The Daidzein Metabolite, 6,7,4'-Trihydroxyisoflavone, Is a Novel Inhibitor of PKCα in Suppressing Solar UV-Induced Matrix Metalloproteinase 1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results
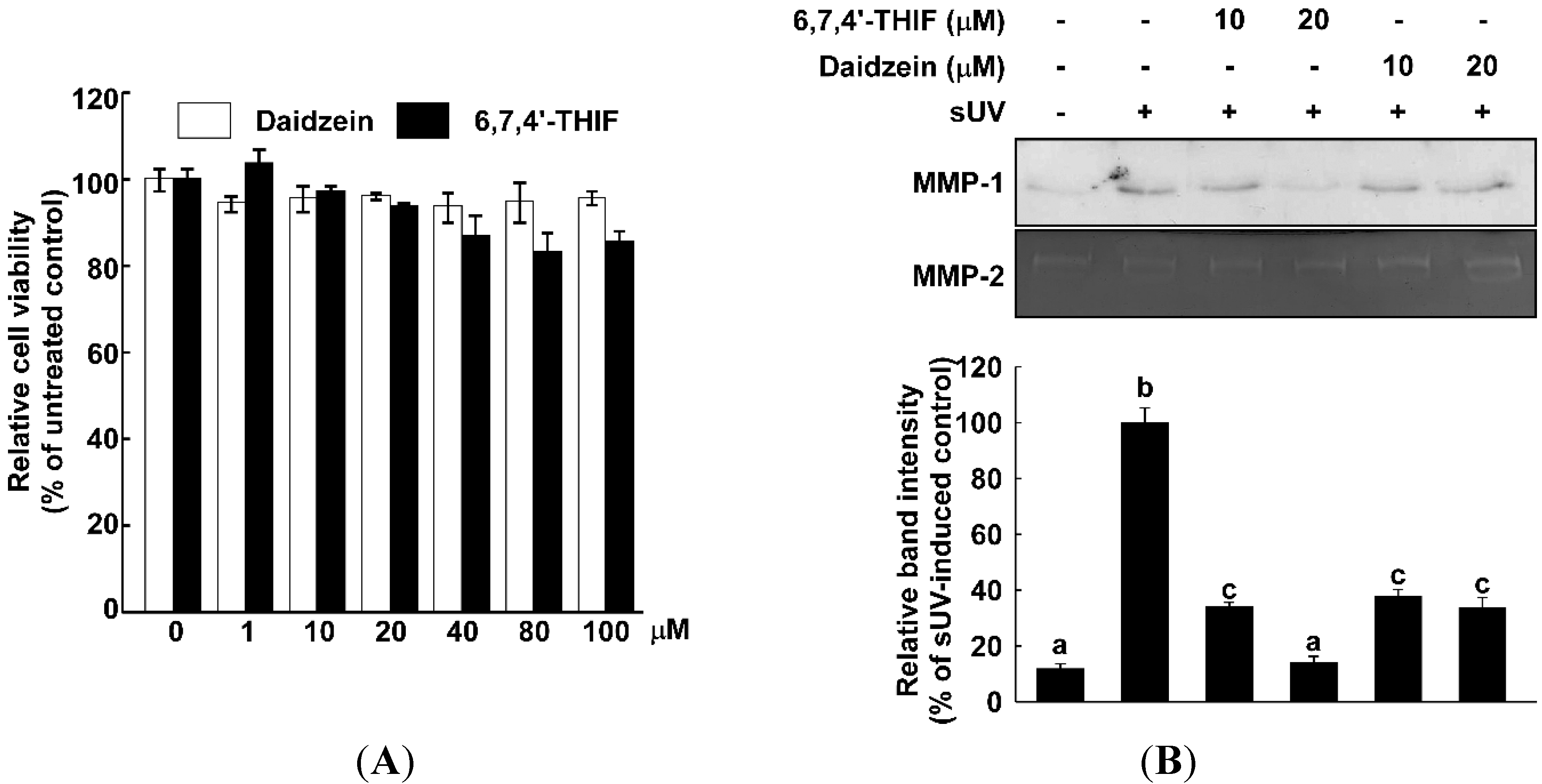
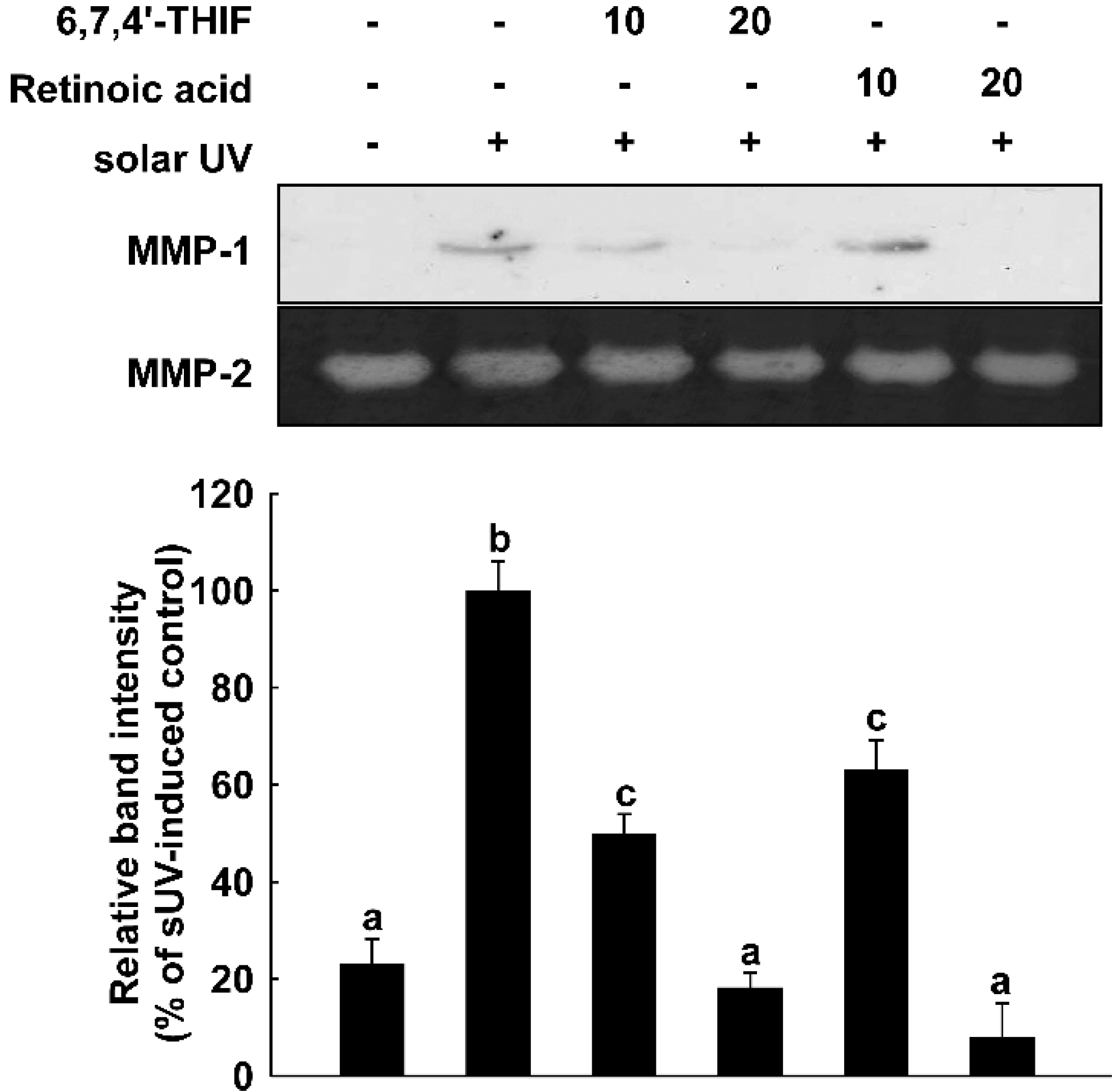
2.1. 6,7,4'-Trihydroxyisoflavone (6,7,4'-THIF) Reduces Solar UV (sUV)-Induced Matrix Metalloproteinase (MMP)-1 Expression in Normal Human Dermal Fibroblasts (NHDF) Cells
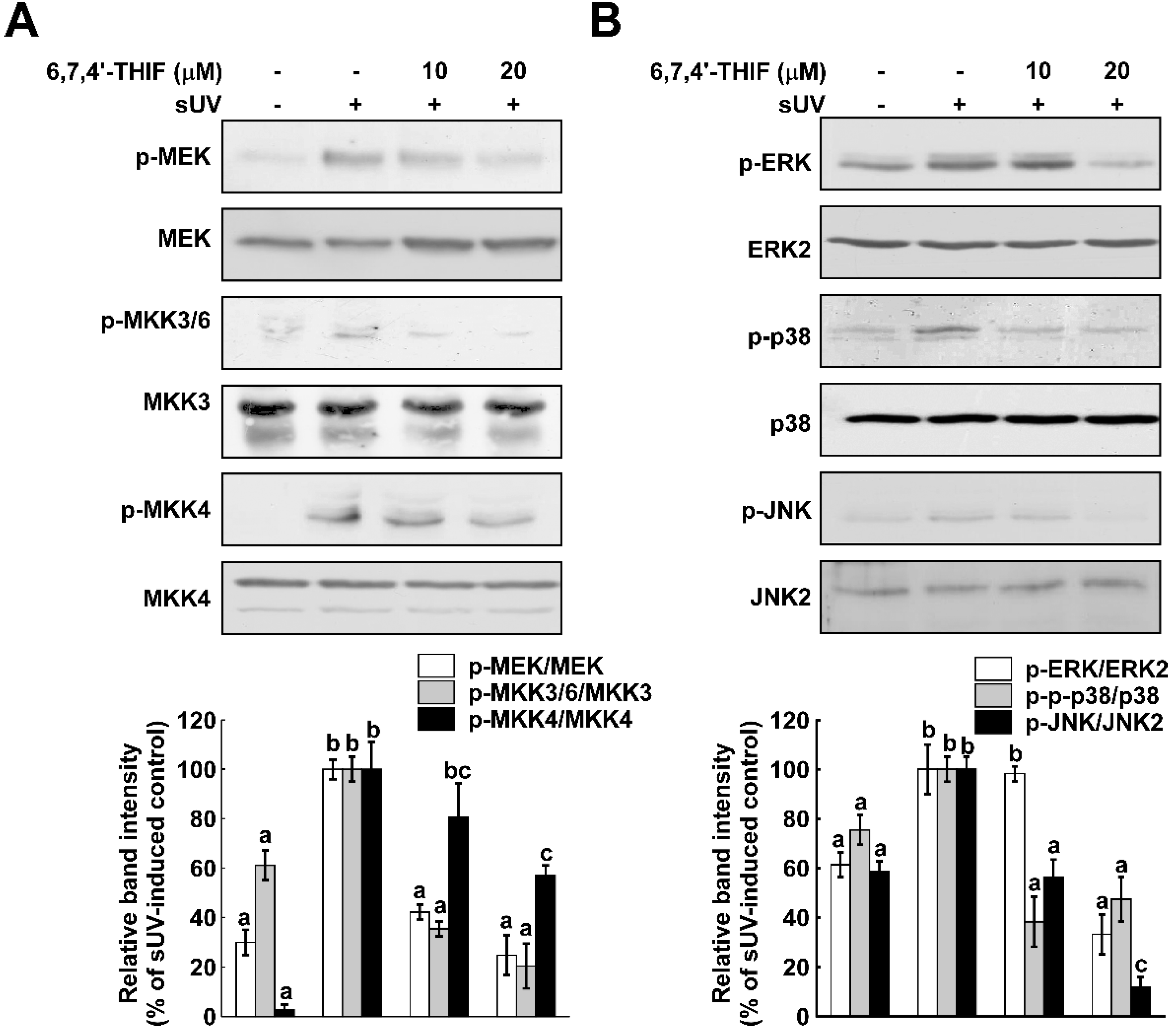
2.2. 6,7,4'-THIF Inhibits sUV-Induced Mitogen-Activated Protein Kinase(MAPK) Signaling Pathways in NHDF Cells

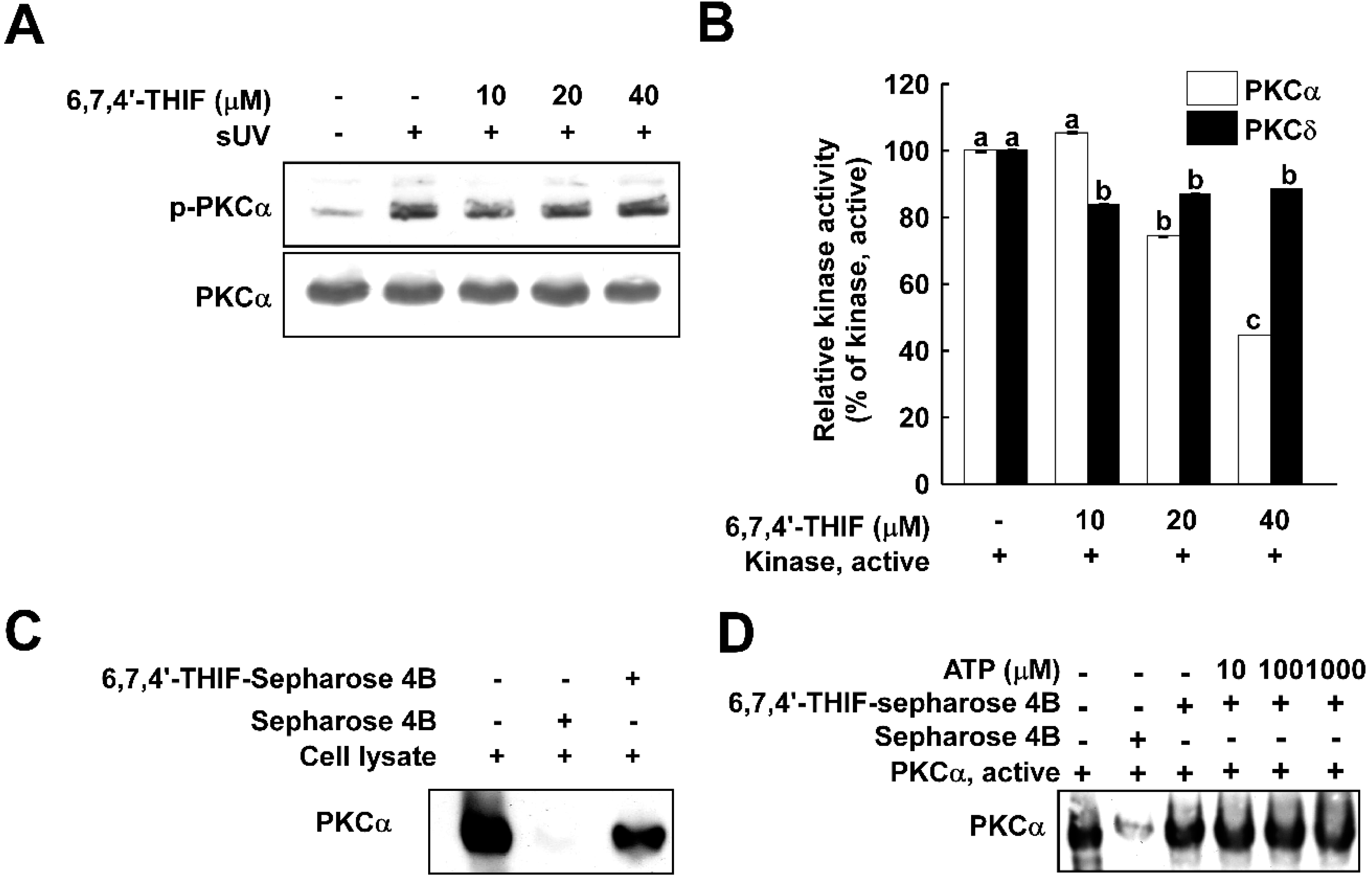
2.3. 6,7,4'-THIF Suppresses Protein Kinase C (PKC)α Kinase Activity with Direct Binding

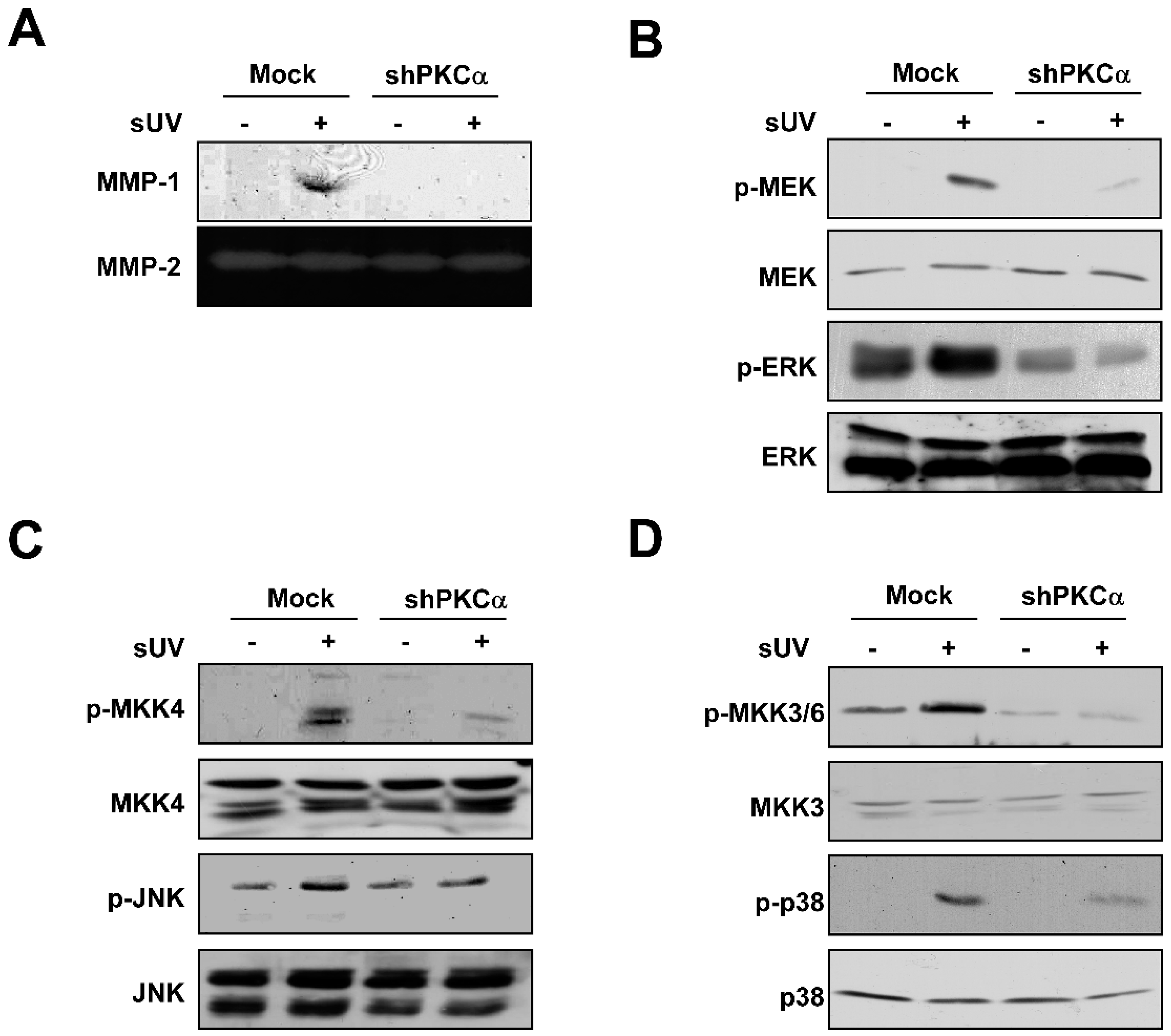
2.4. PKCα Plays a Crucial Role in sUV-Induced MMP-1 Expression in NHDFs


2.5. Discussion

3. Materials and Methods
3.1. Materials
3.2. Cell Culture
3.3. Cell Cytotoxicity
3.4. Solar UV Irradiation
3.5. Western Blot Analysis
3.7. Zymography
3.8. In Vitro Kinase Assay
3.9. 6,7,4'-THIF Pull-Down Assay Using Sepharose 4B
3.10. ATP and PKCα Competition Assay
3.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sjerobabski-Masnec, I.; Situm, M. Skin aging. Acta Clin. Croat. 2010, 49, 515–518. [Google Scholar] [PubMed]
- Gunn, D.A.; Rexbye, H.; Griffiths, C.E.; Murray, P.G.; Fereday, A.; Catt, S.D.; Tomlin, C.C.; Strongitharm, B.H.; Perrett, D.I.; Catt, M.; et al. Why some women look young for their age. PLoS One 2009, 4, e8021. [Google Scholar] [CrossRef] [PubMed]
- Yaar, M.; Gilchrest, B.A. Photoageing: Mechanism, prevention and therapy. Br. J. Dermatol. 2007, 157, 874–887. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, G. Molecular mechanisms of skin ageing. Mech. Ageing Dev. 2002, 123, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Uitto, J. Understanding premature skin aging. N. Engl. J. Med. 1997, 337. [Google Scholar] [CrossRef] [PubMed]
- Yaar, M. Clinical and Histological Features of Intrinsic Versus Extrinsic Skin Aging; Springer: Berlin, Germany, 2006. [Google Scholar]
- Chung, J.H. Photoaging in asians. Photodermatol. Photoimmunol. Photomed. 2003, 19, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.A.; Yi, B.R.; Choi, K.C. Molecular mechanisms and in vivo mouse models of skin aging associated with dermal matrix alterations. Lab. Anim. Res. 2011, 27, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Kim, Y.K.; Jung, J.Y.; Shin, J.E.; Kim, K.H.; Cho, K.H.; Eun, H.C.; Chung, J.H. Intrinsic aging- and photoaging-dependent level changes of glycosaminoglycans and their correlation with water content in human skin. J. Dermatol. Sci. 2011, 62, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Lauer-Fields, J.L.; Juska, D.; Fields, G.B. Matrix metalloproteinases and collagen catabolism. Biopolymers 2002, 66, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.; Bhatti, H.; Nerusu, K.C.; Bhagavathula, N.; Kang, S.; Fisher, G.J.; Varani, J.; Voorhees, J.J. Matrix metalloproteinase-1 is the major collagenolytic enzyme responsible for collagen damage in UV-irradiated human skin. Photochem. Photobiol. 2003, 78, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Lee, D.H.; Lee, S.; Kim, E.J.; Chung, J.H. Uv-induced DNA damage and histone modification may involve MMP-1 gene transcription in human skin in vivo. J. Dermatol. Sci. 2014, 73, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.N.; Lee, Y.H.; Park, J.S.; Jun, W.J.; Kim, H.K.; Hong, B.S.; Shin, D.H.; Cho, H.Y. Protective effects of aucubin isolated from eucommia ulmoides against UVB-induced oxidative stress in human skin fibroblasts. Biol. Pharm. Bull. 2005, 28, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.E.; Lee, K.W.; Jung, S.K.; Lee, E.J.; Hwang, J.A.; Lim, T.G.; Kim, B.Y.; Bode, A.M.; Lee, H.J.; Dong, Z. 6,7,4'-Trihydroxyisoflavone inhibits hct-116 human colon cancer cell proliferation by targeting CDK1 and CDK2. Carcinogenesis 2011, 32, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.E.; Lee, K.W.; Byun, S.; Jung, S.K.; Song, N.; Lim, S.H.; Heo, Y.S.; Kim, J.E.; Kang, N.J.; Kim, B.Y.; et al. 7,3',4'-Trihydroxyisoflavone, a metabolite of the soy isoflavone daidzein, suppresses ultraviolet b-induced skin cancer by targeting cot and MKK4. J. Biol. Chem. 2011, 286, 14246–14256. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hsieh, D.; Yang, Y.L.; Xu, Z.; Peto, C.; Jablons, D.M.; You, L. Coumestrol from the national cancer institute’s natural product library is a novel inhibitor of protein kinase CK2. BMC Pharmacol. Toxicol. 2013, 14, 36. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.G.; Yang, H.; Shin, S.H.; Min, S.; Kim, Y.A.; Yu, J.G.; Lee, D.E.; Chung, M.Y.; Heo, Y.S.; Kwon, J.Y.; et al. A metabolite of daidzein, 6,7,4'-trihydroxyisoflavone, suppresses adipogenesis in 3T3-L1 preadipocytes via ATP-competitive inhibition of PI3K. Mol. Nutr. Food Res. 2013, 57, 1446–1455. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Shi, Y.; Dang, Y.; Zhai, Y.; Ye, X. Daidzein stimulates collagen synthesis by activating the TGF-β/smad signal pathway. Australas. J. Dermatol. 2014. [Google Scholar] [CrossRef]
- Nakamura, Y.; Tsuji, S.; Tonogai, Y. Determination of the levels of isoflavonoids in soybeans and soy-derived foods and estimation of isoflavonoids in the japanese daily intake. J. AOAC Int. 2000, 83, 635–650. [Google Scholar] [PubMed]
- Goh, M.J.; Park, J.S.; Bae, J.H.; Kim, D.H.; Kim, H.K.; Na, Y.J. Effects of ortho-dihydroxyisoflavone derivatives from korean fermented soybean paste on melanogenesis in B16 melanoma cells and human skin equivalents. Phytother. Res. 2012, 26, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.B.; Kim, J.R.; Kim, J.H.; Kim, Y.A.; Park, J.S.; Yeom, M.H.; Joo Lee, H.; Lee, K.W.; Kang, N.J. 7,3',4'-Trihydroxyisoflavone ameliorates the development of dermatophagoides farinae-induced atopic dermatitis in NC/Nga mice. Evid.-Based Complement. Altern. Med. 2013, 2013, 636597. [Google Scholar]
- Lee, Y.M.; Li, W.H.; Kim, Y.K.; Kim, K.H.; Chung, J.H. Heat-induced MMP-1 expression is mediated by TRPV1 through PKCα signaling in HaCaT cells. Exp. Dermatol. 2008, 17, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.G.; Jung, S.K.; Kim, J.E.; Kim, Y.; Lee, H.J.; Jang, T.S.; Lee, K.W. NADPH oxidase is a novel target of delphinidin for the inhibition of UVB-induced MMP-1 expression in human dermal fibroblasts. Exp. Dermatol. 2013, 22, 428–430. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.S.; Hsiao, Y.P.; Lin, J.H.; Hsu, S.C.; Chueh, F.S.; Weng, S.W.; Lai, K.C.; Lin, J.G.; Chung, J.G. Crude extract of Rheum palmatum L. inhibits migration and invasion of LS1034 human colon cancer cells acts through the inhibition of matrix metalloproteinase-2/-9 by MAPK signaling. Environ. Toxicol. 2014. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, H.; Wang, W.; Wang, R.; Liu, Z.L.; Zhu, W.; Lian, S. N-Terminal 5-mer peptide analog P165 of amyloid precursor protein inhibits UVA-induced MMP-1 expression by suppressing the MAPK pathway in human dermal fibroblasts. Eur. J. Pharmacol. 2014, 734C, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ricciarelli, R.; Maroni, P.; Ozer, N.; Zingg, J.M.; Azzi, A. Age-dependent increase of collagenase expression can be reduced by α-tocopherol via protein kinase c inhibition. Free Radic. Biol. Med. 1999, 27, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Muthugounder, S.; Presser, N.; Viswanathan, S. Anticancer therapeutic potential of soy isoflavone, genistein. Adv. Exp. Med. Biol. 2004, 546, 121–165. [Google Scholar] [PubMed]
- Wilson, R. Soybean: Market driven research needs. In Genetics and Genomics of Soybean; Stacey, G., Ed.; Springer: New York, NY, USA, 2008; Volume 2, pp. 3–15. [Google Scholar]
- Chen, K.I.; Erh, M.H.; Su, N.W.; Liu, W.H.; Chou, C.C.; Cheng, K.C. Soyfoods and soybean products: From traditional use to modern applications. Appl. Microbiol. Biotechnol. 2012, 96, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Atherton, K.M.; Mutch, E.; Ford, D. Metabolism of the soyabean isoflavone daidzein by CYP1A2 and the extra-hepatic CYPs 1A1 and 1B1 affects biological activity. Biochem. Pharmacol. 2006, 72, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Do, M.H.; Oh, Y.L.; Cho, D.W.; Kim, S.H.; Kim, S.Y. Dietary fermented soybean suppresses UVB-induced skin inflammation in hairless mice via regulation of the MAPK signaling pathway. J. Agric. Food Chem. 2014, 62, 8962–8972. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Bautista, S.; Nicolas, F.E. Recent patents concerning modulators of protein kinase C. Recent Pat. DNA Gene Seq. 2013, 7, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Basu, A. Protein kinase C-ε promotes emt in breast cancer. Breast Cancer 2014, 8, 61–67. [Google Scholar] [PubMed]
- Chen, Z.; Forman, L.W.; Williams, R.M.; Faller, D.V. Protein kinase C-δ inactivation inhibits the proliferation and survival of cancer stem cells in culture and in vivo. BMC Cancer 2014, 14, 90. [Google Scholar] [CrossRef] [PubMed]
- Mochly-Rosen, D.; Das, K.; Grimes, K.V. Protein kinase C, an elusive therapeutic target? Nat. Rev. Drug Dis. 2012, 11, 937–957. [Google Scholar] [CrossRef]
- Tsao, H.K.; Chiu, P.H.; Sun, S.H. PKC-dependent ERK phosphorylation is essential for P2X7 receptor-mediated neuronal differentiation of neural progenitor cells. Cell Death Dis. 2013, 4, e751. [Google Scholar] [CrossRef] [PubMed]
- Do Carmo, A.; Balca-Silva, J.; Matias, D.; Lopes, M.C. PKC signaling in glioblastoma. Cancer Biol. Ther. 2013, 14, 287–294. [Google Scholar]
- Hu, C.T.; Cheng, C.C.; Pan, S.M.; Wu, J.R.; Wu, W.S. PKC mediates fluctuant ERK-paxillin signaling for hepatocyte growth factor-induced migration of hepatoma cell HepG2. Cell Signal. 2013, 25, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, F.; Benito-Munoz, M.; Panicker, M.; Matute, C. NMDA modulates oligodendrocyte differentiation of subventricular zone cells through PKC activation. Front. Cell. Neurosci. 2013, 7, 261. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.G.; Kwon, J.Y.; Kim, J.; Song, N.R.; Lee, K.M.; Heo, Y.S.; Lee, H.J.; Lee, K.W. Cyanidin-3-glucoside suppresses B[a]PDE-induced cyclooxygenase-2 expression by directly inhibiting Fyn kinase activity. Biochem. Pharmacol. 2011, 82, 167–174. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, T.-G.; Kim, J.-E.; Lee, S.-Y.; Park, J.S.; Yeom, M.H.; Chen, H.; Bode, A.M.; Dong, Z.; Lee, K.W. The Daidzein Metabolite, 6,7,4'-Trihydroxyisoflavone, Is a Novel Inhibitor of PKCα in Suppressing Solar UV-Induced Matrix Metalloproteinase 1. Int. J. Mol. Sci. 2014, 15, 21419-21432. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151121419
Lim T-G, Kim J-E, Lee S-Y, Park JS, Yeom MH, Chen H, Bode AM, Dong Z, Lee KW. The Daidzein Metabolite, 6,7,4'-Trihydroxyisoflavone, Is a Novel Inhibitor of PKCα in Suppressing Solar UV-Induced Matrix Metalloproteinase 1. International Journal of Molecular Sciences. 2014; 15(11):21419-21432. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151121419
Chicago/Turabian StyleLim, Tae-Gyu, Jong-Eun Kim, Sung-Young Lee, Jun Seong Park, Myung Hun Yeom, Hanyong Chen, Ann M. Bode, Zigang Dong, and Ki Won Lee. 2014. "The Daidzein Metabolite, 6,7,4'-Trihydroxyisoflavone, Is a Novel Inhibitor of PKCα in Suppressing Solar UV-Induced Matrix Metalloproteinase 1" International Journal of Molecular Sciences 15, no. 11: 21419-21432. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151121419