Anti-Proliferative Activities and Apoptosis Induction by Triterpenes Derived from Eriobotrya japonica in Human Leukemia Cell Lines

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
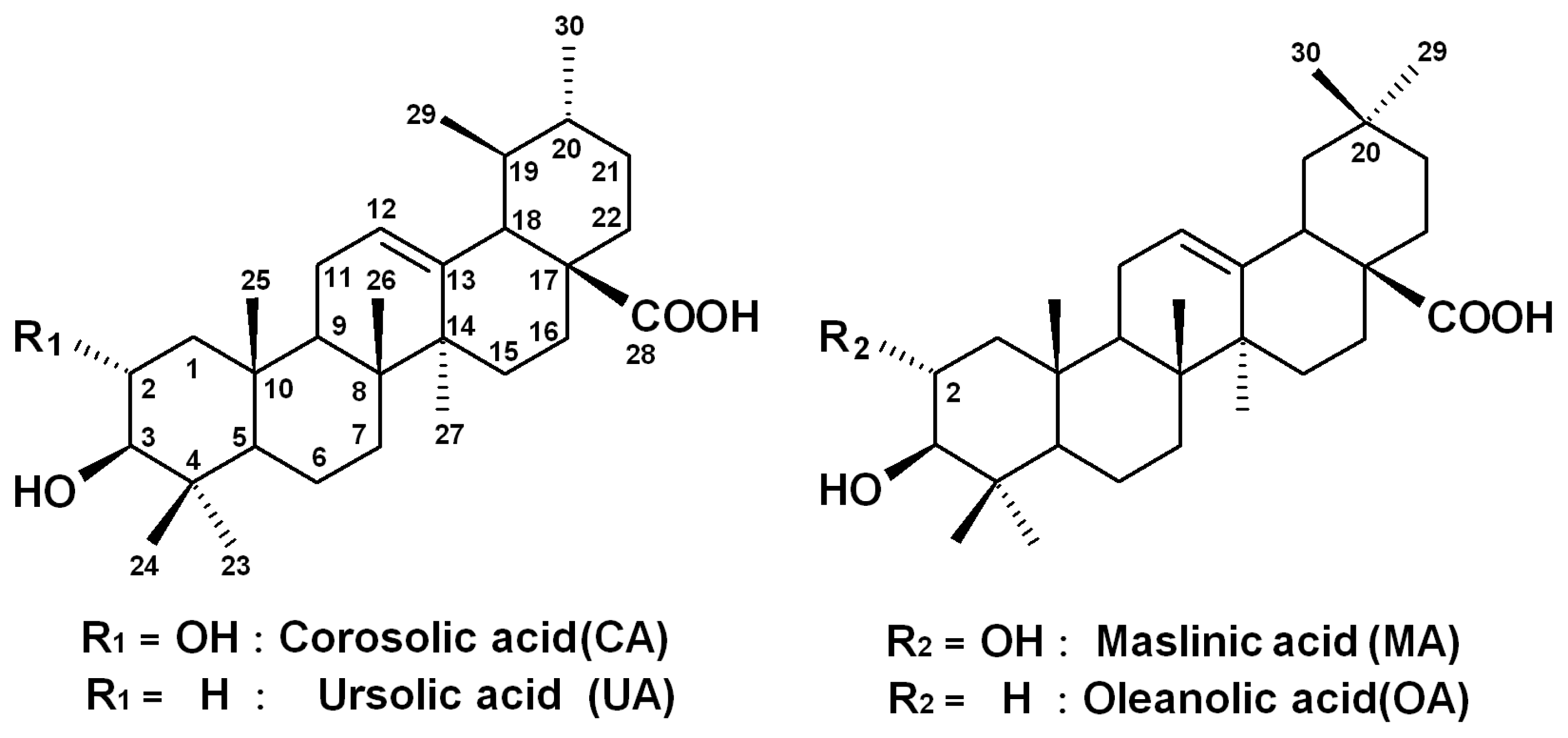
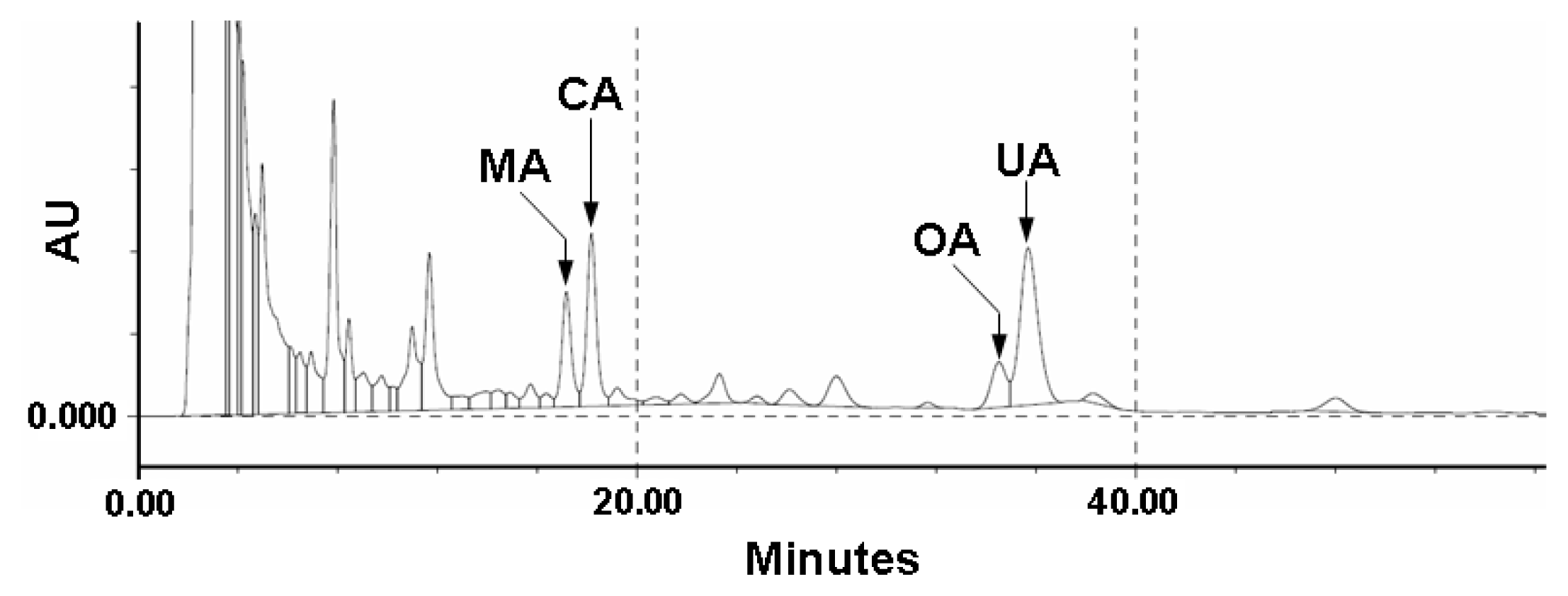
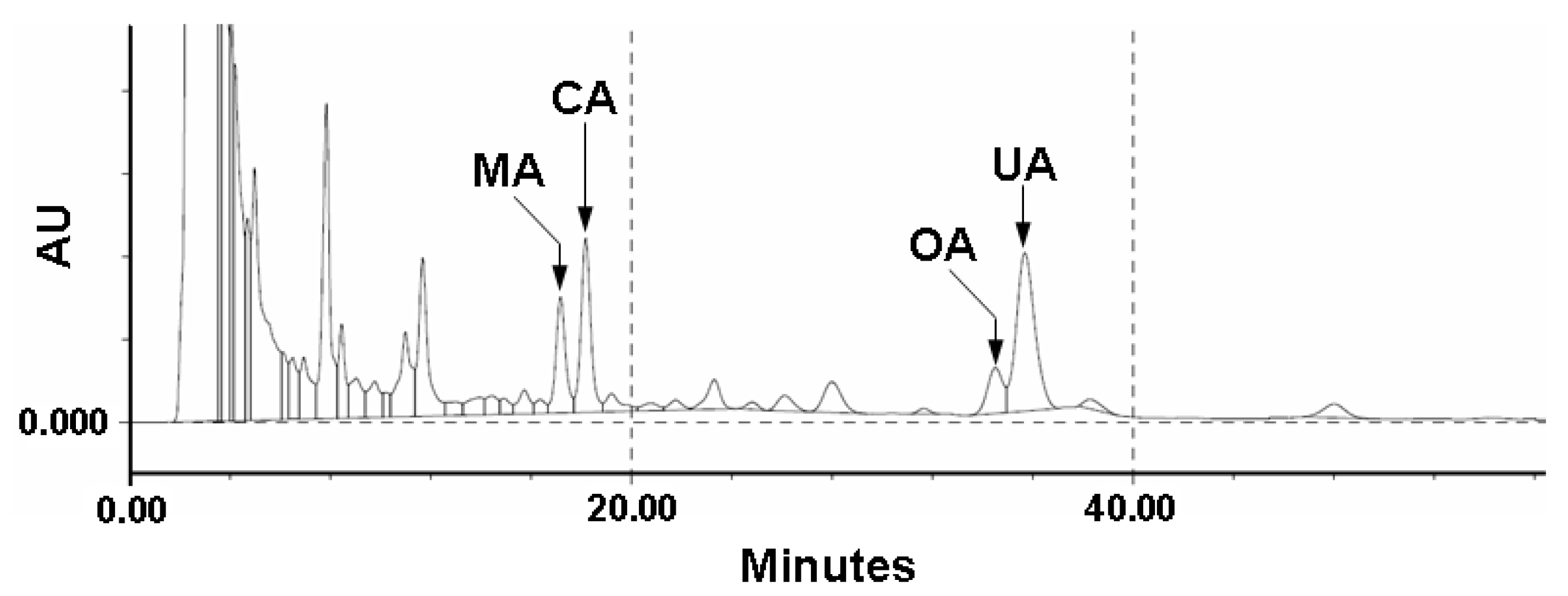
2.1. Quality Control of E. japonica Leaves

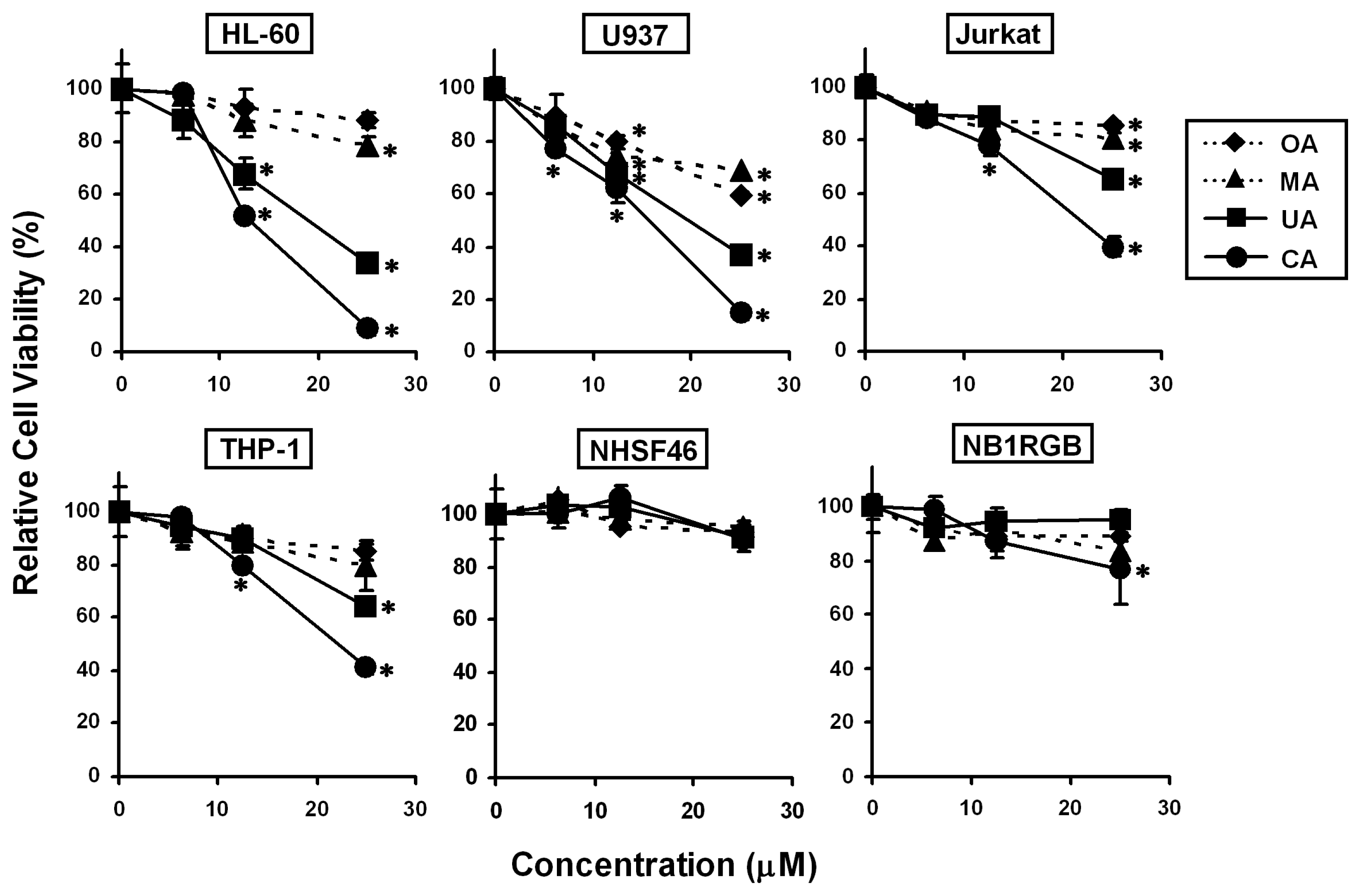
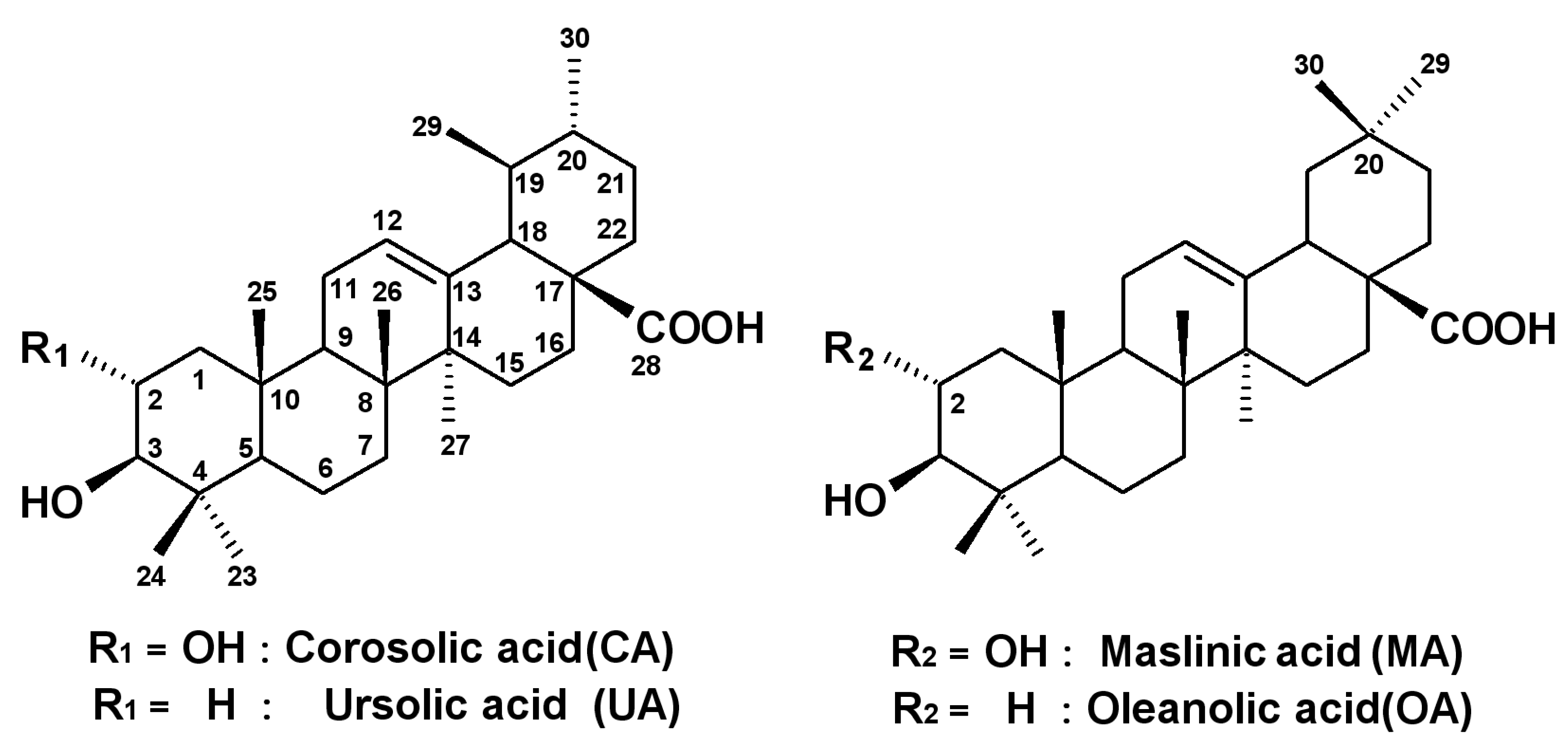
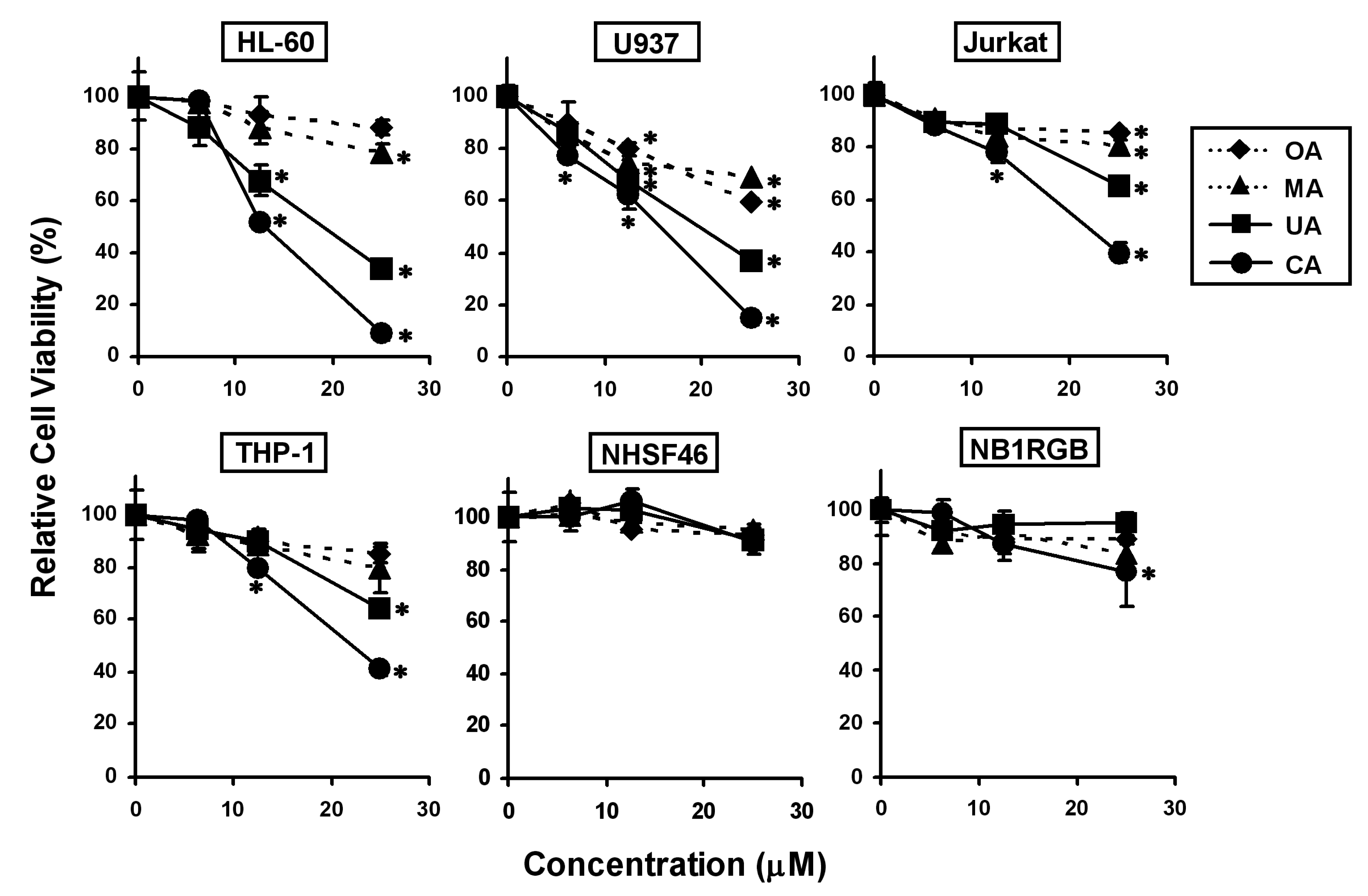
2.2. Effects of the Four Triterpenes from E. japonica Leaves on Cell Proliferation in Human Leukemia and Normal Skin Fibroblast Cell Lines and Their Structure-Activity Correlation
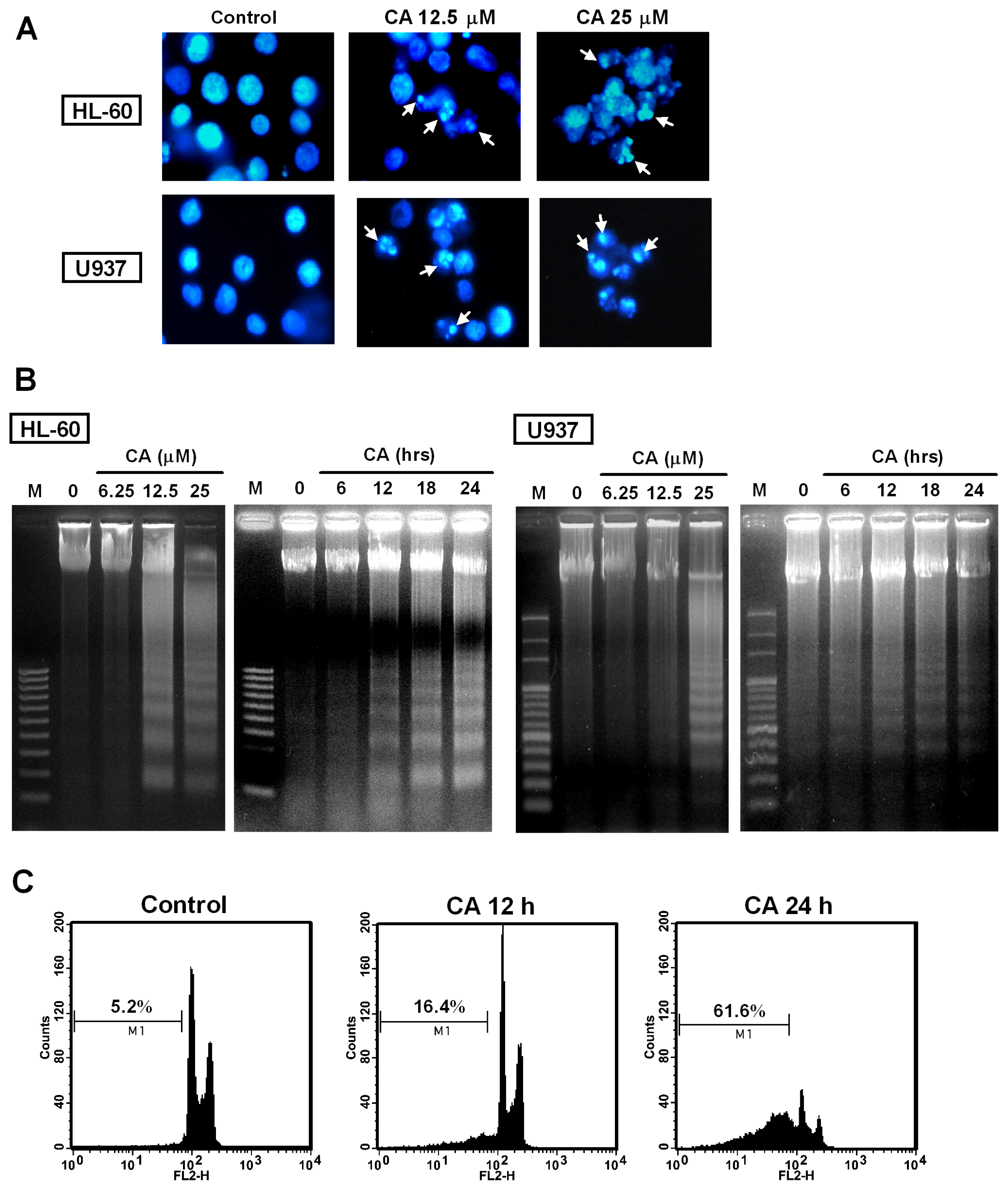
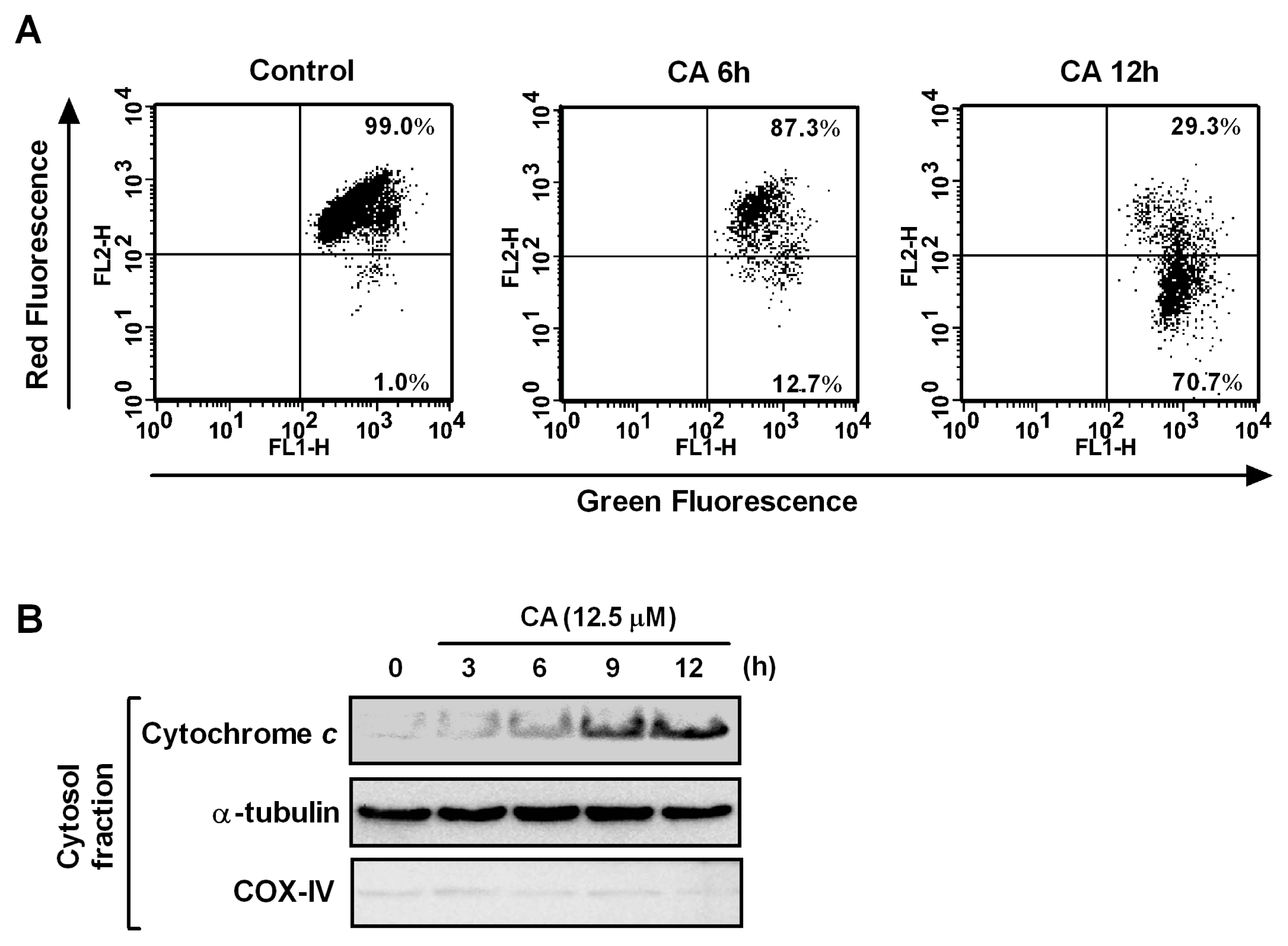
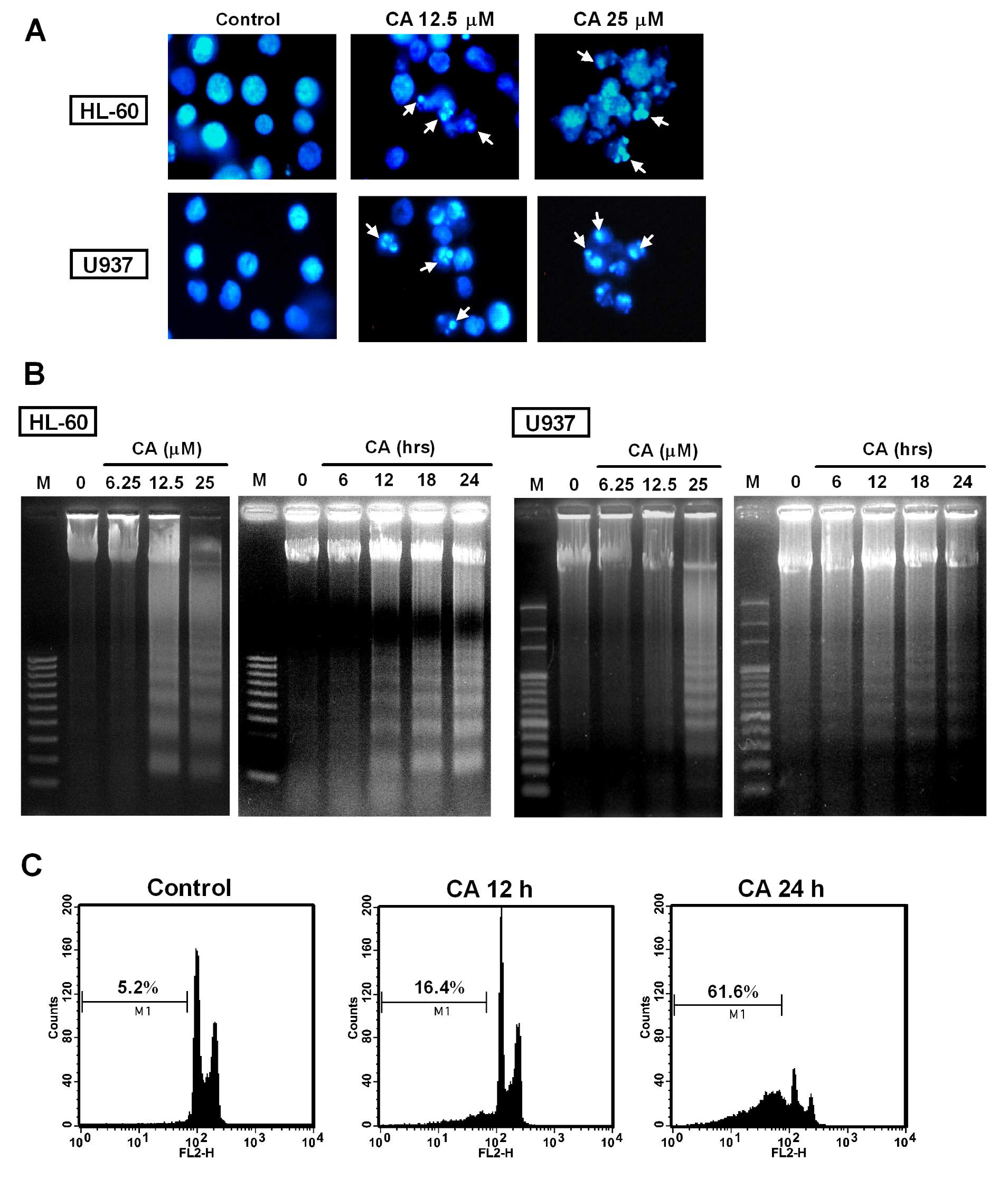
2.3. Effect of CA on Apoptosis Induction

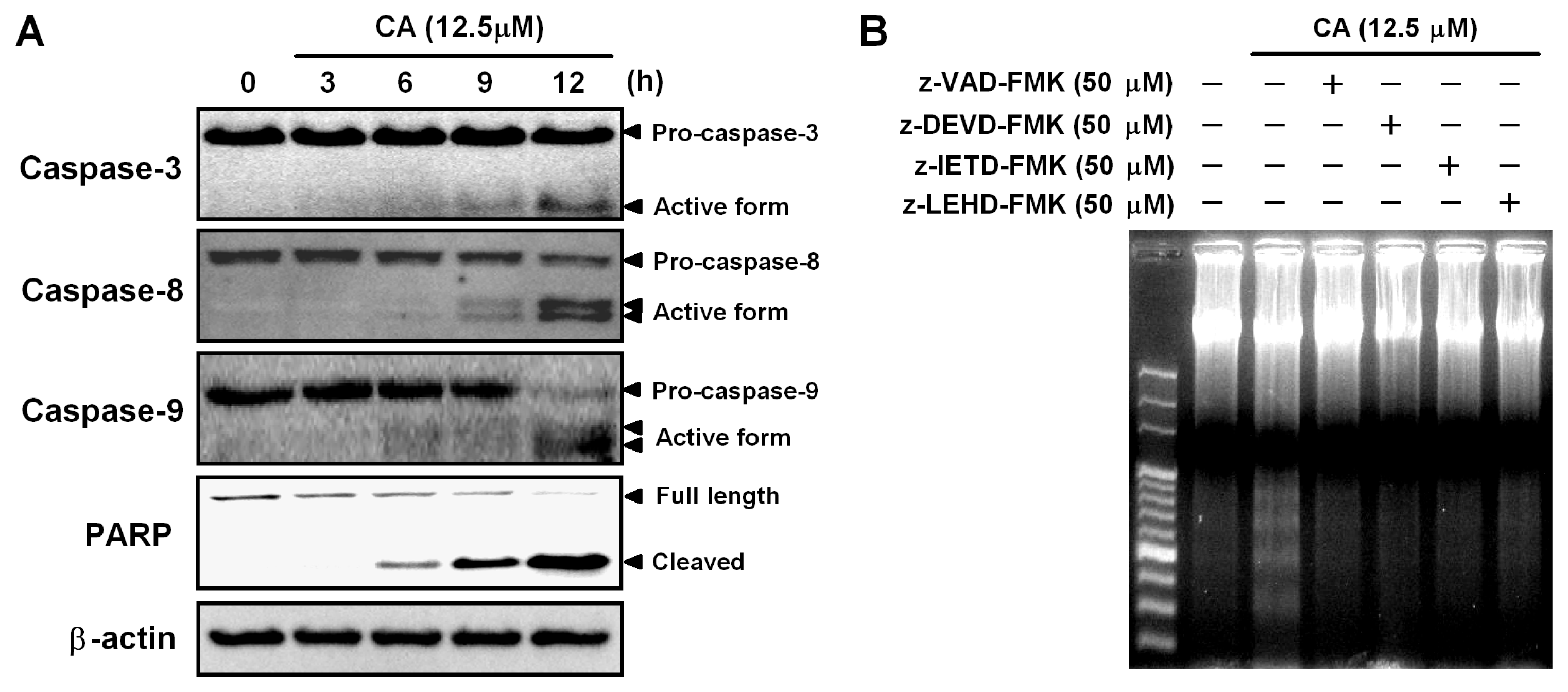
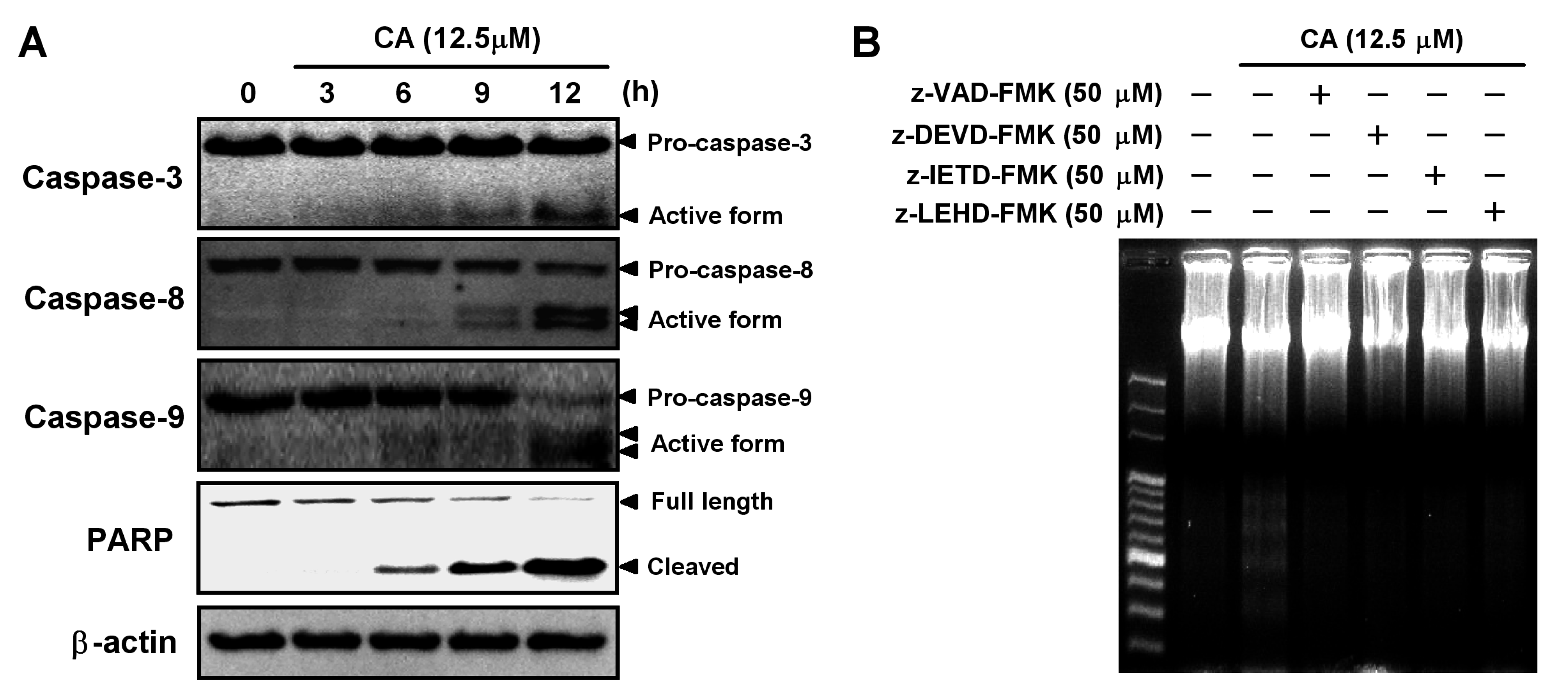
2.4. Involvement of the Caspase Cascade in CA-Induced Apoptosis


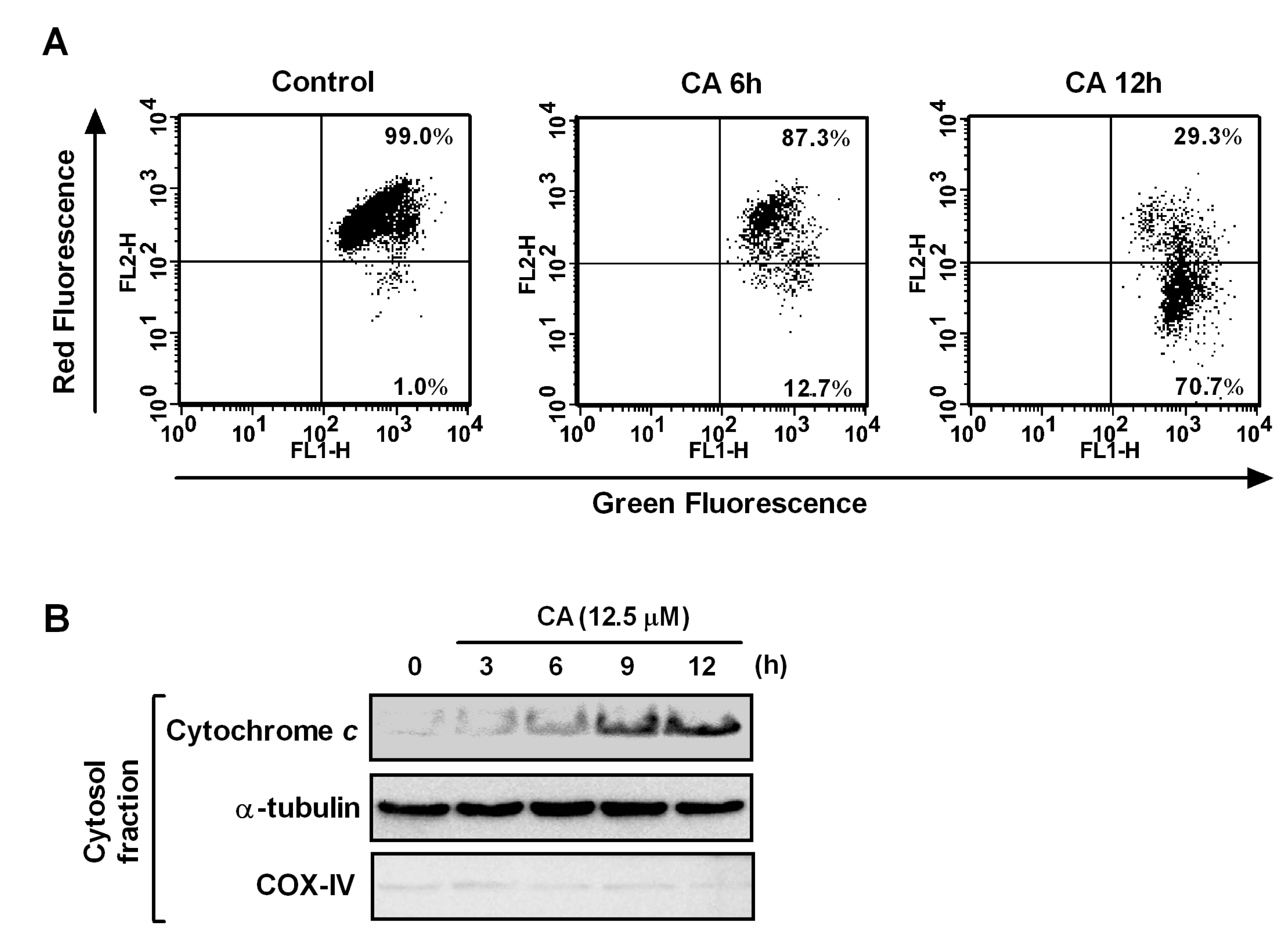
2.5. Effect of CA on Mitochondrial Dysfunction

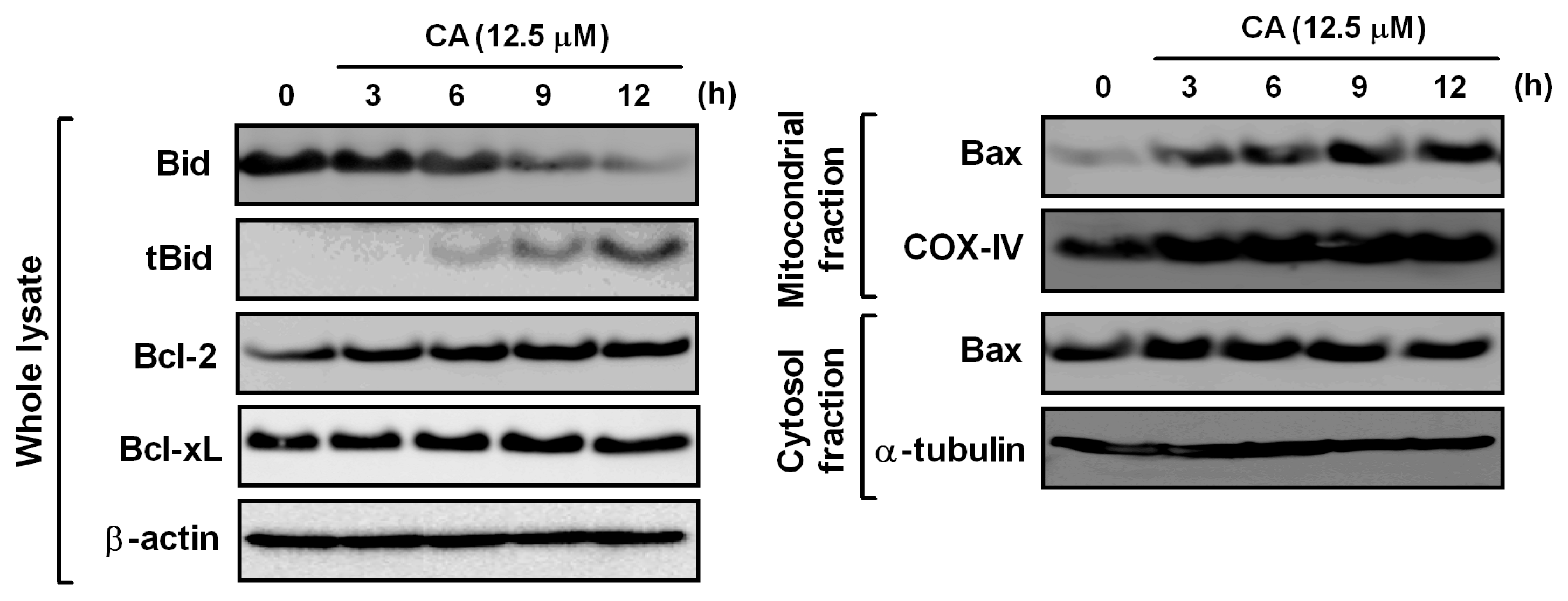
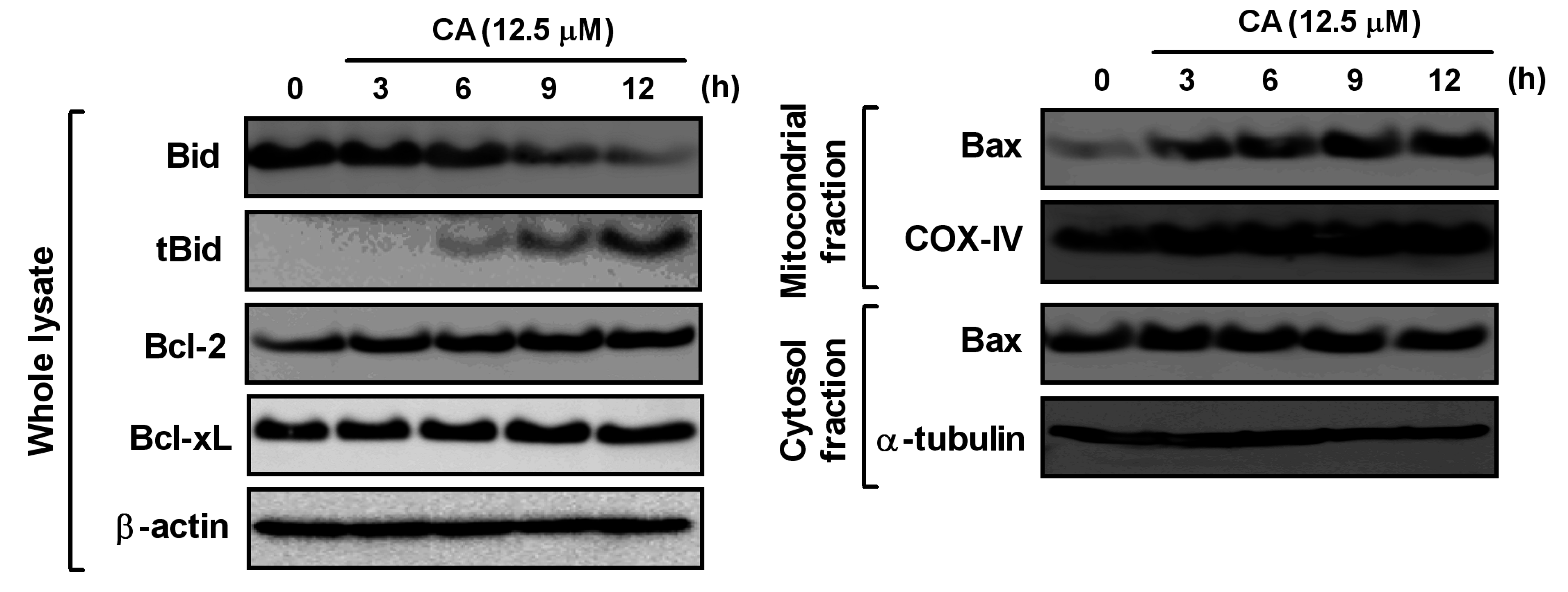
2.6. Effect of CA on Bcl-2 Family Proteins

3. Experimental Section
3.1. Materials
3.2. HPLC Fingerprints of Triterpenes in the Crude Extract from E. japonica Leaves
3.3. Purification and Isolation of Triterpenes by Preparative HPLC
3.4. Cell Culture and Treatment
3.5. Determination of Cell Viability
3.6. Nuclear Staining with Hoechst 33258
3.7. DNA Fragmentation Analysis
3.8. Flow Cytometry Analysis of Apoptotic Cells
3.9. Western Blot Analysis
3.10. Preparation of Cytosolic and Mitochondrial Fractions
3.11. Flow Cytometry Analysis of the Mitochondrial Membrane Potential (∆ψm)
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Perry, L.M. Medicinal Plants of East and Southeast Asia; the MIT Press: Cambridge, MA, USA, 1980; pp. 342–343. [Google Scholar]
- Uto, T.; Suangkaew, N.; Morinaga, O.; Kariyazono, H.; Oiso, S.; Shoyama, Y. Eriobotryae folium extract suppresses LPS-induced iNOS and COX-2 expression by inhibition of NF-κB and MAPK activation in murine macrophages. Am. J. Chin. Med. 2010, 38, 985–994. [Google Scholar] [CrossRef]
- Kang, S.C.; Lee, C.M.; Choi, H.; Lee, J.H.; Oh, J.S.; Kwak, J.H.; Zee, O.P. Evaluation of oriental medicinal herbs for estrogenic and antiproliferative activities. Phytother. Res. 2006, 20, 1017–1019. [Google Scholar] [CrossRef]
- Kim, M.S.; You, M.K.; Rhuy, D.Y.; Kim, Y.J.; Baek, H.Y.; Kim, H.A. Loquat (Eriobotrya japonica) extracts suppress the adhesion, migration and invasion of human breast cancer cell line. Nutr. Res. Pract. 2009, 3, 259–264. [Google Scholar] [CrossRef]
- Cha, D.S.; Shin, T.Y.; Eun, J.S.; Kim, D.K.; Jeon, H. Anti-metastatic properties of the leaves of Eriobotrya japonica. Arch. Pharm. Res. 2011, 34, 425–436. [Google Scholar] [CrossRef]
- Shimizu, M.; Fukumura, H.; Tsuji, H.; Tanaami, S.; Hayashi, T.; Morita, N. Anti-inflammatory constituents of topically applied crude drugs. I. Constituents and anti-inflammatory effect of Eriobotrya japonica LINDL. Chem. Pharm. Bull. 1986, 34, 2614–2617. [Google Scholar] [CrossRef]
- Ito, H.; Kobayashi, E.; Li, S.H.; Hatano, T.; Sugita, D.; Kubo, N.; Shimura, S.; Itoh, Y.; Tokuda, H.; Nishino, H.; et al. Antitumor activity of compounds isolated from leaves of Eriobotrya japonica. J. Agric. Food Chem. 2002, 50, 2400–2403. [Google Scholar]
- Huang, Y.; Li, J.; Cao, Q.; Yu, S.C.; Lv, X.W.; Jin, Y.; Zhang, L.; Zou, Y.H.; Ge, J.F. Anti-oxidative effect of triterpene acids of Eriobotrya japonica (Thunb.) Lindl. leaf in chronic bronchitis rats. Life Sci. 2006, 78, 2749–2757. [Google Scholar] [CrossRef]
- Lü, H.; Chen, J.; Li, W.L.; Ren, B.R.; Wu, J.L.; Kang, H.Y.; Zhang, H.Q.; Adams, A.; de Kimpe, N. Hypoglycemic and hypolipidemic effects of the total triterpene acid fraction from Folium Eriobotryae. J. Ethnopharmacol. 2009, 122, 486–491. [Google Scholar] [CrossRef]
- Banno, N.; Akihisa, T.; Tokuda, H.; Yasukawa, K.; Taguchi, Y.; Akazawa, H.; Ukiya, M.; Kimura, Y.; Suzuki, T.; Nishino, H. Anti-inflammatory and antitumor-promoting effects of the triterpene acids from the leaves of Eriobotrya japonica. Biol. Pharm. Bull. 2005, 28, 1995–1999. [Google Scholar] [CrossRef]
- Lee, C.H.; Wu, S.L.; Chen, J.C.; Li, C.C.; Lo, H.Y.; Cheng, W.Y.; Lin, J.G.; Chang, Y.H.; Hsiang, C.Y.; Ho, T.Y. Eriobotrya japonica leaf and its triterpenes inhibited lipopolysaccharide-induced cytokines and inducible enzyme production via the nuclear factor-κB signaling pathway in lung epithelial cells. Am. J. Chin. Med. 2008, 36, 1185–1198. [Google Scholar] [CrossRef]
- Ge, J.F.; Wang, T.Y.; Zhao, B.; Lv, X.W.; Jin, Y.; Peng, L.; Yu, S.C.; Li, J. Anti-inflammatory effect of triterpenoic acids of Eriobotrya japonica (Thunb.) Lindl. leaf on rat model of chronic bronchitis. Am. J. Chin. Med. 2009, 37, 309–321. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, Y.; Huang, C.; Lv, X.; Liu, L.; Wang, Y.; Li, J. Antifibrosis effects of triterpene acids of Eriobotrya japonica (Thunb.) Lindl. leaf in a rat model of bleomycin-induced pulmonary fibrosis. J. Pharm. Pharmacol. 2012, 64, 1751–1760. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef]
- Martinou, J.C.; Youle, R.J. Mitochondria in apoptosis: Bcl-2 family members and mitochondrial dynamics. Cell 2011, 21, 92–101. [Google Scholar]
- Ho, H.Y.; Lin, W.C.; Kitanaka, S.; Chang, C.T. Analysis of bioactive triterpenes in Eriobotrya japonica lindl. by high-performance liquid chromatography. J. Food Drug Anal. 2008, 16, 41–45. [Google Scholar]
- Taniguchi, S.; Imayoshi, Y.; Kobayashi, E.; Takamatsu, Y.; Ito, H.; Hatano, T.; Sakagami, H.; Tokuda, H.; Nishino, H.; Sugita, D.; et al. Production of bioactive triterpenes by Eriobotrya japonica calli. Phytochemistry 2002, 59, 315–323. [Google Scholar]
- Lee, M.S.; Cha, E.Y.; Thuong, P.T.; Kim, J.Y.; Ahn, M.S.; Sul, J.Y. Down-regulation of human epidermal growth factor receptor 2/neu oncogene by corosolic acid induces cell cycle arrest and apoptosis in NCI-N87 human gastric cancer cells. Biol. Pharm. Bull. 2010, 33, 931–937. [Google Scholar] [CrossRef]
- Xu, Y.; Ge, R.; Du, J.; Xin, H.; Yi, T.; Sheng, J.; Wang, Y.; Ling, C. Corosolic acid induces apoptosis through mitochondrial pathway and caspase activation in human cervix adenocarcinoma HeLa cells. Cancer Lett. 2009, 284, 229–237. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Komohara, Y.; Ikeda, T.; Takeya, M. Corosolic acid inhibits glioblastoma cell proliferation by suppressing the activation of signal transducer and activator of transcription-3 and nuclear factor-κB in tumor cells and tumor-associated macrophages. Cancer Sci. 2011, 102, 206–211. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Wang, X.; Jiang, C. Ursolic acid inhibits proliferation and induces apoptosis in human glioblastoma cell lines U251 by suppressing TGF-β1/miR-21/PDCD4 pathway. Basic Clin. Pharmacol. Toxicol. 2012, 111, 106–112. [Google Scholar]
- Reyes-Zurita, F.J.; Rufino-Palomares, E.E.; Lupiáñez, J.A.; Cascante, M. Maslinic acid, a natural triterpene from Olea europaea L., induces apoptosis in HT29 human colon-cancer cells via the mitochondrial apoptotic pathway. Cancer Lett. 2009, 273, 44–54. [Google Scholar] [CrossRef]
- Juan, M.E.; Planas, J.M.; Ruiz-Gutierrez, V.; Daniel, H.; Wenzel, U. Antiproliferative and apoptosis-inducing effects of maslinic and oleanolic acids, two pentacyclic triterpenes from olives, on HT-29 colon cancer cells. Br. J. Nutr. 2008, 100, 36–43. [Google Scholar]
- Tung, N.H.; Shoyama, Y. Eastern blotting analysis and isolation of two new dammarane-type saponins from American ginseng. Chem. Pharm. Bull. 2012, 60, 1329–1333. [Google Scholar] [CrossRef]
- Shang, M.Y.; Tian, M.; Tanaka, H.; Li, X.W.; Cai, S.Q.; Shoyama, Y. Quality control oftraditional chinese medicine by monoclonal antibody method. Curr. Drug Discov. Technol. 2011, 8, 60–65. [Google Scholar] [CrossRef]
- Tanaka, H.; Fukuda, N.; Yahara, S.; Isoda, S.; Yuan, C.S.; Shoyama, Y. Isolation of ginsenoside Rb1 from Kalopanax pictus by eastern blotting using anti-ginsenoside Rb1 monoclonal antibody. Phytother. Res. 2005, 19, 255–258. [Google Scholar] [CrossRef]
- Tanaka, H.; Fukuda, N.; Shoyama, Y. Eastern blotting and immunoaffinity concentration using monoclonal antibody for ginseng saponins in the field of traditional Chinese medicines. J. Agric. Food Chem. 2007, 55, 3783–3787. [Google Scholar] [CrossRef]
- Ulukaya, E.; Acilan, C.; Yilmaz, Y. Apoptosis: Why and how does it occur in biology? Cell Biochem. Funct. 2011, 29, 468–480. [Google Scholar] [CrossRef]
- Armstrong, J.S. Mitochondria: A target for cancer therapy. Br. J. Pharmacol. 2006, 147, 239–248. [Google Scholar] [CrossRef]
- Von Ahsen, O.; Waterhouse, N.J.; Kuwana, T.; Newmeyer, D.D.; Green, D.R. The “harmless” release of cytochrome c. Cell Death Differ. 2000, 7, 1192–1199. [Google Scholar] [CrossRef]
- Crompton, M. Bax, Bid and the permeabilization of the mitochondrial outer membrane in apoptosis. Curr. Opin. Cell Biol. 2000, 12, 414–419. [Google Scholar] [CrossRef]
- Uto, T.; Morinaga, O.; Tanaka, H.; Shoyama, Y. Analysis of the synergistic effect of glycyrrhizin and other constituents in licorice extract on lipopolysaccharide-induced nitric oxide production using knock-out extract. Biochem. Biophys. Res. Commun. 2012, 417, 473–478. [Google Scholar] [CrossRef]
- Aung, H.H.; Wang, C.Z.; Ni, M.; Fishbein, A.; Mehendale, S.R.; Xie, J.T.; Shoyama, Y.; Yuan, C.S. Crocin from Crocus sativus possesses significant anti-proliferation effects on human colorectal cancer cells. Exp. Oncol. 2007, 29, 175–180. [Google Scholar]
- Tung, N.H.; Du, G.J.; Wang, C.Z.; Yuan, C.S.; Shoyama, Y. Naphthoquinone components from Alkanna tinctoria (L.) Tausch show significant antiproliferative effects on human colorectal cancer cells. Phytother. Res. 2013, 27, 66–70. [Google Scholar] [CrossRef]
- Patlolla, J.M.; Rao, C.V. Triterpenoids for cancer prevention and treatment: Current status and future prospects. Curr. Pharm. Biotechnol. 2012, 13, 147–155. [Google Scholar] [CrossRef]
- Bishayee, A.; Ahmed, S.; Brankov, N.; Perloff, M. Triterpenoids as potential agents for the chemoprevention and therapy of breast cancer. Front Biosci. 2011, 16, 980–996. [Google Scholar] [CrossRef]
- Fulda, S. Modulation of apoptosis by natural products for cancer therapy. Planta Med. 2010, 76, 1075–1079. [Google Scholar] [CrossRef]
- Wilkinson, K.; Boyd, J.D.; Glicksman, M.; Moore, K.J.; El Khoury, J. A high content drug screen identifies ursolic acid as an inhibitor of amyloid beta protein interactions with its receptor CD36. J. Biol. Chem. 2011, 286, 34914–34922. [Google Scholar]
- Murakami, A.; Ohnishi, K. Target molecules of food phytochemicals: Food science bound for the next dimension. Food Funct. 2012, 3, 462–476. [Google Scholar] [CrossRef]
- Ikeda, Y.; Murakami, A.; Fujimura, Y.; Tachibana, H.; Yamada, K.; Masuda, D.; Hirano, K.; Yamashita, S.; Ohigashi, H. Aggregated ursolic acid, a natural triterpenoid, induces IL-1beta release from murine peritoneal macrophages: Role of CD36. J. Immunol. 2007, 178, 4854–4864. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Uto, T.; Sakamoto, A.; Tung, N.H.; Fujiki, T.; Kishihara, K.; Oiso, S.; Kariyazono, H.; Morinaga, O.; Shoyama, Y. Anti-Proliferative Activities and Apoptosis Induction by Triterpenes Derived from Eriobotrya japonica in Human Leukemia Cell Lines. Int. J. Mol. Sci. 2013, 14, 4106-4120. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14024106
Uto T, Sakamoto A, Tung NH, Fujiki T, Kishihara K, Oiso S, Kariyazono H, Morinaga O, Shoyama Y. Anti-Proliferative Activities and Apoptosis Induction by Triterpenes Derived from Eriobotrya japonica in Human Leukemia Cell Lines. International Journal of Molecular Sciences. 2013; 14(2):4106-4120. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14024106
Chicago/Turabian StyleUto, Takuhiro, Ayana Sakamoto, Nguyen Huu Tung, Tsukasa Fujiki, Kenji Kishihara, Shigeru Oiso, Hiroko Kariyazono, Osamu Morinaga, and Yukihiro Shoyama. 2013. "Anti-Proliferative Activities and Apoptosis Induction by Triterpenes Derived from Eriobotrya japonica in Human Leukemia Cell Lines" International Journal of Molecular Sciences 14, no. 2: 4106-4120. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14024106