
Biotechnological Approaches for Production of Artemisinin, an Anti-Malarial Drug from Artemisia annua L.

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Regeneration Studies
2.1. Direct Organogenesis
2.2. Indirect Organogenesis
2.3. Somatic Embryogenesis
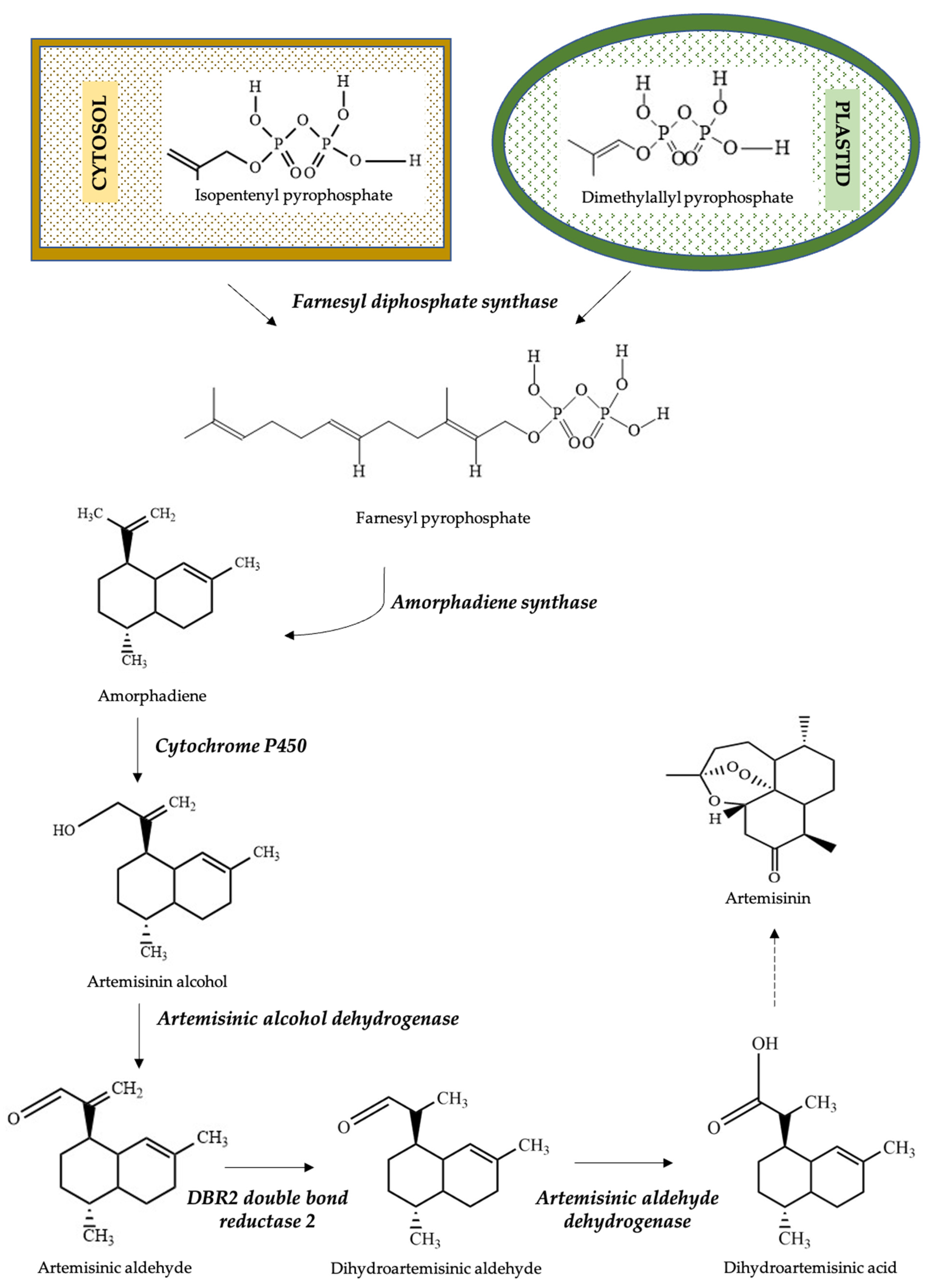
3. Biosynthesis of Artemisinin from Artemisia annua L.
4. In Vitro Production of Artemisinin from Artemisia
4.1. Organ Cultures
4.1.1. Shoot Culture
4.1.2. Hairy Root Culture (Agrobacterium rhizogenes Mediated Transformation)
4.2. Callus and Cell Suspension Cultures
5. Agrobacterium tumefaciens Mediated Transformation for Increased Metabolite Content
6. Elicitation Strategy for Production of Artemisinin
6.1. Biotic Elicitation for Artemisinin Production
6.2. Abiotic Elicitation for Artemisinin Production
7. Bioreactor Scale Production of Artemisinin
8. Metabolic Engineering for Artemisinin Production
9. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woerdenbag, H.; Pras, N.; Alfermann, A. Production of artemisinin in shoot cultures of Artemisia annua. Planta Med. 1991, 57, A91–A92. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Janick, J. Distribution of artemisinin in Artemisia annua. In Progress in New Crops; Janick, J., Ed.; ASHS Press: Arlington, VA, USA, 1996; pp. 579–584. [Google Scholar]
- Eastman, R.T.; Fidock, D.A. Artemisinin-based combination therapies: A vital tool in efforts to eliminate malaria. Nat. Rev. Genet. 2009, 7, 864–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. World Malaria Report 2015. Available online: http://www.who.int/malaria/publications/world-malaria-report-2015/report/en/ (accessed on 2 April 2022).
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Zhou, W.-W.; Meng, J.; Du, X.-L.; Sui, Y.-P.; Dai, L.; Wang, P.-Q.; Huo, H.-R.; Sui, F. The pharmacological activities and mechanisms of artemisinin and its derivatives: A systematic review. Med. Chem. Res. 2017, 26, 867–880. [Google Scholar] [CrossRef]
- Chen, M.; Yan, T.; Shen, Q.; Lu, X.; Pan, Q.; Huang, Y.; Tang, Y.; Fu, X.; Liu, M.; Jiang, W.; et al. Glandular trichome - specific WRKY 1 promotes artemisinin biosynthesis in Artemisia annua. New Phytol. 2016, 214, 304–316. [Google Scholar] [CrossRef]
- Mondal, A.; Chatterji, U. Artemisinin represses telomerase subunits and induces apoptosis in HPV-39 infected human cervical cancer cells. J. Cell. Biochem. 2015, 116, 1968–1981. [Google Scholar] [CrossRef]
- Wu, Y.; He, S.; Bai, B.; Zhang, L.; Xue, L.; Lin, Z.; Yang, X.; Zhu, F.; He, P.; Tang, W.; et al. Therapeutic effects of the artemisinin analog SM934 on lupus-prone MRL/lpr mice via inhibition of TLR-triggered B-cell activation and plasma cell formation. Cell. Mol. Immunol. 2015, 13, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Slade, D.; Galal, A.M.; Gul, W.; Radwan, M.; Ahmed, S.A.; Khan, S.I.; Tekwani, B.L.; Jacob, M.R.; Ross, S.A.; ElSohly, M.A. Antiprotozoal, anticancer and antimicrobial activities of dihydroartemisinin acetal dimers and monomers. Bioorganic Med. Chem. 2009, 17, 7949–7957. [Google Scholar] [CrossRef] [Green Version]
- Obeid, S.; Alen, J.; Nguyen, V.H.; Pham, V.C.; Meuleman, P.; Pannecouque, C.; Le, T.N.; Neyts, J.; Dehaen, W.; Paeshuyse, J. Artemisinin analogues as potent inhibitors of In vitro hepatitis C virus replication. PLoS ONE 2013, 8, e81783. [Google Scholar] [CrossRef] [Green Version]
- Fulzele, D.P.; Sipahimalani, A.T.; Heble, M.R. Tissue cultures of Artemisia annua: Organogenesis and artemisinin production. Phytother. Res. 1991, 5, 149–153. [Google Scholar] [CrossRef]
- Jamaleddine, Z.O.; Lyam, P.; Fajimi, O.; Giwa, A.; Aina, A.; Lawyer, E.F.; Okere, A.U.; Odofin, W.T. In vitro growth response of Artemisia annua seeds to different concentrations of plant growth regulators. Afr. J. Biotechnol. 2011, 10, 17841–17844. [Google Scholar] [CrossRef]
- Blando, F.; Rizzello, F.; Durante, M.; De Paolis, A.; Caretto, S.; Mita, G. In Vitro adventitious regeneration of Artemisia annua L. influencing artemisinin metabolism. Horticulturae 2021, 7, 438. [Google Scholar] [CrossRef]
- Lualon, W.; De-Eknamkul, W.; Tanaka, H.; Shoyama, Y.; Putalun, W. Artemisinin production by shoot regeneration of Artemisia annua L. using thidiazuron. Z. Nat. C. J. Biosci. 2008, 63, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Gupta, M.M.; Khanuja, S.P.S. Maintaining the artemisinin content through direct and indirect in vitro regeneration and their assessment of variations with the field grown mother plants of Artemisia annua L. Nat. Prod. Bioprospecting 2012, 2, 194–199. [Google Scholar] [CrossRef] [Green Version]
- Janarthanam, B.; Rashmi, P.; Sumathi, E. Rapid and efficient plant regeneration from nodal explants of Artemisia annua L. Plant Tissue Cult. Biotechnol. 2012, 22, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Gaurav, S.; Vinay, S.; Veena, A. An efficient micropropagation protocol of an elite clone EC-353508 of Artemisia annua L., an important antimalarial plant. Int. J. Pharma Bio Sci. 2011, 2, 205–214. [Google Scholar]
- Hailu, T.; Abera, B.; Mariam, G. In vitro mass propagation of Artemisia (Artemisia annua L.) cv: Anamed. Plant Tissue Cult. Biotechnol. 2014, 23, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Almaarri, K.; Xie, Y. In vitro direct organogenesis and micropropagation of Artemisia annua. J. Biotechnol. Vegetale. 2010, 26, 327–337. [Google Scholar]
- Lei, C.; Wang, H.; Liu, B.; Ye, H. Effects of silver nitrate on shoot regeneration of Artemisia annua L. Plant Biotechnol. 2014, 31, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Mirzaee, H.; Sharafi, A.; Sohi, H.H. In vitro regeneration and transient expression of recombinant sesquiterpene cyclase (SQC) in Artemisia annua L. South Afr. J. Bot. 2016, 104, 225–231. [Google Scholar] [CrossRef]
- Mathur, A.K.; Kumar, S. Micropropagation of Artemisia annua via the inflorescence. J. Herbs Spices Med. Plants 1996, 4, 61–71. [Google Scholar] [CrossRef]
- Paniego, N.B.; Giulietti, A.M. Artemisia annua L.: Dedifferentiated and differentiated cultures. Plant Cell Tissue Organ Cult. 1994, 36, 163–168. [Google Scholar] [CrossRef]
- Gulati, A.; Bharel, S.; Jain, S.K.; Abdin, M.Z.; Srivastava, P.S. In Vitro micropropagation and flowering in Artemisia annua. J. Plant Biochem. Biotechnol. 1996, 5, 31–35. [Google Scholar] [CrossRef]
- Alejos-Gonzalez, F.; Perkins, K.; Winston, M.I.; Xie, D.-Y. Efficient somatic embryogenesis and organogenesis of self-pollination Artemisia annua progeny and artemisinin formation in regenerated plants. Am. J. Plant Sci. 2013, 04, 2206–2217. [Google Scholar] [CrossRef] [Green Version]
- Zayova, E.; Nedev, T.; Petrova, D.; Zhiponova, M.; Kapchina, V.; Chaneva, G. Tissue culture applications of Artemisia annua L. callus for indirect organogenesis and production phytochemical. Plant Tissue Cult. Biotechnol. 2020, 30, 97–106. [Google Scholar] [CrossRef]
- Zimmerman, J.L. Somatic Embryogenesis: A model for early development in higher plants. Plant Cell 1993, 5, 1411–1423. [Google Scholar] [CrossRef] [Green Version]
- Weathers, P.J.; Arsenault, P.R.; Covello, P.S.; McMickle, A.; Teoh, K.H.; Reed, D.W. Artemisinin production in Artemisia annua: Studies in planta and results of a novel delivery method for treating malaria and other neglected diseases. Phytochem. Rev. 2010, 10, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Teoh, K.H.; Polichuk, D.R.; Reed, D.W.; Nowak, G.; Covello, P.S. Artemisia annua L. (Asteraceae) trichome-specific cDNAs reveal CYP71AV1, a cytochrome P450 with a key role in the biosynthesis of the antimalarial sesquiterpene lactone artemisinin. FEBS Lett. 2006, 580, 1411–1416. [Google Scholar] [CrossRef] [Green Version]
- Weathers, P.J.; Elkholy, S.F.; Wobbe, K.K. Artemisinin: The biosynthetic pathway and its regulation in Artemisia annua, a terpenoid-rich species. Vitr. Cell. Dev. Biol. -Plant 2006, 42, 309–317. [Google Scholar] [CrossRef]
- Ikram, N.K.B.K.; Simonsen, H.T. A Review of biotechnological artemisinin production in plants. Front. Plant Sci. 2017, 8, 1966. [Google Scholar] [CrossRef] [Green Version]
- Farhi, M.; Marhevka, E.; Ben-Ari, J.; Algamas-Dimantov, A.; Liang, Z.; Zeevi, V.; Edelbaum, O.; Spitzer-Rimon, B.; Abeliovich, H.; Schwartz, B.; et al. Generation of the potent anti-malarial drug artemisinin in tobacco. Nat. Biotechnol. 2011, 29, 1072–1074. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.-Y.; Ma, D.-M.; Judd, R.; Jones, A.L. Artemisinin biosynthesis in Artemisia annua and metabolic engineering: Questions, challenges, and perspectives. Phytochem. Rev. 2016, 15, 1093–1114. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, Y.; Wang, Y. Artemisinin: Current state and perspectives for biotechnological production of an antimalarial drug. Appl. Microbiol. Biotechnol. 2006, 72, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, Y.; Ouyang, F.; Ye, H.; Li, G. Production of artemisinin by hairy root cultures of Artemisia annua L in bioreactor. Biotechnol. Lett. 1998, 20, 265–268. [Google Scholar] [CrossRef]
- Weathers, P.J.; Cheetham, R.D.; Follansbee, E.; Teoh, K. Artemisinin production by transformed roots of Artemisia annua. Biotechnol. Lett. 1994, 16, 1281–1286. [Google Scholar] [CrossRef]
- Paniego, N.; Giulietti, A.M. Artemisinin production by Artemisia annua L.-transformed organ cultures. Enzym. Microb. Technol. 1996, 18, 526–530. [Google Scholar] [CrossRef]
- Woerdenbag, H.; Lüers, J.F.J.; Van Uden, W.; Pras, N.; Malingré, T.M.; Alfermann, A.W. Production of the new antimalarial drug artemisinin in shoot cultures of Artemisia annua L. Plant Cell Tissue Organ Cult. 1993, 32, 247–257. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Janick, J. Immunoquantitative analysis of artemisinin from Artemisia annua using polyclonal antibodies. Phytochemistry 1996, 41, 97–104. [Google Scholar] [CrossRef]
- Martinez, B.C.; Staba, E.J. The Production of artemisinin in Artemisia annua L. tissue cultures. Adv. Cell Culture 1988, 6, 69–87. [Google Scholar] [CrossRef]
- Liu, C.; Guo, C.; Wang, Y.; Ouyang, F. Factors influencing artemisinin production from shoot cultures of Artemisia annua L. World J. Microbiol. Biotechnol. 2003, 19, 535–538. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Janick, J. Roots as an enhancing factor for the production of artemisinin in shoot cultures of Artemisia annua. Plant Cell Tissue Organ Cult. 1996, 44, 211–217. [Google Scholar] [CrossRef]
- Ahlawat, S. Influence of Agrobacterium rhizogenes on induction of hairy roots for enhanced production of artemisinin in Artemisia annua L. plants. Afr. J. Biotechnol. 2012, 11, 35. [Google Scholar] [CrossRef]
- Gantait, S.; Mukherjee, E. Hairy root culture technology: Applications, constraints and prospect. Appl. Microbiol. Biotechnol. 2020, 105, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Beigmohammadi, D.M.; Seyyedi, M.; Rostampour, S.; Mohammadi, E.; Sharafi, A. In vitro regeneration and genetically transformed culture of Artemisia diffusa. J. Appl. Biotechnol. Rep. 2021, 8, 312–319. [Google Scholar]
- Timina, O.; Timin, O.; Stepanova, A.; Shchuka, T. Getting bearded roots in Artemisia annua L. In Proceedings of the 90 Years—From Plant to Medicine: Achievements and Prospects, Moscow, Russia, 10 June 2021. (in Russian). [Google Scholar] [CrossRef]
- Dilshad, E.; Cusido, R.M.; Estrada, K.R.; Bonfill, M.; Mirza, B. Genetic transformation of Artemisia carvifolia Buch with rol genes enhances artemisinin accumulation. PLoS ONE 2015, 10, e0140266. [Google Scholar] [CrossRef] [Green Version]
- Brown, G. Production of anti-malarial and anti-migraine drugs in tissue culture of Artemisia annua and Tanacetum parthenium. Acta Hortic. 1993, 269–276. [Google Scholar] [CrossRef]
- Woerdenbag, H.J.; Pras, N.; Van Uden, W.; De Boer, A.; Batterman, S.; Visser, J.F.; Malingré, T.M. High peroxidase activity in cell cultures of Artemisia annua with minute artemisinin contents. Nat. Prod. Lett. 1992, 1, 121–128. [Google Scholar] [CrossRef]
- Liu, K.-S.; Yang, S.-L.; Roberts, M.; Elford, B.; Phillipson, J. Antimalarial activity of Artemisia annua flavonoids from whole plants and cell cultures. Plant Cell Rep. 1992, 11, 637–640. [Google Scholar] [CrossRef]
- Baldi, A.; Dixit, V. Yield enhancement strategies for artemisinin production by suspension cultures of Artemisia annua. Bioresour. Technol. 2008, 99, 4609–4614. [Google Scholar] [CrossRef]
- Kiani, B.H.; Safdar, N.; Mannan, A.; Mirza, B. Comparative Artemisinin analysis in Artemisia dubia transformed with two different Agrobacteria harbouringrol ABC genes. Plant Omics. 2012, 5, 386–391. [Google Scholar]
- Ghosh, B.; Mukherjee, S.; Jha, S. Genetic transformation of Artemisia annua by Agrobacterium tumefaciens and artemisinin synthesis in transformed cultures. Plant Sci. 1997, 122, 193–199. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The genecards suite: From gene data mining to disease genome sequence analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-H.; Ye, H.-C.; Li, G.-F. Expression of a chimeric farnesyl diphosphate synthase gene in Artemisia annua L. transgenic plants via Agrobacterium tumefaciens-mediated transformation. Plant Sci. 2000, 155, 179–185. [Google Scholar] [CrossRef]
- Wen, W.J.; Hao, X.Z.; Xiang, T.R. Elicitation on artemisinin biosynthesis in Artemisia annua hairy roots by the oligosaccharide extract from the Endophytic colletotrichum sp. B501. J. Integr. Plant Biol. 2002, 44, 1233. [Google Scholar]
- Wang, J.W.; Zheng, L.P.; Zhang, B.; Zou, T. Stimulation of artemisinin synthesis by combined cerebroside and nitric oxide elicitation in Artemisia annua hairy roots. Appl. Microbiol. Biotechnol. 2009, 85, 285–292. [Google Scholar] [CrossRef]
- Zheng, L.-P.; Guo, Y.-T.; Wang, J.-W.; Tan, R.-X. Nitric Oxide potentiates Oligosaccharide-induced artemisinin production in Artemisia annua hairy roots. J. Integr. Plant Biol. 2008, 50, 49–55. [Google Scholar] [CrossRef]
- Wang, J.W.; Zhang, Z.; Tan, R.X. Stimulation of artemisinin production in Artemisia annua hairy roots by the elicitor from the endophytic Colletotrichum sp. Biotechnol. Lett. 2001, 23, 857–860. [Google Scholar] [CrossRef]
- Sharma, G.; Agrawal, V. Marked enhancement in the artemisinin content and biomass productivity in Artemisia annua L. shoots co-cultivated with Piriformospora indica. World J. Microbiol. Biotechnol. 2013, 29, 1133–1138. [Google Scholar] [CrossRef]
- Zheng, L.P.; Tian, H.; Yuan, Y.F.; Wang, J.W. The influence of endophytic Penicillium oxalicum B4 on growth and artemisinin biosynthesis of in vitro propagated plantlets of Artemisia annua L. Plant Growth Regul. 2016, 80, 93–102. [Google Scholar] [CrossRef]
- Zhang, B.; Zheng, L.P.; Yi Li, W.; Wen Wang, J. Stimulation of artemisinin production in Artemisia annua hairy roots by Ag-SiO2 core-shell nanoparticles. Curr. Nanosci. 2013, 9, 363–370. [Google Scholar] [CrossRef]
- Ghassemi, B.; Nayeri, F.D.; Hosseini, R. The effects of chitosan nanoparticles on genes expression of artemisinin synthase in suspension culture of Artemisia annua L.: A comparative study. Int. J. Adv. Biol. Biomed. Research 2021, 9, 190–203. [Google Scholar]
- Ghasemi, B.; Hosseini, R.; Nayeri, F.D. Effects of cobalt nanoparticles on artemisinin production and gene expression in Artemisia annua. Turk. J. Bot. 2015, 39, 769–777. [Google Scholar] [CrossRef]
- Yee, M.K.; Ping, W.Y. Successes of artemisinin elicitation in low-artemisinin producing Artemisia annua cell cultures constrained by repression of biosynthetic genes. bioRxiv 2019, 740167. [Google Scholar] [CrossRef] [Green Version]
- Mannan, A.; Liu, C.; Arsenault, P.R.; Towler, M.J.; Vail, D.R.; Lorence, A.; Weathers, P.J. DMSO triggers the generation of ROS leading to an increase in artemisinin and dihydroartemisinic acid in Artemisia annua shoot cultures. Plant Cell Rep. 2009, 29, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Durante, M.; Caretto, S.; Quarta, A.; De Paolis, A.; Nisi, R.; Mita, G. β-Cyclodextrins enhance artemisinin production in Artemisia annua suspension cell cultures. Appl. Microbiol. Biotechnol. 2011, 90, 1905–1913. [Google Scholar] [CrossRef]
- Pontiggia, D.; Ciarcianelli, J.; Salvi, G.; Cervone, F.; De Lorenzo, G.; Mattei, B. Sensitive detection and measurement of oligogalacturonides in Arabidopsis. Front. Plant Sci. 2015, 6, 258. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Zou, T.; Lu, Y.H.; Wang, J.W. Stimulation of artemisinin biosynthesis in Artemisia annua hairy roots by oligogalacturonides. Afr. J. Biotechnol. 2010, 9, 3437–3442. [Google Scholar] [CrossRef]
- Patil, A.; Suryavanshi, P.; Fulzele, D. In vitro regeneration of gamma irradiated callus of Artemisia annua and evaluation of increase artemisinin content by HPLC analysis. J. Anal. Pharm. Res. 2018, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Salehi, M.; Karimzadeh, G.; Naghavi, M.R. Synergistic effect of coronatine and sorbitol on artemisinin production in cell suspension culture of Artemisia annua L. cv. Anamed. Plant Cell Tissue Organ Cult. 2019, 137, 587–597. [Google Scholar] [CrossRef]
- SaghirzadehDarki, B.; Shabani, L.; Pourvaez, R.; Ghannadian, M. Effects of CuSO4 and AgNO3 on artemisinin and phenolic compound in shoot cultures of Artemisia annua L. J. Plant Physiol. 2010, 31, 1–8. [Google Scholar]
- Ahlawat, S.; Saxena, P.; Alam, P.; Wajid, S.; Abdin, M.Z. Modulation of artemisinin biosynthesis by elicitors, inhibitor, and precursor in hairy root cultures of Artemisia annua L. J. Plant Interact. 2014, 9, 811–824. [Google Scholar] [CrossRef]
- Putalun, W.; Luealon, W.; De-Eknamkul, W.; Tanaka, H.; Shoyama, Y. Improvement of artemisinin production by chitosan in hairy root cultures of Artemisia annua L. Biotechnol. Lett. 2007, 29, 1143–1146. [Google Scholar] [CrossRef] [PubMed]
- Patra, N.; Srivastava, A.K.; Sharma, S. Study of various factors for enhancement of artemisinin in Artemisia annua hairy roots. Int. J. Chem. Eng. Appl. 2013, 157–160. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Kiani, B.H.; Mannan, A.; Ismail, T.; Mirza, B. Enhanced production of artemisinin by hairy root cultures of Artemisia dubia. J. Med. Plants Res. 2012, 6. [Google Scholar] [CrossRef]
- Zia, M.; Mannan, A.; Chaudhary, M.F. Effect of growth regulators and amino acids on artemisinin production in the callus of Artemisia absinthium. Pak. J. Bot. 2007, 39, 799–805. [Google Scholar]
- Yancheva, S.; Georgieva, L.; Badjakov, I.; Dincheva, I.; Georgieva, M.; Georgiev, V.; Kondakova, V. Application of bioreactor technology in plant propagation and secondary metabolite production. J. Cent. Eur. Agric. 2019, 20, 321–340. [Google Scholar] [CrossRef]
- Patra, N.; Srivastava, A.K. Artemisinin production by plant hairy root cultures in gas- and liquid-phase bioreactors. Plant Cell Rep. 2015, 35, 143–153. [Google Scholar] [CrossRef]
- Liu, C.-Z.; Guo, C.; Wang, Y.-C.; Ouyang, F. Comparison of various bioreactors on growth and artemisinin biosynthesis of Artemisia annua L. shoot cultures. Process. Biochem. 2003, 39, 45–49. [Google Scholar] [CrossRef]
- Liu, C.-Z.; Wang, Y.-C.; Guo, C.; Ouyang, F.; Ye, H.-C.; Li, G.-F. Production of artemisinin by shoot cultures of Artemisia annua L. in a modified inner-loop mist bioreactor. Plant Sci. 1998, 135, 211–217. [Google Scholar] [CrossRef]
- Wyslouzil, B.E.; Waterbury, R.G.; Weathers, P.J. The growth of single roots of Artemisia annua in nutrient mist reactors. Biotechnol. Bioeng. 2000, 70, 143–150. [Google Scholar] [CrossRef]
- Patra, N.; Srivastava, A.K. Use of model-based nutrient feeding for improved production of artemisinin by hairy roots of Artemisia annua in a modified stirred tank bioreactor. Appl. Biochem. Biotechnol. 2015, 177, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Patra, N.; Srivastava, A.K. Mass scale artemisinin production in a stirred tank bioreactor using hairy roots of Artemisia annua. Int. J. Biosci. Biochem. Bioinform. 2014, 467–474. [Google Scholar] [CrossRef]
- Liu, C.Z.; Wang, Y.C.; Kang, X.Z.; Fan, O.Y.; Ye, H.C.; Li, G.F. Artemisinin production by adventitious shoots of Artemisia annua in a novel mist bioreactor. Acta Bot. Sin. 1999, 41, 524–527. [Google Scholar]
- Kim, Y.; Wyslouzil, B.E.; Weathers, P.J. A comparative study of mist and bubble column reactors in the in vitro production of artemisinin. Plant Cell Rep. 2001, 20, 451–455. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Y.; Ouyang, F.; Ye, H.; Li, G. Production of artemisinin by hairy root cultures of Artemisia annua L. Biotechnol. Lett. 1997, 19, 927–929. [Google Scholar] [CrossRef]
- Martin, V.J.J.; Pitera, D.J.; Withers, S.T.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802. [Google Scholar] [CrossRef]
- Tremblay, R.; Wang, D.; Jevnikar, A.M.; Ma, S. Tobacco, a highly efficient green bioreactor for production of therapeutic proteins. Biotechnol. Adv. 2010, 28, 214–221. [Google Scholar] [CrossRef]
- Staub, J.M.; Garcia, B.; Graves, J.; Hajdukiewicz, P.T.J.; Hunter, P.; Nehra, N.; Paradkar, V.; Schlittler, M.; Carroll, J.A.; Spatola, L.; et al. High-yield production of a human therapeutic protein in tobacco chloroplasts. Nat. Biotechnol. 2000, 18, 333–338. [Google Scholar] [CrossRef]
- Wallaart, T.E.; Bouwmeester, H.J.; Hille, J.; Poppinga, L.; Maijers, N.C.A. Amorpha-4,11-diene synthase: Cloning and functional expression of a key enzyme in the biosynthetic pathway of the novel antimalarial drug artemisinin. Planta 2001, 212, 460–465. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Schalk, M.; Clark, A.; Miles, R.B.; Coates, R.; Chappell, J. Redirection of cytosolic or plastidic isoprenoid precursors elevates terpene production in plants. Nat. Biotechnol. 2006, 24, 1441–1447. [Google Scholar] [CrossRef]
- Van Herpen, T.W.J.M.; Cankar, K.; Nogueira, M.; Bosch, D.; Bouwmeester, H.J.; Beekwilder, J. Nicotiana benthamiana as a production platform for artemisinin precursors. PLoS ONE 2010, 5, e14222. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Explant | Media | Response | References |
|---|---|---|---|
| Seed | MS + 0.1 ppm BA + 1.0 ppm NAA | Shooting | [12] |
| Inflorescence | MS + 1.0 mg/L BAP + 2.0 mg/L IBA | Multiple shooting | [23] |
| Stem | MS + 0.1 mg/L TDZ | Multiple shooting | [15] |
| Leaf, petiole | MS + 1.0 mg/L TDZ | Shooting and rooting | [20] |
| Seed | MS + 0.1 mg/L KN + 0.01 mg/L NAA | Shooting | [13] |
| Nodal stem explants | MS + 0.2 mg/L NAA + 0.2 mg/L BAP | Multiple shooting | [16] |
| Nodal stem explants | Shooting: MS + 4.44 µM BAP Rooting: ½ MS + 2.46 µM IBA | Multiple shooting and rooting | [17] |
| Nodal stem explants | Shooting: MS + 10.0 µM 2-iP; Rooting: 1/2 MS + 5.0 µM NAA | Multiple shooting and rooting | [18] |
| Nodal stem explants | Shooting: MS + 0.8 mg/L BAP + 0.1 mg/L IBA; Multiplication medium: MS + 1.0 mg/L BAP + 0.1 mg/L IBA; Rooting: 1/2 MS + 0.5 mg/L IBA | Multiple shooting and rooting | [19] |
| Leaf | Shooting: MS + 1.0 mg/L BAP + 0.05 mg/L NAA + 2.0 mg/L AgNO3 | Shoot regeneration | [21] |
| Leaf | Shooting: MS + 0.5 mg/L NAA + 2.0 mg/L BA; Rooting: MS + 0.1 mg/L IBA | Shooting and rooting | [22] |
| Seed | Shooting: MS + 4.4 µM BA + 0.35 µM IBA; Multiplication medium: MS + 0.9 µM BA + 0.05 µM NAA | Shooting and multiplication | [14] |
| Explant | Media | Response | References |
|---|---|---|---|
| Leaf, hypocotyl | Callus: MS + 5.4 µM NAA; Shooting: MS + 13.32 µM BA + 1.08 µM NAA | Callusing and organogenesis | [24] |
| Hypocotyl | Callusing and shooting: MS + 0.5 µM NAA + 13 µM BAP + 0.3 µM GA3 | Callusing and multiple shooting | [25] |
| Leaf | Callusing: MS + 0.1 mg/L BAP + 0.05 mg/L NAA; Shooting: 0.4 mg/L BAP + 0.2 mg/L NAA | Callusing and multiple shooting | [16] |
| Leaf | Callusing: MS + 1.0 mg/L BAP + 0.05 mg/L NAA | Callusing and organogenesis | [26] |
| Leaf | Callusing: MS + 0.5 mg/L NAA or 2,4-D + 0.5 mg/L BAP; Shooting: 0.25 mg/L NAA + 1.0 mg/L BAP; Rooting: ½ MS + 0.1 mg/L IBA | Callusing and organogenesis | [27] |
| Elicitor | Culture Type | Culture Conditions | Yield of Artemisinin | Reference |
|---|---|---|---|---|
| Biotic elicitors | ||||
| Cell wall’s oligosaccharide from Colletotrichum sp. B501 | Hairy root culture | MS medium + 20 mg/L elicitor | Increased by 68.29% | [57] |
| Cerebroside from fungal source | Hairy root culture | MS medium + 10–70 µg/mL cerebroside | Increased by 2.3 folds | [58] |
| Oligosaccharide from Fusarium oxysporum mycelium | Hairy root culture | MS medium + 0.3 mg total sugar/mL elicitor | Increased from 0.7 mg/g DW to 1.3 mg/g DW | [59] |
| Mycelial extract of Colletotrichum sp. | Hairy root culture | MS medium + 0.4 mg total sugar/mL elicitor | Increased from 0.8 mg/g DW to 1 mg/g DW | [60] |
| Pencillium oxalium B4 | In vitro grown Rooted plantlets | MS medium + 5.0 mg/L BAP + 1.0 mg/L NAA + P. oxalium B4 (30 days exposure) | Increased by 43.5% | [62] |
| Abiotic elicitors | ||||
| Ag-SiO2 nanoparticles | Hairy root culture | MS medium + 900 mg/L nano elicitor | Increased by 3.9 folds | [63] |
| Chitosan nanoparticles | Cell suspension culture | MS medium + 0.5 mg/l NAA + 0.5 mg/L BAP + 15 mg/L Elicitor | NA | [64] |
| Cobalt nano particles | Callus culture | MS medium + 0.5 mg/L NAA + 0.5 mg/L BAP + 5 mg/L elicitor | Increased by 2.25 folds | [65] |
| Chitosan | Hairy root culture | MS medium + 150 mg/L chitosan | Increased by 6 folds | [75] |
| Oligogalacturonides | Hairy root culture | MS medium + 0.01 mg/L gibberellic acid +60 g/mL elicitor | Increased by 55.2% | [70] |
| Heptakis (2,6-di-O-methyl)-β-cyclodextrin (DIMEB) and methyl jasmonate | Cell suspension culture | MS medium + 2.0 mg/L 2,4-D + 0.15 mg/L BAP + 50 mM DIMEB + 100 µM Methyl jasmonate | Increased by 300 folds (27 umol/g DW) | [68] |
| Methyl jasmonate and mevalonic acid lactone | Cell suspension culture | MS medium + 0.1 mg/L NAA + 0.1 mg/L KN + 50 mg/L mevalonic acid lactone + methyl jasmonate | Increased by 5.93 times | [52] |
| Sorbitol and Coronatine | Cell suspension culture | MS medium + 0.1 mg/L NAA + 0.1 mg/L KN + 30 g/L Sorbitol + 0.05 µM Coronatine | Increased by 8 folds | [72] |
| Culture Type | Type of Bioreactor | Culture Conditions | Yield | References |
|---|---|---|---|---|
| Hairy root culture | Modified nutrient mist bioreactor | ¼ MS + 10 µg/L GA3Batch culture having adequate oxygen supply and nutrient | 1.12 mg/g | [80] |
| Shoot culture | Mist bioreactor | MS + 0.05 mg/L NAA + 0.5 mg/L BAP; 25 days batch | 48.2 mg/L | [81] |
| Shoot culture | Mist nutrient bioreactor | MS + 0.05 mg/L NAA + 0.5 mg/L BAP; 25 days batch | 46.9 mg/L | [82] |
| Hairy root culture | Mist nutrient bioreactor | B5 medium; 1 min on/15 min off mist cycle | NA | [83] |
| Hairy root culture | Modified stirred tank bioreactor | ¼ MS + 10 µg/L GA3; Fed batch for 10–15 days | 0.99 mg/g DW | [84] |
| Hairy root culture | Stirred tank bioreactor | MS medium; 25-day batch. | 0.32 mg/g DW | [85] |
| Adventitious roots | Mist bioreactor | MS medium; 0.5 L/min air, 25-day culture | 46.9 mg DW/L | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Khayri, J.M.; Sudheer, W.N.; Lakshmaiah, V.V.; Mukherjee, E.; Nizam, A.; Thiruvengadam, M.; Nagella, P.; Alessa, F.M.; Al-Mssallem, M.Q.; Rezk, A.A.; et al. Biotechnological Approaches for Production of Artemisinin, an Anti-Malarial Drug from Artemisia annua L. Molecules 2022, 27, 3040. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27093040
Al-Khayri JM, Sudheer WN, Lakshmaiah VV, Mukherjee E, Nizam A, Thiruvengadam M, Nagella P, Alessa FM, Al-Mssallem MQ, Rezk AA, et al. Biotechnological Approaches for Production of Artemisinin, an Anti-Malarial Drug from Artemisia annua L. Molecules. 2022; 27(9):3040. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27093040
Chicago/Turabian StyleAl-Khayri, Jameel M., Wudali N. Sudheer, Vasantha V. Lakshmaiah, Epsita Mukherjee, Aatika Nizam, Muthu Thiruvengadam, Praveen Nagella, Fatima M. Alessa, Muneera Q. Al-Mssallem, Adel A. Rezk, and et al. 2022. "Biotechnological Approaches for Production of Artemisinin, an Anti-Malarial Drug from Artemisia annua L." Molecules 27, no. 9: 3040. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27093040