MiR199b Suppresses Expression of Hypoxia-Inducible Factor 1α (HIF-1α) in Prostate Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
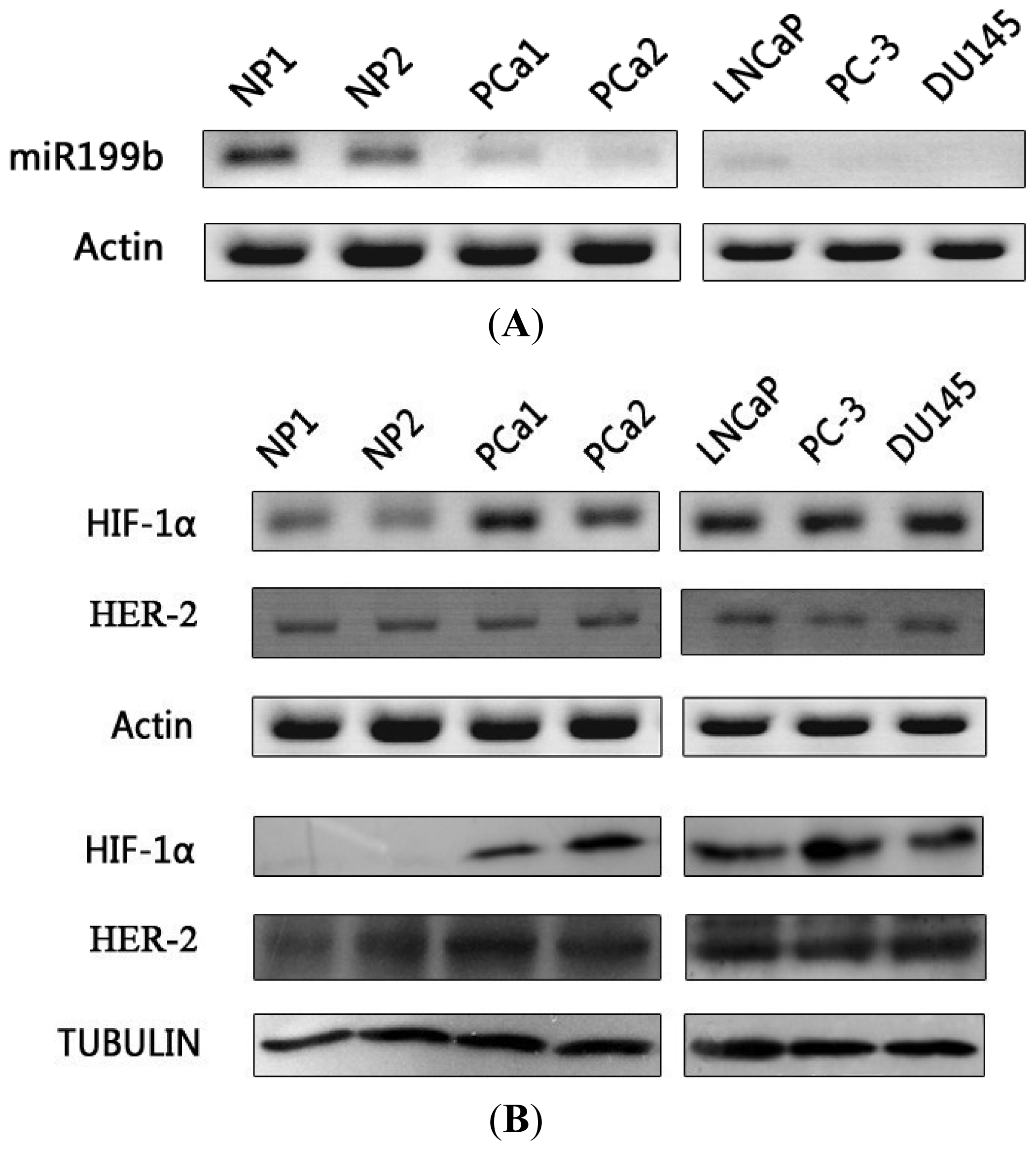
2.1. Expression of miR199b and HIF-1α in Prostate Cancer
2.2. Identification of Potential HIF-1α 3′-UTR Seed Sequences
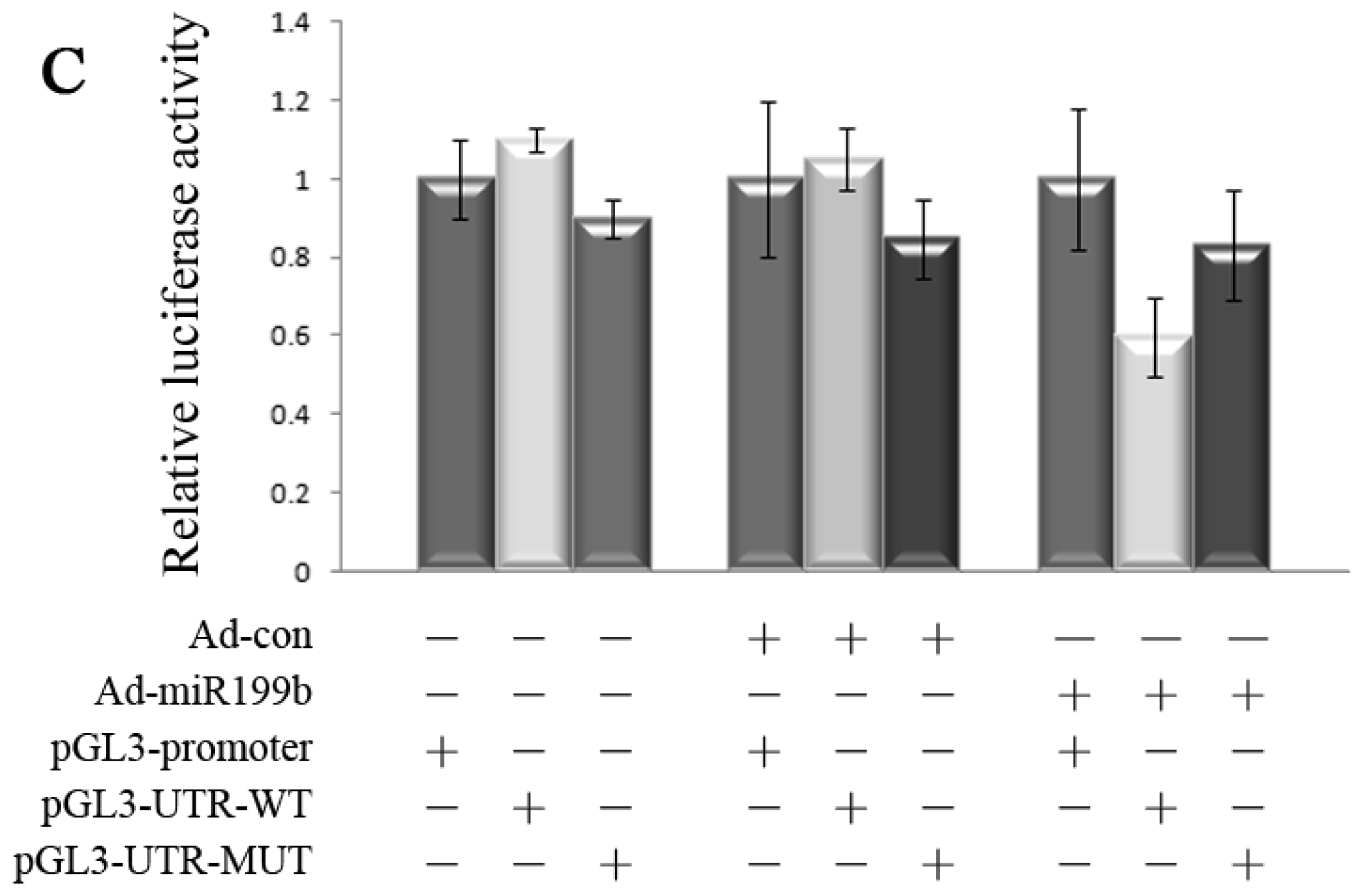
2.3. Dual Reporter Gene Assays Showed Interaction of miR199b with 3′-UTR of HIF-1α
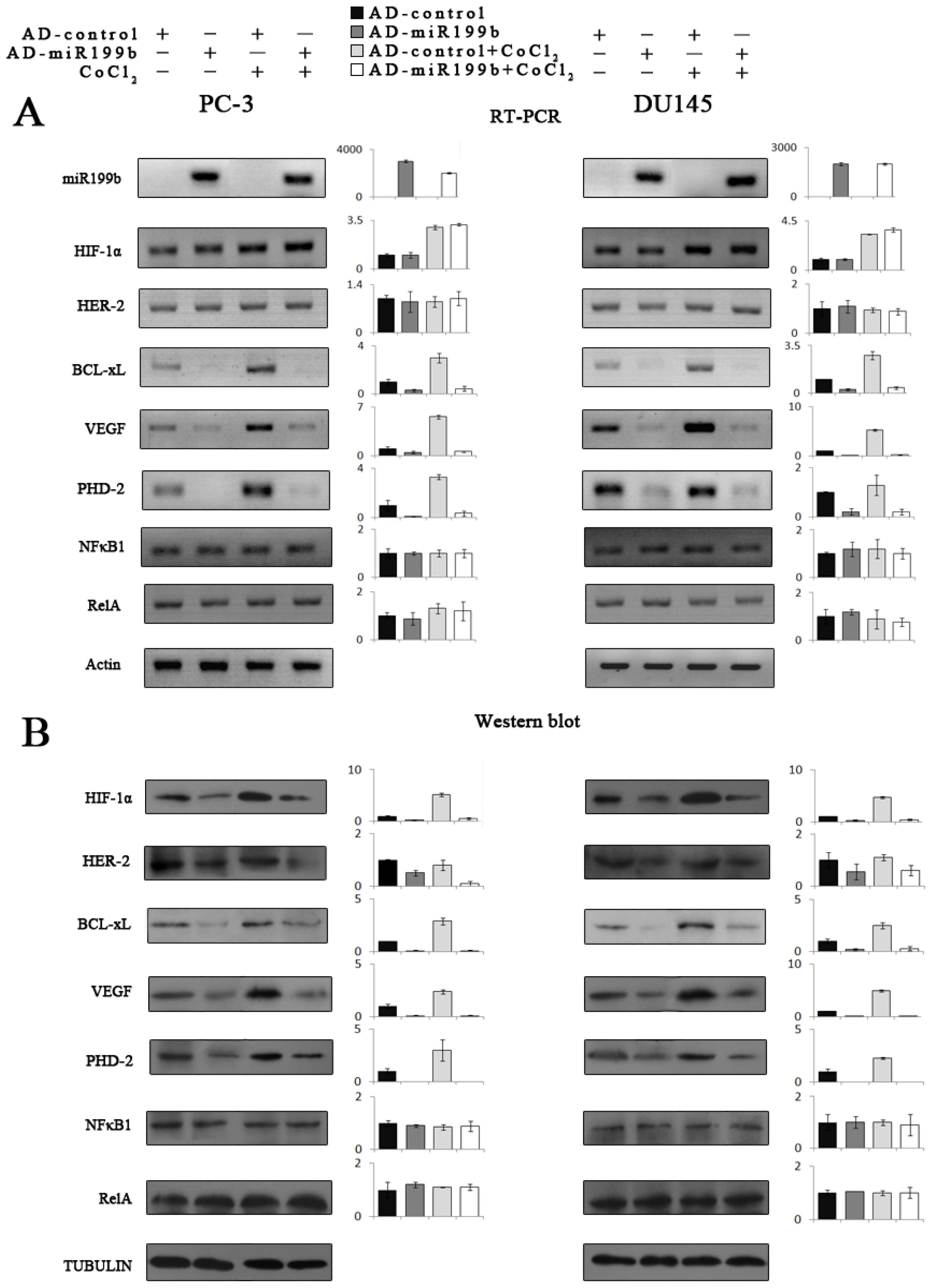
2.5. Artificial Overexpression of miR-199b in HIF-1α Knockdown Cells
3. Experimental Section
3.1. Cell lines and General Reagents
3.2. Stem-Loop Reverse Transcription and Conventional Reverse Transcription-PCR
3.3. Real-Time Quantitative PCR
3.4. Hypoxia Mimetic Treatment of PC-3 and DU145 Cells
3.5. Knockdown of HIF-1α Using Small Interfering RNA (siRNA)
3.6. Western Blot
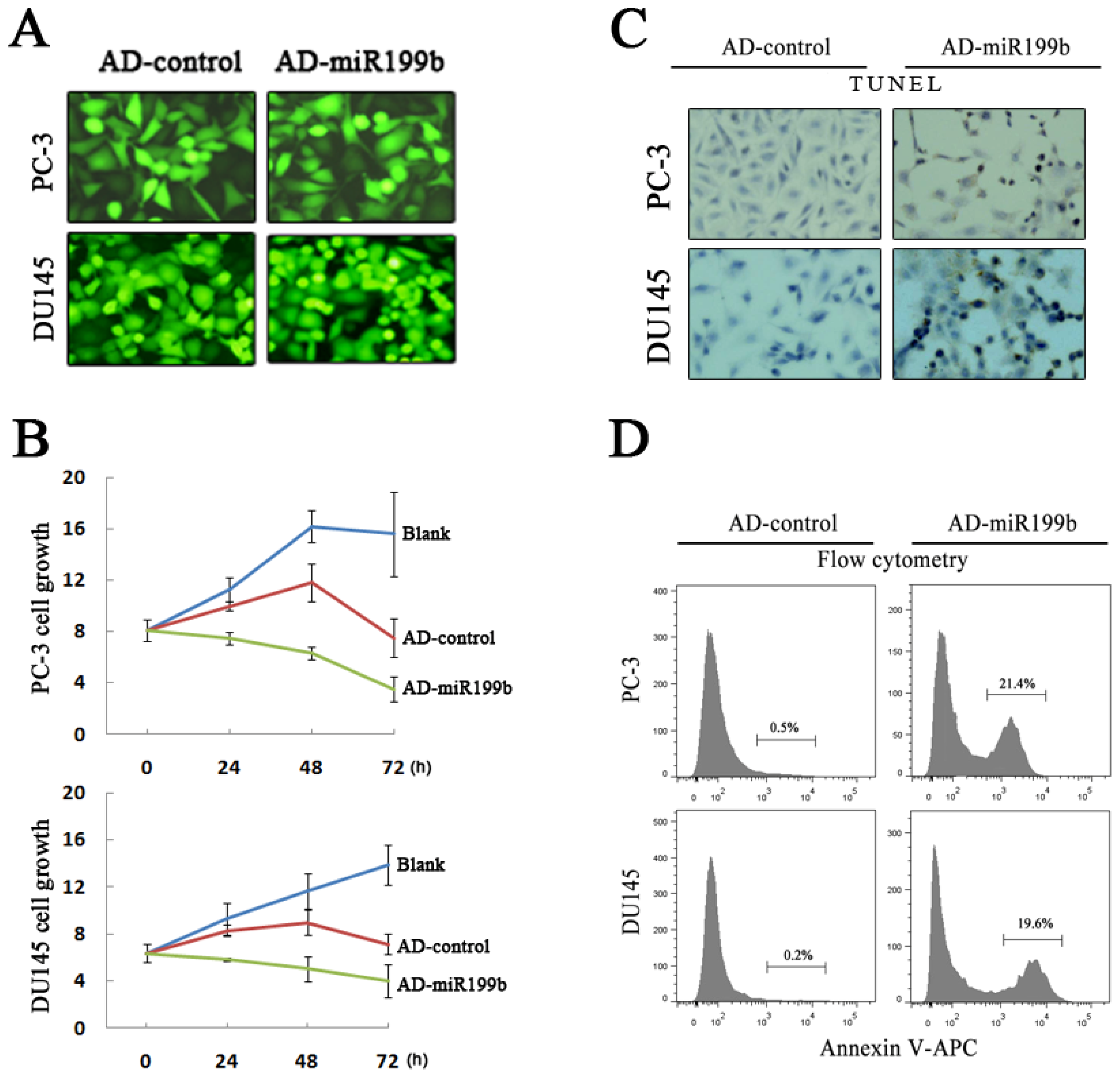
3.8. Cell Viability Assay
3.9. MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium Bromide] Assay
3.10. Terminal Deoxynucleotidyltransferase–Mediated Biotinylated dUTP Nick End-Labeling
3.11. Flow Cytometry
3.12. Luciferase Reporter Constructs and Site-Directed Mutagenesis
3.13. Dual Reporter Gene Assay
3.14. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Hudson, T.; Denis, L.J. Europa uomo: The European prostate cancer coalition. Recent Results Cancer Res 2007, 175, 267–271. [Google Scholar]
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2012. CA Cancer J. Clin 2012, 62, 10–29. [Google Scholar]
- Nelson, W.G.; de Marzo, A.M.; Isaacs, W.B. Prostate cancer. N. Engl. J. Med 2003, 349, 366–381. [Google Scholar]
- Harris, A.L. Hypoxia—A key regulatory factor in tumour growth. Nat. Rev. Cancer 2002, 2, 38–47. [Google Scholar]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar]
- Chen, N.; Chen, X.; Huang, R.; Zeng, H.; Gong, J.; Meng, W.; Lu, Y.; Zhao, F.; Wang, L.; Zhou, Q. BCL-xL is a target gene regulated by hypoxia-inducible factor-1{alpha}. J. Biol. Chem 2009, 284, 10004–10012. [Google Scholar]
- Lin, C.; McGough, R.; Aswad, B.; Block, J.A.; Terek, R. Hypoxia induces HIF-1alpha and VEGF expression in chondrosarcoma cells and chondrocytes. J. Orthop. Res 2004, 22, 1175–1181. [Google Scholar]
- Yu, X.; Deng, L.; Wang, D.; Li, N.; Chen, X.; Cheng, X.; Yuan, J.; Gao, X.; Liao, M.; Wang, M.; et al. Mechanism of TNF-alpha autocrine effects in hypoxic cardiomyocytes: Initiated by hypoxia inducible factor 1alpha, presented by exosomes. J. Mol. Cell Cardiol 2012, 53, 848–857. [Google Scholar]
- Zhong, H.; Semenza, G.L.; Simons, J.W.; de Marzo, A.M. Up-regulation of hypoxia-inducible factor 1alpha is an early event in prostate carcinogenesis. Cancer Detect. Prev 2004, 28, 88–93. [Google Scholar]
- Movsas, B.; Chapman, J.D.; Hanlon, A.L.; Horwitz, E.M.; Greenberg, R.E.; Stobbe, C.; Hanks, G.E.; Pollack, A. Hypoxic prostate/muscle pO2 ratio predicts for biochemical failure in patients with prostate cancer: Preliminary findings. Urology 2002, 60, 634–639. [Google Scholar]
- Cha, S.T.; Chen, P.S.; Johansson, G.; Chu, C.Y.; Wang, M.Y.; Jeng, Y.M.; Yu, S.L.; Chen, J.S.; Chang, K.J.; Jee, S.H.; et al. MicroRNA-519c suppresses hypoxia-inducible factor-1alpha expression and tumor angiogenesis. Cancer Res 2010, 70, 2675–2685. [Google Scholar]
- Zhong, H.; Agani, F.; Baccala, A.A.; Laughner, E.; Rioseco-Camacho, N.; Isaacs, W.B.; Simons, J.W.; Semenza, G.L. Increased expression of hypoxia inducible factor-1alpha in rat and human prostate cancer. Cancer Res 1998, 58, 5280–5284. [Google Scholar]
- Zhong, H.; de Marzo, A.M.; Laughner, E.; Lim, M; Hilton, D.A.; Zagzag, D.; Buechler, P.; Isaacs, W.B.; Semenza, G.L.; Simons, JW. Overexpression of hypoxia-inducible factor 1alpha in common human cancers and their metastases. Cancer Res 1999, 59, 5830–5835. [Google Scholar]
- Gu, S.; Jin, L.; Zhang, F.; Sarnow, P.; Kay, M.A. Biological basis for restriction of microRNA targets to the 3′ untranslated region in mammalian mRNAs. Nat. Struct. Mol. Biol 2009, 16, 144–150. [Google Scholar]
- Ryan, B.M.; Robles, A.I.; Harris, C.C. Genetic variation in microRNA networks: The implications for cancer research. Nat. Rev. Cancer 2010, 10, 389–402. [Google Scholar]
- Suzuki, H.I.; Miyazono, K. Dynamics of microRNA biogenesis: Crosstalk between p53 network and microRNA processing pathway. J. Mol. Med 2010, 88, 1085–1094. [Google Scholar]
- Li, X.; Pan, J.H.; Song, B.; Xiong, E.Q.; Chen, Z.W.; Zhou, Z.S.; Su, Y.P. Suppression of CX43 expression by miR-20a in the progression of human prostate cancer. Cancer Biol. Ther 2012, 13, 890–898. [Google Scholar]
- Maugeri-Sacca, M.; Coppola, V.; Bonci, D.; de Maria, R. MicroRNAs and prostate cancer: From preclinical research to translational oncology. Cancer J 2012, 18, 253–261. [Google Scholar]
- Singh, S.; Chitkara, D.; Mehrazin, R.; Behrman, S.W.; Wake, R.W.; Mahato, R.I. Chemoresistance in prostate cancer cells is regulated by miRNAs and Hedgehog pathway. PLoS One 2012, 7, e40021. [Google Scholar]
- Lee, Y.S.; Dutta, A. MicroRNAs: Small but potent oncogenes or tumor suppressors. Curr. Opin. Investig. Drugs 2006, 7, 560–564. [Google Scholar]
- Visone, R. Croce CM MiRNAs and cancer. Am. J. Pathol 2009, 174, 1131–1138. [Google Scholar]
- DeVere White, R.W.; Vinall, R.L.; Tepper, C.G.; Shi, X.B. MicroRNAs and their potential for translation in prostate cancer. Urol. Oncol 2009, 27, 307–311. [Google Scholar]
- Cho, W.C. MicroRNAs in cancer—From research to therapy. Biochim. Biophys. Acta A 2010, 1805, 209–217. [Google Scholar]
- Cho, W.C. MicroRNAs: Potential biomarkers for cancer diagnosis, prognosis and targets for therapy. Int. J. Biochem. Cell Biol. B 2010, 42, 1273–1281. [Google Scholar]
- Chen, F.; Yu, Z.; Baoyu, G. MiR-199b-5p targets HER2 in breast cancer cells. J. Cell Biochem. 2013. [Google Scholar] [CrossRef]
- Gao, P.; Wong, C.C.; Tung, E.K.; Lee, J.M.; Wong, C.M.; Ng, I.O. Deregulation of microRNA expression occurs early and accumulates in early stages of HBV-associated multistep hepatocarcinogenesis. J. Hepatol 2011, 54, 1177–1184. [Google Scholar]
- Pignot, G.; Cizeron-Clairac, G.; Vacher, S.; Susini, A.; Tozlu, S.; Vieillefond, A.; Zerbib, M.; Lidereau, R.; Debre, B.; Amsellem-Ouazana, D.; et al. Microrna expression profile in a large series of bladder tumors: Identification of a 3-miRNA signature associated with aggressiveness of muscle-invasive bladder cancer. Int. J. Cancer 2013, 132, 2479–2491. [Google Scholar]
- Torres, A.; Torres, K.; Pesci, A.; Ceccaroni, M.; Paszkowski, T.; Cassandrini, P.; Zamboni, G.; Maciejewski, R. Deregulation of miR-100, miR-99a and miR-199b in tissues and plasma coexists with increased expression of mTOR kinase in endometrioid endometrial carcinoma. BMC Cancer 2012, 12. [Google Scholar] [CrossRef]
- Andolfo, I.; Liguori, L.; de Antonellis, P.; Cusanelli, E.; Marinaro, F.; Pistollato, F.; Garzia, L.; de Vita, G.; Petrosino, G.; Accordi, B.; et al. The micro-RNA 199b-5p regulatory circuit involves Hes1, CD15, and epigenetic modifications in medulloblastoma. Neuro-Oncology 2012, 14, 596–612. [Google Scholar]
- Rossing, M.; Borup, R.; Henao, R.; Winther, O.; Vikesaa, J.; Niazi, O.; Godballe, C.; Krogdahl, A.; Glud, M.; Hjort-Sørensen, C.; et al. Down-regulation of microRNAs controlling tumourigenic factors in follicular thyroid carcinoma. J. Mol. Endocrinol. 2012, 48, 11–23. [Google Scholar]
- Wu, X.; Weng, L.; Li, X.; Guo, C.; Pal, S.K.; Jin, J.M.; Li, Y.; Nelson, R.A.; Mu, B.; Onami, S.H.; et al. Identification of a 4-microRNA signature for clear cell renal cell carcinoma metastasis and prognosis. PLoS One 2012, 7, e35661. [Google Scholar]
- Zagzag, D.; Zhong, H.; Scalzitti, J.M.; Laughner, E.; Simons, J.W.; Semenza, G.L. Expression of hypoxia-inducible factor 1alpha in brain tumors: Association with angiogenesis, invasion, and progression. Cancer 2000, 88, 2606–2618. [Google Scholar]
- Giatromanolaki, A.; Koukourakis, M.I.; Sivridis, E.; Turley, H.; Talks, K.; Pezzella, F.; Gatter, K.C.; Harris, A.L. Relation of hypoxia inducible factor 1 alpha and 2 alpha in operable non-small cell lung cancer to angiogenic/molecular profile of tumours and survival. Br. J. Cancer 2001, 85, 881–890. [Google Scholar]
- Saponaro, C.; Malfettone, A.; Ranieri, G.; Danza, K.; Simone, G.; Paradiso, A.; Mangia, A. VEGF, HIF-1alpha expression and MVD as an angiogenic network in familial breast cancer. PLoS One 2013, 8, e53070. [Google Scholar]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar]
- Chao, A.; Tsai, C.L.; Wei, P.C.; Hsueh, S.; Chao, A.S.; Wang, C.J.; Tsai, C.N.; Lee, Y.S.; Wang, T.H.; Lai, C.H. Decreased expression of microRNA-199b increases protein levels of SET (protein phosphatase 2A inhibitor) in human choriocarcinoma. Cancer Lett 2010, 291, 99–107. [Google Scholar]
- Favreau, A.J.; Cross, E.L.; Sathyanarayana, P. miR-199b-5p directly targets PODXL and DDR1 and decreased levels of miR-199b-5p correlate with elevated expressions of PODXL and DDR1 in acute myeloid leukemia. Am. J. Hematol 2012, 87, 442–446. [Google Scholar]
- Garzia, L.; Andolfo, I.; Cusanelli, E.; Marino, N.; Petrosino, G.; de Martino, D.; Esposito, V.; Galeone, A.; Navas, L.; Esposito, S.; et al. MicroRNA-199b-5p impairs cancer stem cells through negative regulation of HES1 in medulloblastoma. PLoS One 2009, 4, e4998. [Google Scholar]
- De Antonellis, P.; Liguori, L.; Falanga, A.; Carotenuto, M.; Ferrucci, V.; Andolfo, I.; Marinaro, F.; Scognamiglio, I.; Virgilio, A.; de Rosa, G.; et al. MicroRNA 199b-5p delivery through stable nucleic acid lipid particles (SNALPs) in tumorigenic cell lines. Naunyn Schmiedebergs Arch. Pharmacol 2013, 386, 287–302. [Google Scholar] [Green Version]
- van Uden, P.; Kenneth, N.S.; Rocha, S. Regulation of hypoxia-inducible factor-1alpha by NF-kappaB. Biochem. J 2008, 412, 477–484. [Google Scholar]
- Maxwell, P.J.; Gallagher, R.; Seaton, A.; Wilson, C.; Scullin, P.; Pettigrew, J.; Stratford, I.J.; Williams, K.J.; Johnston, P.G.; Waugh, D.J. HIF-1 and NF-kappaB-mediated upregulation of CXCR1 and CXCR2 expression promotes cell survival in hypoxic prostate cancer cells. Oncogene 2007, 26, 7333–7345. [Google Scholar]
- Piret, J.P.; Minet, E.; Cosse, J.-P.; Ninane, N.; Debacq, C.; Raes, M.; Michiels, C. Hypoxia-inducible factor-1-dependent overexpression of myeloid cell factor-1 protects hypoxic cells against tert-butyl hydroperoxide-induced apoptosis. J. Biol. Chem 2005, 280, 9336–9344. [Google Scholar]
- Hanze, J.; Eul, B.G.; Savai, R.; Krick, S.; Goyal, P.; Grimminger, F.; Seeger, W.; Rose, F. RNA interference for HIF-1alpha inhibits its downstream signalling and affects cellular proliferation. Biochem. Biophys. Res. Commun 2003, 312, 571–577. [Google Scholar]
- Gillespie, D.L.; Whang, K.; Ragel, B.T.; Flynn, J.R.; Kelly, D.A.; Jensen, R.L. Silencing of hypoxia inducible factor-1alpha by RNA interference attenuates human glioma cell growth in vivo. Clin. Cancer Res 2007, 13, 2441–2448. [Google Scholar]
- Peng, X.H.; Karna, P.; Cao, Z.; Jiang, B.H.; Zhou, M.; Yang, L. Cross-talk between epidermal growth factor receptor and hypoxia-inducible factor-1alpha signal pathways increases resistance to apoptosis by up-regulating survivin gene expression. J. Biol. Chem 2006, 281, 25903–25914. [Google Scholar]
- Metzen, E.; Stiehl, D.P.; Doege, K.; Marxsen, J.H.; Hellwig-Bürgel, T.; Jelkmann, W. Regulation of the prolyl hydroxylase domain protein 2 (phd2/egln-1) gene: Identification of a functional hypoxia-responsive element. Biochem. J 2005, 387, 711–717. [Google Scholar]






© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shang, W.; Chen, X.; Nie, L.; Xu, M.; Chen, N.; Zeng, H.; Zhou, Q. MiR199b Suppresses Expression of Hypoxia-Inducible Factor 1α (HIF-1α) in Prostate Cancer Cells. Int. J. Mol. Sci. 2013, 14, 8422-8436. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14048422
Shang W, Chen X, Nie L, Xu M, Chen N, Zeng H, Zhou Q. MiR199b Suppresses Expression of Hypoxia-Inducible Factor 1α (HIF-1α) in Prostate Cancer Cells. International Journal of Molecular Sciences. 2013; 14(4):8422-8436. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14048422
Chicago/Turabian StyleShang, Weiwei, Xueqin Chen, Ling Nie, Miao Xu, Ni Chen, Hao Zeng, and Qiao Zhou. 2013. "MiR199b Suppresses Expression of Hypoxia-Inducible Factor 1α (HIF-1α) in Prostate Cancer Cells" International Journal of Molecular Sciences 14, no. 4: 8422-8436. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14048422