Disruption of Phosphoinositide-Specific Phospholipases Cγ1 Contributes to Extracellular Matrix Synthesis of Human Osteoarthritis Chondrocytes

Abstract
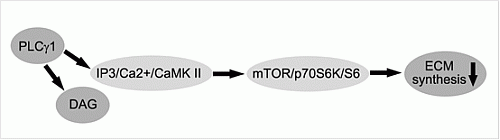
:
1. Introduction
2. Results
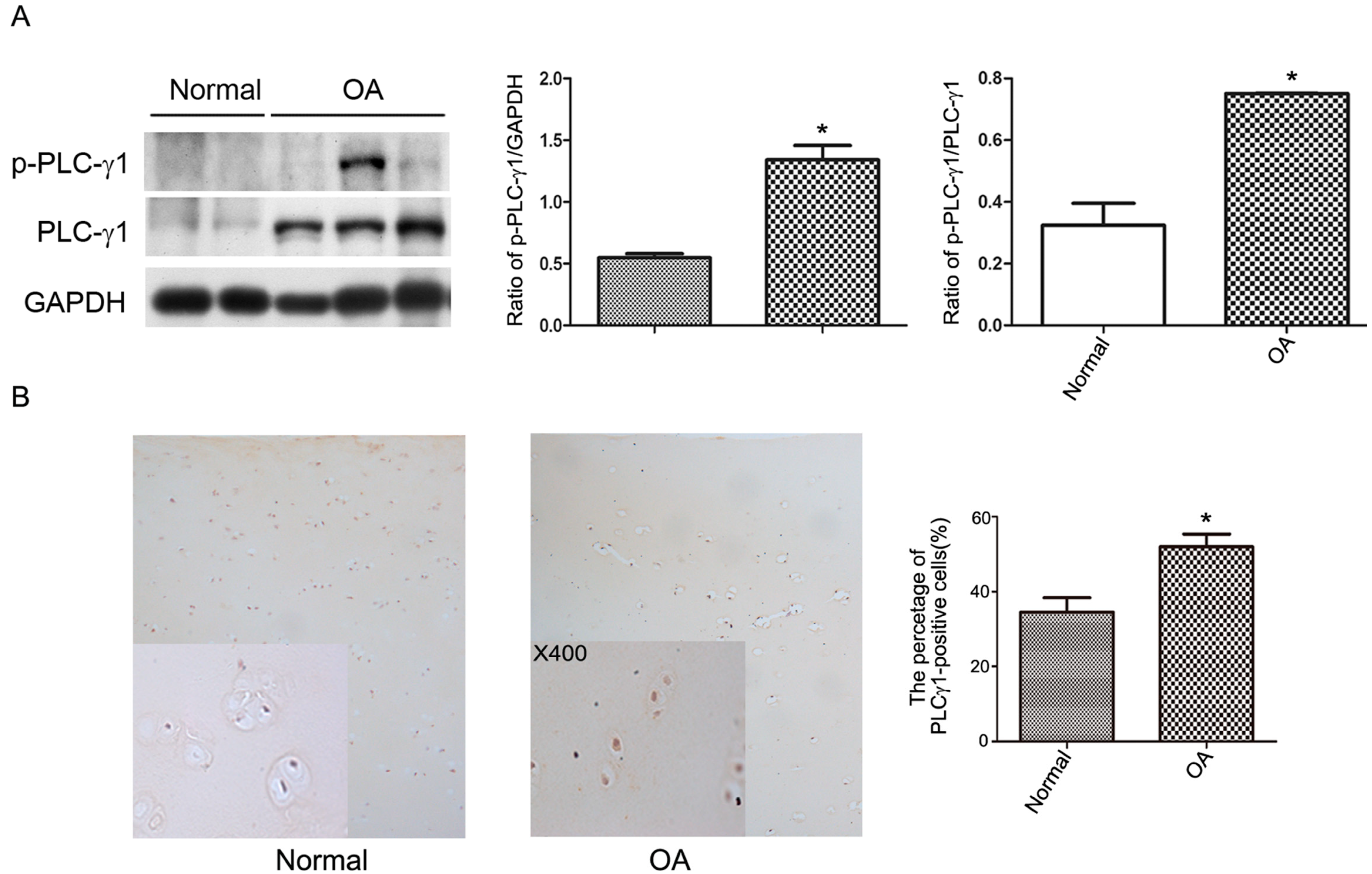
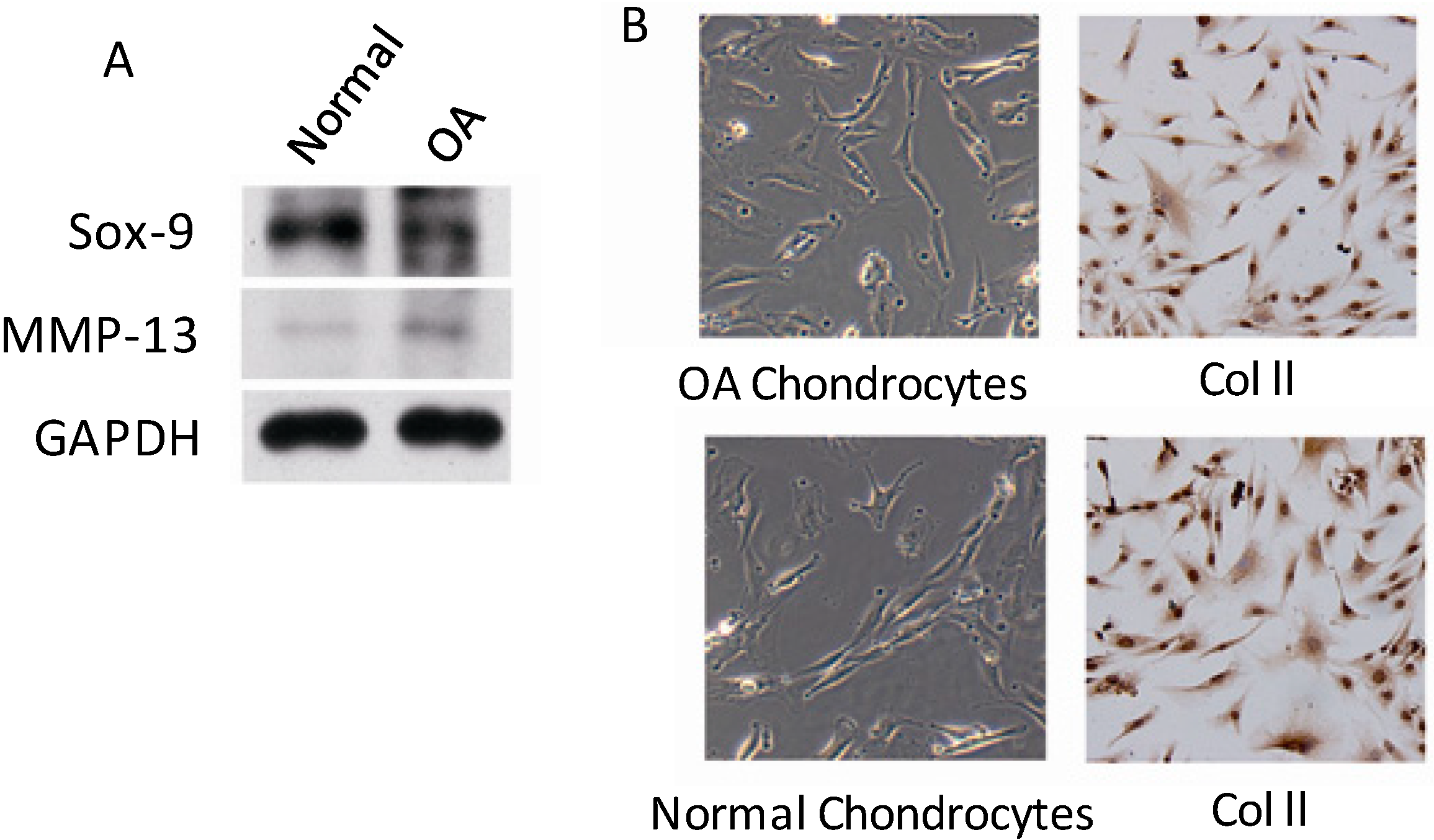
2.1. PLC-γ1 Is Highly Expressed in Human OA Chondrocytes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Year) | Case | Sex | Duration of OA (Year) | * K.L.Image Criterion | Pro-Treatment Arthroscopy | |||
|---|---|---|---|---|---|---|---|---|
| M | F | ≤3 | >3 | III | IV | |||
| 60- | 11 | 1 | 10 | 5 | 6 | 2 | 9 | 5 |
| 65- | 15 | 5 | 10 | 4 | 11 | 4 | 11 | 2 |

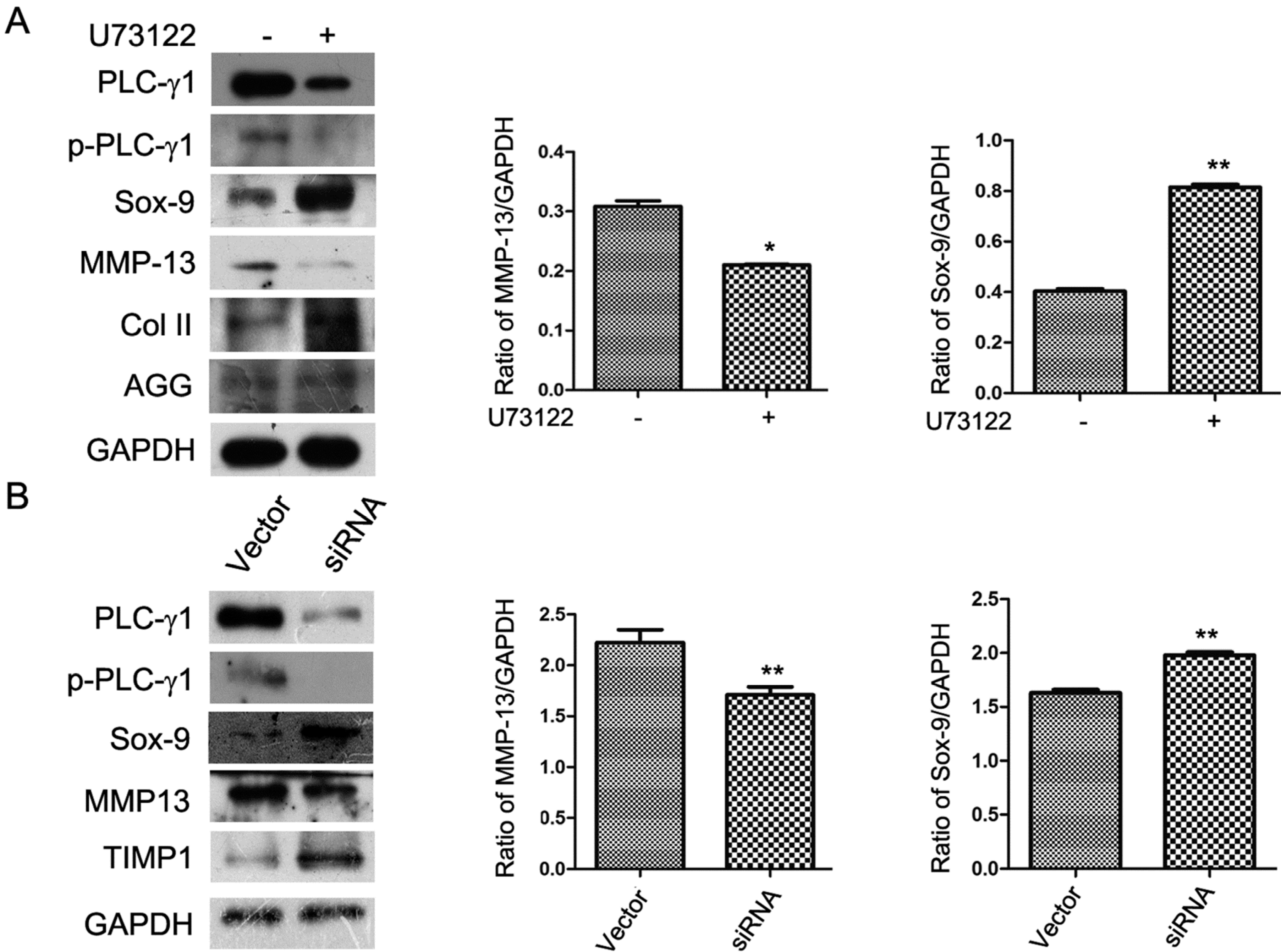
2.2. The ECM Synthesis of Human OA Chondrocytes Partly Depends on PLC-γ1

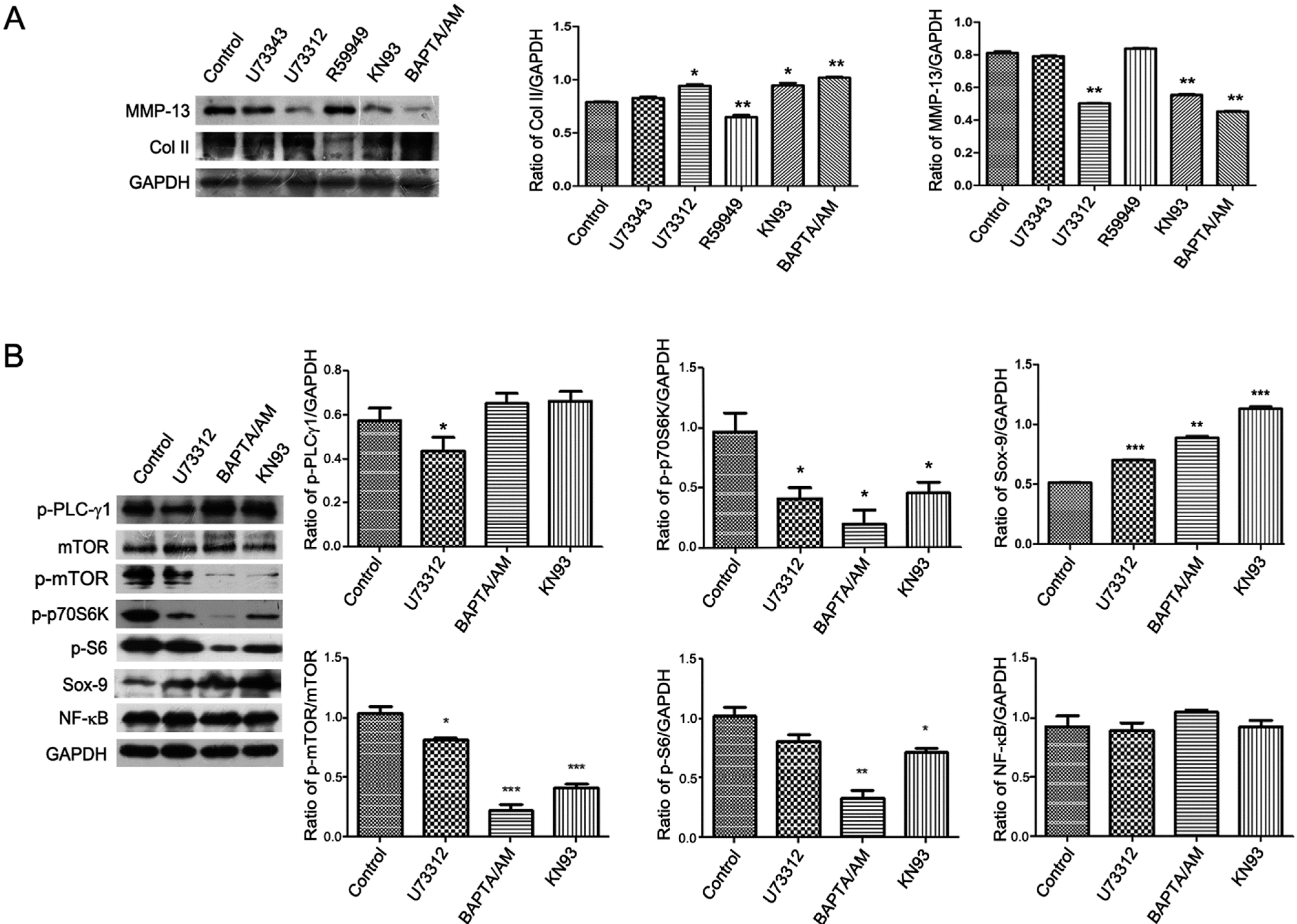
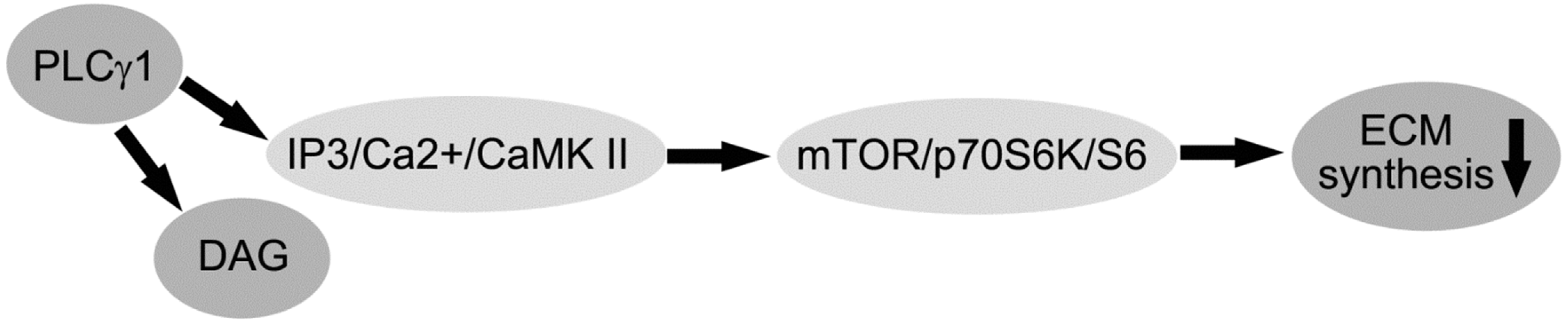
2.3. PLC-γ1/IP3/Ca(2+)/CaMK II Axis Regulates the ECM Synthesis of Human OA Chondrocytes

3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Human Normal and OA Chondrocyte Isolation and Culture
4.3. Immunohistochemistry Technique
4.4. Protein Extraction and Western Blotting Analysis
4.5. Statistical Analysis
5. Conclusions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maldonado, M.; Nam, J. The role of changes in extracellular matrix of cartilage in the presence of inflammation on the pathology of osteoarthritis. Biomed. Res. Int. 2013. [Google Scholar] [CrossRef]
- Jackson, M.T.; Smith, M.M.; Smith, S.M.; Jackson, C.J.; Xue, M.; Little, C.B. Activation of cartilage matrix metalloproteinases by activated protein C. Arthritis Rheumatol. 2009, 60, 780–791. [Google Scholar] [CrossRef]
- Ahmad, R.; Sylvester, J.; Ahmad, M.; Zafarullah, M. Adaptor proteins and Ras synergistically regulate IL-1-induced ADAMTS-4 expression in human chondrocytes. J. Immunol. 2009, 182, 5081–5087. [Google Scholar] [CrossRef]
- Rhee, S.G. Regulation of phosphoinositide-specific phospholipase C. Annu. Rev. Biochem. 2001, 70, 281–312. [Google Scholar] [CrossRef]
- Tvorogov, D.; Wang, X.J.; Zent, R.; Carpenter, G. Integrin-dependent PLC-gamma1 phosphorylation mediates fibronectin-dependent adhesion. J. Cell Sci. 2005, 118, 601–610. [Google Scholar] [CrossRef]
- Timsah, Z.; Ahmed, Z.; Lin, C.C.; Melo, F.A.; Stagg, L.J.; Leonard, P.G.; Jeyabal, P.; Berrout, J.; O’Neil, R.G.; Bogdanov, M.; et al. Competition between Grb2 and Plcγ1 for FGFR2 regulates basal phospholipase activity and invasion. Nat. Struct. Mol. Biol. 2014, 21, 180–188. [Google Scholar]
- Burgdorf, C.; Schäfer, U.; Richardt, G.; Kurz, T. U73122, an aminosteroid phospholipase C inhibitor, is a potent inhibitor of cardiac phospholipase D by a PIP2-dependent mechanism. J. Cardiovasc. Pharmacol. 2010, 55, 555–559. [Google Scholar] [CrossRef]
- Harada, D.; Yamanaka, Y.; Ueda, K.; Nishimura, R.; Morishima, T.; Seino, Y.; Tanaka, H. Sustained phosphorylation of mutated FGFR3 is a crucial feature of genetic dwarfism and induces apoptosis in the ATDC5 chondrogenic cell line via PLCgamma-activated STAT1. Bone 2007, 41, 273–281. [Google Scholar] [CrossRef]
- Biswas, S.; Manikandan, J.; Pushparaj, P.N. Decoding the differential biomarkers of Rheumatoid arthritis and Osteoarthritis: A functional genomics paradigm to design disease specific therapeutics. Bioinformation 2011, 6, 153–157. [Google Scholar] [CrossRef]
- Pearle, A.D.; Warren, R.F.; Rodeo, S.A. Basic science of articular cartilage and osteoarthritis. Clin. Sports Med. 2005, 24, 1–12. [Google Scholar] [CrossRef]
- Shlopov, B.V.; Lie, W.R.; Mainardi, C.L.; Cole, A.A.; Chubinskaya, S.; Hasty, K.A. Osteoarthritic lesions: involvement of three different collagenases. Arthritis Rheumatol. 1997, 40, 2065–2074. [Google Scholar] [CrossRef]
- Nam, J.; Perera, P.; Liu, J.; Rath, B.; Deschner, J.; Gassner, R.; Butterfield, T.A.; Agarwal, S. Sequential alterations in catabolic and anabolic gene expression parallel pathological changes during progression of monoiodoacetate-induced arthritis. PLoS One 2011, 6, e24320. [Google Scholar]
- Lee, J.S.; Im, G.I. SOX trio decrease in the articular cartilage with the advancement of osteoarthritis. Connect. Tissue Res. 2011, 52, 496–502. [Google Scholar] [CrossRef]
- Wang, Q.; Siminovitch, K.A.; Downey, G.P.; McCulloch, C.A. Ras-guanine-nucleotide-releasing factors 1 and 2 interact with PLCγ at focal adhesions to enable IL-1-induced Ca(2+) signalling, ERK activation and MMP-3 expression. Biochem. J. 2013, 449, 771–782. [Google Scholar] [CrossRef]
- Kanaki, T.; Bujo, H.; Mori, S.; Yanjuan, Z.; Takahashi, K.; Yokote, K.; Morisaki, N.; Saito, Y. Functional analysis of aortic endothelial cells expressing mutant PDGF receptors with respect to expression of matrix metalloproteinase-3. Biochem. Biophys. Res. Commun. 2002, 294, 231–237. [Google Scholar]
- Yuan, F.L.; Chen, F.H.; Lu, W.G.; Li, X.; Li, J.P.; Li, C.W.; Xu, R.S.; Wu, F.R.; Hu, W.; Zhang, T.Y.; et al. Inhibition of acid-sensing ion channels in articular chondrocytes by amiloride attenuates articular cartilage destruction in rats with adjuvant arthritis. Inflamm. Res. 2010, 59, 939–947. [Google Scholar] [CrossRef]
- Nguyen, C.; Lieberherr, M.; Bordat, C.; Velard, F.; Côme, D.; Lioté, F.; Ea, H.K. Intracellular calcium oscillations in articular chondrocytes induced by basic calcium phosphate crystals lead to cartilage degradation. Osteoarthr. Cartil. 2012, 20, 1399–1408. [Google Scholar] [CrossRef]
- Markova, B.; Albers, C.; Breitenbuecher, F.; Melo, J.V.; Brümmendorf, T.H.; Heidel, F.; Lipka, D.; Duyster, J.; Huber, C.; Fischer, T. Novel pathway in Bcr-Abl signal transduction involves Akt-independent, PLC-gamma1-driven activation of mTOR/p70S6-kinase pathway. Oncogene 2010, 29, 739–751. [Google Scholar] [CrossRef]
- Kharas, M.G.; Fruman, D.A. ABL oncogenes and phosphoinositide 3-kinase: Mechanism of activation and downstream effectors. Cancer Res. 2005, 65, 2047–2053. [Google Scholar] [CrossRef]
- Edinger, A.L.; Thompson, C.B. Akt maintains cell size and survival by increasing mTOR-dependent nutrient uptake. Mol. Biol. Cell. 2002, 13, 2276–2288. [Google Scholar] [CrossRef]
- Razmara, M.; Heldin, C.H.; Lennartsson, J. Platelet-derived growth factor-induced Akt phosphorylation requires mTOR/Rictor and phospholipase C-γ1, whereas S6 phosphorylation depends on mTOR/Raptor and phospholipase D. Cell. Commun. Signal. 2013, 11. [Google Scholar] [CrossRef]
- Mamane, Y.; Petroulakis, E.; LeBacquer, O.; Sonenberg, N. mTOR, translation initiation and cancer. Oncogene 2006, 25, 6416–6422. [Google Scholar] [CrossRef]
- Zhang, T.; Lu, X.; Li, J.; Chidiac, P.; Sims, S.M.; Feng, Q. Inhibition of Na/K-ATPase promotes myocardial tumor necrosis factor-alpha protein expression and cardiac dysfunction via calcium/mTOR signaling in endotoxemia. Basic Res. Cardiol. 2012, 107, 1435–1803. [Google Scholar]
- Attur, M.G.; Dave, M.; Cipolletta, C.; Kang, P.; Goldring, M.B.; Patel, I.R.; Abramson, S.B.; Amin, A.R. Reversal of autocrine and paracrine effects of interleukin 1 (IL-1) in human arthritis by type II IL-1 decoy receptor. Potential for pharmacological intervention. J. Biol. Chem. 2000, 275, 40307–40315. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, B.; Yi, T.; Xia, C. Increased apoptosis in human knee osteoarthritis cartilage related to the expression of protein kinase B and protein kinase Cɑ in chondrocytes. Folia Histochem. Cytobiol. 2012, 50, 137–143. [Google Scholar] [CrossRef]
- Huang, J.G.; Xia, C.; Zheng, X.P.; Yi, T.T.; Wang, X.Y.; Song, G.; Zhang, B. 17β-Estradiol promotes cell proliferation in rat osteoarthritis model chondrocytes via PI3K/Akt pathway. Cell. Mol. Biol. Lett. 2011, 16, 564–575. [Google Scholar] [CrossRef]
Appendix

© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zeng, G.; Cui, X.; Liu, Z.; Zhao, H.; Zheng, X.; Zhang, B.; Xia, C. Disruption of Phosphoinositide-Specific Phospholipases Cγ1 Contributes to Extracellular Matrix Synthesis of Human Osteoarthritis Chondrocytes. Int. J. Mol. Sci. 2014, 15, 13236-13246. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms150813236
Zeng G, Cui X, Liu Z, Zhao H, Zheng X, Zhang B, Xia C. Disruption of Phosphoinositide-Specific Phospholipases Cγ1 Contributes to Extracellular Matrix Synthesis of Human Osteoarthritis Chondrocytes. International Journal of Molecular Sciences. 2014; 15(8):13236-13246. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms150813236
Chicago/Turabian StyleZeng, Guoqing, Xu Cui, Zejun Liu, Honghai Zhao, Xinpeng Zheng, Bing Zhang, and Chun Xia. 2014. "Disruption of Phosphoinositide-Specific Phospholipases Cγ1 Contributes to Extracellular Matrix Synthesis of Human Osteoarthritis Chondrocytes" International Journal of Molecular Sciences 15, no. 8: 13236-13246. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms150813236