; Ali, M.S.M.; Kamarudin, N.H.A. Insight into improved thermostability of cold-adapted staphylococcal lipase by glycine to cysteine mutation. Molecules 2019, 24, 3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Samaddar, S.; Chatterjee, P.; Krishnamoorthy, R.; Jeon, S.; Sa, T. Structural and functional responses of microbial community with respect to salinity levels in a coastal reclamation land. Appl. Soil Ecol. 2019, 137, 96–105. [Google Scholar] [CrossRef]
- Omini, J.; Wojciechowska, I.; Skirycz, A.; Moriyama, H.; Obata, T. Association of the malate dehydrogenase-citrate synthase metabolon is modulated by intermediates of the Krebs tricarboxylic acid cycle. Sci. Rep. 2021, 11, 18770. [Google Scholar] [CrossRef] [PubMed]
- Trcek, J.; Mira, N.P.; Jarboe, L.R. Adaptation and tolerance of bacteria against acetic acid. Appl. Microbiol. Biotechnol. 2015, 99, 6215–6229. [Google Scholar] [CrossRef]
- Saeki, A.; Matsushita, K.; Takeno, S.; Taniguchi, M.; Toyama, H.; Theeragool, G.; Lotong, N.; Adachi, O. Enzymes responsible for acetate oxidation by acetic acid bacteria. Biosci. Biotechnol. Biochem. 1999, 63, 2102–2109. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J.L. The lipid bilayer membrane and its protein constituents. J. Gen. Physiol. 2018, 150, 1472–1483. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Liu, H.; Chen, X.; Liu, L. Engineering microbial membranes to increase stress tolerance of industrial strains. Metab. Eng. 2019, 53, 24–34. [Google Scholar] [CrossRef]
- Xu, C.; Sun, T.; Li, S.; Chen, L.; Zhang, W. Adaptive laboratory evolution of cadmium tolerance in Synechocystis sp. PCC 6803. Biotechnol. Biofuels 2018, 11, 205. [Google Scholar] [CrossRef]
- Rajaraman, E.; Agrawal, A.; Crigler, J.; Seipelt Thiemann, R.; Altman, E.; Eiteman, M.A. Transcriptional analysis and adaptive evolution of Escherichia coli strains growing on acetate. Appl. Microbiol. Biotechnol. 2016, 100, 7777–7785. [Google Scholar] [CrossRef]
- Li, Z.J.; Qiao, K.; Liu, N.; Stephanopoulos, G. Engineering Yarrowia lipolytica for poly-3-hydroxybutyrate production. J. Ind. Microbiol. Biotechnol. 2017, 44, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.; Bae, J.; Kang, S.; Cho, B.K.; Oh, M.K. Poly-3-hydroxybutyrate production in acetate minimal medium using engineered Methylorubrum extorquens AM1. Bioresour. Technol. 2022, 353, 127127. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Qiu, Y.Q.; Chen, X.T.; Liang, X.F.; Ren, L.H. Metabolomic insights into polyhydroxyalkanoates production by halophilic bacteria with acetic acid as carbon source. Biosci. Biotechnol. Biochem. 2019, 83, 1955–1963. [Google Scholar] [CrossRef]
- Shi, L.L.; Da, Y.Y.; Zheng, W.T.; Chen, G.Q.; Li, Z.J. Production of polyhydroxyalkanoate from acetate by metabolically engineered Aeromonas hydrophilia. J. Biosci. Bioeng. 2020, 130, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, W.; Zhang, Z.Z.; Tan, T.W.; Li, Z.J. Metabolic engineering of Escherichia coli for the synthesis of polyhydroxyalkanoates using acetate as a main carbon source. Microb. Cell Fact. 2018, 17, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.; Xue, Y.S.; Aibaidula, G.; Chen, G.Q. Unsterile and continuous production of polyhydroxybutyrate by Halomonas TD01. Bioresour. Technol. 2011, 102, 8130–8136. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Pühler, A. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in gram negative bacteria. Nat. Biotechnol. 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, H.M.; Chen, X.; Li, T.; Wu, Q.; Ouyang, Q.; Chen, G.Q. Novel T7-like expression systems used for Halomonas. Metab. Eng. 2017, 39, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Silva-Rocha, R.; Martínez-García, E.; Calles, B.; Chavarría, M.; Arce-Rodríguez, A.; de Las Heras, A.; Páez-Espino, A.D.; Durante-Rodríguez, G.; Kim, J.; Nikel, P.I.; et al. The standard european vector architecture (SEVA): A coherent platform for the analysis and deployment of complex prokaryotic phenotypes. Nucleic Acids Res. 2013, 41, D666–D675. [Google Scholar] [CrossRef]
- Sun, Z.; Ramsay, J.A.; Guay, M.; Ramsay, B.A. Carbon-limited fed-batch production of medium-chain-length polyhydroxyalkanoates from nonanoic acid by Pseudomonas putida KT2440. Appl. Microbiol. Biotechnol. 2007, 74, 69–77. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, Y.; Ji, M.; Zhang, X.; Wang, H.; Chen, Y.; Wu, Q.; Chen, G.Q. Hyperproduction of PHA copolymers containing high fractions of 4-hydroxybutyrate (4HB) by outer membrane-defected Halomonas bluephagenesis grown in bioreactors. Microb. Biotechnol. 2022, 15, 1586–1597. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | NCBI Number | Function | Position | Type a | Description b |
|---|---|---|---|---|---|
| phaB | WP_009725067.1 | The rate-limiting enzyme for PHB synthesis | Exonic | SNV SNV | G→C,Ala26Ala G→T,Gly29Cys |
| mdh | WP_186004630.1 | Key enzyme of TCA cycle | Exonic | SNV | C→T,Val14Ala |
| TolA | WP_050801109.1 | Cell envelope integrity protein | Exonic | In | 175429: 30 bp |
| OmpA family protein | WP_009724794.1 | Protect cells from the effects of environmental stresses and maintain osmotic balance in cells [32] | Upstream Upstream Upstream Upstream Upstream Upstream Upstream | SNV SNV SNV SNV SNV SNV SNV | 19715: C→T, 19716: T→G, 19728: G→A, 19731: A→G, 19732: G→A, 20622: A→G, 21072: G→A |
| Strain | Carbon Source | PHB (g/L) | CDW (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|---|
| Y. lipolytica | Acetate | 0.24 | 6.2 | Shake-flasks | [40] |
| Y. lipolytica | Acetate | 7.35 | 72.01 | Fed-batch | [40] |
| M. extorquens | Acetate | 0.7 | 2.06 | Shake-flasks | [41] |
| B. cereus | Acetate | 1.46 | 2.94 | Shake-flasks | [42] |
| A. hydrophilia | Acetate | 0.55 | 2.77 | Shake-flasks | [43] |
| E. coli JM109 (pBHR68+pMCS-pta-ackA) | Acetate | 1.27 | 3.02 | Shake-flasks | [44] |
| H. bluephagenesis B71 | Acetate | 7.27 | 11.36 | Shake-flasks | This study |
| H. bluephagenesis B71 | Acetate | 49.78 | 70.01 | Fed-batch | This study |
| Strains/Plasmids | Genotype/Description | Source/Reference |
|---|---|---|
| Strains | ||
| E. coli S17-1 pir | recA, thi-1. pro, hsdR, RP4-2-Tc::Mu-Km::Tn7, a donor strain used for conjugation | [46] |
| H. bluephagenesis TD1.0 | Wild type H. bluephagenesis TD01 derivative with Mmp1 RNA polymerase integrated into the genome, PJ23110-lacI-Ptac-MmP1 | [47] |
| B71 | Acetate-adapted strain | This study |
| B71-ACS | B71 harboring pN59-PMmp1-ACS | This study |
| Plasmids | ||
| pN59 | A high copy number expression vector, ColE1 replication origin, oriT, CmR | [48] |
| pN59-PMmp1-ACS | pN59 derivates, containing codon-optimized acs gene driven by PMmp1, a T7-like inducible promoter, CmR | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Jin, B.; Fu, J.; Wang, Z.; Chen, T. Adaptive Laboratory Evolution of Halomonas bluephagenesis Enhances Acetate Tolerance and Utilization to Produce Poly(3-hydroxybutyrate). Molecules 2022, 27, 3022. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27093022
Zhang J, Jin B, Fu J, Wang Z, Chen T. Adaptive Laboratory Evolution of Halomonas bluephagenesis Enhances Acetate Tolerance and Utilization to Produce Poly(3-hydroxybutyrate). Molecules. 2022; 27(9):3022. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27093022
Chicago/Turabian StyleZhang, Jing, Biao Jin, Jing Fu, Zhiwen Wang, and Tao Chen. 2022. "Adaptive Laboratory Evolution of Halomonas bluephagenesis Enhances Acetate Tolerance and Utilization to Produce Poly(3-hydroxybutyrate)" Molecules 27, no. 9: 3022. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27093022