Protective Effects of a New Phloretin Derivative against UVB-Induced Damage in Skin Cell Model and Human Volunteers


Abstract
:1. Introduction

2. Results and Discussion
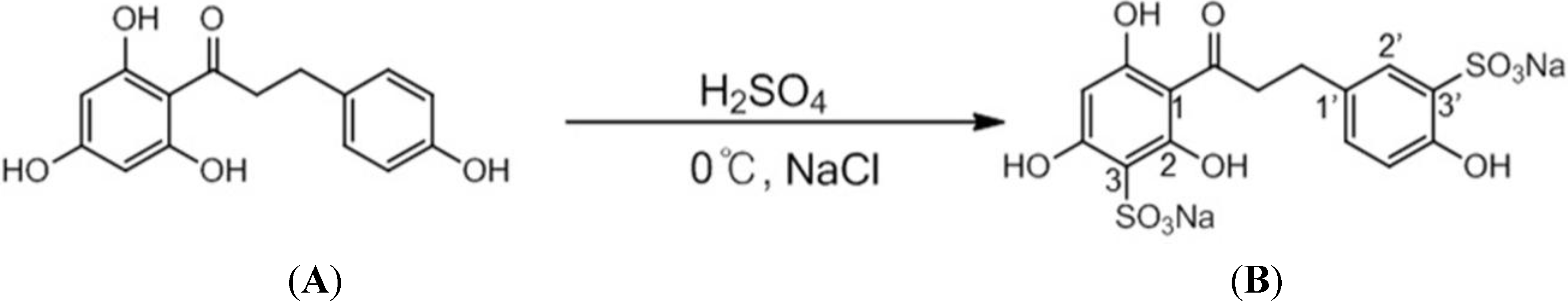
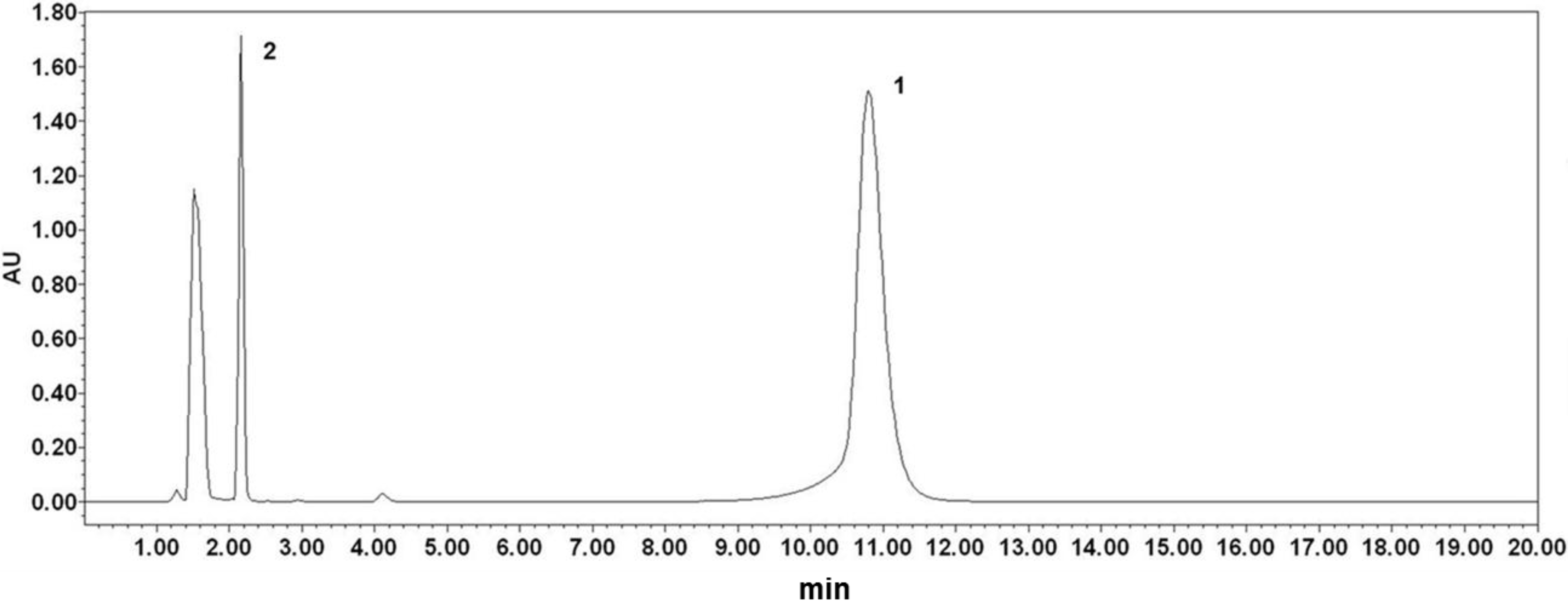
2.1. Analysis of Phloretin and Phloretin 3',3-Disulfonate

2.2. Solubilities of Phloretin and Phloretin 3',3-Disulfonate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Test Material | Solubility in Water (μg/mL) a | Relative Solubility |
|---|---|---|---|
| 1 | Phloretin | 7 ± 1 | 1 |
| 2 | Phloretin 3',3-disulfonate | 187,292 ± 940 | 26,756 |
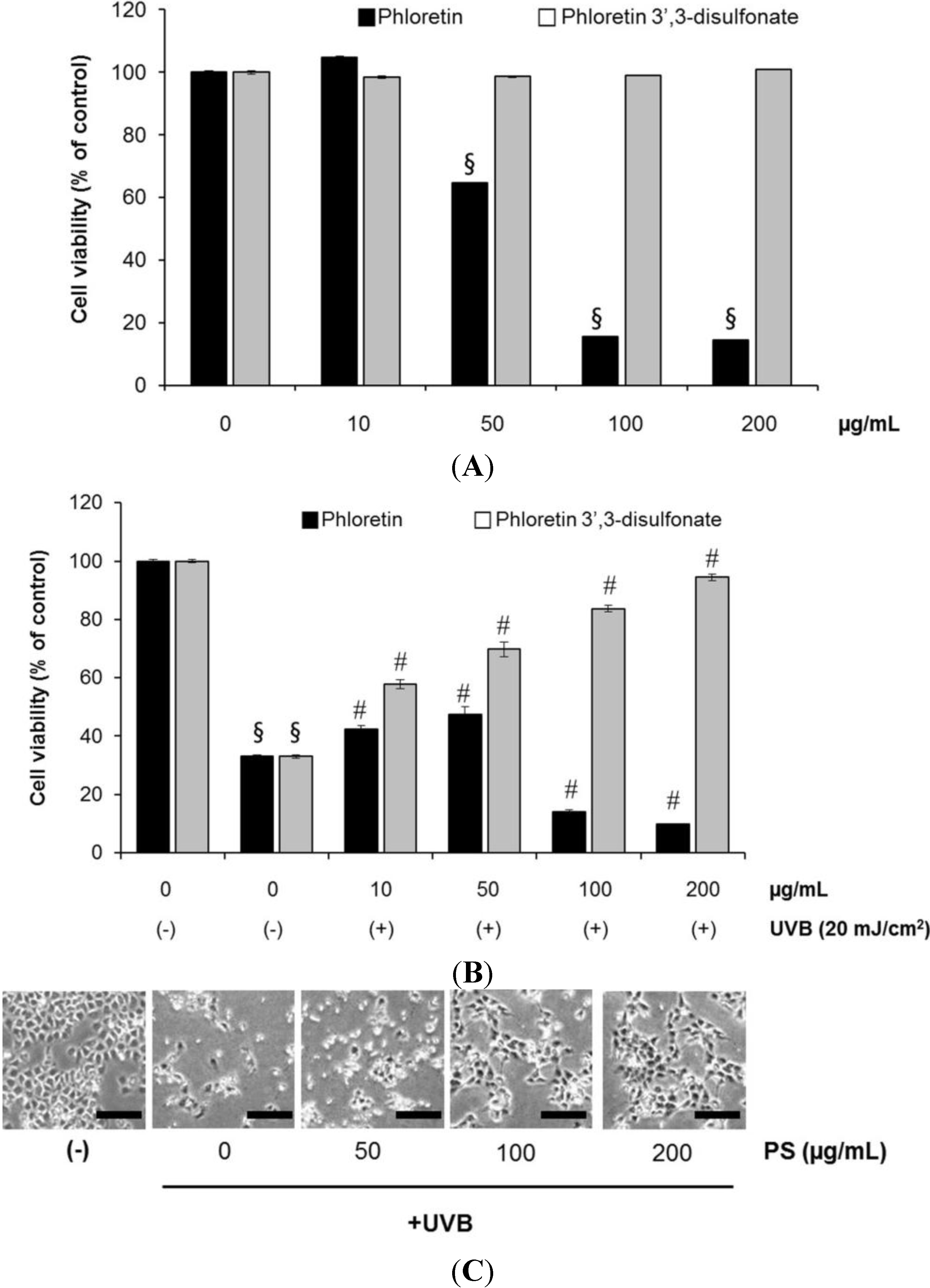
2.3. Phloretin 3',3-Disulfonate Inhibits UV-Induced Damage in HaCaT Cells


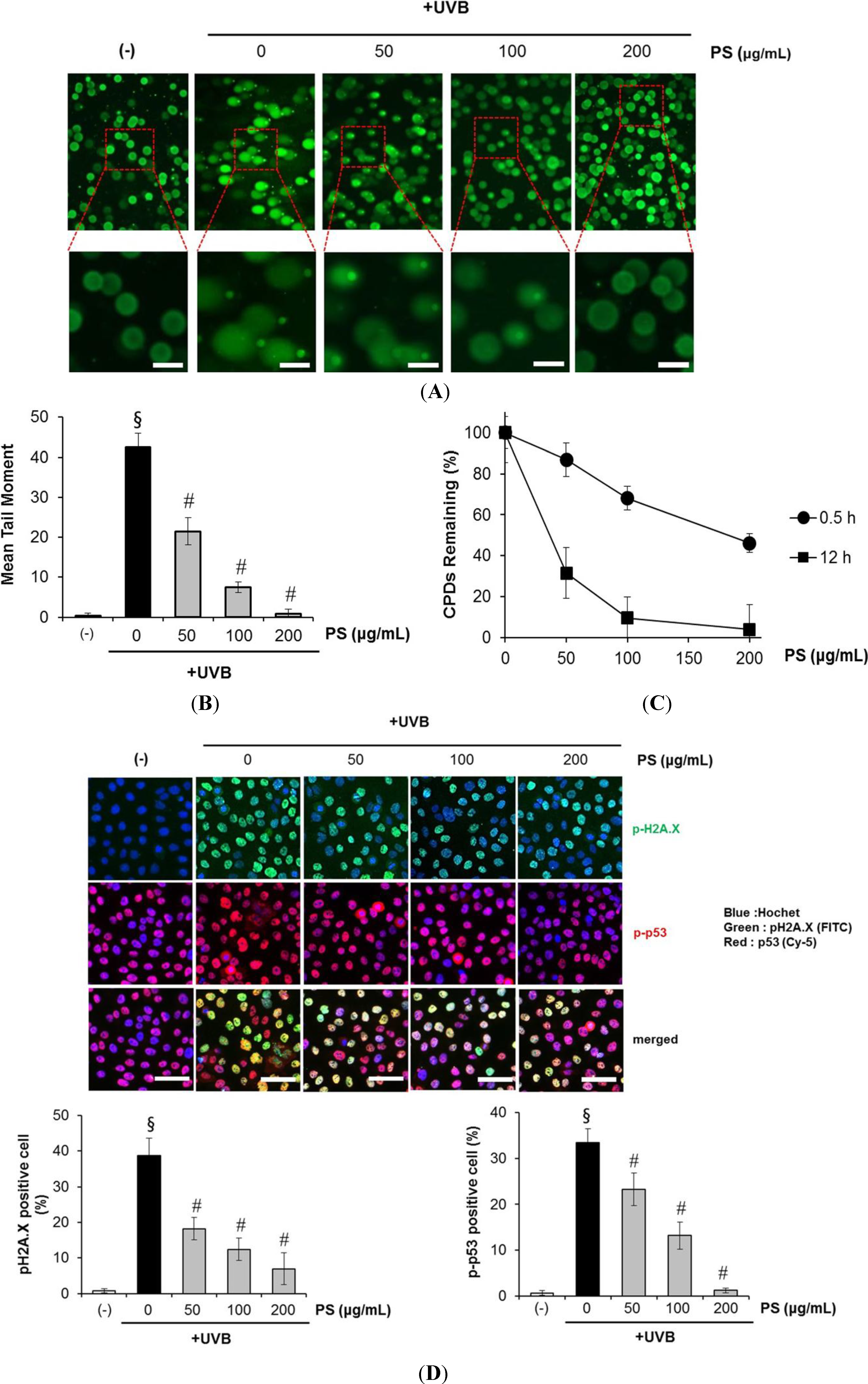
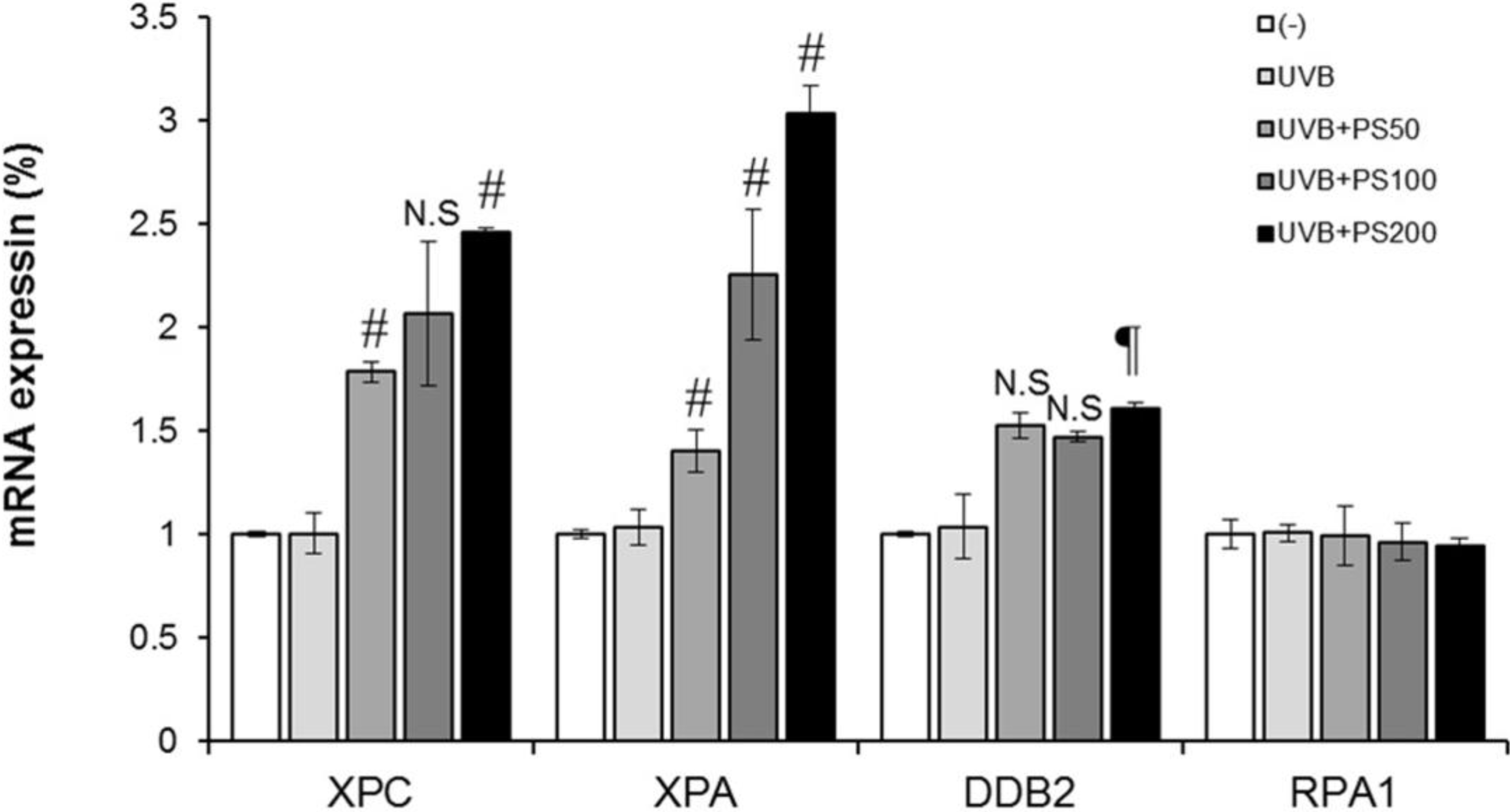
2.4. Phloretin 3',3-Disulfonate Enhances Expression of the NER Gene in UVB-Irradiated Cells

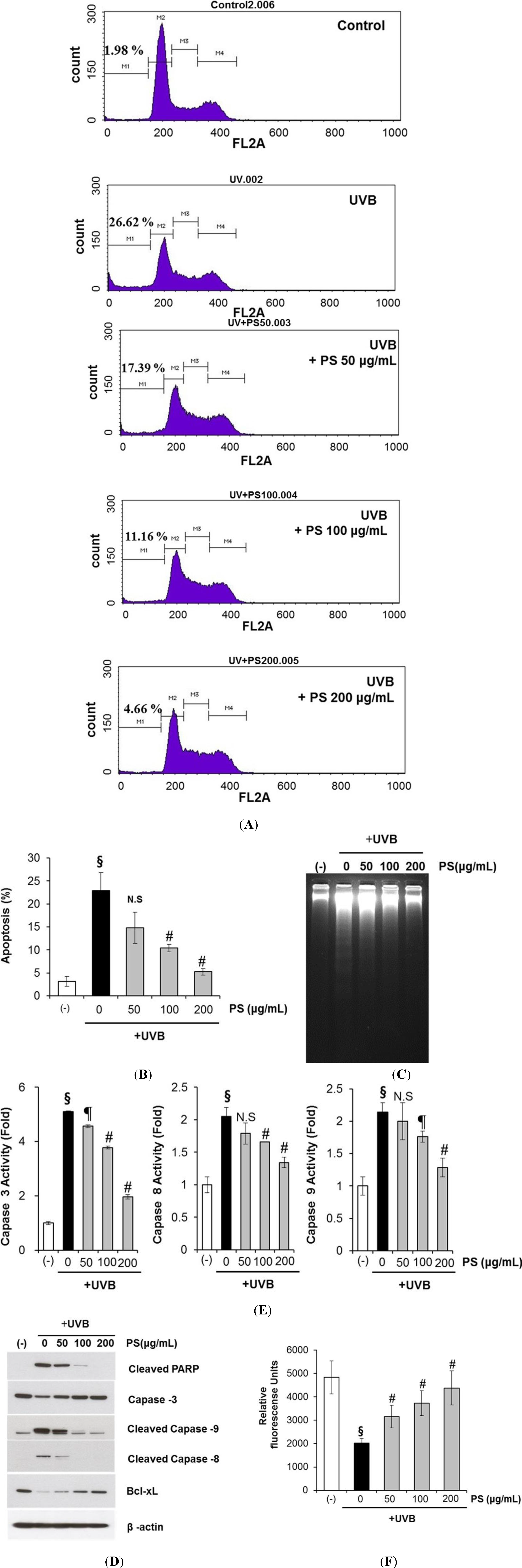
2.5. Phloretin 3',3-Disulfonate Protects HaCaT Cells from UVB-Mediated Apoptosis

2.6. Phloretin 3',3-Disulfonate Prevent UVB-Induced Depletion of GSH in HaCaT Cells
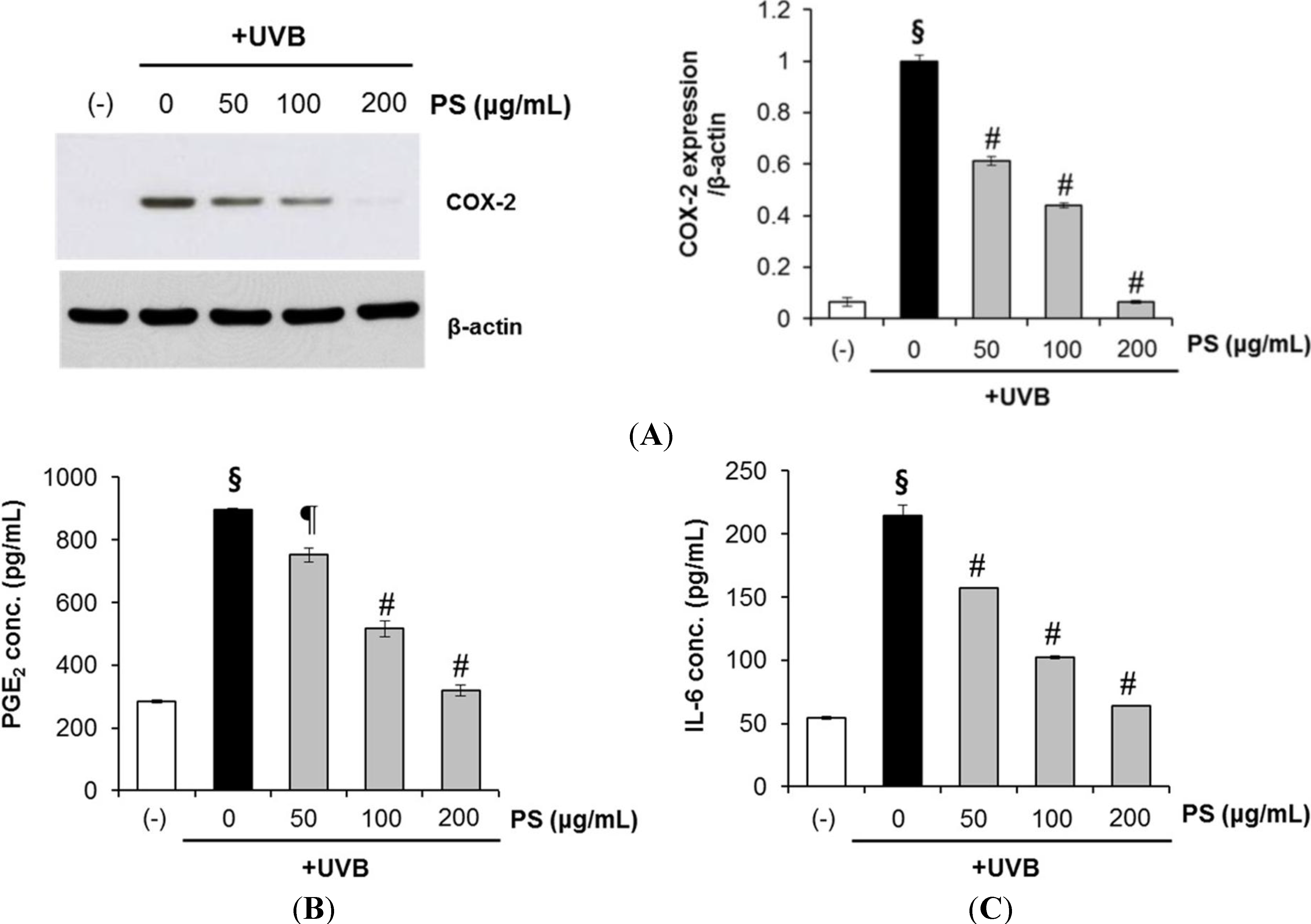
2.7. Phloretin 3',3-Disulfonate Inhibits UVB-Induced Inflammation in Vitro and in Vivo


2.8. Human Skin Primary Irritation Test of Phloretin 3',3-Disulfonate
| No. | Test Material | 48 h | 72 h | Reaction Grade b | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ± | 1+ | 2+ | 3+ | 4+ | ± | 1+ | 2+ | 3+ | 4+ | 48 h | 72 h | Mean | ||
| Squalene | - a | - | - | - | - | - | - | - | - | - | 0 | 0 | 0 | |
| 2 | Phloretin 3',3-disulfonate (0.1%) | - | - | - | - | - | - | - | - | - | - | 0 | 0 | 0 |
3. Experimental Section
3.1. Chemicals and Antibodies
3.2. Modification of Phloretin
3.3. Analysis of Phloretin and Phloretin 3',3-Disulfonate
3.4. Water Solubility Measurement
3.5. Cell Culture
3.6. UV Irradiation and Treatment
3.7. Cell Viability Assay
3.8. Analysis of DNA Damage by the Comet Assay
3.9. Cyclobutane Pyrimidine Dimer (CPD) Quantification
3.10. Immunofluorescence
3.11. FACS Analysis
3.12. DNA Fragmentation Assay
3.13. Western Blot Analysis
3.14. Enzymatic Caspase Activity Assay
3.15. Real Time PCR
3.16. Intracellullar GSH Level
3.17. Inflammatory Cytokine Assay
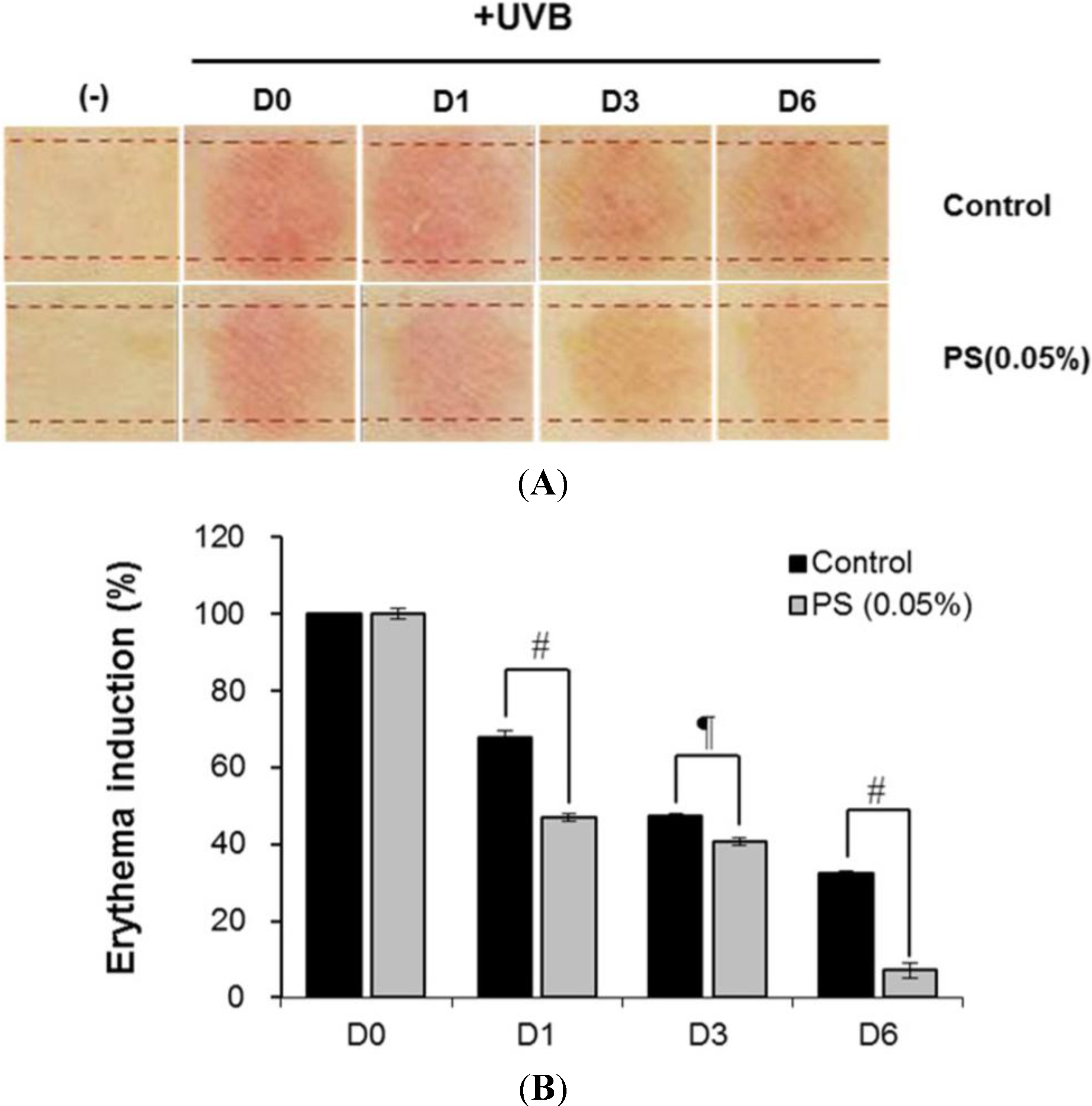
3.18. Clinical Study of Skin Recovery Effect of Phloretin 3',3-Disulfonate on UV-Induced Damage Skin
3.19. Human Skin Primary Irritation Test
3.20. Statistical Analysis
4. Conclusions
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Amaro-Ortiz, A.; Yan, B.; D’Orazio, J.A. Ultraviolet radiation, aging and the skin: Prevention of damage by topical cAMP Manipulation. Molecules 2014, 19, 6202–6219. [Google Scholar] [CrossRef] [PubMed]
- Afaq, F.; Adhami, V.M.; Mukhtar, H. Photochemoprevention of ultraviolet B signaling and photocacinogenesis. Mutat. Res. 2005, 571, 153–173. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.W.; Park, K.; Kweon, G.R.; Jang, B.C.; Baek, W.K.; Suh, M.H.; Kim, C.W.; Lee, K.S.; Suh, S.I. Curcumin inhibits the expression of COX-2 in UVB-irradiated human keratinocyte (HaCaT) by inhibiting activation of AP-1: p38 MAP kinase and JNK as potential upstream targets. Exp. Mol. Med. 2005, 37, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.D.; Lin, J.C.; Yang, S.M.; Tseng, M.J.; Hsu, J.D.; Lee, Y.J.; Cherng, J.M. Curcumin protects against UVB-induced skin cancers in SKH-1 hairless mouse: Analysis of early molecular markers in carcinogenesis. Evid. Based Complement Altern. Med. 2012, 2012, 593952. [Google Scholar]
- Müller-Decker, K. Cyclooxygenase-dependent signaling is causally linked to non-melanoma skin carcinogenesis: Pharmacological, genetic, and clinical evidence. Cancer Metastasis Rev. 2011, 30, 343–361. [Google Scholar] [CrossRef] [PubMed]
- Yaar, M.; Gilchrest, B.A. Photoageing: Mechanism, prevention and therapy. Br. J. Dermatol. 2007, 157, 874–887. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Mukhtar, H. Green tea polyphenol (−)-epigallocatechin-3-gallate treatment to mouse skin prevents UVB-induced infiltration of leukocytes, depletion of antigen-presenting cells, and oxidative stress. J. Leukoc. Biol. 2001, 69, 719–726. [Google Scholar] [PubMed]
- Mittal, A.; Elmets, C.A.; Katiyar, S.K. Dietary feeding of proanthocyanidins from grape seeds prevents photocarcinogenesis in SKH-1 hairless mice: Relationship to decreased fat and lipid peroxidation. Carcinogenesis 2003, 24, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.J.; Lee, K.W.; Shin, B.J.; Jung, S.K.; Hwang, M.K.; Bode, A.M.; Heo, Y.S.; Lee, H.J.; Dong, Z. Caffeic acid, a phenolic phytochemical in coffee, directly inhibits Fyn kinase activity and UVB-induced COX-2 expression. Carcinogenesis 2009, 30, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Staniforth, V.; Huang, W.C.; Aravindaram, K.; Yang, N.S. Ferulic acid, a phenolic phytochemical, inhibits UVB-induced matrix metalloproteinases in mouse skin via posttranslational mechanisms. J. Nutr. Biochem. 2012, 23, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Alvarez-Suarez, J.M.; Tulipani, S.; Gonzàles-Paramàs, A.M.; Santos-Buelga, C.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Battino, M. Photoprotective potential of strawberry (Fragaria × ananassa) extract against UV-A irradiation damage on human fibroblasts. J. Agric. Food Chem. 2012, 60, 2322–2327. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Cheng, G.; Liu, Y.; Yang, H.; Hu, Y.; Huang, W. In vitro evaluation of tectoridin, tectorigenin and tectorigenin sodium sulfonate on antioxidant properties. Food Chem. Toxicol. 2012, 50, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Woznicka, E.; Kuzniar, A.; Nowak, D.; Nykiel, E.; Kopacz, M.; Gruszecka, J.; Golec, K. Comparative study on the antibacterial activity of some flavonoids and their sulfonic derivatives. Acta Pol. Pharm. Drug Res. 2013, 70, 567–571. [Google Scholar]
- Chlebda, E.; Magdalan, J.; Merwid-Lad, A.; Trocha, M.; Kopacz, M.; Kuzniar, A.; Nowak, D.; Szelag, A. Influence of water-soluble flavonoids, quercetin-5'-sulfonic acid sodium salt and morin-5'-sulfonic acid sodium salt, on antioxidant parameters in the subacute cadmium intoxication mouse model. Exp. Toxicol. Pathol. 2010, 62, 105–108. [Google Scholar] [CrossRef]
- Nakamura, Y.; Watanabe, S.; Miyake, N.; Kohno, H.; Osawa, T. Dihydrochalcones: Evaluation as novel radical scavenging antioxidants. J. Agric. Food Chem. 2003, 51, 3309–3312. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C.; Li, C.K.; Lin, A.H.; Yeh, Y.W.; Yao, H.T.; Li, C.C.; Liu, K.L.; Chen, H.W. Induction of glutathione synthesis and heme oxygenase 1 by the flavonoids butein and phloretin is mediated through the ERK/Nrf2 pathway and protects against oxidative stress. Free Radic. Biol. Med. 2011, 51, 2073–2081. [Google Scholar] [CrossRef] [PubMed]
- Barreca, D.; Bellocco, E.; Laqana, G.; Ginestra, G.; Bisiqnano, C. Biochemical and antimicrobial activity of phloretin and its glycosylated derivatives present in apple and kumquat. Food Chem. 2014, 160, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.; Chun, K.S.; Chae, I.G.; Kundu, J.K. Phloretin: An apple polyphenol with cancer chemopreventive potential. Arch. Bas. App. Med. 2014, 2, 17–23. [Google Scholar]
- Szliszka, E.; Czuba, Z.P.; Mazur, B.; Paradysz, A.; Krol, W. Chalcones and dihydrochalcones augment TRAIL-mediated apoptosis in prostate cancer cell. Molecules 2010, 15, 5336–5353. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Huang, W.C.; Liou, C.J. Evaluation of the anti-inflammatory effects of phloretin and phlorizin in lipopolysaccharide-stimulated mouse macrophages. Food Chem. 2012, 134, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.W.; Kundu, J.K.; Surh, Y.J. Phloretin inhibits phorbol ester-induced tumor promotion and expression of cyclooxygenase-2 in mouse skin: Extracellular signal-regulated kinase and nuclear factor-kappaB as potential targets. J. Med. Food 2012, 15, 253–257. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, S.H.; Clark, A.C. Death by caspase dimerization. Adv. Exp. Med. Biol. 2012, 747, 55–73. [Google Scholar] [PubMed]
- Fonseca, Y.M.; Catini, C.D.; Vicentini, F.T.; Nomizo, A.; Gerlach, R.F.; Fonseca, M.J. Protective effect of Calendula officinalis extract against UVB-induced oxidative stress in skin: Evaluation of reduced glutathione levels and matrix metalloproteinase secretion. J. Ethnopharmacol. 2010, 127, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Mizu, H.; Yamauchi, H. Photoinflammatory responses to UV-irradiated ketoprofen mediated by the induction of ROS generation, enhancement of cyclooxygenase-2 expression, and regulation of multiple signaling pathways. Free Radic. Biol. Med. 2010, 48, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Tripp, C.S.; Blomme, E.A.; Chinn, K.S.; Hardy, M.M.; LaCelle, P.; Pentland, A.P. Epidermal COX-2 induction following ultraviolet irradiation: Suggested mechanism for the role of COX-2 inhibition in photoprotection. J. Investig. Dermatol. 2003, 121, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Burford, C.; Tu, J.H.; Mercurio, M.; Carey, D.; Han, R.; Gordon, G.; Niwas, S.; Bell, W.; Elmets, C.A.; Grizzle, W.; et al. Selective cyclooxygenase-2 inhibition produces heterogeneous erythema response to ultraviolet irradiation. J. Investig. Dermatol. 2005, 125, 1317–1320. [Google Scholar] [CrossRef]
- Li, D.; Park, S.H.; Shim, J.H.; Lee, H.S.; Tang, S.Y.; Cheon, S.P.; Park, K.H. In vitro enzymatic modification of puerarin to puerarin glycosides by maltogenic amylase. Cabohydr. Res. 2004, 339, 2789–2797. [Google Scholar] [CrossRef]
- Frosch, P.J.; Kligman, A.M. The soap chamber test. A new method for assessing the irritancy of soaps. J. Am. Acad. Dermatol. 1979, 1, 35–41. [Google Scholar] [CrossRef] [PubMed]
- CTFA Safety Testing Guideline; Cosmetic, Toiletry and Fragrance Association: Washington, DC, USA, 1981; p. 20005.
- Statview 5. In A Statistics Application Released for Apple Macintosh Computers; Abacus Concepts: Piscataway, NJ, USA, 2009.
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.; Kum, H.; Ryu, D.; Kim, M.; Jung, E.; Park, D. Protective Effects of a New Phloretin Derivative against UVB-Induced Damage in Skin Cell Model and Human Volunteers. Int. J. Mol. Sci. 2014, 15, 18919-18940. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151018919
Shin S, Kum H, Ryu D, Kim M, Jung E, Park D. Protective Effects of a New Phloretin Derivative against UVB-Induced Damage in Skin Cell Model and Human Volunteers. International Journal of Molecular Sciences. 2014; 15(10):18919-18940. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151018919
Chicago/Turabian StyleShin, Seoungwoo, Hyunwoo Kum, Dehun Ryu, Minkyung Kim, Eunsun Jung, and Deokhoon Park. 2014. "Protective Effects of a New Phloretin Derivative against UVB-Induced Damage in Skin Cell Model and Human Volunteers" International Journal of Molecular Sciences 15, no. 10: 18919-18940. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151018919