Identification and Molecular Characterization of a Chitin-Binding Protein from the Beet Webworm, Loxostege sticticalis L.

Abstract
:1. Introduction
2. Results and Discussion
2.1. Cloning of the CBP cDNA of Loxostege sticticalis

2.2. Characterization of LstiCBP Chitin-Binding Domains


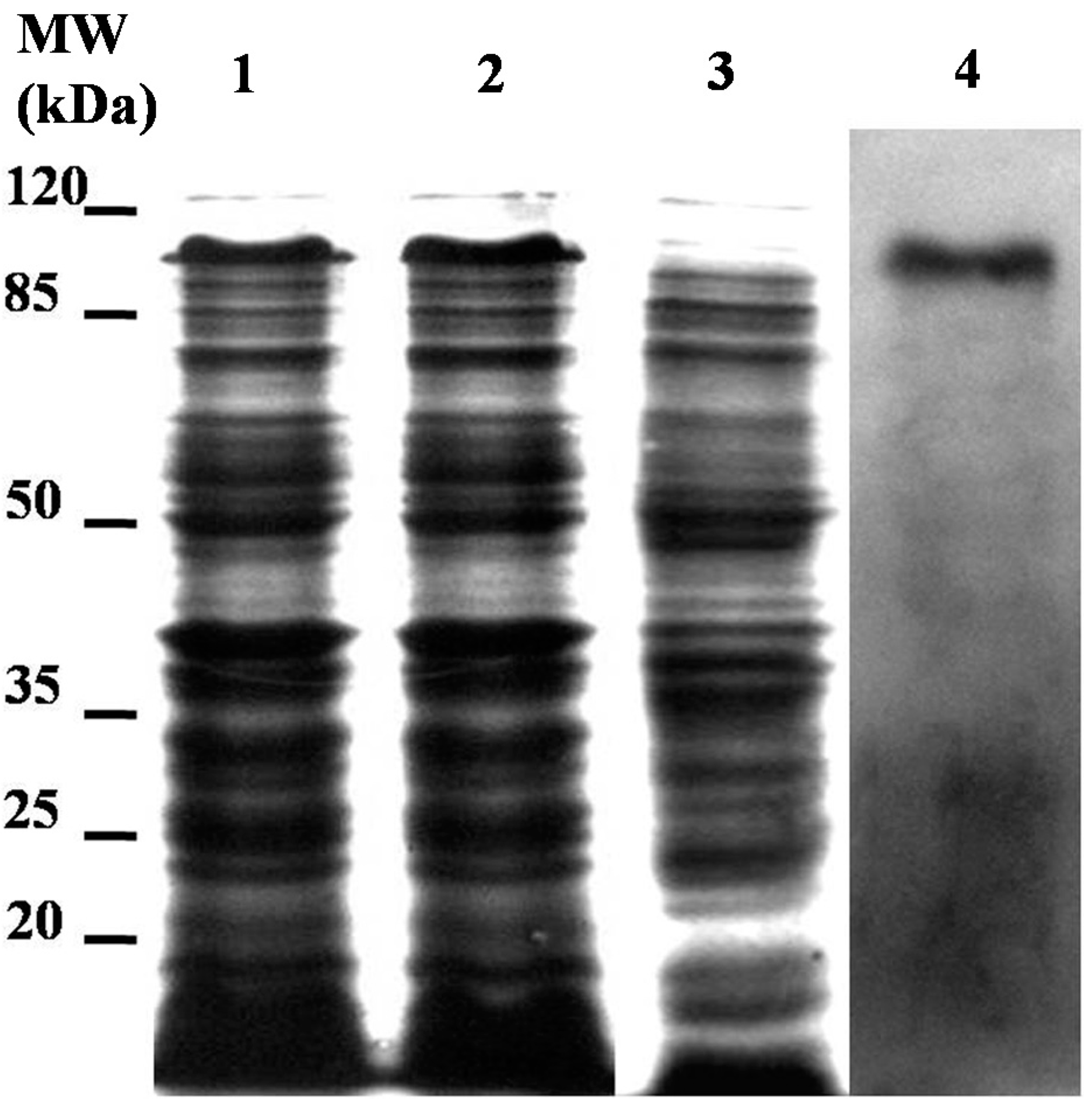
2.3. Expression of Recombinant LstiCBP in E. Coliand Chitin-Binding Analysis


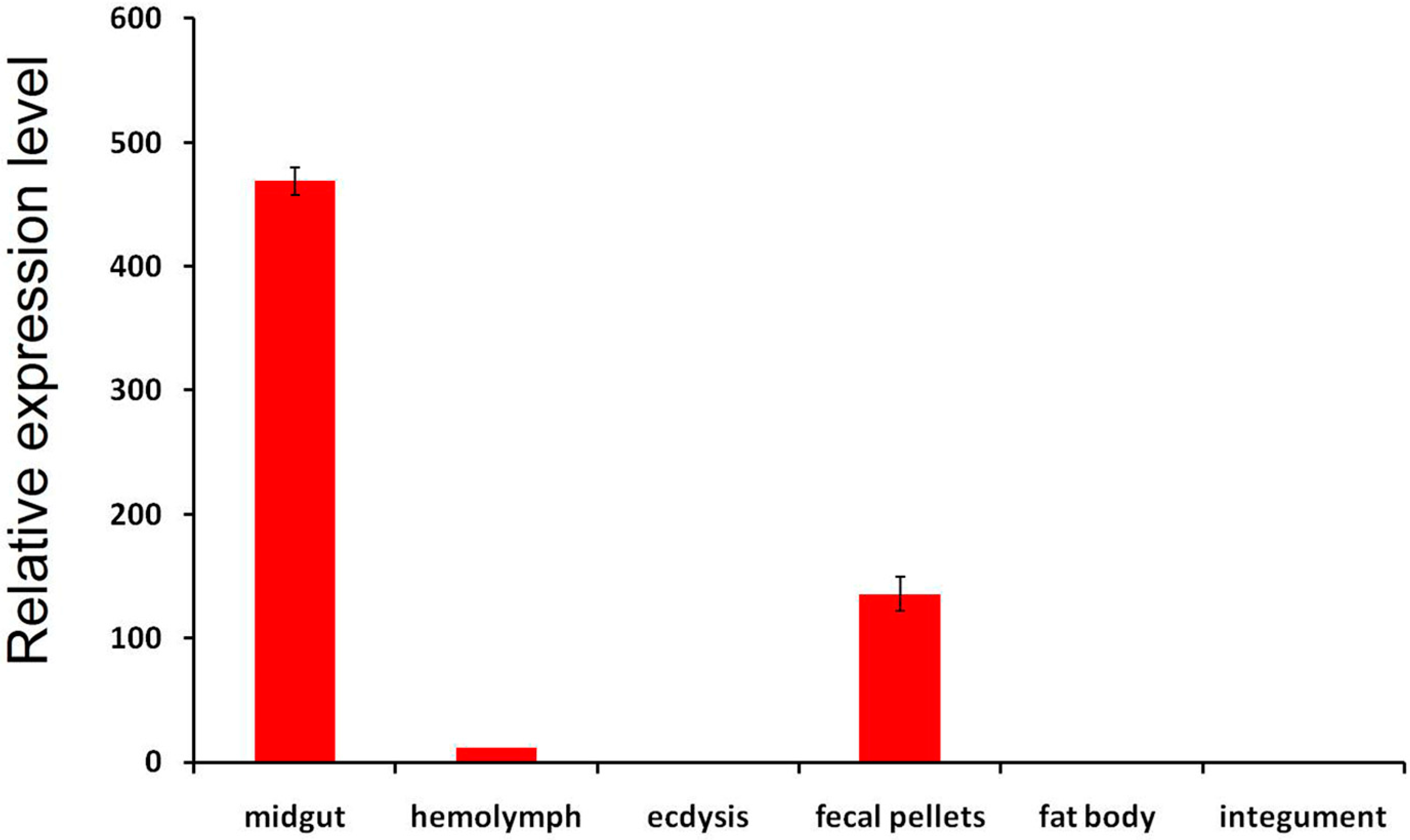
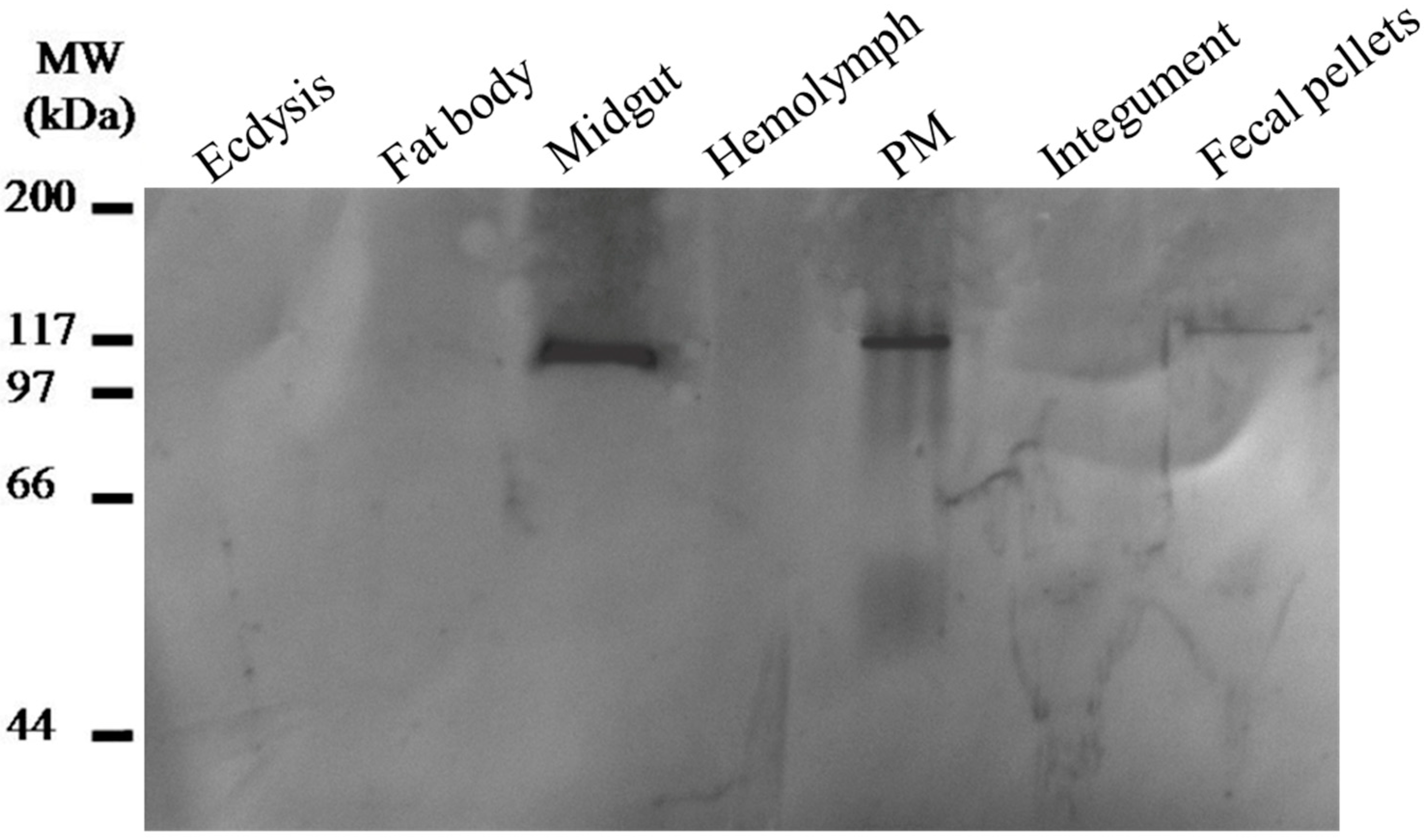
2.4. Expression Profiles and Localization of LstiCBP


3. Experimental Section
3.1. Insect Larvae
3.2. Cloning and Sequencing of LstiCBP

{kind=link}
{kind=link}
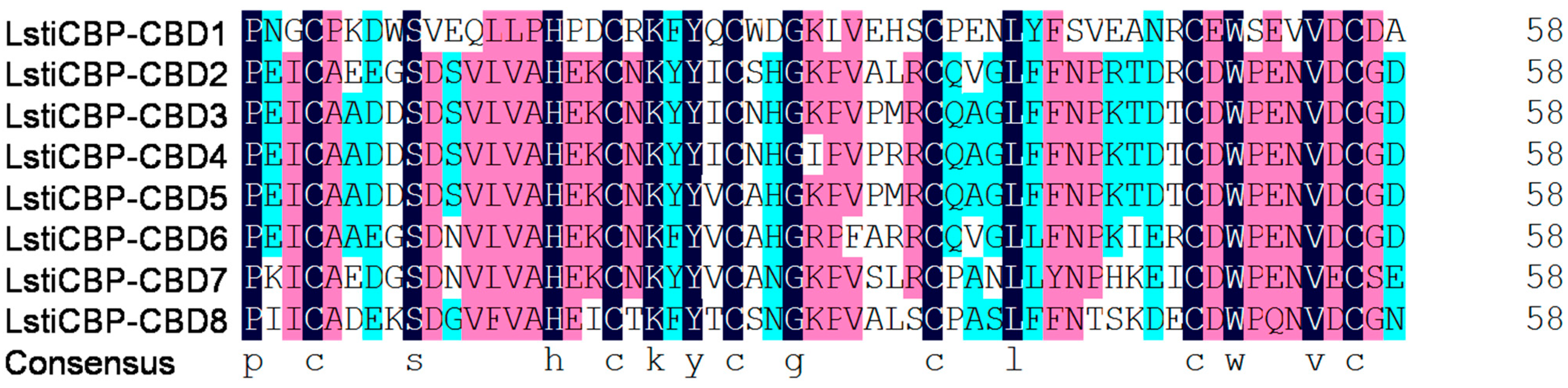{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5'-3') | Position |
|---|---|---|
| CBP-R1 | CATCGGGACAGGCTTTCCGTGGTT | 687–710 |
| CBP-R2 | TCCTCGGTACTGGTATTCCGTGATTACAAA | 991–1020 |
| CBP-R3 | CGGACCACTCACATCGGTTTGCTTC | 198–222 |
| CBP-R4 | CATCGGGACAGGCTTTCCGTGGTT | 687–710 |
| 5'RACE Outer Primer: | CATGGCTACATGCTGACAGCCTA | - |
| 5'RACE Inner Primer: | CGCGGATCCACAGCCTACTGATGATCAGTCGATG | - |
3.3. Expression Pattern of LstiCBP
| Gene Name | Primer Name | Sequence (5'-3') |
|---|---|---|
| CBP | Forward | GCCCATGAAATATGTACCAAGTTCTAT |
| Reverse | CAGTCGCACTCGTCTTTGGA | |
| Probe | FAM-ACGGCAAGCCTGTCGCCCTCA-TAMRA | |
| 18S rRNA | Forward | CAGGCTAGAGTCTCGTTCGTTACC |
| Reverse | CAACACGGGAAATCTCACCAG | |
| Probe | FAM-CAAATCGCTCCACCAACTAAGAACGGC-TAMRA |
3.4. Expression of Recombinant LstiCBP in E. coli and Chitin-Binding Analysis
 AAATCTGGGGATAGTGGTATAAAC-3') and CBP-MRP(5'-G
AAATCTGGGGATAGTGGTATAAAC-3') and CBP-MRP(5'-G  AAACCCATCGCATAAAAGTG-3') (the boxed sequences indicate the BamH I and EcoR I restriction sites, respectively). The PCR product was inserted into the BamH I and EcoR I sites of pET30, and transformed into the E. coli BL21 strain. After 3 h preincubation, the recombinant LstiCBP was induced by adding isopropyl-beta-d-thiogalactopyranoside (IPTG) at a final concentration of 2.0 mM for 4 h. The cells (1 L) were harvested by centrifugation, and the pellets were homogenized in phosphate-buffered saline (PBS, 0.04 M, pH 7.0). After centrifugation at 12,000× g for 20 min at 4 °C, the supernatants were dried and stored at −70 °C until use. Before SDS-PAGE, the cells were thawed and disrupted in PBS by sonication (5 s, 5 passes, 4 °C). The cell lysates from the cells before or after induction with IPTG were mixed with SDS-PAGE sample buffer, boiled for 10 min, centrifugation at 12,000× g for 10 min and supernatants were loaded onto an 8% SDS-PAGE gel. Western blot was performed following the description of Wei et al. [36]. After SDS-PAGE, the proteins were blotted onto a PVDF membrane (Hybond-P, Amersham, Zhengzhou, China), and the membrane was incubated with antibodies to 6 × His for 2 h at 37 °C. After washing in PBST (PBS-Tween, Sigma-aldrich, Shanghai, China), the membrane was incubated with secondary antibodies (HRP-conjugated goat anti-rabbit IgG, dilution 1/2000) for 2 h at 37 °C and then washed thoroughly with PBST. Antibody binding was detected using a DAB stock stain kit (Sino-American Biotechnology Co., Luoyang, China).
AAACCCATCGCATAAAAGTG-3') (the boxed sequences indicate the BamH I and EcoR I restriction sites, respectively). The PCR product was inserted into the BamH I and EcoR I sites of pET30, and transformed into the E. coli BL21 strain. After 3 h preincubation, the recombinant LstiCBP was induced by adding isopropyl-beta-d-thiogalactopyranoside (IPTG) at a final concentration of 2.0 mM for 4 h. The cells (1 L) were harvested by centrifugation, and the pellets were homogenized in phosphate-buffered saline (PBS, 0.04 M, pH 7.0). After centrifugation at 12,000× g for 20 min at 4 °C, the supernatants were dried and stored at −70 °C until use. Before SDS-PAGE, the cells were thawed and disrupted in PBS by sonication (5 s, 5 passes, 4 °C). The cell lysates from the cells before or after induction with IPTG were mixed with SDS-PAGE sample buffer, boiled for 10 min, centrifugation at 12,000× g for 10 min and supernatants were loaded onto an 8% SDS-PAGE gel. Western blot was performed following the description of Wei et al. [36]. After SDS-PAGE, the proteins were blotted onto a PVDF membrane (Hybond-P, Amersham, Zhengzhou, China), and the membrane was incubated with antibodies to 6 × His for 2 h at 37 °C. After washing in PBST (PBS-Tween, Sigma-aldrich, Shanghai, China), the membrane was incubated with secondary antibodies (HRP-conjugated goat anti-rabbit IgG, dilution 1/2000) for 2 h at 37 °C and then washed thoroughly with PBST. Antibody binding was detected using a DAB stock stain kit (Sino-American Biotechnology Co., Luoyang, China).3.5. Preparation of Antibodies that React to LstiCBP
3.6. Western Blot Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lehane, M.J. Peritrophic matrix structure and function. Annu. Rev. Entomol. 1997, 42, 525–550. [Google Scholar] [CrossRef] [PubMed]
- Tellam, R.L.; Wijffels, G.; Willadsen, P. Peritrophic membrane proteins. Insect Biochem. Mol. Biol. 1999, 29, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, R.; Terra, W.R.; Ribeiro, A.F.; Ferreira, C. The peritrophic membrane of Spodoptera frugiperda: Secretion of peritrophins and role in immobilization and recycling digestive enzymes. Arch. Insect Biochem. Physiol. 2001, 47, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.H.; Yuan, Z.M. Peritrophic membrane: A potential target for biocontrol of pest insects. Acta Entomol. Sin. 2005, 48, 968–974. [Google Scholar]
- Casu, R.; Eisemann, C.H.; Pearson, R.D.; Riding, G.A.; East, L.J.; Donaldson, A.; Cadogan, L.; Tellam, R.L. Antibody-mediated inhibition of the growth of larvae from an insect causing sutaneous myiasis in a mammalian host. Proc. Natl. Acad. Sci. USA 1997, 94, 8939–8944. [Google Scholar] [CrossRef] [PubMed]
- Elvin, C.M.; Vuocolo, T.; Pearson, R.D.; East, L.J.; Riding, G.A.; Eisemann, C.H.; Tellam, R.L. Characterization of a major peritrophic membrane protein, peritrophin-44, from the larvae of Lucilia cuprina: cDNA and deduced amino acid sequences. J. Biol. Chem. 1996, 271, 8925–8935. [Google Scholar] [CrossRef] [PubMed]
- Schorderet, S.; Pearson, R.D.; Vuocolo, T.; Eisemann, C.H.; Riding, G.A.; Tellam, R.L. cDNA and deduced amino acid sequences of a peritrophic membrane glycoprotein, “peritrophin-48”, from the larvae of Lucilia cuprina. Insect Biochem. Mol. Biol. 1998, 28, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Granados, R.R. Molecular cloning and sequencing of a novel invertebrate intestinal mucin cDNA. J. Biol. Chem. 1997, 272, 16663–16669. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Jacobs-Lorena, M. A type I peritrophic matrix protein from the malaria vector Anopheles gambiae binds to chitin. Cloning, expression and characterization. J. Biol. Chem. 1998, 273, 17665–17670. [Google Scholar] [CrossRef]
- Wang, P.; Granados, R.R. Molecular structure of the peritrophic membrane (PM): Identification of potential PM target sites for insect control. Arch. Insect Biochem. Physiol. 2001, 47, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Vuocolo, T.; Eisemann, C.H.; Pearson, R.D.; Willadsen, P.; Tellam, R.L. Identification and molecular characterization of a peritrophin gene, peritrophin 48, from the myiasis fly Chrysomya bezziana. Insect Biochem. Mol. Biol. 2001, 31, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Wijffels, G.; Eisemann, C.H.; Riding, G.A.; Pearson, R.D.; Jones, A.; Willadsen, P.; Tellam, R.L. A novel family of chitin-binding proteins from insect type 2 peritrophic matrix. cDNA sequences, chitin binding activity, and cellular localization. J. Biol. Chem. 2001, 276, 15527–15536. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Aksoy, S. Proventriculus-specific cDNAs characterized from the tsetse, Glossina morsitans morsitans. Insect Biochem. Mol. Biol. 2002, 32, 1663–1671. [Google Scholar] [CrossRef]
- Tellam, R.L.; Vuocolo, T.; Eisemann, C.H.; Briscoe, S.; Riding, G.A.; Elvin, C.M.; Pearson, R.D. Identification of an immuno-protective mucin-like protein, peritrophin-55, from the peritrophic matrix of Lucilia cuprina larvae. Insect Biochem. Mol. Biol. 2003, 33, 239–252. [Google Scholar] [CrossRef]
- Sarauer, B.L.; Gillott, C.; Hegedust, D. Characterization of an intestinal mucin from the peritrophic matrix of the diamondback moth, Plutella xylostella. Insect Mol. Biol. 2003, 12, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chamankhah, M.; Visal-Shah, S.; Hemmingsen, S.M.; Erlandson, M.; Braun, L.; Alting-mees, M.; Khachatourians, G.G.; Grady, M.O.; Hegedus, D.D. Modeling the structure of the type I peritrophic matrix: Characterization of a Mamestra configurata intestinal mucin and a novel peritrophin containing 19 chitin binding domains. Insect Biochem. Mol. Biol. 2004, 34, 1101–1115. [Google Scholar] [CrossRef]
- Devenport, M.; Fujioka, H.; Donnelly-Doman, M.; Shen, Z.; Jacobs-Lorena, M. Storage and secretion of Ag-Aper14, a novel peritrophic matrix protein, and Ag-Muc1 from the mosquito Anopheles gambiae. Cell Tissue Res. 2005, 320, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Li, G.; Pang, Y.; Wang, P. A novel chitin-binding protein identified from the peritrophic membrane of the cabbage looper, Trichoplusiani. Insect Biochem. Mol. Biol. 2005, 35, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, J.; Guo, W.; Li, G.X. A Method of rapid and efficient isolation of highly specific antibodies from an antiserum against a pool of proteins. Acta Agric. Boreali-Sin. 2008, 23, 111–113. [Google Scholar]
- Yang, H.; Zhou, F.; Malik, F.A.; Bhaskar, R.; Li, X.; Hu, J.; Sun, C.; Miao, Y. Identification and characterization of two chintin-binding proteins from the peritrophic membrane of the silkworm, Bombyx mori L. Arch. Insect Biochem. Physiol. 2010, 75, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Guo, W. Isolation and Identification of Insect Intestinal Mucin Haiim86—The New Target for Helicoverpa Armigera Biocontrol. Int. J. Biol. Sci. 2011, 7, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Chen, L.; Xiang, X.; Yang, R.; Yu, S.; Wu, X. Proteomic analysis of peritrophic membrane (PM) from the midgut of fifth-instar larvae, Bombyx mori. Mol. Biol. Rep. 2012, 39, 3427–3434. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Zhang, L.; Zou, Y.; Yi, Q.; Zhao, P.; Xia, Q.; Xiang, Z. Shotgun analysis on theperitrophic membrane of the silkworm Bombyx mori. BMB Rep. 2012, 45, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Chen, L.; Yang, R.; Xiang, X.; Yu, S.; Wu, X. Molecular characterization of a peritrophic membrane protein from the silkworm, Bombyx mori. Mol. Biol. Rep. 2013, 40, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Guo, W.; Xu, D.; Zhao, D.; Sun, W.; Shan, M. Eukaryotic expression and analysis of chitin-binding of intestinal mucin HaIIM72 from Helicoverpa armigera (Hübner) Lepidoptera Noctuidae. J. Agric. Univ. Hebei 2013, 36, 92–96. [Google Scholar]
- Zhong, X.W.; Wang, X.H.; Tan, X.; Xia, Q.Y.; Xiang, Z.H.; Zhao, P. Identification and Molecular Characterization of a Chitin Deacetylase from Bombyx mori Peritrophic Membrane. Int. J. Mol. Sci. 2014, 15, 1946–1961. [Google Scholar] [CrossRef]
- Liu, X.; Li, J.; Guo, W.; Li, R.; Zhao, D.; Li, X. A new type I peritrophic membrane protein from larval Holotrichia oblita (Coleoptera: Melolonthidae) Binds to Chitin. Int. J. Mol. Sci. 2014, 15, 6831–6842. [Google Scholar] [CrossRef] [PubMed]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New insights into peritrophic matrix synthesis, architecture, and function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Granados, R.R. An intestinal mucin is the target substrate for a baculovirus enhancin. Proc. Natl. Acad. Sci. USA 1997, 94, 6977–6982. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E. Chitin biochemistry: Synthesis, hydrolysis and inhibition. Adv. Insect Physiol. 2010, 38, 5–74. [Google Scholar]
- Qu, X.F.; Shao, Z.R.; Wang, J.Q. Analysis of periodic outbreak of meadow moth in agricultural and pastoral area of North China. Entomol. Knowl. 1999, 36, 11–14. [Google Scholar]
- Wang, P.; Li, G.; Granado, R.R. Identification of two new peritrophic membrane proteins from larval Trichoplusia ni: Structural characteristics and their functions in the protease rich insect gut. Insect Biochem. Mol. Biol. 2004, 34, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; John, M.W., Ed.; Humana Press: Clifton, NJ, USA, 2005. [Google Scholar]
- Jasrapuria, S.; Arakane, Y.; Osman, G.; Kramer, K.J.; Beeman, R.W.; Muthukrishnan, S. Genes encoding proteins with peritrophin A-type chitin-binding domains in Tribolium castaneum are grouped into three distinct families based on phylogeny, expression and function. Insect Biochem. Mol. Biol. 2010, 40, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression datausing real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.J.; Zhang, Q.R.; Kang, L.; Xu, W.H.; Denlinger, D.L. Molecular characterization and expression of prothoracicotropic hormone during developmentand pupal diapause in the cotton bollworm, Helicoverpa armigera. J. Insect Physiol. 2005, 51, 691–700. [Google Scholar] [CrossRef]
- Molano, J.; Durán, A.; Cabib, E. A rapid and sensitive assay for chitinase using tritiated chitin. Anal. Biochem. 1977, 83, 648–656. [Google Scholar] [CrossRef]
- Wang, P.; Granados, R.R. Rapid and efficient isolation of highly specific antibodies from an antiserum against a pool of proteins. Biotech. Histochem. 2003, 78, 201–205. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, J.; Yang, S.; Li, K.; Guo, W.; Cao, Y. Identification and Molecular Characterization of a Chitin-Binding Protein from the Beet Webworm, Loxostege sticticalis L. Int. J. Mol. Sci. 2014, 15, 19147-19161. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151019147
Yin J, Yang S, Li K, Guo W, Cao Y. Identification and Molecular Characterization of a Chitin-Binding Protein from the Beet Webworm, Loxostege sticticalis L. International Journal of Molecular Sciences. 2014; 15(10):19147-19161. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151019147
Chicago/Turabian StyleYin, Jiao, Shuang Yang, Kebin Li, Wei Guo, and Yazhong Cao. 2014. "Identification and Molecular Characterization of a Chitin-Binding Protein from the Beet Webworm, Loxostege sticticalis L." International Journal of Molecular Sciences 15, no. 10: 19147-19161. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151019147