Nitric Oxide Functions as a Signal in Ultraviolet-B-Induced Baicalin Accumulation in Scutellaria baicalensis Suspension Cultures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
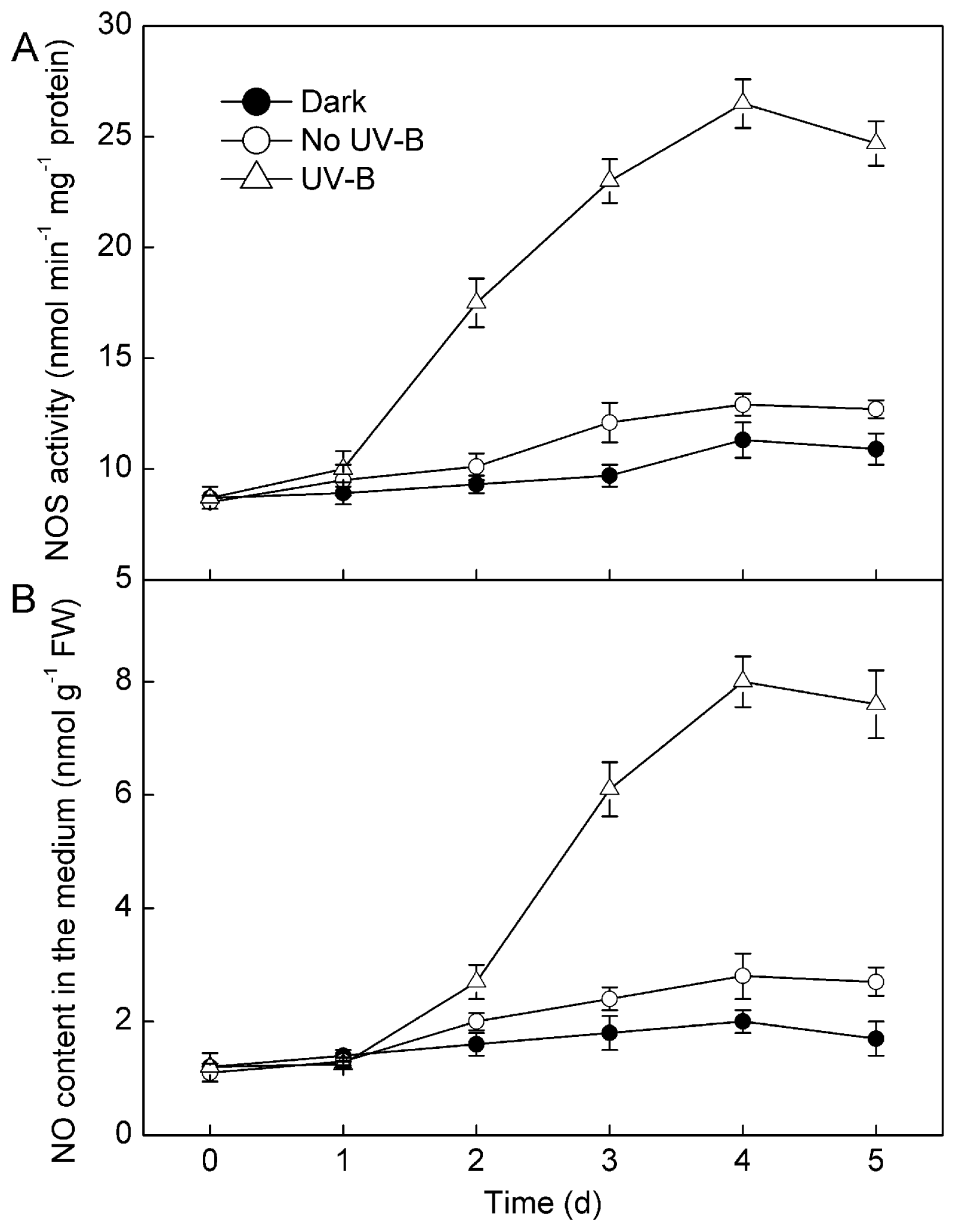
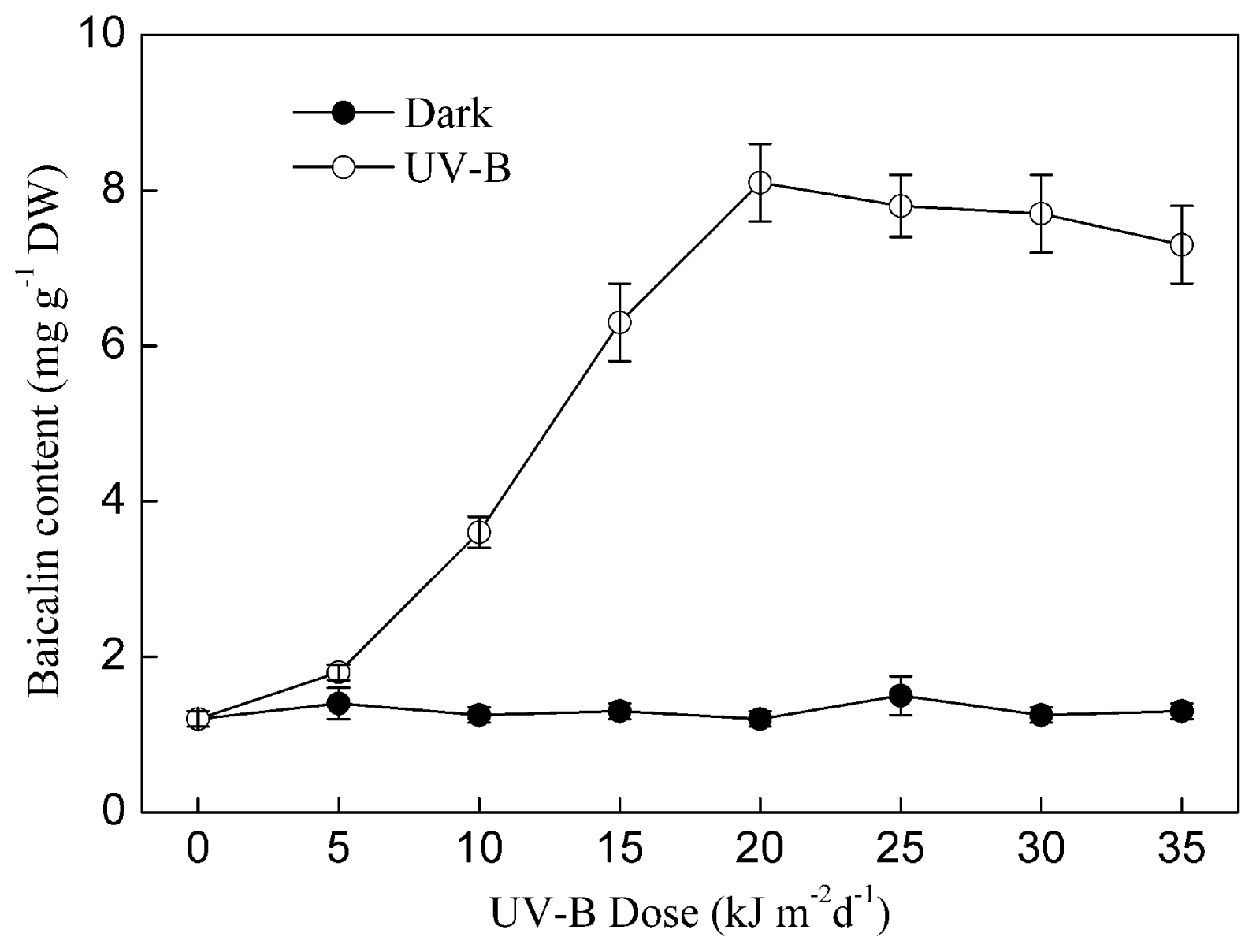
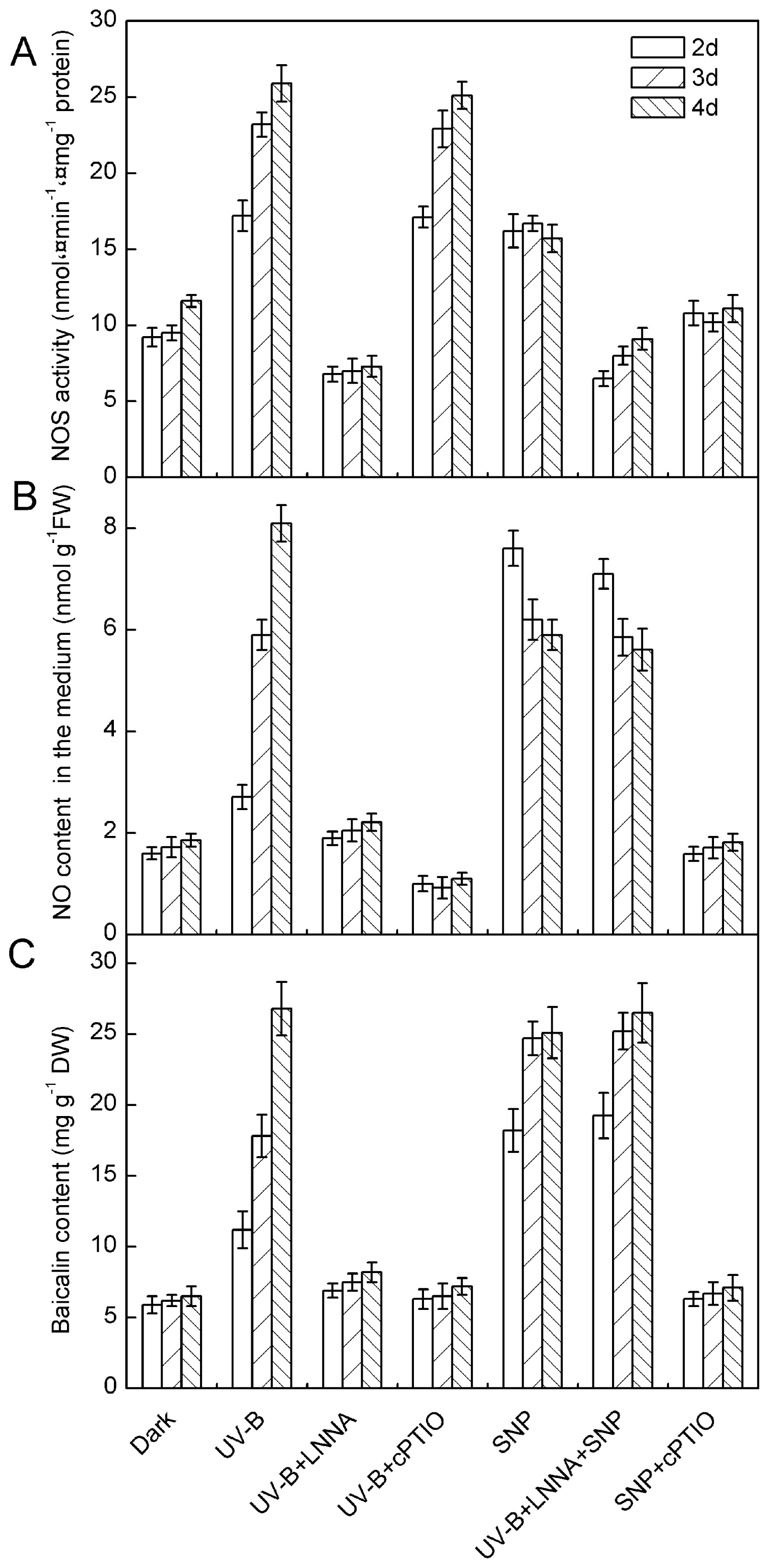
2.1. Results
2.2. Discussion
3. Experimental Section
3.1. Plant Cell Culture
3.2. Experimental Conditions
3.3. Measurement of NO Production
3.4. NOS Activity Assay
3.5. Baicalin Determination
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Lidon, F.C.; Ramalho, J.C. Impact of UV-B irradiation on photosynthetic performance and chloroplast membrane components in Oryza sativa L. J. Photochem. Photobiol. B 2011, 104, 457–466. [Google Scholar]
- Kottuparambila, S.; Shinb, W.; Brownc, M.T.; Han, T. UV-B affects photosynthesis, ROS production and motility of the freshwater flagellate Euglena agilis Carter. Aquat. Toxicol. 2012, 122–123, 206–213. [Google Scholar]
- Frohnmeyer, H.; Staiger, D. Ultraviolet-B radiation-mediated responses in plants Balancing damage and protection. Plant Physiol. 2003, 133, 1420–1428. [Google Scholar]
- Brechner, M.L.; Albright, L.D.; Weston, L.A. Effects of UV-B on secondary metabolites of St John’s Wort (Hypericum perforatum L) grown in controlled environments. Photochem. Photobiol. 2011, 87, 680–684. [Google Scholar]
- Mewis, I.; Schreiner, M.; Nguyen, C.N.; Krumbein, A.; Ulrichs, C.; Marc Lohse, M.; Zrenner, R. UV-B irradiation changes specifically the secondary metabolite profile in broccoli sprouts: induced signaling overlaps with defense response to biotic stressors. Plant Cell Physiol. 2012, 53, 1546–1560. [Google Scholar]
- Grün, S.; Lindermayr, C.; Sell, S.; Durner, J. Nitric oxide and gene regulation in plants. J. Exp. Bot. 2005, 57, 507–516. [Google Scholar]
- Krasylenko, Y.A.; Yemets, A.I.; Blume, Y.B. Functional role of nitric oxide in plants. Russ. J. Plant Physiol. 2010, 57, 451–461. [Google Scholar]
- Fan, H.; Li, T.; Guan, L.; Li, Z.; Guo, N.; Cai, Y.; Lin, Y. Effects of exogenous nitric oxide on antioxidation and DNA methylation of Dendrobium huoshanense grown under drought stress. Plant Cell Tissue Organ Cult. 2012, 109, 307–314. [Google Scholar]
- Petřivalský, M.; Vanickova, P.; Ryzi, M.; Navratilova, B.; Piterkova, J.; Sedlarova, M.; Luhova, L. The effects of reactive nitrogen and oxygen and oxygen species on the regeneration and growth of cucumber cells from isolated protoplasts. Plant Cell Tissue Organ Cult. 2012, 108, 237–249. [Google Scholar]
- Bogatek, R.; Gniazdowska, A. Nitric oxide and HCN reduce deep dormancy of apple seeds. Acta Physiol. Plant. 2006, 28, 281–287. [Google Scholar]
- Hong, J.K.; Yun, B.W.; Kang, J.G.; Raja, M.U.; Kwon, E.; Sorhagen, K.; Chu, C.; Wang, Y.; Loake, G.J. Nitric oxide function and signaling in plant disease resistance. J. Exp. Bot. 2008, 59, 147–154. [Google Scholar]
- Zheng, W.; Miao, K.; Zhang, Y.; Pan, S.; Zhang, M.; Jiang, H. Nitric oxide mediates the fungal-elicitor-enhanced biosynthesis of antioxidant polyphenols in submerged cultures of Inonotus obliquus. Microbiology 2009, 155, 3440–3448. [Google Scholar]
- Sun, J.; Li, L.; Liu, M.; Wang, M.; Ding, M.; Deng, S.; Lu, C.; Zhou, X.; Shen, X.; Chen, S. Hydrogen peroxide and nitric oxide mediate K+/Na+ homeostasis and antioxidant defense in NaCl-stressed callus cells of two contrasting poplars. Plant Cell Tissue Organ Cult. 2010, 103, 205–215. [Google Scholar]
- Simaei, M.; Khavarinejad, R.A.; Saadatmand, S.; Bernard, F.; Fahimi, H. Interactive effects of salicylic acid and nitric oxide on soybean plants under NaCl salinity. Russ. J. Plant Physiol. 2011, 58, 783–790. [Google Scholar]
- Yu, Q.; Sun, L.; Jin, H.; Chen, Q.; Chen, Z.; Xu, M. Lead-induced nitric oxide generation plays a critical role in lead uptake by Pogonatherum crinitum root cells. Plant Cell Physiol. 2012, 53, 1728–1736. [Google Scholar]
- Mayer, B.; Hemmens, B. Biosynthesis and action of nitric oxide in mammalian cells. Trends Biochem. Sci. 1997, 22, 477–481. [Google Scholar]
- Corpas, F.J.; Palma, J.M.; del Río, L.A.; Barroso, J.B. Evidence supporting the existence of l-arginine-dependent nitric oxide synthase activity in plants. New Phytol. 2009, 184, 9–14. [Google Scholar]
- Sekiya, K.; Okuda, H. Selective inhibition of platelet lipoxygenase by baicalin. Biochem. Biophys. Res. Commun. 1982, 105, 1090–1095. [Google Scholar]
- Jung, H.S.; Kim, M.H.; Gwak, N.G.; Im, Y.S.; Lee, K.Y.; Sohn, Y.; Choi, H.; Yang, W.M. Antiallergic effects of Scutellaria baicalensis on inflammation in vivo and in vitro. J. Ethnopharmacol. 2012, 141, 345–349. [Google Scholar]
- Waisundara, V.Y.; Hsu, A.; Huang, D.; Tan, B.K. Scutellaria baicalensis enhances the anti-diabetic activity of metformin in streptozotocin-induced diabetic Wistar rats. Am. J. Chin. Med. 2008, 36, 517–540. [Google Scholar]
- Shen, Y.C.; Chiou, W.F.; Chou, Y.C.; Chen, C.F. Mechanisms in mediating the anti-inflammatory effects of baicalin and baicalein in human leukocytes. Eur. J. Pharmacol 2003, 465, 171–181. [Google Scholar]
- Franek, K.J.; Zhou, Z.; Zhang, W.D.; Chen, W.Y. In vitro studies of baicalin alone or in combination with Salvia miltiorrhiza extract as a potential anti-cancer agent. Int. J. Oncol. 2005, 26, 217–224. [Google Scholar]
- Shieh, D.E.; Liu, L.T.; Lin, C.C. Antioxidant and free radical scavenging effects of baicalein baicalin and wogonin. Anticancer Res. 2000, 20, 2861–2865. [Google Scholar]
- Woo, A.Y.; Cheng, C.H.; Wave, M.M. Baicalein protects rat cardiomyocytes from hypoxia/reoxygenation damage via a prooxidant mechanism. Cardiovasc. Res. 2005, 65, 244–253. [Google Scholar]
- Chang, W.T.; Shao, Z.H.; Yin, J.J.; Mehendale, S.; Wang, C.Z.; Qin, Y.; Li, J.; Chen, W.J.; Chien, C.T.; Becker, L.B.; et al. Comparative effects of flavonoids on oxidant scavenging and ischemia-reperfusion injury in cardiomyocytes. Eur. J. Pharmacol. 2007, 566, 58–66. [Google Scholar]
- Macherness, S.A.H.; John, C.F.; Jordan, B.; Thomas, B. Early signaling components in ultraviolet-B response: distinct roles for different reactive oxygen species and nitric oxide. FEBS Lett. 2001, 489, 237–242. [Google Scholar]
- Qu, Y.; Feng, H.Y.; Wang, Y.B.; Zhang, M.; Cheng, J.; Wang, X.; An, L. Nitric oxide function as a signal in untraviolet-B induced inhibition of pea stems elongation. Plant Sci. 2006, 170, 994–1000. [Google Scholar]
- Hao, G.; Du, X.; Zhao, F.; Shi, R.; Wang, J. Role of nitric oxide in UV-B-induced activation of PAL and stimulation of flavonoid biosynthesis in Ginkgo biloba callus. Plant Cell Tissue Organ Cult. 2009, 97, 175–185. [Google Scholar]
- Zhang, M.; Dong, J.F.; Sun, L.N.; Xu, M.J. Ultraviolet-B-induced flavonoid accumulation in Betula pendula leaves is dependent upon nitrate reductase-mediated nitric oxide signaling. Tree Physiol. 2011, 31, 798–807. [Google Scholar]
- Zhang, B.; Zheng, L.P.; Wang, J.W. Nitric oxide elicitation for secondary metabolite production in cultured plant cells. Appl. Microbiol. Biotechnol. 2012, 93, 455–466. [Google Scholar]
- Edreva, A.; Velikova, V.; Tsonev, V.; Dagnon, S.; Gürel, A.L.; Aktaş, L.; Gesheva, E. Stress-protective role of secondary metabolities: Diversity of functions and mechanisms. Gen. Appl. Plant Physiol. 2008, 34, 67–78. [Google Scholar]
- Gyula, N.; Schafer, E.; Nagy, F. Light perception and signalling in higher plants. Curr. Opin. Plant Biol. 2003, 6, 446–452. [Google Scholar]
- Christie, J.M. Phototropin blue-light receptors. Annu. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar]
- Rizzini, L.; Favory, J.J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schäfer, E.; Nagy, F.; Jenkins, G.I.; et al. Perception of UV-B by the Arabidopsis UVR8 Protein. Science 2011, 332, 103–106. [Google Scholar]
- Logemann, E.; Tavernaro, A.; Schulz, W.; Somssich, I.E.; Hahlbrock, K. UV light selectively coinduces supply pathways from primary metabolism and flavonoid secondary product fo parsley. Proc. Natl. Acad. Sci. USA 2000, 97, 1903–1907. [Google Scholar]
- Ohl, S.; Hahlbrock, K.; Schäfer, E. A stable blue-light-derived signal modulates ultraviolet-light-induced activation of the chalcone-synthase gene in cultured parsley cells. Planta 1989, 177, 228–236. [Google Scholar]
- Tossi, V.; Lombardo, C.; Cassia, R.; Lamattina, L. Nitric oxide and flavonoids are systemically induced by UV-B in maize leaves. Plant Sci. 2012, 193–194, 103–109. [Google Scholar]
- Jenkins, G.I. Signal transduction in responses to UV-B radiation. Annu. Rev. Plant Biol. 2009, 60, 407–431. [Google Scholar]
- Liu, Z.; Liu, Y.; Pu, Z.; Wang, J.; Zheng, Y.; Li, Y.; Wei, Y. Regulation evolution and functionality of flavonoids in cereal crops. Biotechnol. Lett. 2013, 35, 1765–1780. [Google Scholar]
- Liu, P.-F.; Han, F.-G.; Duan, B.-B.; Deng, T.-S.; Hou, X.-L.; Zhao, M.-Q. Purification and antioxidant activities of baicalin isolated from the root of huangqin (Scutellaria baicalensis gcorsi). J. Food Sci. Technol. 2013, 50, 615–619. [Google Scholar]
- Wendehenne, D.; Pugin, A.; Klessig, D.F.; Durner, J. Nitric oxide: Comparative synthesis and signaling in animal and plant cells. Trends Plant Sci. 2001, 6, 177–183. [Google Scholar]
- Xu, M.J.; Dong, J.F.; Zhu, M.Y. Involvement of NO in fungal elicitor-induced activation of PAL and stimulation of taxol biosynthesis in Taxus chinensis suspension cells. Chin. Sci. Bull. 2004, 49, 1038–1043. [Google Scholar]
- Corpas, F.J.; Carreras, A.; Valderrama, R.; Palma, J.M.; León, A.M.; Sandalio, L.M.; del Río, L.A. Constitutive arginine-dependent nitric oxide synthase activity in different organs of pea seedlings during plant development. Planta 2006, 224, 246–254. [Google Scholar]
- Zhao, M.G.; Tian, Q.Y.; Zhang, W.H. Nitric oxide synthase-dependent nitric oxide production is associated with salt tolerance in Arabidopsis. Plant Physiol. 2007, 14, 206–217. [Google Scholar]
- Guo, F.Q.; Okamoto, M.; Crawford, N.M. Identification of a plant nitric oxide synthase gene involved in hormonal signaling. Science 2003, 302, 100–103. [Google Scholar]
- Ribeiro, E.A.; Cunha, F.Q.; Tamashiro, W.M.S.C.; Martins, I.S. Growth phase-dependent subcellular localization of nitric oxide synthase in maize cells. FEBS Lett. 1999, 445, 283–286. [Google Scholar]
- Barroso, J.B.; Cprpas, F.J.; Carreras, A.; Sandalio, L.M.; Valderrama, R.; Palma, J.M.; Lupiáñez, J.A.; del Río, L.A. Localization of nitric-oxide synthase in plant peroxisomes. J. Biol. Chem. 1999, 274, 36729–36733. [Google Scholar]
- Del Río, L.A.; Corpas, F.J.; Barroso, J.B. Nitric oxide and nitric oxide synthase activity in plants. Phytochemistry 2004, 65, 783–792. [Google Scholar]
- Corpas, F.J.; Barroso, J.B.; Carreras, A.; Quirós, M.; León, A.M.; Romero-Puertas, M.C.; Esteban, F.J.; Valderrama, R.; Palma, J.M.; Sandalio, L.M.; et al. Cellular and subcellular localization of endogenous nitric oxide in young and senescent pea plants. Plant Physiol. 2004, 136, 2722–2733. [Google Scholar]
- Foresi, N.; Correa-Aragunde, N.; Parisi, G.; Caló, G.; Salerno, G.; Lamattina, L. Characterization of a nitric oxide synthase from the plant kingdom: NO generation from the green alga Ostreococcus tauri is light irradiance and growth phase dependent. Plant Cell 2010, 22, 3816–3830. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Physiol. Plant 1962, 15, 473–497. [Google Scholar]
- Koti, S.; Reddy, K.R.; Reddy, V.R.; Kakani, V.G.; Zhao, D. Interactive effects of carbon dioxide temperature and ultraviolet-B radiation on soybean (Glycine max L) flower and pollen morphology pollen production germination and tube lengths. J. Exp. Bot. 2005, 56, 725–736. [Google Scholar]
- Caldwell, M.M. Solar UV-B irradiation and the growth and development of higher plant. In Photophysiology; Giese, A.C., Ed.; Academic Press: New York, NY, USA, 1971; Volume 6, pp. 131–171. [Google Scholar]
- Zhang, J.J.; Xu, M.J. Effects of nitric oxide and Methyl Jasmonate on the baicalin production and cell growth in suspension cultures of Scutellaria baicalensis. Chin. Bull. Bot. 2006, 23, 374–379. [Google Scholar]
- De Bisschop, C.; Kiger, L.; Marden, M.C.; Ajata, A.; Huez, S.; Faoro, V.; Martinot, J.B.; Naeije, R.; Guénard, H. Pulmonary capillary blood volume and membrane conductance in Andeans and lowlanders at high altitude: A cross-sectional study. Nitric Oxide 2010, 23, 187–193. [Google Scholar]
- Kojima, H.; Nagano, T. Fluorescent indicators for nitric oxide. Adv. Mater. 2000, 12, 763–765. [Google Scholar]
- Ninnemann, H.; Maier, J. Indication for occurrence of nitric oxide synthases in fungi and plants and involvement in photoconidiation of Neurospora crassa. Photochem. Photobiol. 1996, 64, 393–398. [Google Scholar]
- Zhang, J.J.; Xu, M.J.; Yu, J. Determination of baicalin content in suspension culture of Scutellaria baicalensis Georgi cells. West China J. Pharm. Sci. 2006, 21, 575–576. [Google Scholar]







© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, J.-J.; Li, X.-Q.; Sun, J.-W.; Jin, S.-H. Nitric Oxide Functions as a Signal in Ultraviolet-B-Induced Baicalin Accumulation in Scutellaria baicalensis Suspension Cultures. Int. J. Mol. Sci. 2014, 15, 4733-4746. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms15034733
Zhang J-J, Li X-Q, Sun J-W, Jin S-H. Nitric Oxide Functions as a Signal in Ultraviolet-B-Induced Baicalin Accumulation in Scutellaria baicalensis Suspension Cultures. International Journal of Molecular Sciences. 2014; 15(3):4733-4746. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms15034733
Chicago/Turabian StyleZhang, Jin-Jie, Xue-Qin Li, Jun-Wei Sun, and Song-Heng Jin. 2014. "Nitric Oxide Functions as a Signal in Ultraviolet-B-Induced Baicalin Accumulation in Scutellaria baicalensis Suspension Cultures" International Journal of Molecular Sciences 15, no. 3: 4733-4746. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms15034733