Betulinic Acid Derivatives NVX-207 and B10 for Treatment of Glioblastoma—An in Vitro Study of Cytotoxicity and Radiosensitization


 ,
,
Abstract
:1. Introduction

2. Results and Discussion
2.1. Results
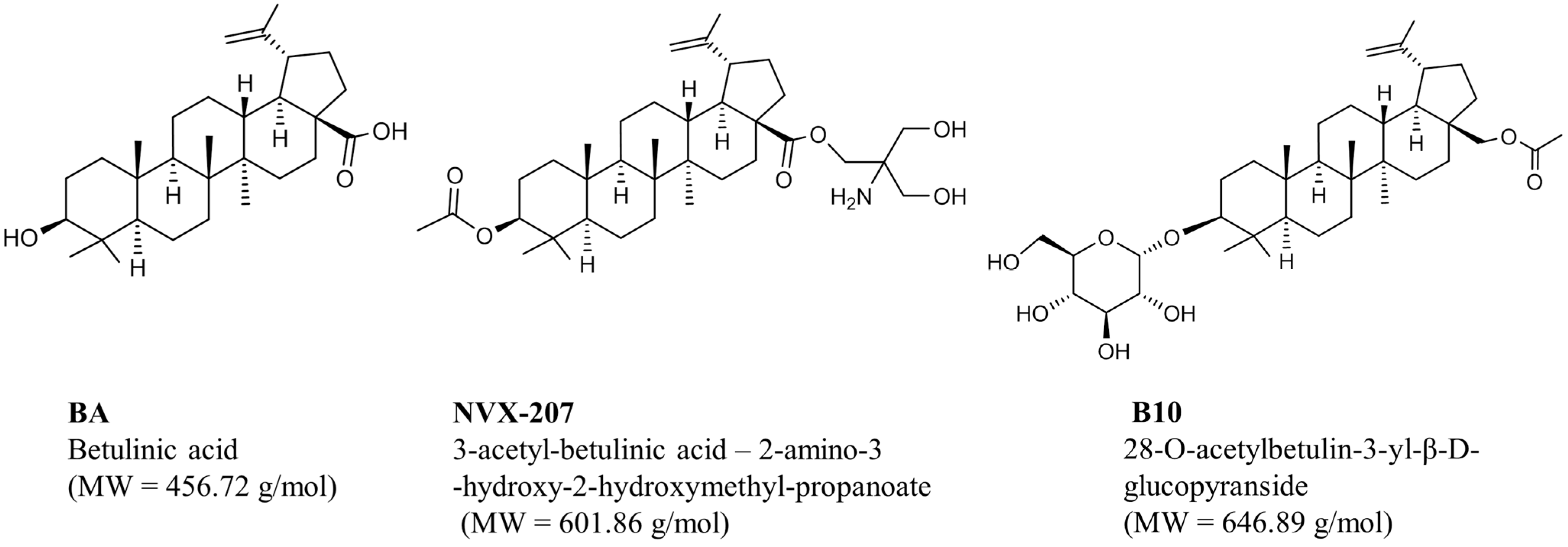
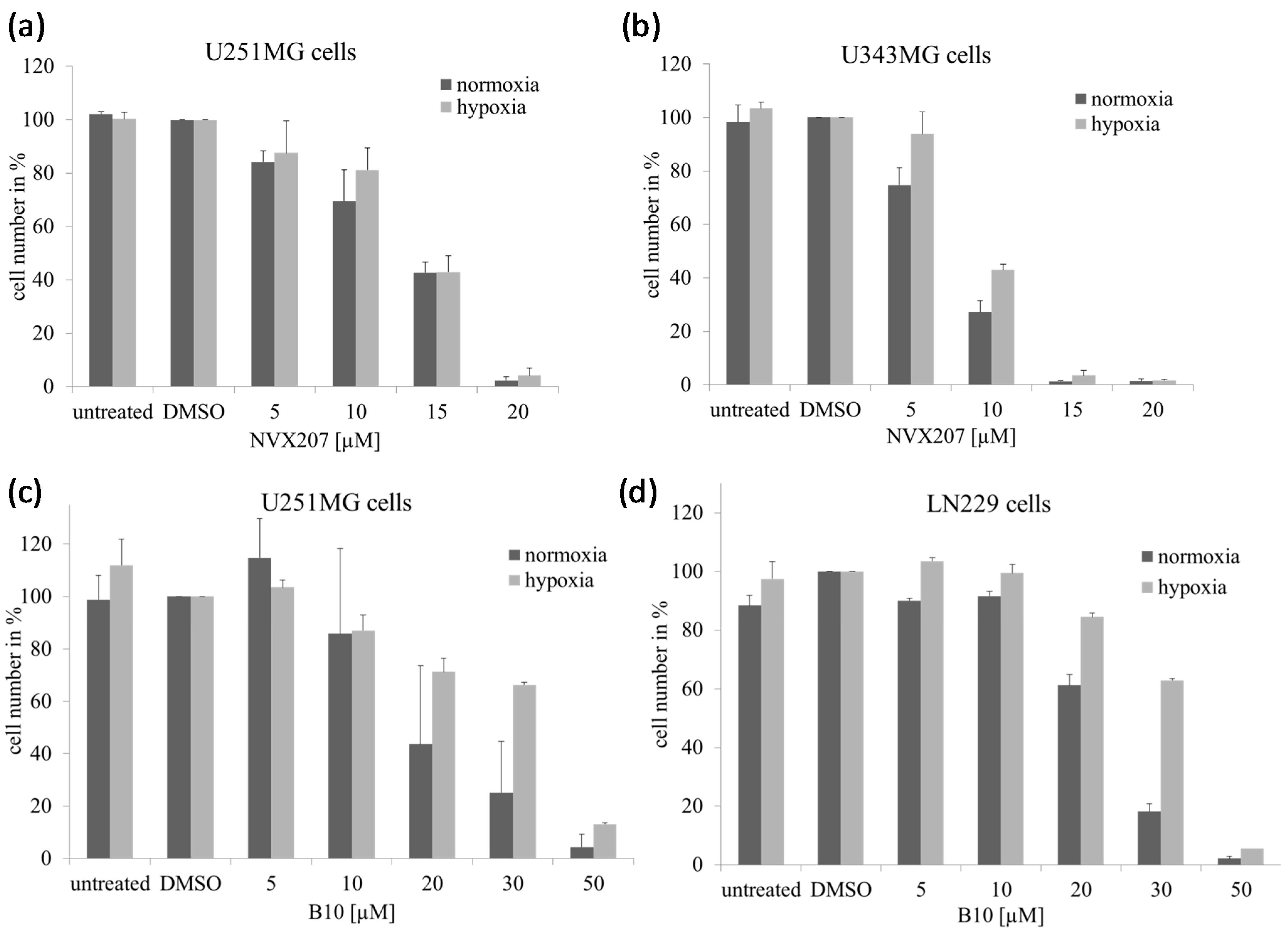
2.1.1. Effects of Betulinic Acid (BA) and BA Derivatives on the Cytotoxicity and Protein Expression of Malignant Glioma Cell Lines under Normoxic and Hypoxic Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | IC50 (µM) | |||||
|---|---|---|---|---|---|---|
| U251MG | LN229 | U343MG | ||||
| N | H | N | H | N | H | |
| BA | 18.4 ± 3.7 | 7.6 ± 1.0 | 23.1 ± 5.3 | 8.5 ± 0.7 | 20.2 ± 4.4 | 7.0 ± 1.2 |
| NVX-207 | 7.6 ± 0.4 | 9.6 ± 0.9 | 7.9 ± 0.5 | 8.5 ± 0.3 | 8.5 ± 0.8 | 8.3 ± 0.1 |
| B10 | 8.1 ± 0.8 | 11.4 ± 3.7 | 17.2 ± 1.5 | 22.4 ± 3.7 | 15.8 ± 1.7 | 24.9 ± 4.8 |



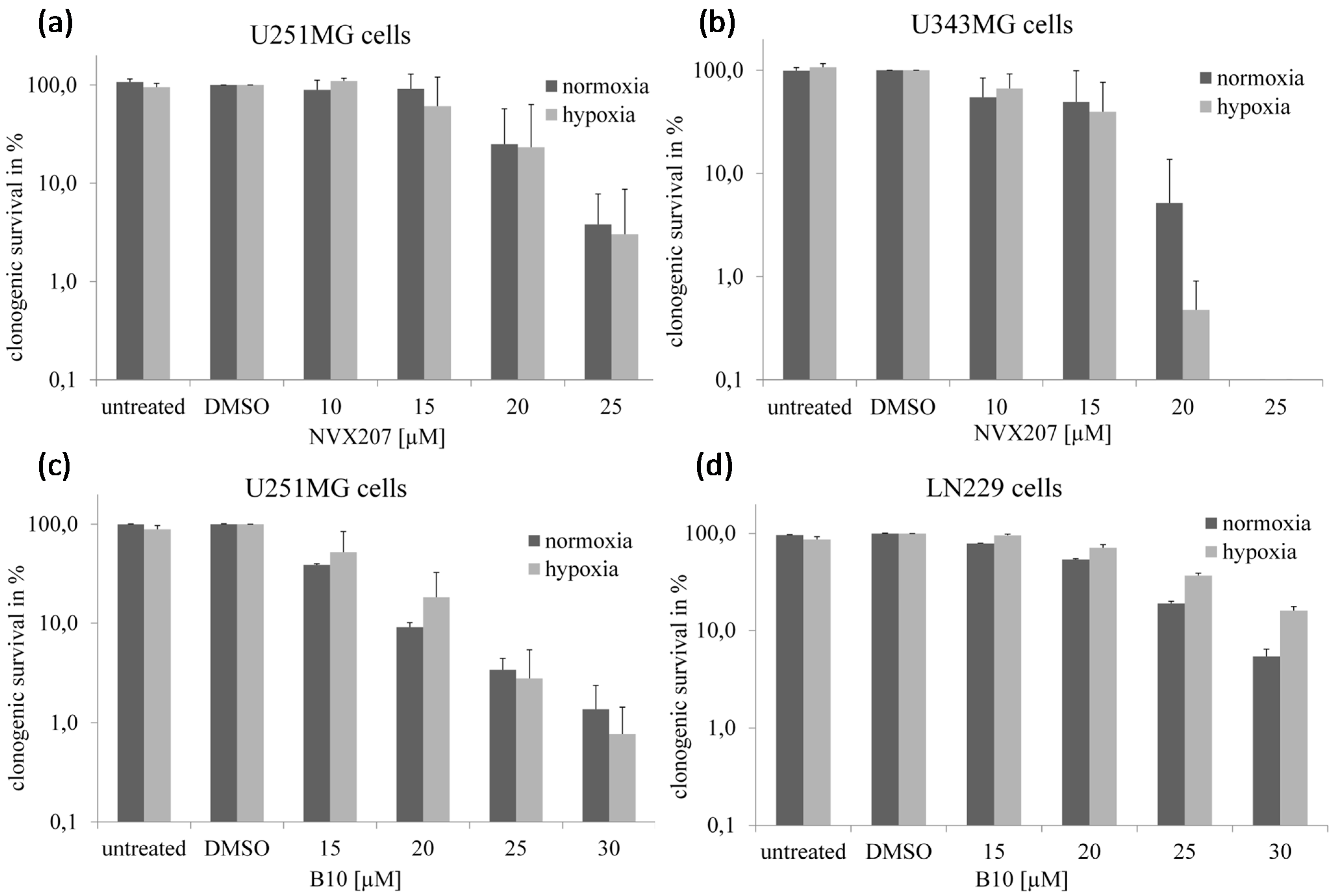
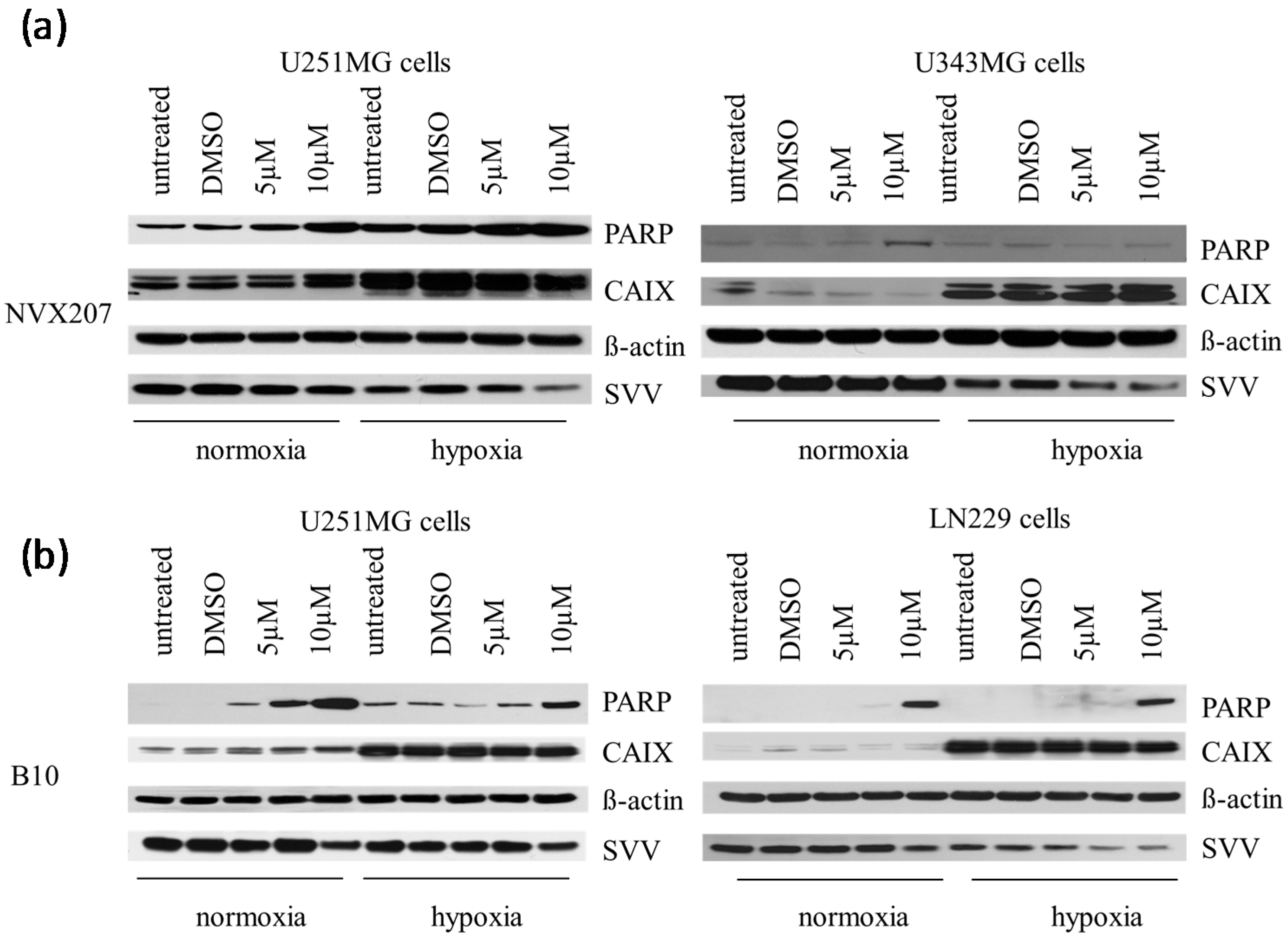
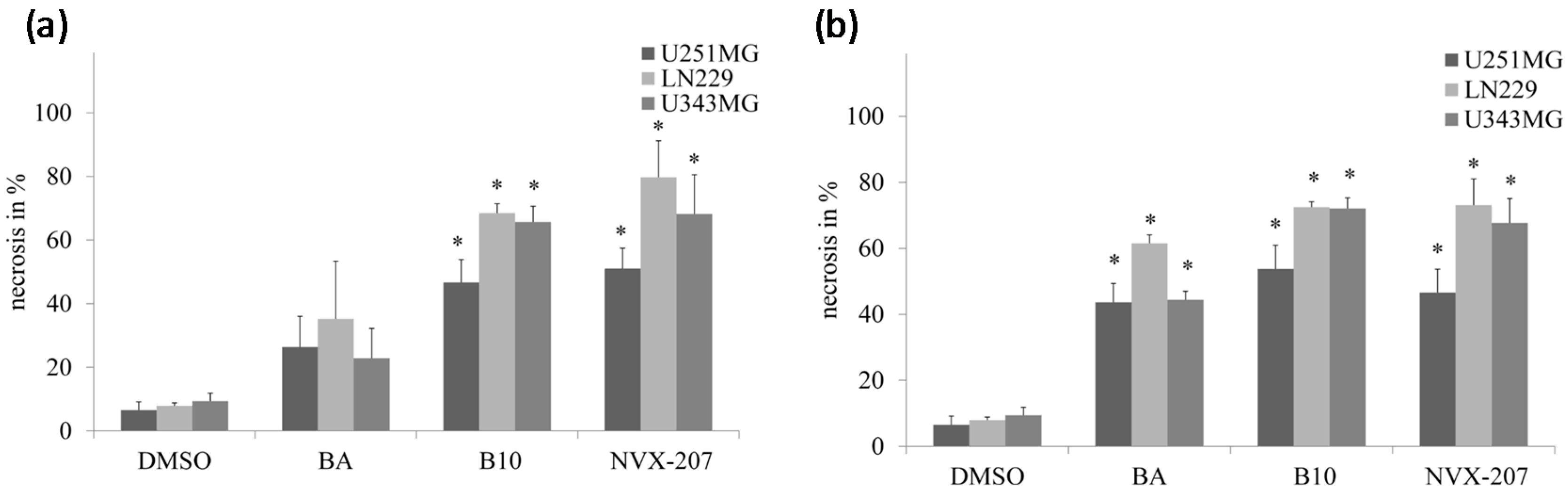
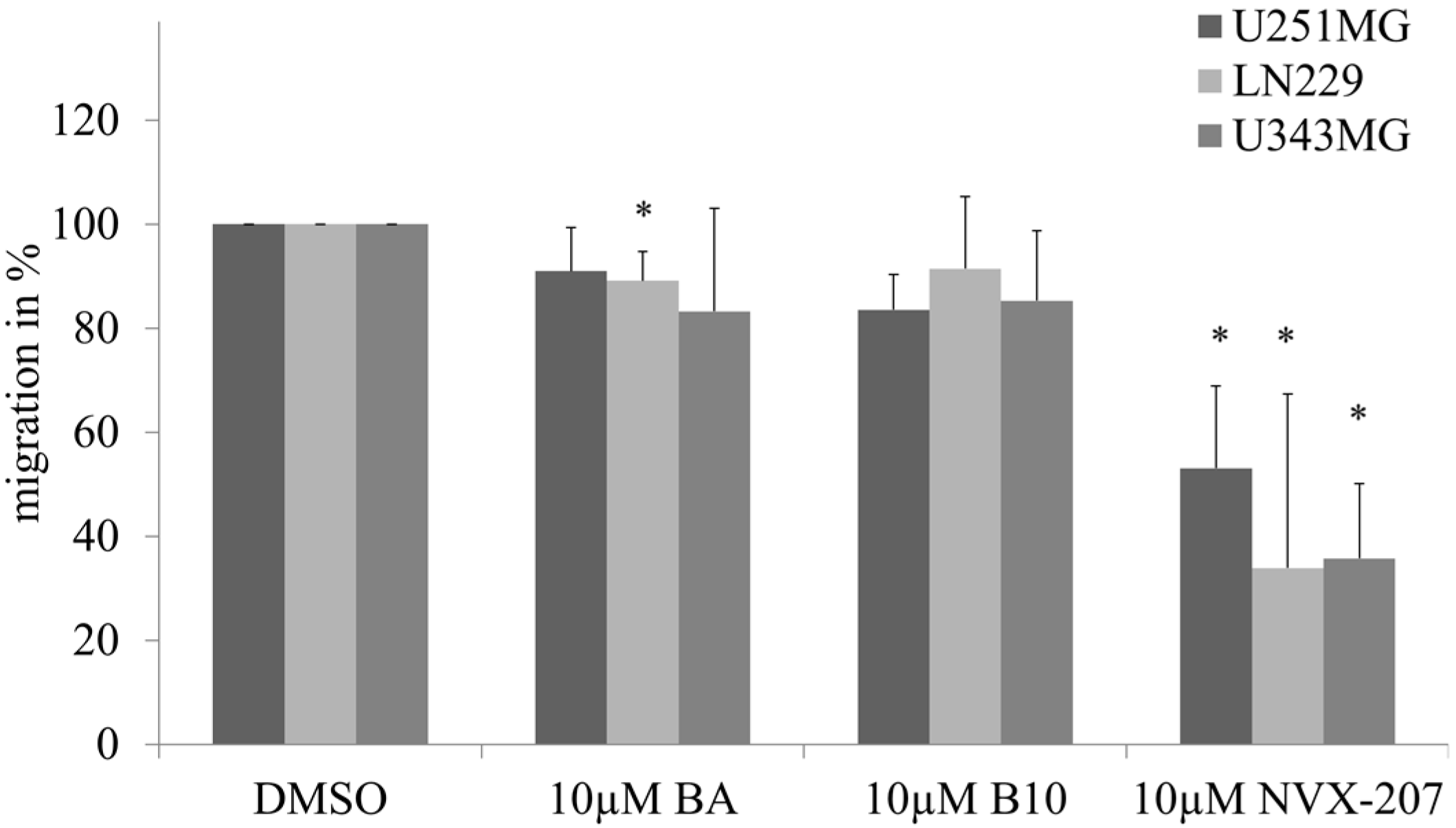
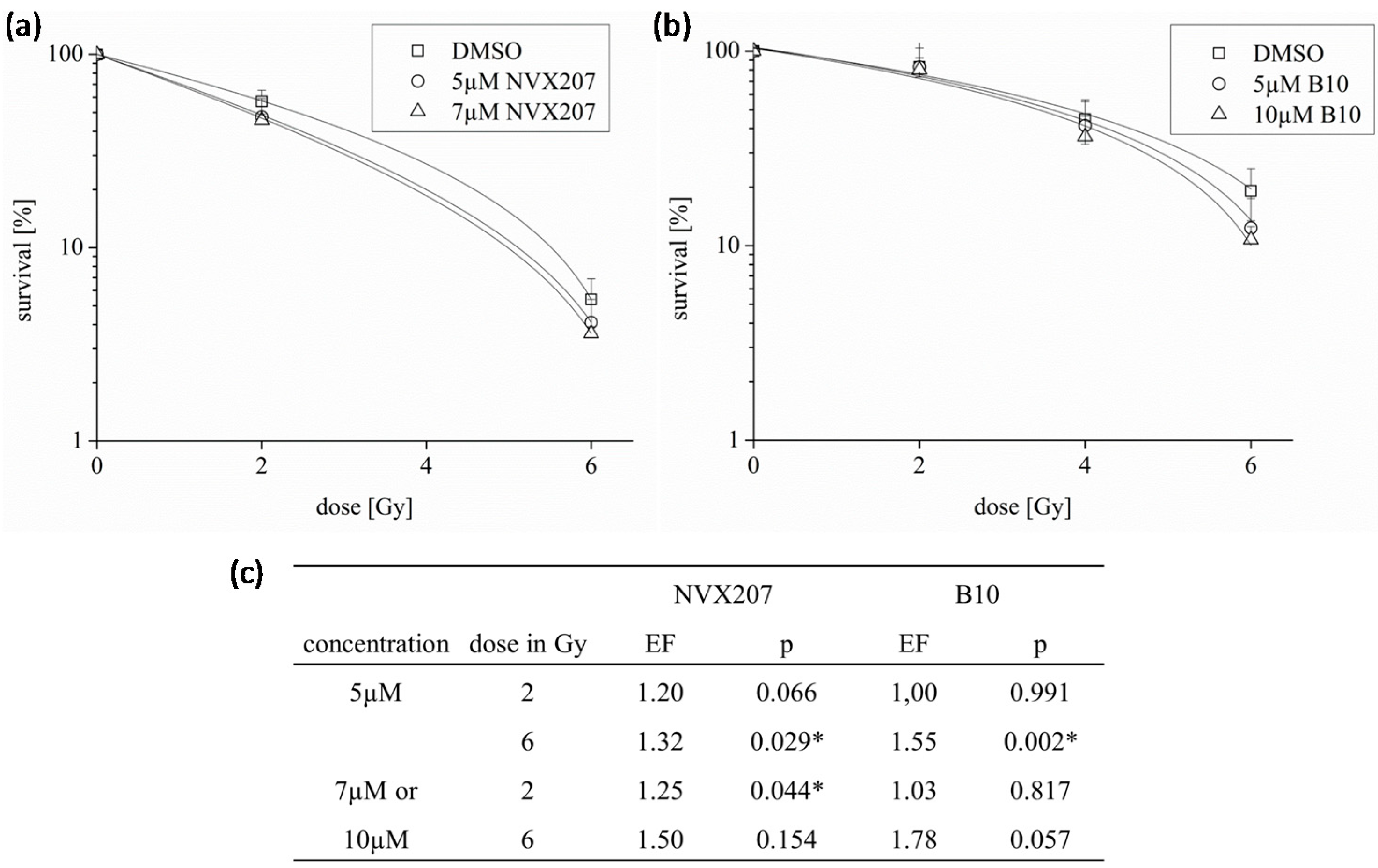
2.1.2. Effects of BA and BA Derivatives on Necrosis, Migration, and Radiosensitivity in Malignant Glioma Cell Lines



2.2. Discussion
3. Experimental Section
3.1. Cell Lines, Culture Conditions, and Treatments with BA and Irradiation
3.2. Sulforhodamine-B (SRB) Assay
3.3. Lactate Dehydrogenase (LDH) Assay
3.4. Scratch Assay
3.5. Western Blotting
3.6. Clonogenic Survival Assays and Radiosensitivity
3.7. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kortmann, R.D.; Jeremic, B.; Weller, M.; Plasswilm, L.; Bamberg, M. Radiochemotherapy of malignant glioma in adults. Clinical experiences. Strahlenther. Onkol. 2003, 179, 219–232. [Google Scholar]
- Henriksson, R.; Asklund, T.; Poulsen, H.S. Impact of therapy on quality of life, neurocognitive function and their correlates in glioblastoma multiforme: A review. J. Neurooncol. 2011, 104, 639–646. [Google Scholar]
- Nieder, C.; Mehta, M.P. Advances in translational research provide a rationale for clinical re-evaluation of high-dose radiotherapy for glioblastoma. Med. Hypotheses 2011, 76, 410–413. [Google Scholar] [CrossRef]
- Shirai, K.; Chakravarti, A. Towards personalized therapy for patients with glioblastoma. Expert Rev. Anticancer Ther. 2011, 11, 1935–1944. [Google Scholar] [CrossRef]
- Alakurtti, S.; Mäkelä, T.; Koskimies, S.; Yli-Kauhaluoma, J. Pharmacological properties of the ubiquitous natural product betulin. Eur. J. Pharm. Sci. 2006, 29, 1–13. [Google Scholar] [CrossRef]
- Kessler, J.H.; Mullauer, F.B.; de Roo, G.M.; Medema, J.P. Broad in vitro efficacy of plant-derived betulinic acid against cell lines derived from the most prevalent human cancer types. Cancer Lett. 2007, 251, 132–145. [Google Scholar] [CrossRef]
- Fulda, S.; Jeremias, I.; Steiner, H.H.; Pietsch, T.; Debatin, K.M. Betulinic acid: A new cytotoxic agent against malignant brain-tumor cells. Int. J. Cancer 1999, 82, 435–441. [Google Scholar] [CrossRef]
- Wick, W.; Grimmel, C.; Wagenknecht, B.; Dichgans, J.; Weller, M. Betulinic acid-induced apoptosis in glioma cells: A sequential requirement for new protein synthesis, formation of reactive oxygen species, and caspase processing. J. Pharmacol. Exp. Ther. 1999, 289, 1306–1312. [Google Scholar]
- Jeremias, I.; Steiner, H.H.; Benner, A.; Debatin, K.M.; Herold-Mende, C. Cell death induction by betulinic acid, ceramide and TRAIL in primary glioblastoma multiforme cells. Acta Neurochir. 2004, 146, 721. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Sensitization for anticancer drug-induced apoptosis by betulinic acid. Neoplasia 2005, 7, 162–170. [Google Scholar] [CrossRef]
- Jung, G.R.; Kim, K.J.; Choi, C.H.; Lee, T.B.; Han, S.I.; Han, H.K.; Lim, S.C. Effect of betulinic acid on anticancer drug-resistant colon cancer cells. Basic. Clin. Pharmacol. Toxicol. 2007, 101, 277–285. [Google Scholar] [CrossRef]
- Selzer, E.; Pimentel, E.; Wacheck, V.; Schlegel, W.; Pehamberger, H.; Jansen, B.; Kodym, R. Effects of betulinic acid alone and in combination with irradiation in human melanoma cells. J. Investig. Dermatol. 2000, 114, 935–940. [Google Scholar] [CrossRef]
- Eder-Czembirek, C.; Erovic, B.M.; Czembirek, C.; Brunner, M.; Selzer, E.; Pötter, R.; Thurnher, D. Betulinic acid a radiosensitizer in head and neck squamous cell carcinoma cell lines. Strahlenther. Onkol. 2010, 186, 143–148. [Google Scholar] [CrossRef]
- Bache, M.; Zschornak, M.P.; Passin, S.; Kessler, J.; Wichmann, H.; Kappler, M.; Paschke, R.; Kaluđerović, G.N.; Kommera, H.; Taubert, H.; et al. Increased betulinic acid induced cytotoxicity and radiosensitivity in glioma cells under hypoxic conditions. Radiat. Oncol. 2011, 6, 111. [Google Scholar] [CrossRef]
- Kumar, V.; Rani, N.; Aggarwal, P.; Sanna, V.K.; Singh, A.T.; Jaggi, M.; Joshi, N.; Sharma, P.K.; Irchhaiya, R.; Burman, A.C. Synthesis and cytotoxic activity of heterocyclic ring-substituted betulinic acid derivatives. Bioorg. Med. Chem. Lett. 2008, 18, 5058–5062. [Google Scholar] [CrossRef]
- Bar, F.M.; Khanfar, M.A.; Elnagar, A.Y.; Liu, H.; Zaghloul, A.M.; Badria, F.A.; Sylvester, P.W.; Ahmad, K.F.; Raisch, K.P.; el Sayed, K.A. Rational design and semisynthesis of betulinic acid analogues as potent topoisomerase inhibitors. J. Nat. Prod. 2009, 72, 1643–1650. [Google Scholar] [CrossRef]
- Santos, R.C.; Salvador, J.A.; Marín, S.; Cascante, M. Novel semisynthetic derivatives of betulin and betulinic acid with cytotoxic activity. Bioorg. Med. Chem. 2009, 17, 6241–6250. [Google Scholar] [CrossRef]
- Smith, P.F.; Ogundele, A.; Forrest, A.; Wilton, J.; Salzwedel, K.; Doto, J.; Allaway, G.P.; Martin, D.E. Phase I and II study of the safety, virologic effect, and pharmacokinetics/pharmacodynamics of single-dose 3-o-(3',3'-dimethylsuccinyl)betulinic acid (bevirimat) against human immunodeficiency virus infection. Antimicrob. Agents Chemother. 2007, 51, 3574–3581. [Google Scholar] [CrossRef]
- Willmann, M.; Wacheck, V.; Buckley, J.; Nagy, K.; Thalhammer, J.; Paschke, R.; Triche, T.; Jansen, B.; Selzer, E. Characterization of NVX-207, a novel betulinic acid-derived anti-cancer compound. Eur. J. Clin. Investig. 2009, 39, 384–394. [Google Scholar] [CrossRef]
- Gauthier, C.; Legault, J.; Lebrun, M.; Dufour, P.; Pichette, A. Glycosidation of lupane-type triterpenoids as potent in vitro cytotoxic agents. Bioorg. Med. Chem. 2006, 14, 6713–6725. [Google Scholar] [CrossRef]
- Kommera, H.; Kaluderović, G.N.; Bette, M.; Kalbitz, J.; Fuchs, P.; Fulda, S.; Mier, W.; Paschke, R. In vitro anticancer studies of α- and β-d-glucopyranose betulin anomers. Chem. Biol. Interact. 2010, 185, 128–136. [Google Scholar]
- Gonzalez, P.; Mader, I.; Tchoghandjian, A.; Enzenmüller, S.; Cristofanon, S.; Basit, F.; Debatin, K.M.; Fulda, S. Impairment of lysosomal integrity by B10, a glycosylated derivative of betulinic acid, leads to lysosomal cell death and converts autophagy into a detrimental process. Cell Death Differ. 2012, 19, 1337–1346. [Google Scholar] [CrossRef]
- Kommera, H.; Kaluderović, G.N.; Kalbitz, J.; Dräger, B.; Paschke, R. Small structural changes of pentacyclic lupane type triterpenoid derivatives lead to significant differences in their anticancer properties. Eur. J. Med. Chem. 2010, 45, 3346–3353. [Google Scholar] [CrossRef]
- Mullauer, F.B.; Kessler, J.H.; Medema, J.P. Betulinic acid, a natural compound with potent anticancer effects. Anticancer Drugs 2010, 21, 215–227. [Google Scholar] [CrossRef]
- Goff, R.D.; Thorson, J.S. Enhancing the divergent activities of betulinic acid via neoglycosylation. Org. Lett. 2009, 11, 461–464. [Google Scholar] [CrossRef]
- Enzenmüller, S.; Gonzalez, P.; Karpel-Massler, G.; Debatin, K.M.; Fulda, S. GDC-0941 enhances the lysosomal compartment via TFEB and primes glioblastoma cells to lysosomal membrane permeabilization and cell death. Cancer Lett. 2013, 329, 27–36. [Google Scholar] [CrossRef]
- Fischer, S.; Ronellenfitsch, M.W.; Thiepold, A.L.; Harter, P.N.; Reichert, S.; Kögel, D.; Paschke, R.; Mittelbronn, M.; Weller, M.; Steinbach, J.P.; et al. Hypoxia enhances the antiglioma cytotoxicity of b10, a glycosylated derivative of betulinic acid. PLoS One 2014, 9, e94921. [Google Scholar] [CrossRef] [PubMed]
- Karna, E.; Szoka, L.; Palka, J.A. Betulinic acid inhibits the expression of hypoxia-inducible factor 1α and vascular endothelial growth factor in human endometrial adenocarcinoma cells. Mol. Cell. Biochem. 2010, 340, 15–20. [Google Scholar] [CrossRef]
- Shin, J.; Lee, H.J.; Jung, D.B.; Jung, J.H.; Lee, H.J.; Lee, E.O.; Lee, S.G.; Shim, B.S.; Choi, S.H.; Ko, S.G.; et al. Suppression of STAT3 and HIF-1α mediates anti-angiogenic activity of betulinic acid in hypoxic PC-3 prostate cancer cells. PLoS One 2011, 6, e21492. [Google Scholar] [CrossRef]
- Dehelean, C.A.; Feflea, S.; Ganta, S.; Amiji, M. Anti-angiogenic effects of betulinic acid administered in nanoemulsion formulation using chorioallantoic membrane assay. J. Biomed. Nanotechnol. 2011, 7, 317–324. [Google Scholar] [CrossRef]
- Ishii, N.; Maier, D.; Merlo, A.; Tada, M.; Sawamura, Y.; Diserens, A.C.; van Meir, E.G. Frequent co-alterations of TP53, p16/CDKN2A, p14ARF, PTEN tumor suppressor genes in human glioma cell lines. Brain Pathol. 1999, 9, 469–479. [Google Scholar] [CrossRef]
- Berger, B.; Capper, D.; Lemke, D.; Pfenning, P.N.; Platten, M.; Weller, M.; von Deimling, A.; Wick, W.; Weiler, M. Defective p53 antiangiogenic signaling in glioblastoma. Neuro Oncol. 2010, 12, 894–907. [Google Scholar] [CrossRef]
- Kappler, M.; Rot, S.; Taubert, H.; Greither, T.; Bartel, F.; Dellas, K.; Hänsgen, G.; Trott, K.R.; Bache, M. The effects of knockdown of wild-type survivin, survivin-2B or survivin-delta3 on the radiosensitization in a soft tissue sarcoma cells in vitro under different oxygen conditions. Cancer Gene Ther. 2007, 14, 994–1001. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bache, M.; Bernhardt, S.; Passin, S.; Wichmann, H.; Hein, A.; Zschornak, M.P.; Kappler, M.; Taubert, H.; Paschke, R.; Vordermark, D. Betulinic Acid Derivatives NVX-207 and B10 for Treatment of Glioblastoma—An in Vitro Study of Cytotoxicity and Radiosensitization. Int. J. Mol. Sci. 2014, 15, 19777-19790. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151119777
Bache M, Bernhardt S, Passin S, Wichmann H, Hein A, Zschornak MP, Kappler M, Taubert H, Paschke R, Vordermark D. Betulinic Acid Derivatives NVX-207 and B10 for Treatment of Glioblastoma—An in Vitro Study of Cytotoxicity and Radiosensitization. International Journal of Molecular Sciences. 2014; 15(11):19777-19790. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151119777
Chicago/Turabian StyleBache, Matthias, Stephan Bernhardt, Sarina Passin, Henri Wichmann, Anja Hein, Martin P Zschornak, Matthias Kappler, Helge Taubert, Reinhard Paschke, and Dirk Vordermark. 2014. "Betulinic Acid Derivatives NVX-207 and B10 for Treatment of Glioblastoma—An in Vitro Study of Cytotoxicity and Radiosensitization" International Journal of Molecular Sciences 15, no. 11: 19777-19790. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151119777