Epigenetic Silencing of DKK3 in Medulloblastoma

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
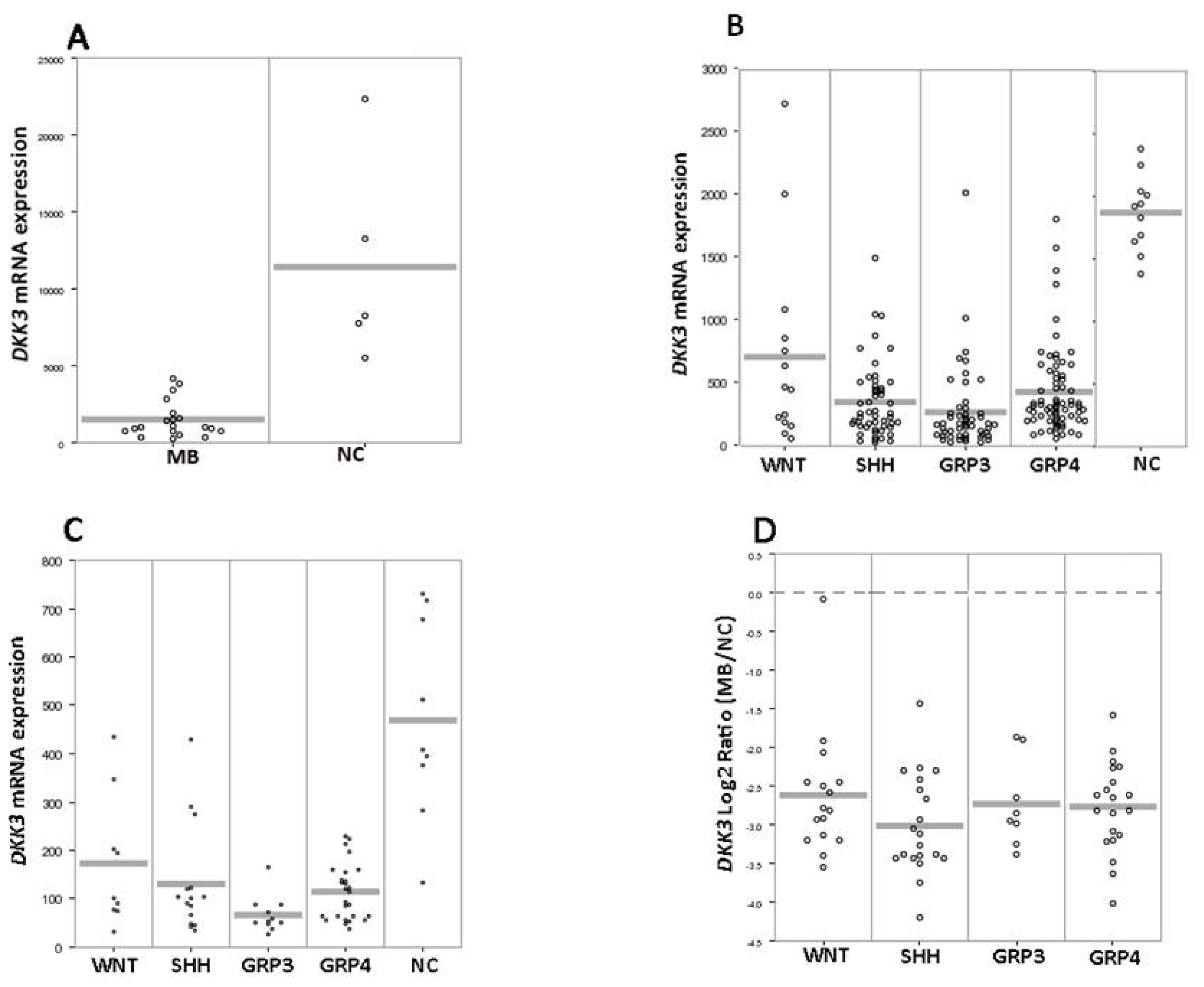
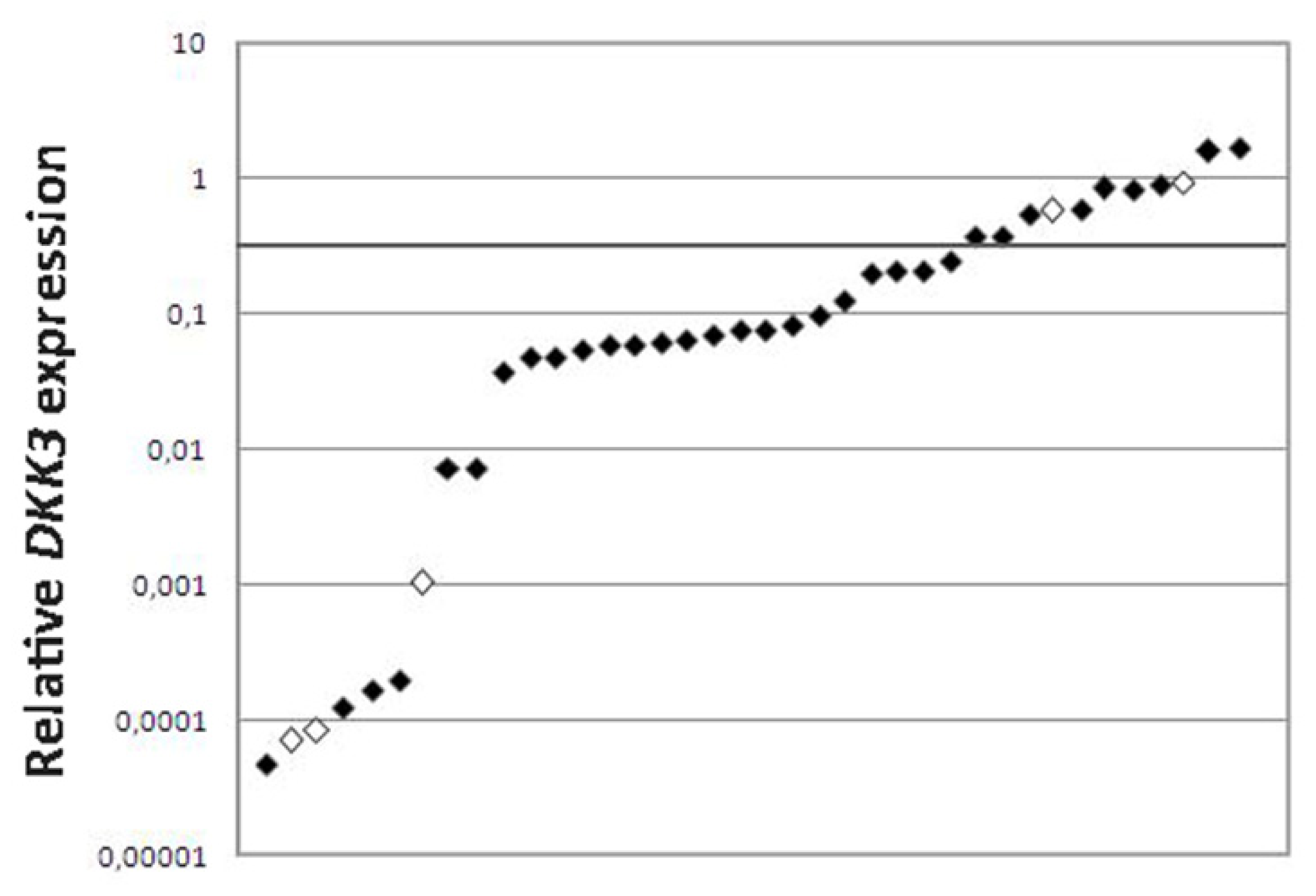
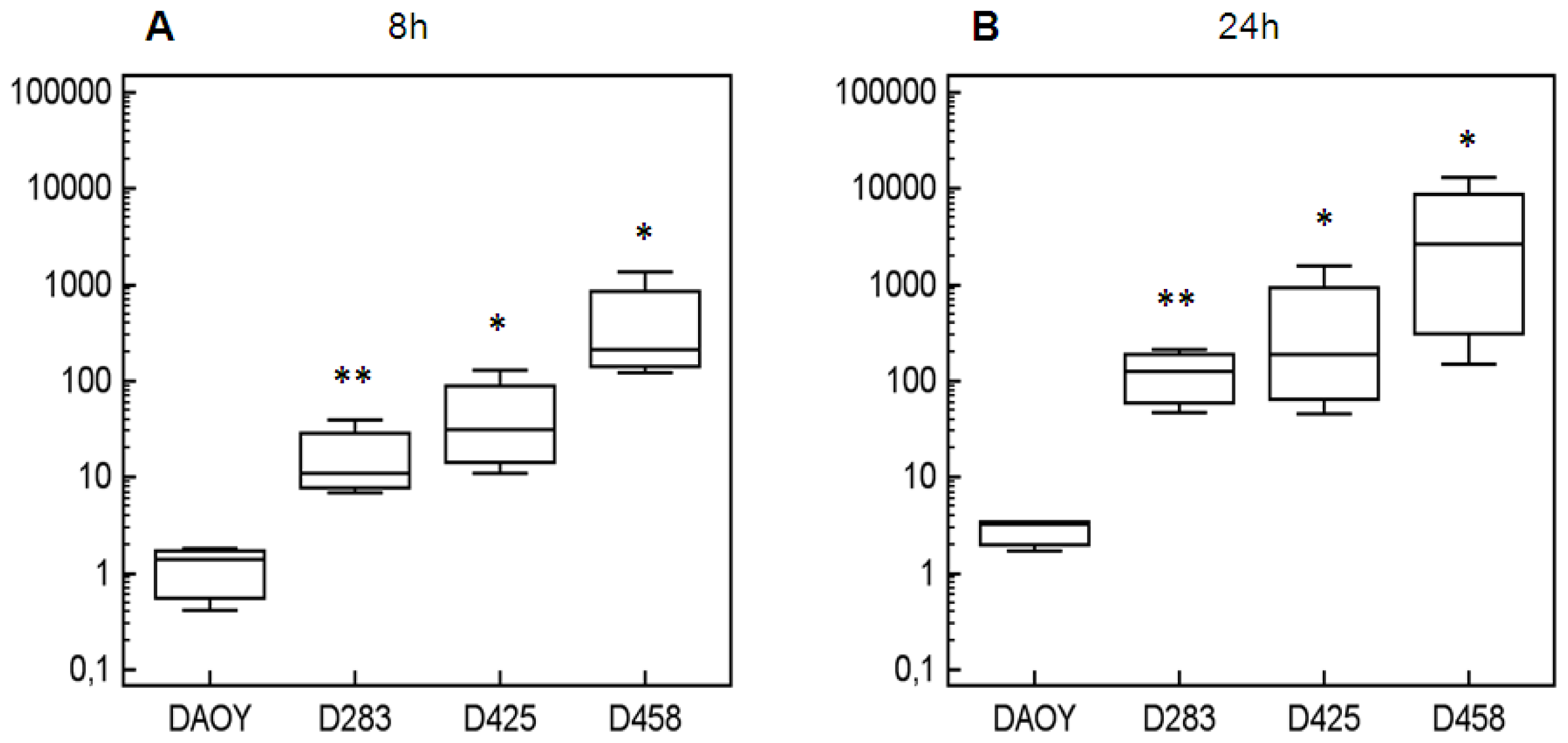
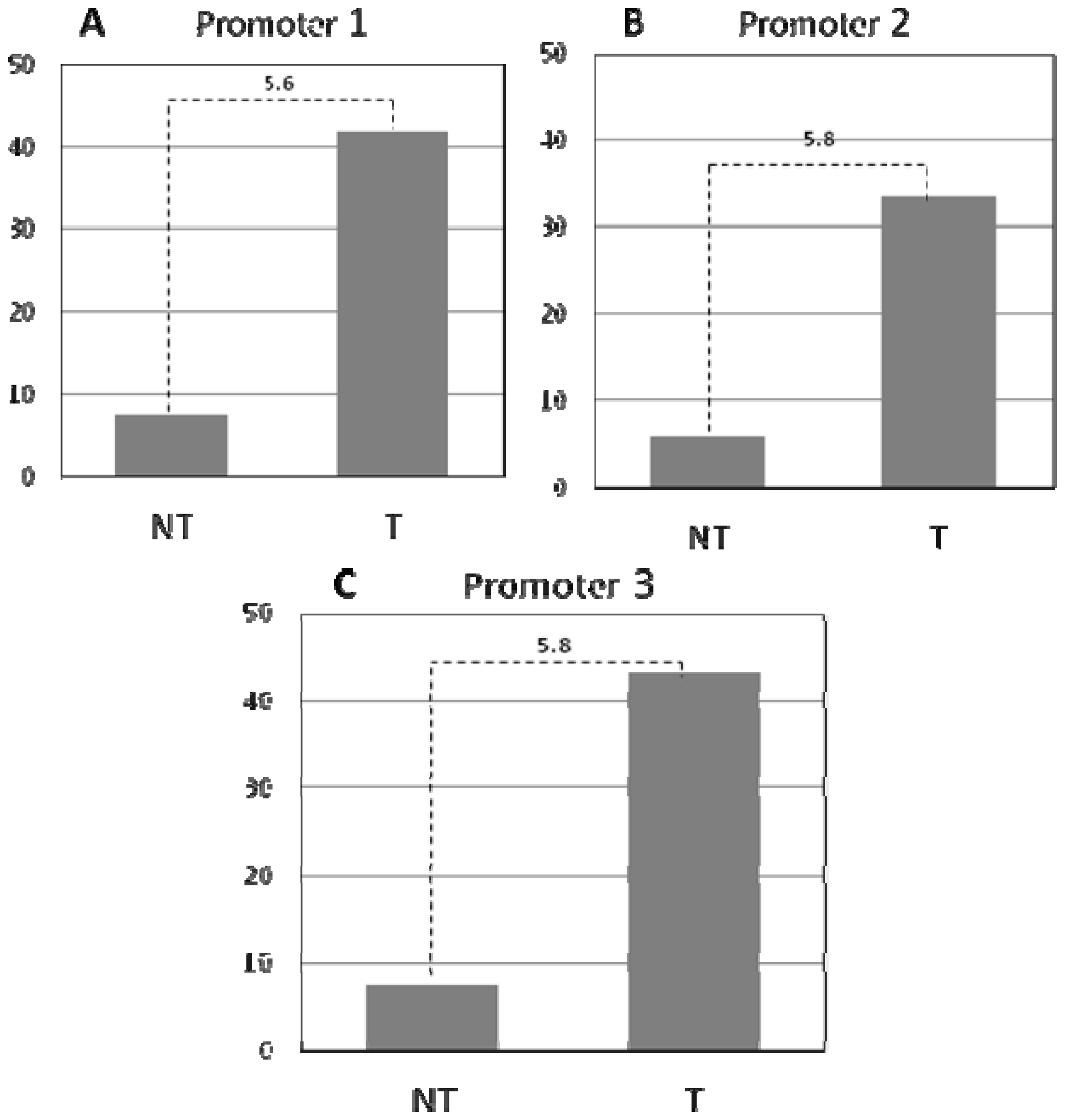
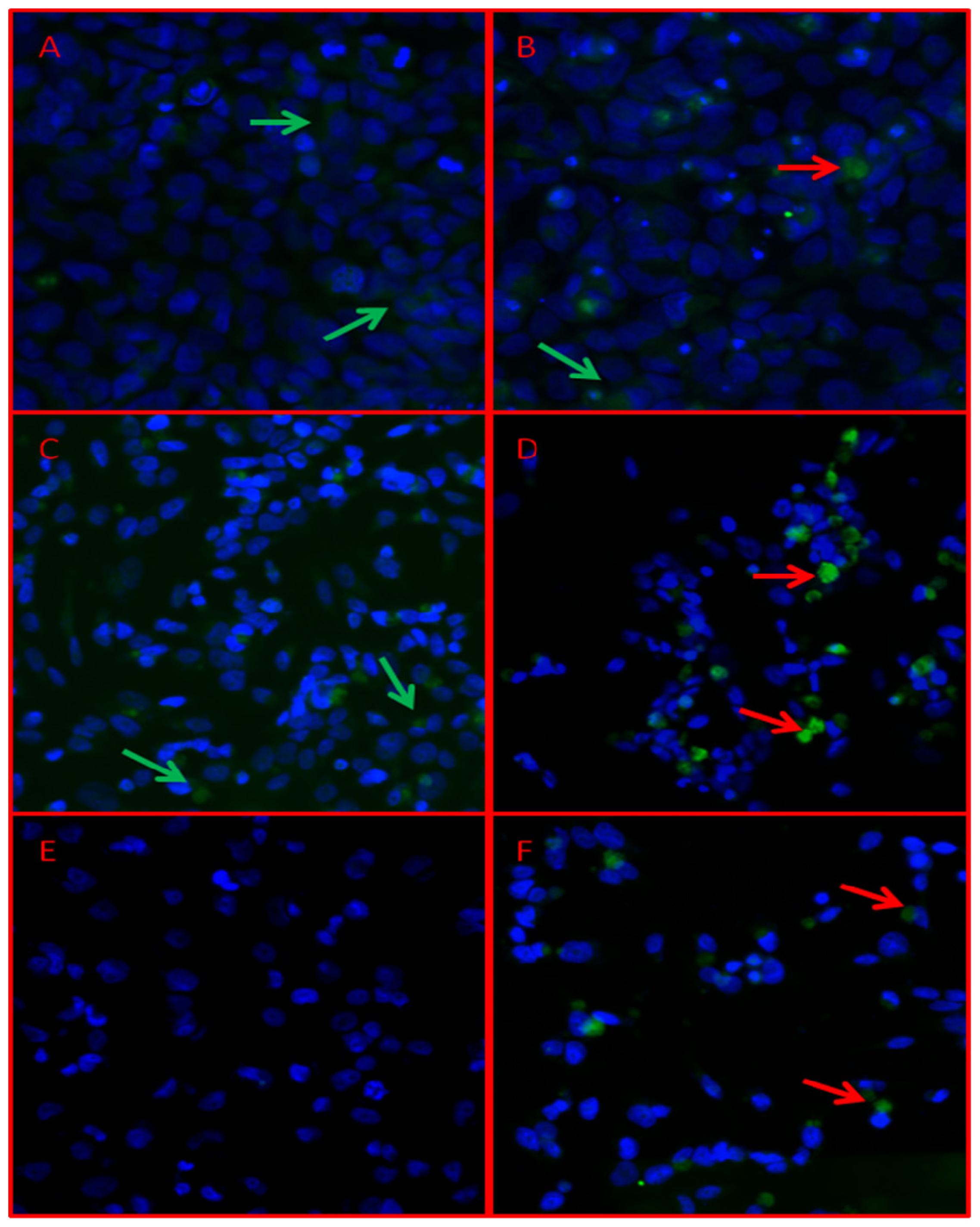
2. Results and Discussion
3. Experimental Section
3.1. Tumor Samples and Cell Lines and Acid Nucleic Isolation
3.2. Gene Expression Profiling, Array-CGH Data and miRNA
3.3. qPCR Analysis
3.4. Trichostatin a Treatment and Chromatin Immuno-Precipitation (ChIP) Assay
3.5. Methylation Analysis
3.6. Immunofluorescence Analysis
4. Conclusions
Supplementary Information
ijms-14-07492-s001.pdfAcknowledgments
Conflict of Interest
References
- Packer, R.J.; Cogen, P.; Vezina, G.; Rorke, L.B. Medulloblastoma: Clinical and biologic aspects. Neuro Oncol 1999, 1, 232–250. [Google Scholar]
- Kool, M.; Koster, J.; Bunt, J.; Hasselt, N.E.; Lakeman, A.; van Sluis, P.; Troost, D.; Meeteren, N.S.; Caron, H.N.; Cloos, J.; et al. Integrated genomics identifies five medulloblastoma subtypes with distinct genetic profiles, pathway signatures and clinicopathological features. PLoS One 2008, 3, 3088. [Google Scholar]
- Northcott, P.A.; Korshunov, A.; Witt, H.; Hielscher, T.; Eberhart, C.G.; Mack, S.; Bouffet, E.; Clifford, S.C.; Hawkins, C.E.; French, P.; et al. Medulloblastoma comprises four distinct molecular variants. J. Clin. Oncol 2011, 10, 1408–1414. [Google Scholar]
- Cho, Y.J.; Tsherniak, A.; Tamayo, P.; Santagata, S.; Ligon, A.; Greulich, H.; Berhoukim, R.; Amani, V.; Goumnerova, L.; Eberhart, C.G.; et al. Integrative genomic analysis of medulloblastoma identifies a molecular subgroup that drives poor clinical outcome. J. Clin. Oncol 2011, 29, 1424–1430. [Google Scholar]
- Remke, M.; Hielscher, T.; Korshunov, A.; Nothcott, P.A.; Bender, S.; Kool, M.; Westermann, F.; Benner, A.; Ryzhova, M.; Sturm, D.; et al. FSTL5 is a marker of poor prognosis in non-Wnt/non.SHH Medulloblastoma. J. Clin. Oncol 2011, 29, 3852–3861. [Google Scholar]
- Polkinghorn, W.R.; Tarbel, N.J. Medulloblastoma: Tumorigenesis, current clinical paradigm, and efforts to improve risk stratification. Nat. Clin. Pract. Oncol 2007, 4, 295–304. [Google Scholar]
- Taylor, M.D.; Northcott, P.A.; Korshunov, A.; Remke, M.; Cho, Y.J.; Clifford, S.C.; Eberhart, C.G.; Parsons, D.W.; Rutkowski, S.; Gajjar, A.; et al. Molecular subgroups of medulloblastoma: The current consensus. Acta Neuropathol 2012, 123, 465–472. [Google Scholar]
- Kool, M.; Korshunov, A.; Remke, M.; Jones, D.T.; Schlanstein, M.; Northcott, P.A.; Cho, Y.J.; Koster, J.; Schouten-van Meeteren, A.; van Vuurden, D.; et al. Molecular subgroups of medulloblastoma: An international meta-analysis of transcriptome, genetic aberrations, and clinical data of WNT, SHH, Group 3, and Group 4 medulloblastomas. Acta Neuropathol 2012, 123, 473–484. [Google Scholar]
- Marino, S. Medulloblastoma: Developmental mechanisms out of control. Trends Mol. Med 2005, 11, 17–22. [Google Scholar]
- Kawano, Y.; Kypta, R. Secreted antagonists of the Wnt signalling pathway. J. Cell Sci 2003, 116, 2627–2634. [Google Scholar]
- Katoh, M.; Katoh, M. WNT signaling pathway and stem cell signaling network. Clin. Cancer Res 2007, 13, 4042–4045. [Google Scholar]
- Kobayashi, K.; Ouchida, M.; Tsuji, T.; Hanafusa, H.; Miyazaki, M.; Namba, M.; Shimizu, N.; Shimizu, K. Reduced expression of the REIC/Dkk3 gene by promoter-hypermethylation in human tumor cells. Gene 2002, 2, 151–158. [Google Scholar]
- Hsieh, S.Y.; Hsieh, P.S.; Chiu, C.-T.; Chen, W.Y. Dickkopf-3/REIC functions as a suppressor gene of tumor growth. Oncogene 2004, 2, 9183–9189. [Google Scholar]
- Kuphal, S.; Lodermeyer, S.; Bataille, F.; Schuierer, M.; Hoang, B.H.; Bosserhoff, A.K. Expression of Dickkopf genes is strongly reduced in malignant Melanoma. Oncogene 2006, 25, 5027–5036. [Google Scholar]
- Chung, M.T.; Lai, H.C.; Sytwu, H.K.; Yan, M.D.; Shih, Y.L.; Chang, C.C.; Yu, M.H.; Liu, H.S.; Chu, D.W.; Lin, Y.W. SFRP1 and SFRP2 suppress the transformation and invasion abilities of cervical cancer cells through Wnt signaling pathway. Ginecol. Oncol 2009, 112, 646–653. [Google Scholar]
- Sato, H.; Suzuki, H.; Toyotak, M.; Nojima, M.; Maruyama, R.; Sasaki, S.; Takagi, H.; Sogabe, Y.; Sasaki, Y.; Idogawa, M.; et al. Frequent epigenetic inactivation of DICKKOPF family genes in human gastrointestinal tumors. Carcinogenesis 2007, 28, 2459–2466. [Google Scholar]
- Vibhakar, R.; Foltz, G.; Yoon, J.G.; Field, L.; Lee, H.; Ryu, G.Y.; Pierson, J.; Davidson, B.; Madan, A. Dickkopf-1 is an epigenetically silenced candidate tumor suppressor gene in medulloblastoma. Neuro Oncol 2007, 4, 135–144. [Google Scholar]
- Facchetti, F.; Previdi, S.; Ballarini, M.; Minucci, S.; Perego, P.; La Porta, C.A. Modulation of pro-and anti-apoptotic factors in human melanoma cells exposed to histone deacetylase inhibitors. Apoptosis 2004, 9, 573–582. [Google Scholar]
- Yoshikawa, M.; Hishikawa, K.; Idei, M.; Fujita, T. Trichostatin a prevents TGF-beta1-induced apoptosis by inhibiting ERK activation in human renal tubular epithelial cells. Eur. J. Pharmacol 2010, 642, 28–36. [Google Scholar]
- Roth, R.B.; Hevezi, P.; Lee, J.; Willhite, D.; Lechner, S.M.; Foster, A.C.; Zlotnik, A. Gene expression analyses reveal molecular relationships among 20 regions of the human CNS. Neurogenetics 2006, 7, 67–80. [Google Scholar]
- Northcott, P.A.; Shih, D.J.; Peacock, J.; Garzia, L.; Morrissy, A.S.; Zichner, T.; Stütz, A.M.; Korshunov, A.; Reimand, J.; Schumacher, S.E.; et al. Subgroup-specific structural variation across 1000 medulloblastoma genomes. Nature 2012, 488, 49–56. [Google Scholar]
- Haug, B.H.; Henriksen, J.R.; Buechner, J.; Geerts, D.; Tømte, E.; Kogner, P.; Martinsson, T.; Flægstad, T.; Sveinbjørnsson, B.; Einvik, C. MYCN-regulated miRNA-92 inhibits secretion of the tumor suppressor DICKKOPF-3 (DKK3) in neuroblastoma. Carcinogenesis 2011, 32, 1005–1012. [Google Scholar]
- Aldosari, N.; Bigner, S.H.; Burger, P.C.; Becker, L.; Kepner, J.L.; Friedman, H.S.; McLendon, R.E. MYCC and MYCN oncogene amplification in medulloblastoma. A fluorescence in situ hybridization study on paraffin sections from the Children’s Oncology Group. Arch. Pathol. Lab. Med 2002, 126, 540–544. [Google Scholar]
- Kongkham, P.N.; Northcott, P.A.; Croul, S.E.; Smith, C.A.; Taylor, M.D.; Rutka, J.T. The SFRP family of WNT inhibitors function as novel tumor suppressor genes epigenetically silenced in medulloblastoma. Oncogene 2010, 29, 3017–3024. [Google Scholar]
- Häcker, S.; Dittrich, A.; Mohr, A.; Schweitzer, T.; Rutkowski, S.; Krauss, J.; Debatin, K.M.; Fulda, S. Histone deacetylase inhibitors cooperate with IFN-gamma to restore caspase-8 expression and overcome TRAIL resistance in cancers with silencing of caspase-8. Oncogene 2009, 28, 3097–3110. [Google Scholar]
- Yang, B.; Du, Z.; Gao, Y.T.; Lou, C.; Zhang, S.G.; Bai, T.; Wang, Y.J.; Song, W.Q. Methylation of Dickkopf-3 as a prognostic factor in cirrhosis-related hepatocellular carcinoma. World J. Gastroenterol 2010, 16, 755–763. [Google Scholar]
- Roman-Gomez, J.; Jimenez-Velasco, A.; Agirre, X.; Castillejo, J.A.; Navarro, G.; Barrios, M.; Andreu, E.J.; Prosper, F.; Heiniger, A.; Torres, A. Transcriptional silencing of the Dickkopfs-3 (Dkk-3) gene by CpG hypermethylation in acute lymphoblastic leukaemia. Br. J. Cancer 2004, 91, 707–713. [Google Scholar]
- Veeck, J.; Bektas, N.; Hartmann, A.; Kristiansen, G.; Heindrichs, U.; Knuchel, R.; Dahl, E. Wnt signalling in human breast cancer: Expression of the putative Wnt inhibitor Dickkopf-3 (DKK3) is frequently suppressed by promoter hypermethylation in mammary tumours. Breast Cancer Res 2008, 10, R82. [Google Scholar]
- Cameron, E.E.; Bachman, K.E.; Myöhänen, S.; Herman, J.G.; Baylin, S.B. Synergy of demethylation and histone deacetylase inhibition in the re-expression of genes silenced in cancer. Nat. Genet 1999, 21, 103–107. [Google Scholar]
- Brigati, C.; Banelli, B.; Casciano, I.; di Vinci, A.; Matis, S.; Cutrona, G.; Forlani, A.; Allemanni, G.; Romani, M. Epigenetic mechanisms regulate ΔNP73 promoter function in human tonsil B cells. Mol. Immunol 2011, 48, 408–414. [Google Scholar]
- Coco, S.; Valdora, F.; Bonassi, S.; Scaruffi, P.; Stigliani, S.; Oberthuer, A.; Berthold, F.; Adolfo, I.; Servidei, T.; Riccardi, R.; et al. Chromosome 9q and 16q loss identified by genome-wide pooled analysis are associated with tumor aggressiveness in patients with classic medulloblastoma. J. Integr. Biol 2011, 15, 273–280. [Google Scholar]
- Pfister, S.M.; Remke, M.; Benner, A.; Mendrzyk, F.; Toedt, G.; Felsberg, J.; Wittmann, A.; Devens, F.; Gerber, N.U.; Joos, S.; et al. Outcome prediction in pediatric medulloblastoma based on DNA copy-number aberrations of chromosomes 6q and 17q and the MYC and MYCN loci. J. Clin. Oncol 2009, 27, 1627–1636. [Google Scholar]
- Banelli, B.; Bonassi, S.; Casciano, I.; Mazzocco, K.; di Vinci, A.; Scaruffi, P.; Brigati, C.; Allemanni, G.; Borzì, L.; Tonini, G.P.; et al. Outcome prediction and risk assessment by quantitative pyrosequencing methylation analysis of the SFN gene advanced stage, high-risk, neuroblastic tumor patients. Int. J. Cancer 2010, 126, 656–668. [Google Scholar]
- Del Grosso, F.; Coco, S.; Scaruffi, P.; Stigliani, S.; Valdora, F.; Benelli, R.; Salvi, S.; Boccardo, S.; Truini, M.; Croce, M.; et al. Role of CXCL13-CXCR5 crosstalk between malignant neuroblastoma cells and Schwannian stromal cells in neuroblastic tumors. Mol. Cancer Res 2011, 9, 815–823. [Google Scholar]
- Giangaspero, F.; Eberhart, C.; Haapasalo, H.; Pietsch, T.; Wiestler, O.D.; Ellison, D.W. Medulloblastoma. In WHO Classification of Tumours of the Central Nervous System; Louis, D.N., Ohgaki, H., Wiestler, O.D., Cavenee, W.K., Eds.; IARC Press: Lyon, France, 2007. [Google Scholar]
- Chang, C.H.; Housepian, E.M.; Herbert, C., Jr. An operative staging system and a megavoltage radiotherapeutic technic for cerebellar medulloblastomas. Radiology 1969, 93, 13561–1359. [Google Scholar]





© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Valdora, F.; Banelli, B.; Stigliani, S.; Pfister, S.M.; Moretti, S.; Kool, M.; Remke, M.; Bai, A.H.C.; Brigati, C.; Hielscher, T.; et al. Epigenetic Silencing of DKK3 in Medulloblastoma. Int. J. Mol. Sci. 2013, 14, 7492-7505. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14047492
Valdora F, Banelli B, Stigliani S, Pfister SM, Moretti S, Kool M, Remke M, Bai AHC, Brigati C, Hielscher T, et al. Epigenetic Silencing of DKK3 in Medulloblastoma. International Journal of Molecular Sciences. 2013; 14(4):7492-7505. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14047492
Chicago/Turabian StyleValdora, Francesca, Barbara Banelli, Sara Stigliani, Stefan M. Pfister, Stefano Moretti, Marcel Kool, Marc Remke, Alfa H.C. Bai, Claudio Brigati, Thomas Hielscher, and et al. 2013. "Epigenetic Silencing of DKK3 in Medulloblastoma" International Journal of Molecular Sciences 14, no. 4: 7492-7505. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14047492