Use of Biotinylated Ubiquitin for Analysis of Rat Brain Mitochondrial Proteome and Interactome

 ,
,
Abstract
:1. Introduction
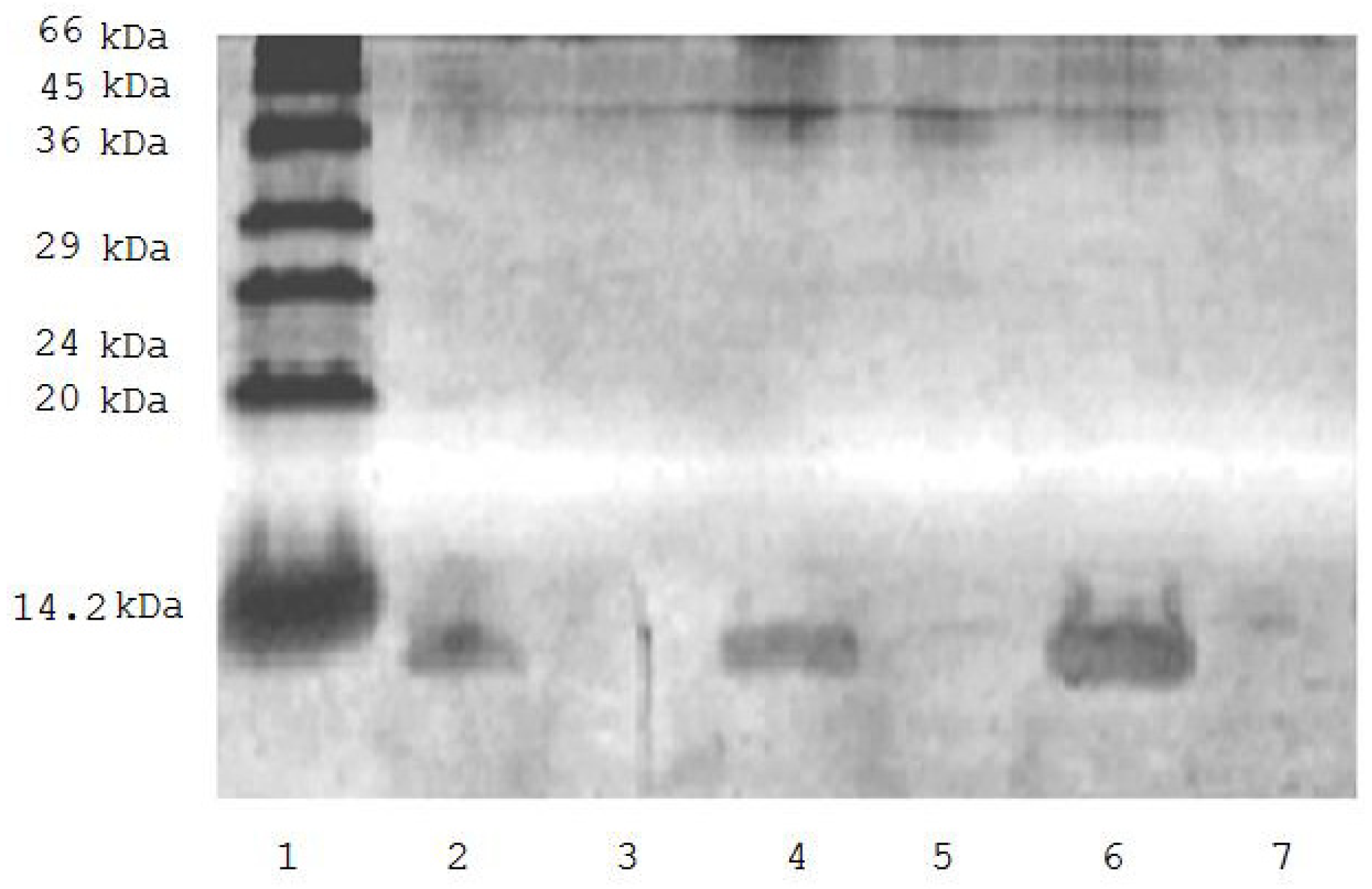
2. Results
3. Discussion
4. Experimental Section
4.1. Chemicals
4.2. Animals and Preparation of Brain Mitochondrial Fraction
4.3. Biotinylated Ubiquitin
4.4. SDS-PAGE
4.5. Sample Preparation
4.6. Avidin Agarose Chromatography
4.7. Proteomic Identification of Ubiquitinated Proteins
4.8. Identification of Ubiquitination Sites
4.9. Prediction of a Mitochondrial Interactome/Protein Interaction Network
5. Conclusions
Acknowledgments
References
- Hershko, A.; Ciechanover, A. The ubiquitin system. Annu. Rev. Biochem 1998, 67, 425–479. [Google Scholar]
- Schwartz, A.L.; Ciechanover, A. Targeting proteins for destruction by the ubiquitin system: Implications for human pathobiology. Annu. Rev. Pharmacol. Toxicol 2009, 49, 73–96. [Google Scholar]
- Ciechanover, A. The ubiquitin-mediated proteolytic pathway: Mechanisms of action and cellular physiology. Biol. Chem. Hoppe-Seyler 1994, 375, 565–581. [Google Scholar]
- Schwartz, A.L.; Ciechanover, A. Ubiquitin-mediated protein modification and degradation. Am. J. Respir. Cell Mol. Biol 1992, 7, 463–468. [Google Scholar]
- Wilkinson, K.D. Ubiquitination and deubiquitination: Targeting of proteins for degradation by the proteasome. Semin. Cell Dev. Biol 2000, 11, 141–148. [Google Scholar]
- Peng, J. Evaluation of proteomic strategies for analyzing ubiquitinated proteins. BMB Rep 2008, 41, 177–183. [Google Scholar]
- Ciechanover, A.; Orian, A.; Schwartz, A.L. Ubiquitin-mediated proteolysis: Biological regulation via destruction. Bioessays 2000, 22, 442–451. [Google Scholar]
- Hershko, A. The ubiquitin system for protein degradation and some of its roles in the control of the cell-division cycle (Nobel lecture). Angew. Chem Int. Ed 2005, 44, 5932–5943. [Google Scholar]
- Peng, J.; Schwartz, D.; Elias, J.E.; Thoreen, C.C.; Cheng, D.; Marsischky, G.; Roelofs, J.; Finley, D.; Gygi, S.P. A proteomics approach to understanding protein ubiquitination. Nat. Biotechnol 2003, 21, 921–926. [Google Scholar]
- Medvedev, A.E.; Kopylov, A.T.; Buneeva, O.A.; Zgoda, V.G.; Archakov, A.I. Affinity-based proteomic profiling: Problems and achievements. Proteomics 2012, 12, 621–637. [Google Scholar]
- Riederer, I.M.; Schiffrin, M.; Kövari, E.; Bouras, C.; Riederer, B.M. Ubiquitination and cysteine nitrosylation during aging and Alzheimer’s disease. Brain Res. Bull 2009, 80, 233–241. [Google Scholar]
- Shimada, Y.; Fukuda, T.; Aoki, K.; Yukawa, T. A protocol for immunoaffinity separation of the accumulated ubiquitin-protein conjugates solubilized with sodium dodecyl sulfate. Anal. Biochem 2008, 377, 77–82. [Google Scholar]
- Gitlin, G.; Bayer, E.A.; Wilchek, M. Studies on the biotin-binding site of avidin. Lysine residues involved in the active site. Biochem. J 1987, 242, 923–926. [Google Scholar]
- Magnani, M.; Serafini, G.; Antonelli, A.; Malatesta, M.; Gazzanelli, G. Evidence for a particulate location of ubiquitin conjugates and ubiquitin-conjugating enzymes in rabbit brain. J. Biol. Chem 1991, 266, 21018–21024. [Google Scholar]
- Kemeny, S.; Dery, D.; Loboda, Y.; Rovner, M.; Lev, T.; Zuri, D.; Finberg, J.M.P.; Larisch, S. Parkin Promotes Degradation of the Mitochondrial Pro-Apoptotic ARTS Protein. PLoS One 2012, 7, e38837. [Google Scholar]
- Buneeva, O.A.; Medvedeva, M.V.; Medvedev, A.E. Incorporation of ubiquitin into rat brain mitochondria is accompanied by increased proteolytic digestibility of MAO. Neurobiology 1999, 7, 257–261. [Google Scholar]
- Buneeva, O.A.; Medvedeva, M.V.; Medvedev, A.E. Ubiquitin and proteolytic degradation of monoamine oxidases. Biochem. (Moscow) Suppl. Ser. B Biomed. Chem 2008, 2, 101–104. [Google Scholar]
- Weekes, J.; Morrison, K.; Mullen, A.; Wait, R.; Barton, P.; Dunn, M.J. Hyperubiquitination of proteins in dilated cardiomyopathy. Proteomics 2003, 3, 208–216. [Google Scholar]
- Jeon, H.B.; Choi, E.S.; Yoon, J.H.; Hwang, J.H.; Chang, J.W.; Lee, E.K.; Choi, H.W; Park, Z.Y.; Yoo, Y.J. A proteomics approach to identify the ubiquitinated proteins in mouse heart. Biochem. Biophys. Res. Comm. 2007, 357, 731–736. [Google Scholar]
- Sullivan, P.G.; Dragicevic, N.B.; Deng, J.H.; Bai, Y.; Dimayuga, E.; Ding, Q.; Chen, Q.; Bruce-Keller, A.J.; Keller, J.N. Proteasome inhibition alters neural mitochondrial homeostasis and mitochondria turnover. J. Biol. Chem 2004, 279, 20699–20707. [Google Scholar]
- Nakamura, N.; Hirose, S. Regulation of mitochondrial morphology by USP30, a deubiquitinating enzyme present in the mitochondrial outer membrane. Mol. Biol. Cell 2008, 19, 1903–1911. [Google Scholar]
- Cohen, M.M.; Leboucher, G.P.; Livnat-Levanon, G.; Glickman, M.H.; Weissman, A.M. Ubiquitin-proteasome dependent degradation of a mitofusin, a critical regulator of mitochondrial fusion. Mol. Biol. Cell 2008, 19, 2457–2464. [Google Scholar]
- Gagnon, P. Technology trends in antibody purification. J. Chrom. A 2012, 1221, 57–70. [Google Scholar]
- Takuma, T.; Arakawa, T.; Tajima, Y. Interaction of SNARE proteins in rat parotid acinar cells. Arch. Oral. Biol 2000, 45, 369–375. [Google Scholar]
- Jacobsson, G.; Meister, B. Molecular components of the exocytotic machinery in the rat pituitary gland. Endocrinology 1996, 137, 5344–5356. [Google Scholar]
- Nakahara, T.; Nakamura, K.; Tsutsumi, T.; Hashimoto, K.; Hondo, H.; Hisatomi, S.; Motomura, K.; Uchimura, H. Effect of chronic haloperidol treatment on synaptic protein mRNAs in the rat brain. Mol. Brain Res 1998, 61, 238–242. [Google Scholar]
- Cho, W.J.; Jeremic, A.; Jena, B.P. Direct interaction between SNAP-23 and l-type Ca2+ channel. J. Cell Mol. Med 2005, 9, 380–386. [Google Scholar]
- Abonyo, B.O.; Gou, D.; Wang, P.; Narasaraju, T.; Wang, Z.; Liu, L. Syntaxin 2 and SNAP-23 are required for regulated surfactant secretion. Biochemistry 2004, 43, 3499–3506. [Google Scholar]
- Kanno, E.; Fukuda, M.J. Increased plasma membrane localization of O-glycosylation-deficient mutant of synaptotagmin I in PC12 cells. J. Neurosci Res 2008, 86, 1036–1043. [Google Scholar]
- Fukuda, M.J. Vesicle-associated membrane protein-2/synaptobrevin binding to synaptotagmin I promotes O-glycosylation of synaptotagmin I. Biol. Chem 2002, 277, 30351–30358. [Google Scholar]
- Thomas, S.G.; Takahashi, M.; Neill, J.D.; Clarke, I.J. Components of the neuronal exocytotic machinery in the anterior pituitary of the ovariectomised ewe and the effects of oestrogen in gonadotropes as studied with confocal microscopy. Neuroendocrinology 1998, 67, 244–259. [Google Scholar]
- Risinger, C.; Bennett, M.K. Differential phosphorylation of syntaxin and synaptosome-associated protein of 25 kDa (SNAP-25) isoforms. J. Neurochem 1999, 72, 614–624. [Google Scholar]
- Martínez-López, I.; García, C.; Barber, T.; Viña, J.R.; Miralles, V.J. The l-glutamate transporters GLAST (EAAT1) and GLT-1 (EAAT2): Expression and regulation in rat lactating mammary gland. Mol. Membr. Biol 1998, 15, 237–242. [Google Scholar]
- Schlag, B.D.; Vondrasek, J.R.; Munir, M.; Kalandadze, A.; Zelenaia, O.A.; Rothstein, J.D.; Robinson, M.B. Regulation of the glial Na+-dependent glutamate transporters by cyclic AMP analogs and neurons. Mol. Pharmacol 1998, 53, 355–369. [Google Scholar]
- Swanson, R.A.; Liu, J.; Miller, J.W.; Rothstein, J.D.; Farrell, K.; Stein, B.A.; Longuemare, M.C. Neuronal regulation of glutamate transporter subtype expression in astrocytes. J. Neurosci 1997, 17, 932–940. [Google Scholar]
- Boyer, S.; Maunoury, R.; Gomès, D.; de Néchaud, B.; Hill, A.M.; Dupouey, P. Expression of glial fibrillary acidic protein and vimentin in mouse lens epithelial cells during development in vivo and during proliferation and differentiation in vitro: Comparison with the developmental appearance of GFAP in the mouse central nervous system. J. Neurosci. Res. 1990, 27, 55–64. [Google Scholar]
- Verderber, L.; Johnson, W.; Mucke, L.; Sarthy, V. Differential regulation of a glial fibrillary acidic protein-LacZ transgene in retinal astrocytes and Müller cells. Invest. Ophthalmol. Vis. Sci 1995, 36, 1137–1143. [Google Scholar]
- Yeh, C.Y.; Vadhwana, B.; Verkhratsky, A.; Rodríguez, J.J. Early astrocytic atrophy in the entorhinal cortex of a triple transgenic animal model of Alzheimer’s disease. ASN Neuro 2011, 3, 271–279. [Google Scholar]
- Salmaso, N.; Cossette, M.P.; Woodside, B. Pregnancy and maternal behavior induce changes in glia, glutamate and its metabolism within the cingulate cortex. PLoS One 2011, 6, e23529. [Google Scholar]
- Ardais, A.P.; Viola, G.G.; Costa, M.S.; Nunes, F.; Behr, G.A.; Klamt, F.; Moreira, J.C.; Souza, D.O.; Rocha, J.B.; Porciúncula, L.O. Acute treatment with diphenyl diselenide inhibits glutamate uptake into rat hippocampal slices and modifies glutamate transporters, SNAP-25, and GFAP immunocontent. Toxicol. Sci 2010, 113, 434–443. [Google Scholar]
- Cassano, T.; Serviddio, G.; Gaetani, S.; Romano, A.; Dipasquale, P.; Cianci, S.; Bellanti, F.; Laconca, L.; Romano, A.D.; Padalino, I; et al. Glutamatergic alterations and mitochondrial impairment in a murine model of Alzheimer disease. Neurobiol. Aging 2012, 33, 1121.e1–1121.e12. [Google Scholar]
- Medvedev, A.E.; Gorkin, V.Z. Endogenous stimulation of lipid peroxidation in brain increases proteolytic inactivation of mitochondrial monoamine oxidases. Int. J. Dev. Neurosci 1994, 12, 151–155. [Google Scholar]
- Kantor, A. Biotinylation of Antibodies protocol. Available online: http://www.drmr.com/abcon/Biotin.html accessed on 7 July 2004.
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]
- Spector, T. Refinement of the Coomassie Blue method of protein quantitation. Anal. Biochem 1978, 86, 142–146. [Google Scholar]
- Walker, J.M. The protein Protocols Handbook; Humana Press Inc: Totowa, NY, USA, 2002; p. 1176. [Google Scholar]
- Kapp, E.A.; Schultz, F.; Conolly, F.M.; Chakel, J.A.; Meza, J.E.; Miller, C.A.; Fenyo, D.; Eng, J.K.; Adkins, J.N.; Omenn, G.S.; et al. An evaluation, comparison, and accurate benchmarking of several publicly available MS/MS search algorithms: sensitivity and specificity analysis. Proteomics 2005, 5, 3475–3490. [Google Scholar]
- Jensen, L.J.; Kuhn, M.; Stark, M.; Chaffron, S.; Creevey, C.; Muller, J.; Doerks, T.; Julien, P.; Roth, A.; Simonovic, M.; et al. STRING 8—A global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res 2009, 37, D412–D416. [Google Scholar]
- Merbl, Y.; Kirschner, M.W. Large-scale detection of ubiquitination substrates using cell extracts and protein microarrays. Proc. Natl. Acad. Sci. USA 2009, 106, 2543–2548. [Google Scholar]
- Hjerpe, R.; Rodríguez, M.S. Efficient approaches for characterizing ubiquitinated proteins. Biochem. Soc. Trans 2008, 36, 823–827. [Google Scholar]
- Correia, M.A.; Sadeghi, S.; Mundo-Paredes, E. Cytochrome P450 ubiquitination: branding for the proteolytic slaughter? Annu. Rev. Pharmacol. Toxicol 2005, 45, 439–464. [Google Scholar]
- Merbl, Y.; Kirschner, M.W. Large-scale detection of ubiquitination substrates using cell extracts and protein microarrays. Proc. Natl. Acad. Sci. USA 2009, 106, 2543–2548. [Google Scholar]



{kind=link}
{kind=link}
| No. | Protein name | Database Accession Number | Sequence coverage (%) | Search score | Localization |
|---|---|---|---|---|---|
| Proteins/enzymes involved in energy generation and carbohydrate metabolism (n = 13) | |||||
| 1 | Pyruvate carboxylase, mitochondrial precursor | P52873 | 25 | 285.19 | M |
| 2 | ATP synthase subunit beta, mitochondrial precursor | P10719 | 47 | 231.07 | M |
| 3 | ATP synthase subunit alpha, mitochondrial precursor | P15999 | 30 | 163.32 | M |
| 4 | Sodium/potassium-transporting ATP-ase subunit alpha-3 | P06687 | 13 | 130.82 | PM |
| 5 | Glyceraldehyde-3-phosphate dehydrogenase | P04797 | 14 | 48.89 | EM |
| 6 | ADP/ATP translocase 2 | Q09073 | 11 | 42.59 | M |
| 7 | Creatine kinase M-type | P00554 | 12 | 56.1 | M |
| 8 | Sodium/potassium-transporting ATP-ase subunit alpha-2 precursor | P06686 | 5 | 37.89 | PM |
| 9 | ATP synthase subunit O, mitochondrial precursor | Q06647 | 11 | 34.75 | M |
| 10 | Sodium/potassium-transporting ATP-ase subunit alpha-1, precursor | P06685 | 2 | 21,46 | PM |
| 11 | ATP synthase subunit d, mitochondrial | P31399 | 10 | 16.97 | M |
| 12 | Pyruvate dehydrogenase E1 component subunit beta, mitochondrial precursor | P49432 | 4 | 15.32 | M |
| 13 | Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex, mitochondrial precursor | P08461 | 2 | 14.02 | M |
| Proteins involved in cytoskeleton formation and exocytosis (n = 24) | |||||
| 1 | Keratin type I cytoskeletal 10 | Q6IFW6 | 19 | 176.37 | EM |
| 2 | Keratin type II cytoskeletal 5 | Q6P6Q2 | 14 | 165.65 | EM |
| 3 | Keratin type II cytoskeletal 1 | Q6IMF3 | 9 | 124.82 | EM |
| 4 | Keratin type I cytoskeletal 42 | Q6IFU7 | 16 | 118.43 | EM |
| 5 | Keratin type II cytoskeletal 6A | Q4FZU2 | 13 | 115.79 | EM |
| 6 | Tubulin alpha-1B chain | Q6P9V9 | 27 | 111.39 | EM |
| 7 | Keratin type I cytoskeletal 14 | Q6IFV1 | 9 | 80.83 | EM |
| 8 | Neurofilament light polypeptide | P19527 | 13 | 71.54 | EM |
| 9 | Tubulin beta-2B chain | Q3KRE8 | 12 | 62,92 | EM |
| 10 | Tubulin beta-2A chain | Q85108 | 12 | 62,92 | EM |
| 11 | Glial fibrillary acidic protein | P47819 | 14 | 57.15 | EM |
| 12 | Tubulin beta-5 chain | P69897 | 12 | 54.58 | EM |
| 13 | Tubulin beta-3 chain | Q4QRB4 | 10 | 48.33 | EM |
| 14 | Myosin-4 | Q29RW1 | 2 | 45.07 | EM |
| 15 | Alpha-internexin | P23565 | 11 | 43.84 | EM |
| 16 | Actin, alpha skeletal muscle | P68136 | 11 | 43.17 | EM |
| 17 | Tubulin alpha-1B chain | Q6P9V9 | 9 | 39.35 | EM |
| 18 | Tubulin alpha-1A chain | P68370 | 9 | 39.35 | EM |
| 19 | Myosin-6 | P02563 | 2 | 37.23 | EM |
| 20 | Neurofilament medium polypeptide | P12839 | 5 | 35.80 | EM |
| 21 | Myosin-3 | P12847 | 1 | 34.23 | EM |
| 22 | Syntaxin-1B | P61265 | 10 | 26.19 | EM |
| 23 | Synaptotagmin-1 | P21707 | 3 | 18.57 | EM |
| 24 | Spectrin alpha chain, brain | P16086 | 1 | 17.02 | EM |
| Protective proteins (n = 1) | |||||
| 1 | 60 kDa heat shock protein, mitochondrial precursor | P63039 | 9 | 29.87 | M |
| Enzymes involved in signal transduction and regulation of enzyme activity (n = 3) | |||||
| 1 | Calcium/calmodulin-dependent protein kinase | P11275 | 7 | 39.65 | EM |
| 2 | 2′,3′-cyclic-nucleotide 3′-phosphodiesterase | P13233 | 8 | 46.16 | EM |
| 3 | Pro-associated protein kinase 2 | Q62868 | 1 | 19.53 | EM |
| Enzymes involved in fatty acid metabolism (n = 2) | |||||
| 1 | Propionyl-CoA carboxylase beta chain, mitochondrial precursor | P07633 | 13 | 67.94 | M |
| 2 | Propionyl-CoA carboxylase alpha chain, mitochondrial precursor | P14882 | 5 | 37.33 | M |
| Transporters (n = 4) | |||||
| 1 | Excitatory amino acid transporter 1 | P24942 | 6 | 34.57 | PM |
| 2 | Excitatory amino acid transporter 2 | P31596 | 2 | 26.35 | PM |
| 3 | Glial high affinity glutamate transporter isoform a | G3V6R2 | 2 | 25.7 | PM |
| 4 | sodium-dependent high affinity glutamate transporter GLT-1A | G3V6R0 | 2 | 26.0 | PM |
| Ubiquitin and related proteins (n = 3) | |||||
| polyubiquitin | P0CG51 | 51 | 54.99 | EM/M ? | |
| ubiquitin C | Q63429 | 4 | 44.01 | EM/M ? | |
| Ubc protein | Q5FWT0 | 10 | 44.01 | EM/M ? | |
| No | Protein Name | Database Accession Number | Sequence Coverage (%) | Search Score |
|---|---|---|---|---|
| 1 | rCG50775 | EDL86881 | 11 | 102.68 |
| 2 | type I keratin KA17 | Q6IFU8 | 14 | 88.22 |
| 3 | cytokeratin-8 | Q80WH8 | 9 | 86.42 |
| 4 | keratin complex 2, basic, gene 8 | Q10758 | 9 | 86.42 |
| 5 | hypothetical protein LOC683313 | Q63282 | 11 | 85.15 |
| 6 | rCG50690 | EDL86882 | 27 | 76.25 |
| 7 | Keratin, type II cytoskeletal 75 | Q6IG05 | 11 | 71.38 |
| 8 | keratin complex 1, acidic, gene 14 | NP_001008751 | 8 | 63.05 |
| 9 | type I keratin KA22 | Q6IFU7 | 9 | 62.98 |
| 10 | similar to keratin complex 2, basic, gene 6a isoform 1 | NP_001094477 | 7 | 61.64 |
| 11 | rCG50520 | EDL86873 | 4 | 60.01 |
| 12 | rCG23467, isoform CRA a | G3V8B0 | 2 | 59.00 |
| 13 | rCG23609, isoform CRA b | EDM14209 | 2 | 59.00 |
| 14 | type II keratin Kb15 | Q6IFZ4 | 7 | 56.92 |
| 15 | type I keratin KA15 | Q6IFV3 | 8 | 56.63 |
| 16 | type II keratin Kb39 | Q6IG01 | 4 | 56.21 |
| 17 | rCG34382, isoform CRA_a | G3V6D8 | 2 | 56.00 |
| 18 | type I keratin KA16 | Q6IFU9 | 9 | 55.18 |
| 19 | rCG33578 | EDM06024 | 12 | 52.72 |
| 20 | type II keratin Kb36 | Q6IG03 | 6 | 46.15 |
| 21 | type II keratin Kb1 | Q6IMF3 | 7 | 45.54 |
| 22 | type I keratin KA11 | Q6IFV0 | 5 | 45.00 |
| 23 | Krt1–19 protein | Q63279 | 5 | 41.32 |
| 24 | keratin complex 1, acidic, gene 19, isoform CRA b | EDM06018 | 8 | 41.32 |
| 25 | rCG34382, isoform CRA b | EDM04783 | 2 | 40.63 |
| 26 | rCG33626 | EDM04786 | 2 | 37.50 |
| 27 | myelin basic protein, isoform CRA a | Q63327 | 10 | 34.60 |
| 28 | rCG23467, isoform CRA b | EDM14212 | 2 | 29.34 |
| 29 | PREDICTED: similar to type II keratin Kb36 | NP_001008808 | 6 | 28.42 |
| 30 | keratin complex 1, acidic, gene 19, isoform CRA c | EDM06019 | 6 | 27.10 |
| 31 | myelin basic protein isoform 1 | P02688-1 | 10 | 27.03 |
| 32 | myelin basic protein | Q80Z98 | 10 | 27.03 |
| 33 | myelin basic protein isoform 2 | P02688-2 | 12 | 27.03 |
| 34 | myelin basic protein isoform 4 | P02688-4 | 13 | 27.03 |
| 35 | rCG34505 | EDM05987 | 2 | 25.35 |
| 36 | myelin basic protein, isoform CRA d | Q5XFW | 7 | 23.60 |
| 37 | myelin basic protein, isoform CRA c | G5E945 | 8 | 23.60 |
| 38 | keratin 13 | Q6IFV4 | 3 | 23.44 |
| 39 | rCG33575 | EDM04790 | 2 | 22.89 |
| 40 | proteolipid protein | P63081 | 7 | 20.85 |
| 41 | type II keratin Kb2 | Q6IG02 | 2 | 20.30 |
| 42 | Tubulin, alpha 1A | P68370 | 5 | 19.83 |
| 43 | tubulin, alpha 1C | Q6AYZ1 | 5 | 19.83 |
| 44 | rCG50513, isoform CRA b | EDL87016 | 7 | 19.83 |
| 45 | rCG50513, isoform CRA a | EDL87015 | 7 | 19.83 |
| 46 | keratin complex 1, acidic, gene 12 | Q5BJY9 | 1 | 17.88 |
| 47 | rCG34869 | EDM05985 | 3 | 17.88 |
| 48 | keratin 19 | Q63279 | 3 | 17.88 |
| 49 | type II keratin Kb9 | Q6IFZ5 | 3 | 17.11 |
| 50 | rCG62531, isoform CRA d | EDM00590 | 9 | 16.42 |
| 51 | myelin basic protein isoform 3 | P02688-3 | 6 | 16.03 |
| 52 | myelin basic protein isoform 5 | P02688-5 | 7 | 16.03 |
| 53 | rCG44184, isoform CRA a | EDL98317 | 4 | 15.98 |
| 54 | rCG29914, isoform CRA b | EDM01924 | 5 | 15.23 |
| 55 | Tpi 1 protein | P48500 | 6 | 15.23 |
| 56 | rCG29914, isoform CRA a | EDM01923 | 6 | 15.23 |
| 57 | rCG47746, isoform CRA b | EDM13157 | 13 | 14.99 |
| 58 | keratin 20 | P25030 | 2 | 14.22 |
| 59 | rCG33887, isoform CRA a | EDM06013 | 14 | 14.22 |
| No | Protein Name | Database Accession Number | Acronym name | No. of peptides | Sequence Coverage (%) | Search score | Sequence/position of a.a. residue/* |
|---|---|---|---|---|---|---|---|
| 1 | Pyruvate carboxylase, mitochondrial precursor | P52873 | Pc | 18 | 25 | 285.19 | SGEGMGIRLDNASAFQGAVISPHYDSLLVKVIAHG KDHPTAATKMSR/K442/EVKKAYVEANQMLGDLIKVTPSSKIVGDLAQFMV QNGLSR/K891/SSTAPVASPNVRRLEYKPIKKVMVANR/K35/ |
| 2 | ATP synthase subunit alpha, mitochondrial precursor | P15999 | Atp5a1 | 11 | 30 | 163.32 | ELIIGDRQTGKTSIAIDTIINQK/K218 VGSAAQTRAmKQVAGTMKLELAQYREVAAFAQF GSDLDAATQQLLSR/K434/VVDALGNAIDGKGPVGSKIR/K161/AMKQVAGTMKLELAQYREVAAFAQFGSDLDAAT QQLLSR/K427/ |
| 3 | Sodium/potassiumtransporting ATP-ase subunit alpha-3 | P06687 | Atp1a3 | 9 | 13 | 130.82 | EKLENMKKEMEMNDHQLSVSELEQKYQTSATK/K 63/RDTAGDASESALLKCIELSCGSVRKMR/K456/GFAFDCDDVNFTTDNLCFVGLMSMIDPPRAAVPDA VGKCR/K595/ |
| 4 | Keratin type I cytoskeletal 10 | Q6IFW6 | Krt10 | 14 | 19 | 176.37 | NELTEMKRTLQTLEIELQSLLAMK/K298/TIEDLKSKILAATVDNANVLLQIDNAR/K182/ |
| 5 | Keratin type II cytoskeletal 5 | Q6P6Q2 | Krt5 | 20 | 14 | 165.65 | IKTQEREQIKTLNNKFASFIDK/K172/EYQELMNTKLALDVEIATYR/K456/NKYEDEINKRTEmENEFVLIKK/K185/ALNNKFASFIDKVRFLEQQNQVLETKWELLQQLDQ NNSR/K165/ |
| 6 | Keratin type II cytoskeletal 1 | Q6IMF3 | Krt1 | 9 | 9 | 124.82 | LALDMEIATYRKLLEGEEIRMSGECTPNVSVSVSTS HTSMSGTSSR/K483/MLREHQELMSMKLALDIEIATYR/K423/TTAENEFVVLKKDVDAAYMSKVELQAKVDALDGE IK/K139/ |
| 7 | Myosin-4 | Q29RW1 | Myh1 | 4 | 2 | 45.07 | NTQGMLKDTQLHLDDALRGQDDLKEQLAMVERR/K1655/IKKKMEGDLNEMEIQLNHANRQAAEAIR/K1619/SGLYKLTGAVMHYGNMKFKQK/K366/KLEGDLKLAQESTMDIENDKQQLDEKLK/K1083/AFMGVKNWPWMKLYFKIKPLLK/K838/FQLEAKIKEVTERAEDEEEINAELTAK/K919/KMEGDLNEMEIQLSHANRMAAEAQKQVKEAR/K1 644/GKAEAHFSLVHYAGTVDYNIIGWLDKNK/K599/ |
| 8 | Actin, alpha skeletal muscle | P68136 | Acta2 | 3 | 11 | 43.17 | KDLYANNVMSGGTTMYPGIADRMQKEITALAPST MK/K293/AVFPSIVGRPRHQGVMVGMGQKDSYVGDEAQSKR/K50/ |
| 9 | Myosin-6 | P02563 | Myh6 | 3 | 2 | 37.23 | QREEQAEPDGTEDADKSAYLMGLNSADLLK/K383/ |
| 10 | Spectrin alpha chain, brain | P16086 | Spna2 | 2 | 1 | 17.02 | MQHNLEQQIQARNTTGVTEEALKEFSMMFKHFDK/K2333/SCKKFMLFREANELQQWINEKEAALTSEEVGADLE QVEVLQK/K1110/DLNSQADSLMTSSAFDTSQVKEKR/K1738/ |
| 11 | 2′,3′-cyclic-nucleotide 3′-phosphodiesterase | P13233 | Cpn | 3 | 8 | 46.16 | GKLYSLGKGRWMLSLAKK/K370/ |
| 12 | Excitatory amino acid transporter 2 | P31596 | Slc1a2 | 2 | 2 | 26.35 | MASTEGANNMPKQVEVRMHDSHLSSEEPKHRNLGMR/K12/ |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Buneeva, O.A.; Medvedeva, M.V.; Kopylov, A.T.; Zgoda, V.G.; Medvedev, A.E. Use of Biotinylated Ubiquitin for Analysis of Rat Brain Mitochondrial Proteome and Interactome. Int. J. Mol. Sci. 2012, 13, 11593-11609. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms130911593
Buneeva OA, Medvedeva MV, Kopylov AT, Zgoda VG, Medvedev AE. Use of Biotinylated Ubiquitin for Analysis of Rat Brain Mitochondrial Proteome and Interactome. International Journal of Molecular Sciences. 2012; 13(9):11593-11609. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms130911593
Chicago/Turabian StyleBuneeva, Olga A., Marina V. Medvedeva, Arthur T. Kopylov, Victor G. Zgoda, and Alexei E. Medvedev. 2012. "Use of Biotinylated Ubiquitin for Analysis of Rat Brain Mitochondrial Proteome and Interactome" International Journal of Molecular Sciences 13, no. 9: 11593-11609. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms130911593