Low Oxygen Response Mechanisms in Green Organisms

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Plant Metabolic Regulations under Low Oxygen
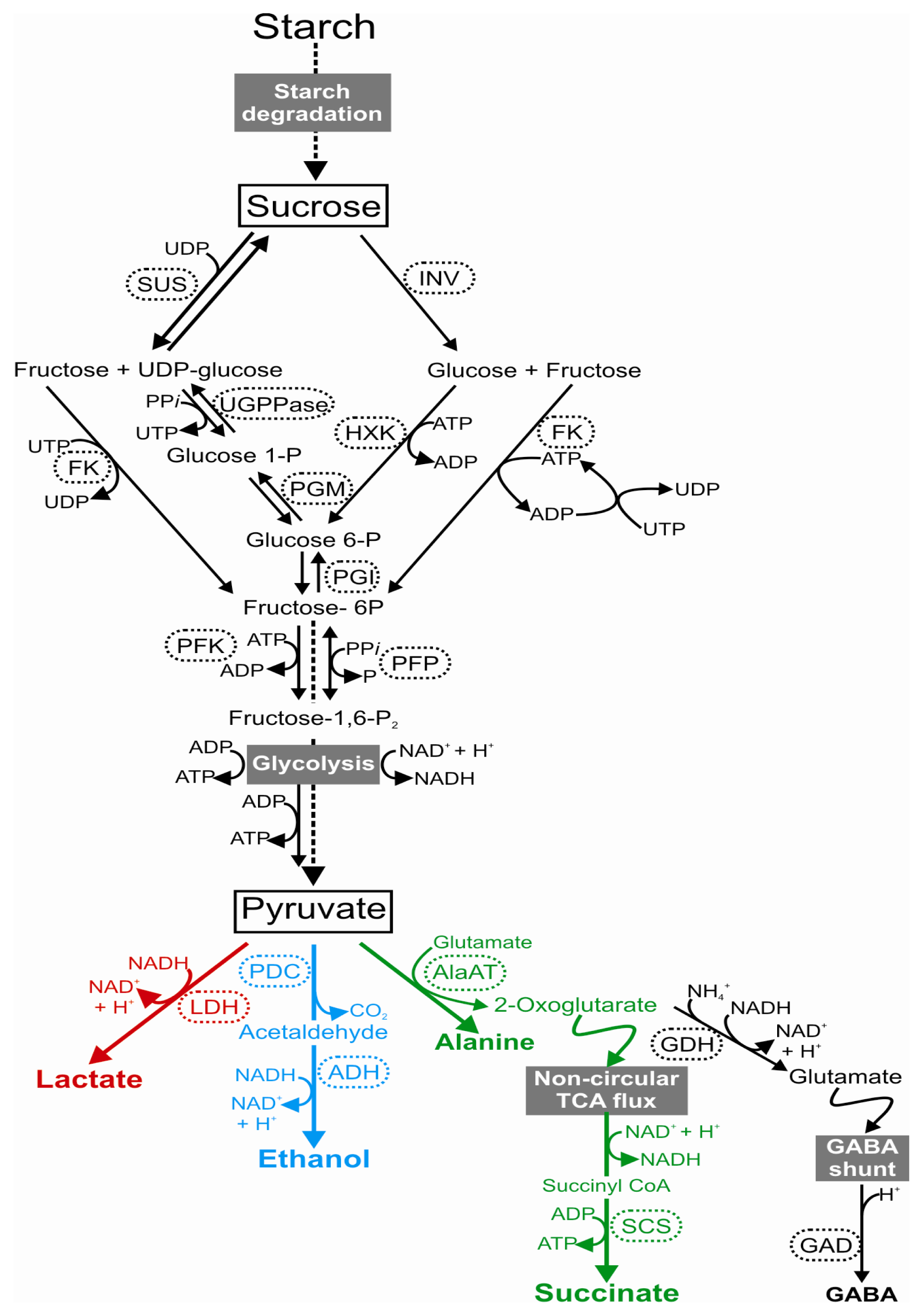
1.1. A Shift towards the Fermentation Pathway
1.2. Sugar Metabolism under Low Oxygen
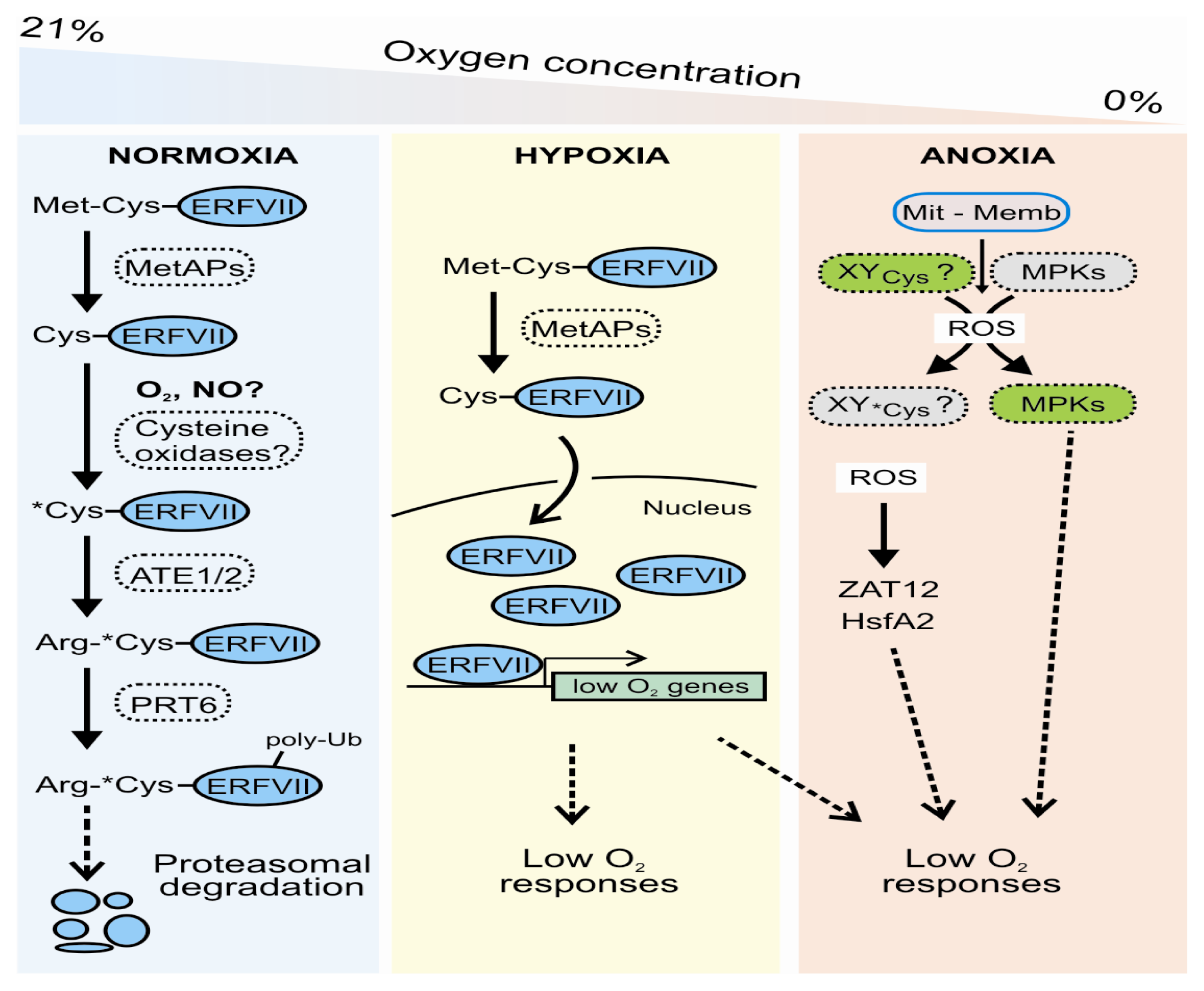
2. Low Oxygen Sensing and Signaling in Arabidopsis thaliana: The N-End Rule Pathway (NERP) and Side Defense Mechanisms
2.1. Oxygen Sensing in Plants
2.2. A Role for Reactive Oxygen and Nitrogen Species (ROS/NOS)
2.3. Low Oxygen and Heat Stress Convergence
3. Low Oxygen Tolerance Mechanisms in Oryza sativa
3.1. Rice Germination under Low Oxygen
3.2. Strategies to Survive Low Oxygen in Adult Rice Plants
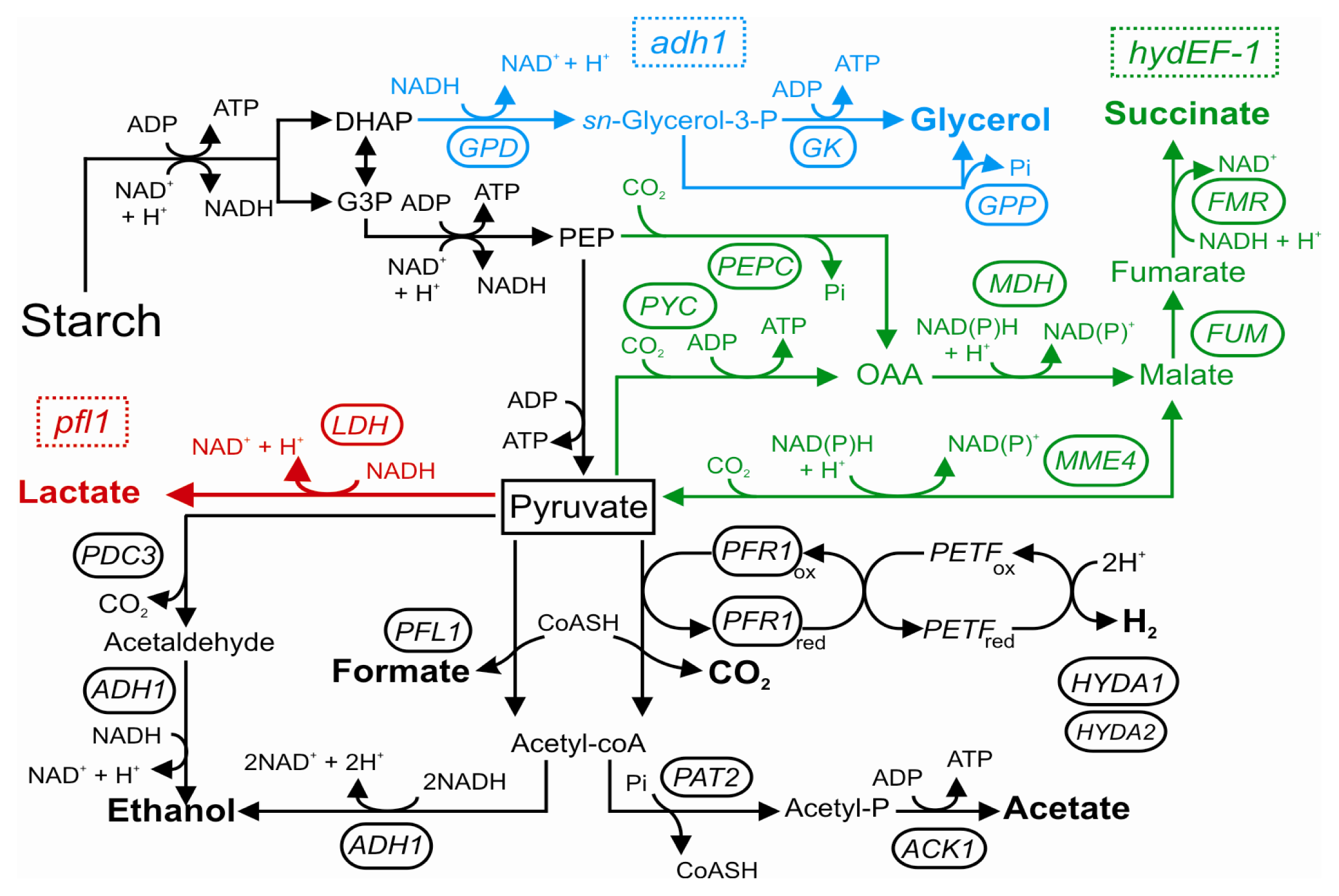
4. Anaerobic Metabolism in Photosynthetic Microorganisms: The case of Chlamydomonas reinhardtii
5. Concluding Remarks
Acknowledgments
References
- Drew, M.C. Oxygen deficiency and root metabolism: Injury and acclimation under hypoxia and anoxia. Annu. Rev. Plant Physiol. Plant Mol. Biol 1997, 48, 223–250. [Google Scholar]
- Bailey-Serres, J.; Fukao, T.; Gibbs, D.J.; Holdsworth, M.J.; Lee, S.C.; Licausi, F.; Perata, P.; Voesenek, L.A.C.J.; van Dongen, J.T. Making sense of low oxygen sensing. Trends Plant Sci 2012, 17, 129–138. [Google Scholar]
- Gupta, K.J.; Zabalza, A.; van Dongen, J.T. Regulation of respiration when the oxygen availability changes. Physiol. Plantarum 2009, 137, 383–391. [Google Scholar]
- Zabalza, A.; van Dongen, J.T.; Froehlich, A.; Oliver, S.N.; Faix, B.; Gupta, K.J.; Schmalzlin, E.; Igal, M.; Orcaray, L.; Royuela, M.; et al. Regulation of respiration and fermentation to control the plant internal oxygen concentration. Plant Physiol 2009, 149, 1087–1098. [Google Scholar]
- Kennedy, R.A.; Rumpho, M.E.; Fox, T.C. Anaerobic metabolism in plants. Plant Physiol 1992, 100, 1–6. [Google Scholar]
- Ricard, B.; Couee, I.; Raymond, P.; Saglio, P.H.; Saint-Ges, V.; Pradet, A. Plant metabolism under hypoxia and anoxia. Plant Physiol. Biochem 1994, 32, 1–10. [Google Scholar]
- Gibbs, J.; Morrell, S.; Valdez, A.; Setter, T.L.; Greenway, H. Regulation of alcoholic fermentation in coleoptiles of two rice cultivars differing in tolerance to anoxia. J. Exp. Bot 2000, 51, 758–796. [Google Scholar]
- Geigenberger, P. Response of plant metabolism to too little oxygen. Curr. Opin. Plant Biol 2003, 6, 247–256. [Google Scholar]
- Rivoal, J.; Ricard, B.; Pradet, A. Lactate dehydrogenase in Oryza sativa L. seedlings and roots: Identification and partial characterization. Plant Physiol 1991, 95, 682–686. [Google Scholar]
- Rivoal, J.; Hanson, A. Metabolic control of anaerobic glycolysis: Overexpression of lactate dehydrogenase in transgenic tomato roots supports the Davies-Roberts hypothesis and points to a critical role for lactate secretion. Plant Physiol 1994, 106, 1179–1185. [Google Scholar]
- Christopher, M.E.; Good, A.G. Characterization of hypoxically inducible lactate dehydrogenase in maize. Plant Physiol 1996, 112, 1015–1022. [Google Scholar]
- Sweetlove, L.J.; Dunford, R.; Ratcliffe, R.G.; Kruger, N.J. Lactate metabolism in potato tubers deficient in lactate dehydrogenase activity. Plant Cell Environ 2000, 23, 873–881. [Google Scholar]
- Roberts, J.K.; Callis, J.; Jardetzky, O.; Walbot, V.; Freeling, M. Cytoplasmic acidosis as a determinant of flooding intolerance in plants. Proc. Natl. Acad. Sci. USA 1984, 81, 6029–6033. [Google Scholar]
- Felle, H.H. pH regulation in anoxic plants. Ann. Bot 2005, 96, 519–532. [Google Scholar]
- Licausi, F.; Perata, P. Low oxygen signaling and tolerance in plants. Adv. Bot. Res 2009, 50, 139–198. [Google Scholar]
- Choi, W.G.; Roberts, D.M. Arabidopsis NIP2;1, a major intrinsic protein transporter of lactic acid induced by anoxic stress. J. Biol. Chem 2007, 282, 24209–24218. [Google Scholar]
- Dolferus, R.; Wolansky, M.; Carroll, R.R.; Miyashita, R.Y.; Ismond, K.K.; Good, A. Functional analysis of lactate dehydrogenase during hypoxic stress in Arabidopsis. Funct. Plant Biol 2008, 35, 131–140. [Google Scholar]
- Perata, P.; Alpi, A. Plant responses to anaerobiosis. Plant Sci 1993, 93, 1–17. [Google Scholar]
- Dolferus, R.; Ellis, M.; de Bruxelles, G.; Trevaskis, B.; Hoeren, F.; Dennis, E.S.; Peacock, W.J. Strategies of gene action in Arabidopsis during hypoxia. Ann. Bot 1997, 79, 21–31. [Google Scholar]
- Ellis, M.H.; Dennis, E.S.; Peacock, W.J. Arabidopsis roots and shoots have different mechanisms for hypoxic stress tolerance. Plant Physiol 1999, 119, 57–64. [Google Scholar]
- Dolferus, R.; Klok, E.J.; Delessert, C.; Wilson, S.; Ismond, K.P.; Good, A.G.; Peacock, W.J.; Dennis, E.S. Enhancing the anaerobic response. Ann. Bot 2003, 91, 111–117. [Google Scholar]
- Visser, E.J.W.; Voesenek, L.A.C.J.; Vartapetian, B.B.; Jackson, M.B. Flooding and plant growth. Ann. Bot 2003, 91, 107–109. [Google Scholar]
- Loreti, E.; Poggi, A.; Novi, G.; Alpi, A.; Perata, P. A genome-wide analysis of the effects of sucrose on gene expression in Arabidopsis seedlings under anoxia. Plant Physiol 2005, 137, 1130–1138. [Google Scholar]
- Bucher, M.; Brädle, R.; Khuhlemeier, C. Ethanolic fermentation in transgenic tobacco expressing Zymomonas mobilis pyruvate decarboxylase. EMBO J 1994, 13, 2755–2763. [Google Scholar]
- Kürsteiner, O.; Dupuis, I.; Kuhlemeier, C. The pyruvate decarboxylase1 gene of Arabidopsis is required during anoxia but not other environmental stresses. Plant Physiol 2003, 132, 968–978. [Google Scholar]
- Mithran, M.; Paparelli, E.; Novi, G.; Perata, P.; Loreti, E. Analysis of the role of the pyruvate decarboxylase gene family in Arabidopsis thaliana under low oxygen conditions. Plant Biol. 2013. [Google Scholar] [CrossRef] [Green Version]
- Ismond, K.P.; Dolferus, R.; de Pauw, M.; Dennis, E.S.; Good, A.G. Enhanced low oxygen survival in Arabidopsis through increased metabolic flux in the fermentative pathway. Plant Physiol 2003, 132, 1292–1302. [Google Scholar]
- Jacobs, M.; Dolferus, R.; van Den Bosshe, V.B. Isolation and biochemical analysis of ethyl methyl sulfonate induced alcohol dehydrogenase null mutants of Arabidopsis thaliana (L.) Heynh. Biochem. Genet 1988, 26, 102–112. [Google Scholar]
- Matsumura, H.; Takano, T.; Yoshida, K.T.; Takeda, G. A rice mutant lacking alcohol dehydorogenase. Breeding Sci 1995, 45, 365–367. [Google Scholar]
- Matsumura, H.; Takano, T.; Takeda, G.; Uchimiya, H. Adh1 is transcriptionally active but its translational product is reduced in a rad mutant of rice (Oryza sativa L.), which is vulnerable to submergence stress. Theor. Appl. Genet 1998, 97, 1197–1203. [Google Scholar]
- Johnson, J.R.; Cobb, B.G.; Drew, M.C. Hypoxic induction of anoxia tolerance in roots of Adh null Zea mays. Plant Physiol 1994, 105, 61–67. [Google Scholar]
- Shiao, T.; Ellis, M.H.; Dolferus, R.; Dennis, E.S.; Doran, P.M. Overexpression of alcohol dehydrogenase or pyruvate decarboxylase improves the growth of hairy roots under hypoxia. Biotechnol. Bioeng 2002, 77, 455–461. [Google Scholar]
- Magneschi, L.; Perata, P. Rice germination and seedling growth in the absence of oxygen. Ann. Bot 2009, 103, 181–196. [Google Scholar]
- Rocha, M.; Licausi, F.; Araújo, W.L.; Nunes-Nesi, A.; Sodek, L.; Fernie, A.R.; van Dongen, J.T. Glycolysis and the tricarboxylic acid cycle are linked by alanine aminotransferase during hypoxia induced by waterlogging of Lotus japonicus. Plant Physiol 2010, 152, 1501–1513. [Google Scholar]
- Branco-Price, C.; Kaiser, K.A.; Jang, C.J.; Larive, C.K.; Bailey-Serres, J. Selective mRNA translation coordinates energetic and metabolic adjustments to cellular oxygen deprivation and reoxygenation in Arabidopsis thaliana. Plant J 2008, 56, 743–755. [Google Scholar]
- Ricoult, C.; Cliquet, J.B.; Limami, A.M. Stimulation of alanine amino transferase (AlaAT) gene expression and alanine accumulation in embryo axis of the model legume Medicago truncatula contribute to anoxia stress tolerance. Physiol. Plantarum 2005, 123, 30–39. [Google Scholar]
- Miyashita, Y.; Dolferus, R.; Ismond, K.P.; Good, A.G. Alanine aminotransferase catalyses the breakdown of alanine after hypoxia in Arabidopsis thaliana. Plant J 2007, 49, 1108–1121. [Google Scholar]
- Aurisano, N.; Bertani, A.; Reggiani, R. Involvement of calcium and calmodulin in protein and amino acid metabolism in rice roots under anoxia. Plant Cell Physiol 1995, 36, 1525–1529. [Google Scholar]
- Breitkreuz, K.E.; Allan, W.L.; van Cauwenberghe, O.R.; Jakobs, C.; Talibi, D.; Andre, B.; Shelp, B.J. Anovel γ-hydroxybutyrate dehydrogenase: Identification and expression of an Arabidopsis cDNA and potential role under oxygen deficiency. J. Biol. Chem 2003, 278, 41552–41556. [Google Scholar]
- Sweetlove, L.J.; Beard, K.F.M.; Nunes-Nesi, A.; Fernie, A.R.; Ratcliffe, R.G. Not just a circle: Flux modes in the plant TCA cycle. Trends Plant Sci 2010, 15, 462–470. [Google Scholar]
- Gibbs, D.J.; Lee, S.C.; Isa, N.M.; Gramuglia, S.; Fukao, T.; Bassel, G.W.; Correia, C.S.; Corbineau, F.; Theodoulou, F.L.; Bailey-Serres, J.; et al. Homeostatic response to hypoxia is regulated by the N-end rule pathway in plants. Nature 2011, 479, 415–418. [Google Scholar]
- Licausi, F.; Kosmacz, M.; Weits, D.A.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.A.C.J.; Perata, P.; van Dongen, J.T. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nature 2011, 479, 419–422. [Google Scholar]
- Roitsch, T. Source-sink regulation by sugar and stress. Curr. Opin. Plant Biol 1999, 2, 198–206. [Google Scholar]
- Blasing, O.E.; Gibon, Y.; Gunther, M.; Hohne, M.; Morcuende, R. Sugars and circadian regulation make major contributions to the global regulation of diurnal gene expression in Arabidopsis. Plant Cell 2005, 17, 3257–3281. [Google Scholar]
- Xia, J.H.; Saglio, P.H. Lactic Acid efflux as a mechanism of hypoxic acclimation of maize root tips to anoxia. Plant Physiol 1992, 100, 40–46. [Google Scholar]
- Sairam, R.K.; Dharmar, K.; Chinnusamy, V.; Meena, R.C. Waterlogging-induced increase in sugar mobilization, fermentation, and related gene expression in the roots of mung bean (Vigna radiata). J. Plant Physiol 2009, 166, 602–616. [Google Scholar]
- Vartapetian, B.B.; Jackson, M.B. Plant adaptation to anaerobic stress. Ann. Bot 1997, 79, 3–20. [Google Scholar]
- Sturm, A.; Hess, D.; Lee, H.S.; Lienhard, S. Neutral invertase is a novel type of sucrose–cleaving enzyme. Physiol. Plant 1999, 107, 159–165. [Google Scholar]
- Bailey-Serres, J.; Voesenek, L.A.C.J. Flooding stress: Acclimations and genetic diversity. Annu. Rev. Plant Biol 2008, 59, 313–339. [Google Scholar]
- Barclay, A.M.; Crawford, R.M. The effect of anaerobiosis on carbohydrate levels in storage tissues of wetland plants. Ann. Bot 1983, 51, 255–259. [Google Scholar]
- Bertrand, A.; Castonguay, Y.; Nadeau, P.; Laberge, S.; Michaud, R.; Bélanger, G.; Rochette, P. Oxygen deficiency affects carbohydrate reserves in overwintering forage crops. J. Exp. Bot 2003, 54, 1721–1730. [Google Scholar]
- Stitt, M. Fructose-2,6-bisphosphate as a regulatory molecule in plants. Trends Plant Sci 1990, 41, 153–185. [Google Scholar]
- Smith, A.M.; Zeeman, S.C.; Smith, S.M. Starch degradation. Ann. Rev. Plant Biol 2005, 56, 73–98. [Google Scholar]
- Germain, V.; Raymond, P.; Ricard, B. Differential expression of two tomato lactate dehydrogenase genes in response to oxygen deficit. Plant Mol. Biol 1997, 35, 711–721. [Google Scholar]
- Zeng, Y.; Wu, Y.; Avigne, W.T.; Koch, K.E. Rapid repression of maize invertases by low oxygen. Invertase/sucrose synthase balance, sugar signaling potential, and seedling survival. Plant Physiol 1999, 121, 599–608. [Google Scholar]
- Van Dongen, J.T.; Roeb, G.W.; Dautzenberg, M.; Froehlich, A.; Vigeolas, H.; Minchin, P.E.H.; Geigenberger, P. Phloem import and storage metabolism are highly coordinated by the low oxygen concentrations within developing wheat seeds. Plant Physiol 2004, 135, 1809–1821. [Google Scholar]
- Mustroph, A.; Albrecht, G.; Hajirezaei, M.; Grimm, B.; Biemelt, S. Low levels of pyrophosphate in transgenic potato plants expressing E. coli pyrophosphatase lead to decreased vitality under oxygen deficiency. Ann. Bot 2005, 96, 717–726. [Google Scholar]
- Zeng, Y.; Wu, Y.; Avigne, W.T.; Koch, K.E. Differential regulation of sugar-sensitive sucrose synthases by hypoxia and anoxia indicate complementary transcriptional and posttranscriptional responses. Plant Physiol 1998, 116, 1573–1583. [Google Scholar]
- Baud, S.; Vaultier, M.N.; Rochat, C. Structure and expression profile of the sucrose synthase multigene family in Arabidopsis. J. Exp. Bot 2004, 55, 397–409. [Google Scholar]
- Bieniawska, Z.; Paul Barratt, D.H.; Garlick, A.P.; Thole, V.; Kruger, N.J. Analysis of the sucrose synthase gene family in Arabidopsis. Plant J 2007, 49, 810–828. [Google Scholar]
- Guglielminetti, L.; Morita, A.; Yamaguchi, J.; Loreti, E.; Perata, P.; Alpi, A. Differential expression of two fructokinases in Oryza sativa seedlings grown under aerobic and anaerobic conditions. J. Plant Res 2006, 119, 351–356. [Google Scholar]
- Cho, J.I.; Ryoo, N.; Ko, S.; Lee, S.K.; Lee, J.; Jung, K.H.; Lee, Y.H.; Bhoo, S.; Winderickx, J.; An, G.; et al. Structure, expression, and functional analysis of the hexokinase gene family in rice (Oryza sativa L.). Planta 2006, 224, 598–611. [Google Scholar]
- Lasanthi-Kudahettige, R.; Magneschi, L.; Loreti, E.; Gonzali, S.; Licausi, F.; Novi, G.; Beretta, O.; Vitulli, F.; Alpi, A.; Perata, P. Transcript profiling of the anoxic rice coleoptile. Plant Physiol 2007, 144, 218–231. [Google Scholar]
- Mertens, E.; Larondelle, Y.; Hers, H.G. Induction of pyrophosphate: Fructose 6-phosphate 1-phosphotransferase by anoxia in rice seedlings. Plant Physiol 1990, 93, 584–587. [Google Scholar]
- Mohanty, B.; Wilson, P.M.; ap Rees, T. Effects of anoxia on growth and carbohydrate metabolism in suspension cultures of soybean and rice. Phytochemistry 1993, 34, 75–82. [Google Scholar]
- Faiz-Ur-Rahman, A.T.M.; Trewavas, A.J.; Davies, D.D. The Pasteur effect in carrot root tissue. Planta 1974, 118, 195–210. [Google Scholar]
- Moons, A.; Valcke, R.; van Montagu, M. Low-oxygen stress and water deficit induce cytosolic pyruvate orthophosphate dikinase (PPDK) expression in roots of rice, a C3 plant. Plant J 1998, 15, 89–98. [Google Scholar]
- Kaelin, W.G., Jr; Ratcliffe, P.J. Oxygen sensing by metazoans: The central role of the HIF hydroxylase pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar]
- Loenarz, C.; Coleman, M.L.; Boleininger, A.; Schierwater, B.; Holland, P.W.H.; Ratcliffe, P.J.; Schofield, C.J. The hypoxia-inducible transcription factor pathway regulates oxygen sensing in the simplest animal, Trichoplax. adhaerens. EMBO Rep 2011, 12, 63–70. [Google Scholar]
- Sasidharan, R.; Mustroph, A. Plant oxygen sensing is mediated by the N-end rule pathway: A milestone in plant anaerobiosis. Plant Cell 2011, 23, 4173–4183. [Google Scholar]
- Mus, F.; Dubini, A.; Seibert, M.; Posewitz, M.C.; Grossman, A.R. Anaerobic acclimation in Chlamydomonas reinhardtii, anoxic gene expression, hydrogenase induction, and metabolic pathways. J. Biol. Chem 2007, 282, 25475–25486. [Google Scholar]
- Bachmair, A.; Finley, D.; Varshavsky, A. In vivo half-life of a protein is a function of its amino-terminal residue. Science 1986, 234, 179–186. [Google Scholar]
- Varshavsky, A. The N-end rule pathway and regulation by proteolysis. Protein Sci 2011, 20, 1298–1345. [Google Scholar]
- Prakash, S.; Tian, L.; Ratliff, K.S.; Lehotzky, R.E.; Matouschek, A. An unstructured initiation site is required for efficient proteasome-mediated degradation. Nat. Struct. Mol. Biol 2004, 11, 830–837. [Google Scholar]
- Ditzel, M.; Wilson, R.; Tenev, T.; Zachariou, A.; Paul, A.; Deas, E.; Meier, P. Degradation of DIAP1 by the N-end rule pathway is essential for regulating apoptosis. Nat. Cell Biol 2003, 5, 467–473. [Google Scholar]
- Mogk, A.; Schmidt, R.; Bukau, B. The N-end rule pathway for regulated proteolysis: Prokaryotic and eukaryotic strategies. Trends Cell Biol 2007, 17, 165–172. [Google Scholar]
- Graciet, E.; Mesiti, F.; Wellmer, F. Structure and evolutionary conservation of the plant N-end rule pathway. Plant J 2010, 61, 741–751. [Google Scholar]
- Tasaki, T.; Kwon, Y.T. The mammalian N-end rule pathway: New insights into its components and physiological roles. Trends Biochem. Sci 2007, 32, 520–528. [Google Scholar]
- Graciet, E.; Wellmer, F. The plant N-end rule pathway: Structure and functions. Trends Plant Sci 2010, 15, 447–453. [Google Scholar]
- Yoshida, S.; Ito, M.; Callis, J.; Nishida, I.; Watanabe, A. A delayed leaf senescence mutant is defective in arginyl-tRNA: Proteinarginyltransferase, a component of the N-end rule pathway in Arabidopsis. Plant J 2002, 32, 129–137. [Google Scholar]
- Graciet, E.; Walter, F.; Maoileidigh, D.O.; Pollmann, S.; Meyerowitz, E.M.; Varshavsky, A.; Wellmer, F. The N-end rule pathway controls multiple functions during Arabidopsis shoot and leaf development. Proc. Natl. Acad. Sci. USA 2009, 106, 13618–13623. [Google Scholar]
- Holman, T.J.; Jones, P.D.; Russell, L.; Medhurst, A.; Ubeda, T.S.; Talloji, P.; Marquez, J.; Schmuths, H.; Tung, S.A.; Taylor, I.; et al. The N-end rule pathway promotes seed germination and establishment through removal of ABA sensitivity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 4549–4554. [Google Scholar]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol 2006, 140, 411–432. [Google Scholar]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar]
- Papdi, C.; Abraham, E.; Joseph, M.P.; Popescu, C.; Koncz, C.; Szabados, L. Functional identification of Arabidopsis stress regulatory genes using the controlled cDNA overexpression system. Plant Physiol. 2008, 147, 528–542. [Google Scholar]
- Hattori, Y.; Nagai, K.; Furukawa, S.; Song, X.-J.; Kawano, R.; Sakakibara, H.; Wu, J.; Matsumoto, T.; Yoshimura, A.; Kitano, H.; et al. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 2009, 460, 1026–1030. [Google Scholar]
- Licausi, F.; van Dongen, J.T.; Giuntoli, B.; Novi, G.; Santaniello, A.; Geigenberger, P.; Perata, P. HRE1 and HRE2, two hypoxia-inducible ethylene response factors, affect anaerobic responses in Arabidopsis thaliana. Plant J 2010, 62, 302–315. [Google Scholar]
- Hu, R.-G.; Sheng, J.; Qi, X.; Xu, Z.; Takahashi, T.T.; Varshavsky, A. The N-end rule pathway as a nitric oxide sensor controlling the levels of multiple regulators. Nature 2005, 437, 981–986. [Google Scholar]
- Mustroph, A.; Lee, S.C.; Oosumi, T.; Zanetti, M.E.; Yang, H.; Ma, K.; Yaghoubi-Masihi, A.; Fukao, T.; Bailey-Serres, J. Cross-kingdom comparison of transcriptomic adjustments to low oxygen stress highlights conserved and plant-specific responses. Plant Physiol 2010, 152, 1484–1500. [Google Scholar]
- Pucciariello, C.; Parlanti, S.; Banti, V.; Novi, G.; Perata, P. Reactive oxygen species-driven transcription in Arabidopsis under oxygen deprivation. Plant Physiol 2012, 159, 184–196. [Google Scholar]
- Vandenbroucke, K.; Robbens, S.; Vandepoele, K.; Inzè, D.; van de Peer, Y.; van Breusegem, F. Hydrogen peroxide-induced gene expression across kingdoms: A comparative analysis. Mol. Biol. Evol 2008, 25, 507–516. [Google Scholar]
- Banti, V.; Mafessoni, F.; Loreti, E.; Alpi, A.; Perata, P. The heat-inducible transcription factor HsfA2 enhances anoxia tolerance in Arabidopsis. Plant Physiol 2010, 152, 1471–1483. [Google Scholar]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci 2004, 9, 490–498. [Google Scholar]
- Davletova, S.; Schlauch, K.; Coutu, J.; Mittler, R. The zinc-finger protein Zat12 plays a central role in reactive oxygen and abiotic stress signaling in Arabidopsis. Plant Physiol 2005, 133, 847–856. [Google Scholar]
- Baxter-Burrell, A.; Yang, Z.; Springer, P.S.; Bailey-Serres, J. RopGAP4-dependent Rop GTPase rheostat control of Arabidopsis oxygen deprivation tolerance. Science 2002, 296, 2026–2028. [Google Scholar]
- Chang, R.; Jang, C.J.H.; Branco-Price, C.; Nghiem, P.; Bailey-Serres, J. Transient MPK6 activation in response to oxygen deprivation and reoxygenation is mediated by mitochondria and aids seedling survival in Arabidopsis. Plant Mol. Biol 2012, 78, 109–122. [Google Scholar]
- Pucciariello, C.; Perata, P. How plants sense low oxygen. Plant Signal. Behav 2012, 7, 1–4. [Google Scholar]
- Licausi, F.; Pucciariello, C.; Perata, P. New role for an old rule: N-end rule mediated degradation of ERF proteins governs low oxygen response in plants. J. Integr. Plant Biol 2012, 55, 31–39. [Google Scholar]
- Miki, H.; Funato, Y. Regulation of intracellular signalling through cysteine oxidation by reactive oxygen species. J. Biochem 2012, 151, 255–261. [Google Scholar]
- Rhee, S.G.; Chae, H.Z.; Kim, K. Peroxiredoxins: A historical overview and speculative preview of novel mechanisms and emerging concepts in cell signaling. Free Radic. Biol. Med 2005, 38, 1543–1552. [Google Scholar]
- Gupta, R.; Luan, S. Redox control of protein tyrosine phosphatases and mitogen-activated protein kinases in plants. Plant Physiol 2003, 132, 1149–1152. [Google Scholar]
- Blanchetot, C.; Tertoolen, L.G.J.; Hertog, J.D. Regulation of receptor protein-tyrosine phosphatase a by oxidative stress. EMBO J 2002, 21, 493–503. [Google Scholar]
- Igamberdiev, A.U.; Bykova, N.V.; Shah, J.K.; Hill, R.D. Anoxic nitric oxide cycling in plants: Participating reactions and possible mechanisms. Physiol. Plantarum 2010, 138, 393–404. [Google Scholar]
- Gupta, K.J.; Igamberdiev, A.U.; Manjunatha, G.; Segu, S.; Moran, J.F.; Neelawarne, B.; Bauwe, H.; Kaiser, W.M. The emerging roles of nitric oxide (NO) in plant mitochondria. Plant Sci 2011, 181, 520–526. [Google Scholar]
- Li, F.; Sonveaux, P.; Rabbani, Z.N.; Liu, S.; Yan, B.; Huang, Q.; Vujaskovic, Z.; Dewhirst, M.W.; Li, C.Y. Regulation of HIF-1 alpha stability through S-nitrosylation. Mol. Cell 2007, 26, 63–74. [Google Scholar]
- Ho, J.J.D.; Man, H.S.J.; Marsden, P.A. Nitric oxide signaling in hypoxia. J. Mol. Med 2012, 90, 217–231. [Google Scholar]
- Banti, V.; Loreti, E.; Novi, G.; Santaniello, A.; Alpi, A.; Perata, P. Heat acclimation and cross tolerance against anoxia in Arabidopsis. Plant Cell Environ 2008, 31, 1029–1037. [Google Scholar]
- Kim, H.S.; Snesrud, E.C.; Moy, L.P.; Haas, B.J.; Nierman, W.C.; Quackenbush, J. Transcriptional divergence of the duplicated oxidative stress-responsive genes in the Arabidopsis genome. Plant J 2005, 41, 212–220. [Google Scholar]
- Gadjev, I.; Vanderauwera, S.; Gechev, T.S.; Laloi, C.; Minkov, I.N.; Shulaev, V.; Apel, K.; Inzè, D.; Mittler, R.; van Breusegem, F. Transcriptomic footprints disclose specificity of reactive oxygen species signalling in Arabidopsis. Plant Physiol 2006, 141, 436–445. [Google Scholar]
- Van Dongen, J.T.; Fröhlic, A.; Ramìrez-Aguilar, S.J.; Schauer, N.; Fernie, A.R.; Erban, A.; Kopka, J.; Clark, J.; Langer, A.; Geigenberger, P. Transcript and metabolic profiling of the adaptive response to mild decreases in oxygen concentration in the roots of Arabidopsis plants. Ann. Bot. Lond 2008, 103, 269–280. [Google Scholar]
- Vierling, E. The role of heat shock proteins in plants. Ann. Rev. Plant Physiol 1991, 42, 579–620. [Google Scholar]
- Nishizawa, A.; Yabuta, Y.; Yoshida, E.; Maruta, T.; Yoshimura, K.; Shigeoka, S. Arabidopsis heat shock transcription factor A2 as a key regulator in response to several types of environmental stress. Plant J 2006, 48, 535–547. [Google Scholar]
- Schramm, F.; Ganguli, A.; Kiehlmann, E.; Englich, G.; Walch, D.; von Koskull-Döring, P. The heat stress transcription factor HsfA2 serves as a regulatory amplifier of a subset of genes in the heat stress response of Arabidopsis. Plant Mol. Biol 2006, 60, 759–772. [Google Scholar]
- Ogawa, D.; Yamaguchi, K.; Nishiuchi, T. High-level overexpression of the Arabidopsis HsfA2 gene confers not only increased themotolerance but also salt/osmotic stress tolerance and enhanced callus growth. J. Exp. Bot 2007, 58, 3373–3383. [Google Scholar]
- Nishizawa-Yokoi, A.; Tainaka, H.; Yoshida, E.; Tamoi, M.; Yabuta, Y.; Shigeoka, S. The 26S proteasome function and Hsp90 activity involved in the regulation of HsfA2 expression in response to oxidative stress. Plant Cell Physiol 2010, 51, 486–496. [Google Scholar]
- Nishizawa, A.; Nosaka, Y.R.; Hayashi, H.; Tainaka, H.; Maruta, T.; Tamoi, M.; Ikeda, M.; Takagi, M.O.; Yoshimura, K.; Yabuta, Y.; et al. HsfA1d and HsfA1e involved in the transcriptional regulation of HsfA2 function as key regulators for the Hsf signaling network in response to environmental stress. Plant Cell Physiol 2011, 52, 933–945. [Google Scholar]
- Yu, H.D.; Yang, X.F.; Chen, S.T.; Wang, Y.T.; Li, J.K.; Shen, Q.; Liu, X.L.; Guo, F.Q. Downregulation of chloroplast RPS1 negatively modulates nuclear heat-responsive expression of HsfA2 and its target genes in Arabidopsis. PLoS Genet 2012, 8, e1002669. [Google Scholar]
- Zhang, L.; Li, Y.; Xing, D.; Gao, C. Characterization of mitochondrial dynamics and subcellular localization of ROS reveal that HsfA2 alleviates oxidative damage caused by heat stress in Arabidopsis. J. Exp. Bot 2009, 60, 2073–2091. [Google Scholar]
- Bissoli, G.; Niñoles, R.; Fresquet, S.; Palombieri, S.; Bueso, E.; Rubio, L.; García-Sánchez, M.J.; Fernández, J.A.; Mulet, J.M.; Serrano, R. Peptidyl-prolyl cis-trans isomerase ROF2 modulates intracellular pH homeostasis in Arabidopsis. Plant J 2012, 70, 704–716. [Google Scholar]
- Pucciariello, C.; Banti, V.; Perata, P. ROS signaling as common element in low oxygen and heat stresses. Plant Physiol. Bioch 2012, 59, 3–10. [Google Scholar] [Green Version]
- Blokhina, O.B.; Chirkova, T.V.; Fagerstedt, K.V. Anoxic stress leads to hydrogen peroxide formation in plant cells. J. Exp. Bot 2001, 52, 1179–1190. [Google Scholar]
- Fukao, T.; Bailey-Serres, J. Plant responses to hypoxia—Is survival a balancing act? Trends Plant Sci 2004, 9, 449–456. [Google Scholar]
- Tsuji, H. Growth and metabolism in plants under anaerobic conditions. Environ. Control. Biol 1973, 11, 79–84. [Google Scholar]
- Huang, S.B.; Greenway, H.; Colmer, T.D. Anoxia tolerance in rice seedlings: Exogenous glucose improves growth of an anoxia-“intolerant”, but not of a “tolerant” genotype. J. Exp. Bot 2003, 54, 2363–2373. [Google Scholar]
- Murata, T.; Akazawa, T.; Fukuchi, S. Enzymic mechanism of starch breakdown in germinating rice seeds: 1. An analytical study. Plant Physiol 1968, 43, 1899–1905. [Google Scholar]
- Dunn, G. A model for starch breakdown in higher plants. Phytochemistry 1974, 13, 1341–1346. [Google Scholar]
- Sun, Z.; Henson, C.A. A quantitative assessment of the importance of barley seed α-amylase, debranching enzyme, and α-glucosidase in starch degradation. Arch. Biochem. Biophys 1991, 284, 298–305. [Google Scholar]
- Perata, P.; Pozuetaromero, J.; Akazawa, T.; Yamaguchi, J. Effect of anoxia on starch breakdown in rice and wheat seeds. Planta 1992, 188, 611–618. [Google Scholar]
- Guglielminetti, L.; Yamaguchi, J.; Perata, P.; Alpi, A. Amylolytic activities in cereal seeds under aerobic and anaerobic conditions. Plant Physiol 1995, 109, 1069–1076. [Google Scholar]
- Perata, P.; Matsukura, C.; Vernieri, P.; Yamaguchi, J. Sugar repression of a gibberellin-dependent signaling pathway in barley embryos. Plant Cell 1997, 9, 2197–2208. [Google Scholar]
- Morita, A.; Umemura, T.; Kuroyanagi, M.; Futsuhara, Y.; Perata, P.; Yamaguchi, J. Functional dissection of a sugar-repressed α-amylase gene (RAmy1A) promoter in rice embryos. FEBS Lett 1998, 423, 81–85. [Google Scholar]
- Loreti, E.; Yamaguchi, J.; Alpi, A.; Perata, P. Gibberellins are not required for rice germination under anoxia. Plant Soil 2003, 253, 137–143. [Google Scholar]
- Lu, C.A.; Ho, T.H.D.; Ho, S.L.; Yu, S.M. Three novel MYB proteins with one DNA binding repeat mediate sugar and hormone regulation of α-amylase gene expression. Plant Cell 2002, 14, 1963–1980. [Google Scholar]
- Lu, C.-A.; Lin, C.-C.; Lee, K.-W.; Chen, J.-L.; Huang, L.-F.; Ho, S.-L.; Liu, H.-J.; Hsing, Y.-I.; Yu, S.-M. The SnRK1A protein kinase plays a key role in sugar signaling during germination and seedling growth of rice. Plant Cell 2007, 19, 2484–2499. [Google Scholar]
- Lee, K.-W.; Chen, P.-W.; Lu, C.-A.; Chen, S.; Ho, T.-H.D.; Yu, S.-M. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding. Sci. Signal. 2009, 2, ra61. [Google Scholar]
- Kudahettige, N.P.; Pucciariello, C.; Parlanti, S.; Alpi, A.; Perata, P. Regulatory interplay of the Sub1A and CIPK15 pathways in the regulation of α-amylase production in flooded rice plants. Plant Biol 2011, 13, 611–619. [Google Scholar]
- Lu, C.-A.; Lim, E.-K.; Yu, S.-M. Sugar response sequence in the promoter of a rice α-amylase gene serves as a transcriptional enhancer. J. Biol. Chem 1998, 273, 10120–10131. [Google Scholar]
- Toyofuku, K.; Umemura, T; Yamaguchi, J. Promoter elements required for sugar-repression of the RAmy3D gene for α-amylase in rice. FEBS Lett. 1998, 428, 275–280. [Google Scholar]
- Chen, P.-W.; Lu, C.-A.; Yu, T.-S.; Tseng, T.-H.; Wang, C.-S.; Yu, S.-M. Rice α-amylase transcriptional enhancers direct multiple mode regulation of promoters in transgenic rice. J. Biol. Chem 2002, 277, 13641–13649. [Google Scholar]
- Cosgrove, D.J. Enzymes and other agents that enhance cell wall extensibility. Annu. Rev. Plant Physiol. Plant Mol. Biol 1999, 50, 391–417. [Google Scholar]
- Huang, J.; Takano, T.; Akita, S. Expression of α-expansin genes in young seedlings of rice (Oryza sativa L.). Planta 2000, 211, 467–473. [Google Scholar]
- Magneschi, L.; Kudahettige, R.L.; Alpi, A.; Perata, P. Expansin gene expression and anoxic coleoptile elongation in rice cultivars. J. Plant Physiol 2009, 166, 1576–1580. [Google Scholar] [Green Version]
- Choi, D.S.; Lee, Y.; Cho, H.T.; Kende, H. Regulation of expansin gene expression affects growth and development in transgenic rice plants. Plant Cell 2003, 15, 1386–1398. [Google Scholar]
- Li, Z.-X.; Septiningsih, E.M.; Quilloy-Mercado, S.M.; McNally, K.L.; Mackill, D.J. Identification of SUB1A alleles from wild rice Oryza rufipogon Griff. Genet. Resour. Crop. Ev 2011, 58, 1237–1242. [Google Scholar]
- Niroula, R.K.; Pucciariello, C.; Ho, V.T.; Novi, G.; Fukao, T.; Perata, P. SUB1A-dependent and -independent mechanisms are involved in the flooding tolerance of wild rice species. Plant J 2012, 72, 282–293. [Google Scholar]
- Vergara, B.S.; Mazaredo, A. Screening for resistance to submergence under greenhouse conditions. In Proceedings of the International Seminar on Deepwater Rice; Bangladesh Rice Research Institute: Dhaka, Bangladesh, 1975; pp. 67–70. [Google Scholar]
- Nagai, K.; Hattori, Y.; Ashikari, M. Stunt or elongate? Two opposite strategies by which rice adapts to floods. J. Plant Res 2010, 123, 303–309. [Google Scholar]
- Xu, K.; Mackill, D.J. A major locus for submergence tolerance mapped on rice chromosome 9. Mol. Breed 1996, 2, 219–224. [Google Scholar]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A variable cluster of ethylene response factor- like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar]
- Fukao, T.; Bailey-Serres, J. Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice. Proc. Natl. Acad. Sci. USA 2008, 105, 16814–16819. [Google Scholar]
- Septiningsih, E.M.; Pamplona, A.M.; Sanchez, D.L.; Neeraja, C.N.; Vergara, G.V.; Heuer, S.; Ismail, A.M.; Mackill, D.J. Development of submergence-tolerant rice cultivars: The Sub1 locus and beyond. Ann. Bot 2009, 103, 151–160. [Google Scholar]
- Singh, N.; Dang, T.; Vergara, G.; Pandey, D.; Sanchez, D.; Neeraja, C.; Septiningsih, E.; Mendioro, M.; Tecson-Mendoza, R.; Ismal, A.; et al. Molecular marker survey and expression analyses of the rice submergence-tolerance genes SUB1A and SUB1C. Theor. Appl. Genet 2010, 121, 1441–1453. [Google Scholar]
- Grossman, A.R.; Catalanotti, C.; Yang, W.; Dubini, A.; Magneschi, L.; Subramanian, V.; Posewitz, M.C.; Seibert, M. Multiple facets of anoxic metabolism and hydrogen production in the unicellular green alga Chlamydomonas reinhardtii. New Phytol 2011, 190, 279–288. [Google Scholar]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 2007, 318, 245–250. [Google Scholar]
- Harris, E.H. Chlamydomonas as a model organism. Annu. Rev. Plant Physiol. Plant Mol. Biol 2001, 52, 363–406. [Google Scholar]
- Grossman, A.R. In the grip of algal genomics. Adv. Exp. Med. Biol 2007, 616, 54–76. [Google Scholar]
- Purton, S. Tools and techniques for chloroplast transformation of Chlamydomonas Adv. Exp. Med. Biol 2007, 616, 34–45. [Google Scholar]
- Molnar, A.; Bassett, A.; Thuenemann, E.; Schwach, F.; Karkare, S.; Ossowski, S.; Weigel, D.; Baulcombe, D. Highly specific gene silencing by artificial microRNAs in the unicellular alga Chlamydomonas reinhardtii. Plant J 2009, 58, 165–174. [Google Scholar]
- Gonzalez-Ballester, D.; Pootakham, W.; Mus, F.; Yang, W.; Catalanotti, C.; Magneschi, L.; de Montaigu, A.; Higuera, J.J.; Prior, M.; Galván, A.; et al. Reverse genetics in Chlamydomonas, a platform for isolating insertional mutants. Plant Methods 2011, 7, 24. [Google Scholar]
- Pootakham, W.; Gonzalez-Ballester, D.; Grossman, A.R. Identification and regulation of plasma membrane sulfate transporters in Chlamydomonas. Plant Physiol 2010, 153, 1653–1668. [Google Scholar]
- Schmollinger, S.; Strenkert, D.; Schroda, M. An inducible artificial microRNA system for Chlamydomonas reinhardtii confirms a key role for heat shock factor 1 in regulating thermotolerance. Curr. Genet 2010, 56, 383–389. [Google Scholar]
- Godman, J.E.; Molnár, A.; Baulcombe, D.C.; Balk, J. RNA silencing of hydrogenase(-like) genes and investigation of their physiological roles in the green alga Chlamydomonas reinhardtii. Biochem. J 2010, 431, 345–351. [Google Scholar]
- Meuser, J.E.; D’Adamo, S.; Jinkerson, R.E.; Mus, F.; Yang, W.; Ghirardi, M.L.; Seibert, M.; Grossman, A.R.; Posewitz, M.C. Genetic disruption of both Chlamydomonas reinhardtii [FeFe]-hydrogenases, insight into the role of HYDA2 in H2 production. Biochem. Biophys. Res. Commun 2012, 417, 704–709. [Google Scholar]
- Forestier, M.; King, P.; Zhang, L.; Posewitz, M.; Schwarzer, S.; Happe, T.; Ghirardi, M.L.; Seibert, M. Expression of two [Fe]-hydrogenases in Chlamydomonas reinhardtii under anaerobic conditions. Eur. J. Biochem 2003, 270, 2750–2758. [Google Scholar]
- Happe, T.; Kaminski, A. Differential regulation of the Fe-hydrogenase during anaerobic adaptation in the green alga Chlamydomonas reinhardtii. Eur. J. Biochem 2002, 269, 1022–1032. [Google Scholar]
- Posewitz, M.C.; King, P.W.; Smolinski, S.L.; Zhang, L.; Seibert, M.; Ghirardi, M.L. Discovery of two novel radical S-adenosylmethionine proteins required for the assembly of an active [Fe]-hydrogenase. J. Biol. Chem 2004, 279, 25711–25720. [Google Scholar]
- Posewitz, M.C.; King, P.W.; Smolinski, S.L.; Smith, R.D.; Ginley, A.R.; Ghirardi, M.L.; Seibert, M. Identification of genes required for hydrogenase activity in Chlamydomonas reinhardtii. Biochem. Soc. Trans 2005, 33, 102–104. [Google Scholar]
- Atteia, A.; van Lis, R.; Gelius-Dietrich, G.; Adrait, A.; Garin, J.; Joyard, J.; Rolland, N.; Martin, W. Pyruvate formate-lyase and a novel route of eukaryotic ATP synthesis in Chlamydomonas mitochondria. J. Biol. Chem 2006, 281, 9909–9918. [Google Scholar]
- Terashima, M.; Specht, M.; Naumann, B.; Hippler, M. Characterizing the anaerobic response of Chlamydomonas reinhardtii by quantitative proteomics. Mol. Cell Proteomics 2010, 9, 1514–1532. [Google Scholar]
- Philipps, G.; Krawietz, D.; Hemschemeier, A.; Happe, T. A pyruvate formate lyase-deficient Chlamydomonas reinhardtii strain provides evidence for a link between fermentation and hydrogen production in green algae. Plant J 2011, 66, 330–340. [Google Scholar]
- Terashima, M.; Specht, M.; Hippler, M. The chloroplast proteome, a survey from the Chlamydomonas reinhardtii perspective with a focus on distinctive features. Curr. Genet 2011, 57, 151–168. [Google Scholar]
- Catalanotti, C.; Dubini, A.; Subramanian, V.; Yang, W.; Magneschi, L.; Mus, F.; Seibert, M.; Posewitz, M.C.; Grossman, A.R. Altered fermentative metabolism in Chlamydomonas reinhardtii mutants lacking pyruvate formate lyase and both pyruvate formate lyase and alcohol dehydrogenase. Plant Cell 2012, 24, 692–707. [Google Scholar]
- Magneschi, L.; Catalanotti, C.; Subramanian, V.; Dubini, A.; Yang, W.; Mus, F.; Posewitz, M.C.; Seibert, M.; Perata, P.; Grossman, A.R. A mutant in the ADH1 gene of Chlamydomonas reinhardtii elicits metabolic restructuring during anaerobiosis. Plant Physiol 2012, 158, 1293–1305. [Google Scholar]
- Gfeller, R.P.; Gibbs, M. Fermentative metabolism of Chlamydomonas reinhardtii, I. Analysis of fermentative products from starch in dark and light. Plant Physiol 1984, 75, 212–218. [Google Scholar]
- Kreuzberg, K. Starch fermentation via formate producing pathway in Chlamydomonas reinhardtii; Chlorogonium elongatum and Chlorella fusca. Physiol. Plantarum 1984, 61, 87–94. [Google Scholar]
- Ohta, S.; Miyamoto, K.; Miura, Y. Hydrogen evolution as a consumption mode of reducing equivalents in green algal fermentation. Plant Physiol 1987, 83, 1022–1026. [Google Scholar]
- Tsygankov, A.A.; Kosourov, S.; Seibert, M.; Ghirardi, M.L. Hydrogen photoproduction under continuous illumination by sulfur-deprived, synchronous Chlamydomonas reinhardtii cultures. Int. J. Hydrog. Energ 2002, 27, 1239–1244. [Google Scholar]
- Kosourov, S.; Seibert, M.; Ghirardi, M.L. Effects of extracellular pH on the metabolic pathways in sulfur-deprived; H2-producing Chlamydomonas reinhardtii cultures. Plant Cell Physiol 2003, 44, 146–155. [Google Scholar]
- Dubini, A.; Mus, F.; Seibert, M.; Grossman, A.R.; Posewitz, M.C. Flexibility in anaerobic metabolism as revealed in a mutant of Chlamydomonas reinhardtii lacking hydrogenase activity. J. Biol. Chem 2009, 284, 7201–7213. [Google Scholar]
- Melis, A.; Zhang, L.; Forestier, M.; Ghirardi, M.L.; Seibert, M. Sustained photobiological hydrogen gas production upon reversible inactivation of oxygen evolution in the green alga Chlamydomonas reinhardtii. Plant Physiol 2000, 122, 127–136. [Google Scholar]
- Tolleter, D.; Ghysels, B.; Alric, J.; Petroutsos, D.; Tolstygina, I.; Krawietz, D.; Happe, T.; Auroy, P.; Adriano, JM.; Beyly, A.; et al. Plant Cell 2011, 23, 2619–2630.
- Terashima, M.; Petroutsos, D.; Hüdig, M.; Tolstygina, I.; Trompelt, K.; Gäbelein, P.; Fufezan, C.; Kudla, J.; Weinl, S.; Finazzi, G.; et al. Calcium-dependent regulation of cyclic photosynthetic electron transfer by a CAS, ANR1, and PGRL1 complex. Proc. Natl. Acad. Sci. USA 2012, 109, 17717–17722. [Google Scholar]
- Antal, T.K.; Krendeleva, T.E.; Laurinavichene, T.V.; Makarova, V.V.; Ghirardi, M.L.; Rubin, A.B.; Tsygankov, A.A.; Seibert, M. The dependence of algal H2 production on Photosystem II and O2 consumption activities in sulfur-deprived Chlamydomonas reinhardtii cells. BBA 2003, 1607, 153–160. [Google Scholar]
- Bailey-Serres, J.; Chang, R. Sensing and signalling in response to oxygen deprivation in plants and other organisms. Ann. Bot 2005, 96, 519–532. [Google Scholar]
- Guzy, R.D.; Schumacker, P.T. Oxygen sensing by mitochondria at complex III, the paradox of increased reactive oxygen species during hypoxia. Exp. Physiol 2006, 91, 807–819. [Google Scholar]
- Bell, E.L.; Chandel, N.S. Mitochondrial oxygen sensing, regulation of hypoxia-inducible factor by mitochondrial generated reactive oxygen species. Essays Biochem 2007, 43, 17–27. [Google Scholar]
- Whitney, L.A.; Loreti, E.; Alpi, A.; Perata, P. Alcohol dehydrogenase and hydrogenase transcript fluctuations during a day-night cycle in Chlamydomonas reinhardtii, the role of anoxia. New Phytol 2011, 190, 488–498. [Google Scholar]
- Whitney, L.A.; Novi, G.; Perata, P.; Loreti, E. Distinct mechanisms regulating gene expression coexist within the fermentative pathways in Chlamydomonas reinhardtii. ScientificWorldJournal 2012. [Google Scholar] [CrossRef]
- Quinn, J.M.; Eriksson, M.; Moseley, J.L.; Merchant, S. Oxygen deficiency responsive gene expression in Chlamydomonas reinhardtii through a copper-sensing signal transduction pathway. Plant Physiol 2002, 128, 463–471. [Google Scholar]
- Eriksson, M.; Moseley, J.L.; Tottey, S.; Del Campo, J.A.; Quinn, J.; Kim, Y.; Merchant, S. Genetic dissection of nutritional copper signaling in Chlamydomonas distinguishes regulatory and target genes. Genetics 2004, 168, 795–807. [Google Scholar]
- Kropat, J.; Tottey, S.; Birkenbihl, R.P.; Depège, N.; Huijser, P.; Merchant, S. A regulator of nutritional copper signaling in Chlamydomonas is an SBP domain protein that recognizes the GTAC core of copper response element. Proc. Natl. Acad. Sci. USA 2005, 102, 18730–18735. [Google Scholar]
- Lambertz, C.; Hemschemeier, A.; Happe, T. Anaerobic expression of the ferredoxin-encoding FDX5 gene of Chlamydomonas reinhardtii is regulated by the Crr1 transcription factor. Eukaryot. Cell 2010, 9, 1747–1754. [Google Scholar]
- Pape, M.; Lambertz, C.; Happe, T.; Hemschemeier, A. Differential expression of the Chlamydomonas [FeFe]-hydrogenase-encoding HYDA1 gene is regulated by the copper response regulator 1. Plant Physiol 2012, 159, 1700–1712. [Google Scholar]
- Castruita, M.; Casero, D.; Karpowicz, S.J.; Kropat, J.; Vieler, A.; Hsieh, S.I.; Yan, W.; Cokus, S.; Loo, J.A.; Benning, C.; et al. Systems biology approach in Chlamydomonas reveals connections between copper nutrition and multiple metabolic steps. Plant Cell 2011, 23, 1273–1292. [Google Scholar]
- Quinn, J.M.; Barraco, P.; Eriksson, M.; Merchant, S. Coordinate copper- and oxygen-responsive Cyc6 and Cpx1 expression in Chlamydomonas is mediated by the same element. J. Biol. Chem 2000, 275, 6080–6089. [Google Scholar]



© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Banti, V.; Giuntoli, B.; Gonzali, S.; Loreti, E.; Magneschi, L.; Novi, G.; Paparelli, E.; Parlanti, S.; Pucciariello, C.; Santaniello, A.; et al. Low Oxygen Response Mechanisms in Green Organisms. Int. J. Mol. Sci. 2013, 14, 4734-4761. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14034734
Banti V, Giuntoli B, Gonzali S, Loreti E, Magneschi L, Novi G, Paparelli E, Parlanti S, Pucciariello C, Santaniello A, et al. Low Oxygen Response Mechanisms in Green Organisms. International Journal of Molecular Sciences. 2013; 14(3):4734-4761. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14034734
Chicago/Turabian StyleBanti, Valeria, Beatrice Giuntoli, Silvia Gonzali, Elena Loreti, Leonardo Magneschi, Giacomo Novi, Eleonora Paparelli, Sandro Parlanti, Chiara Pucciariello, Antonietta Santaniello, and et al. 2013. "Low Oxygen Response Mechanisms in Green Organisms" International Journal of Molecular Sciences 14, no. 3: 4734-4761. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14034734