Modulation of Nitrosative Stress via Glutathione-Dependent Formaldehyde Dehydrogenase and S-Nitrosoglutathione Reductase

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Subcloning and Construction of Codon Optimized Tc-Glutathione-Dependent Formaldehyde Dehydrogenase (Tc-GFD-O)

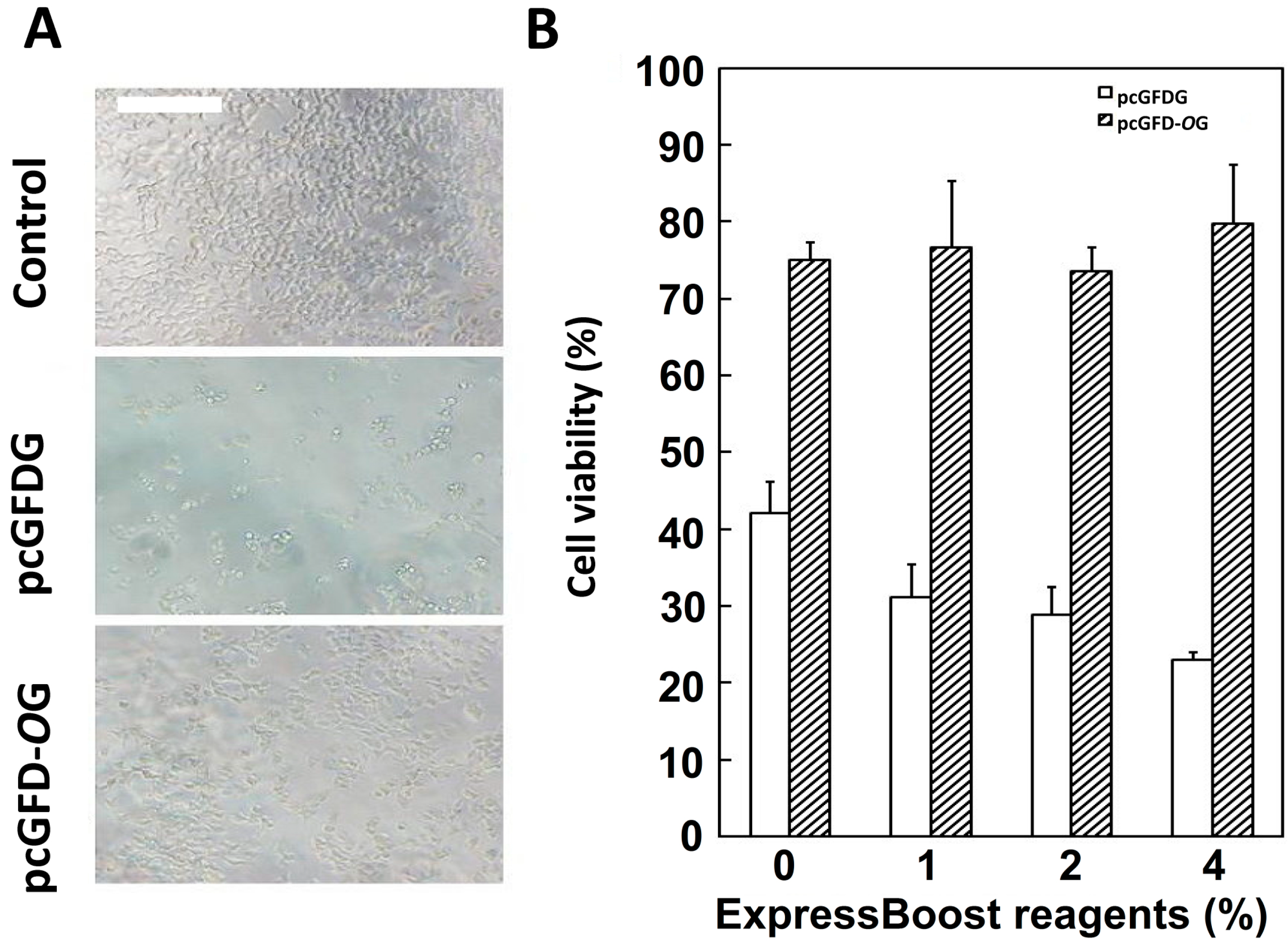
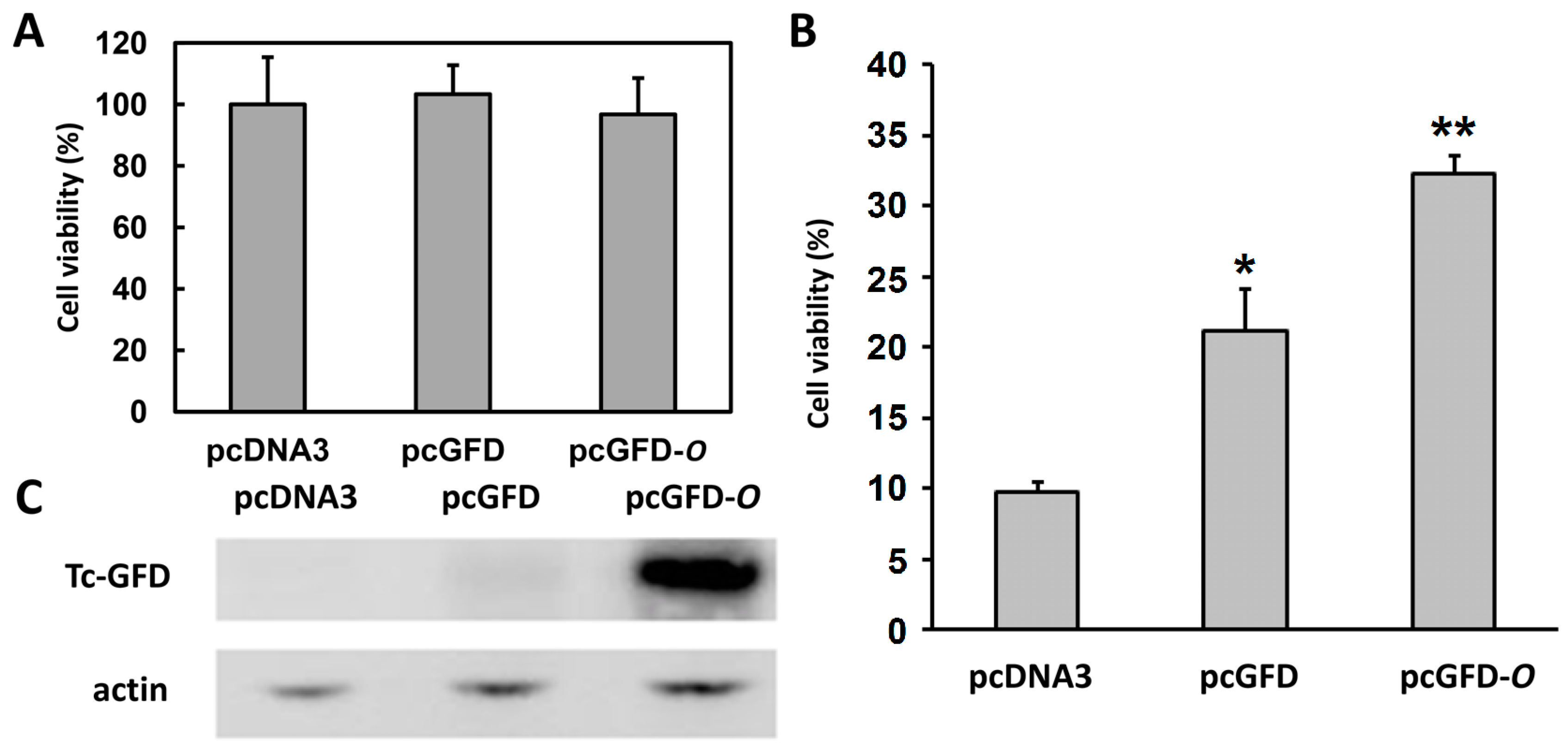
2.2. Improved Viability of 293T Cells Transfected with Codon Optimized Tc-GFD-OG

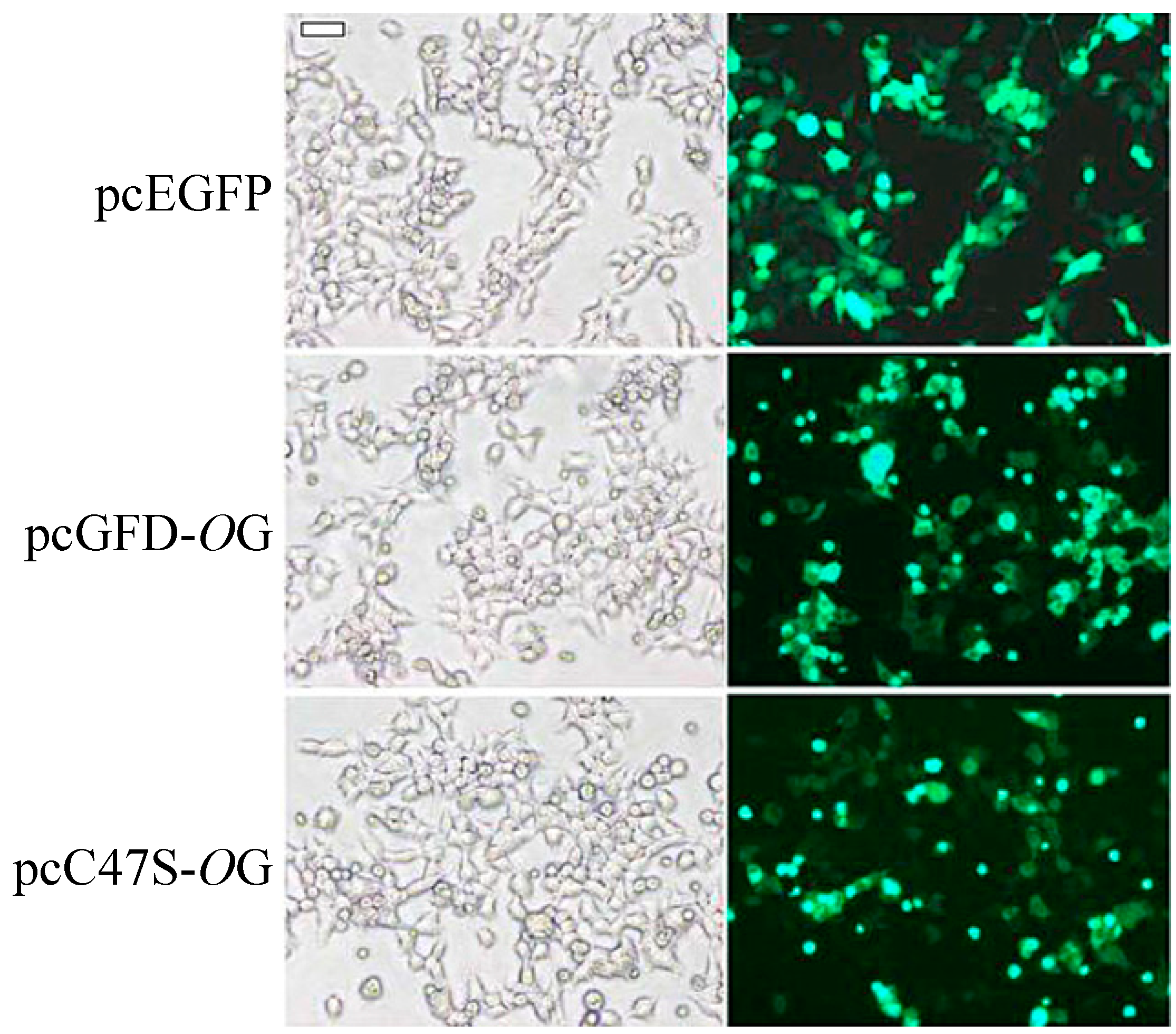
2.3. Transfection and Expression of Green Fluorescent Protein (GFP)-Fused Tc-GFD Protein and C47S Mutant in 293T

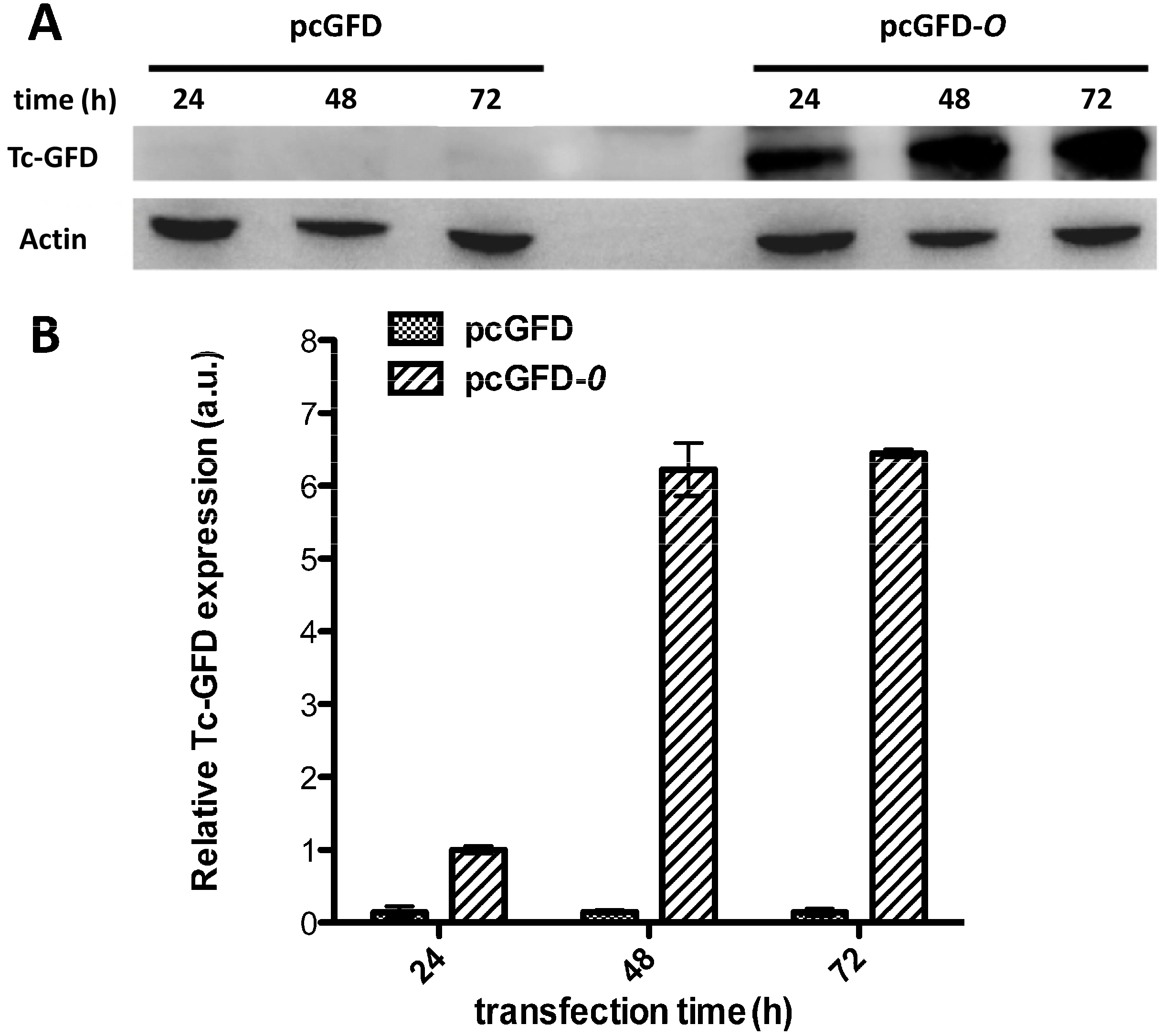
2.4. Expression of the Recombinant Tc-GFD-O in 293T Cells
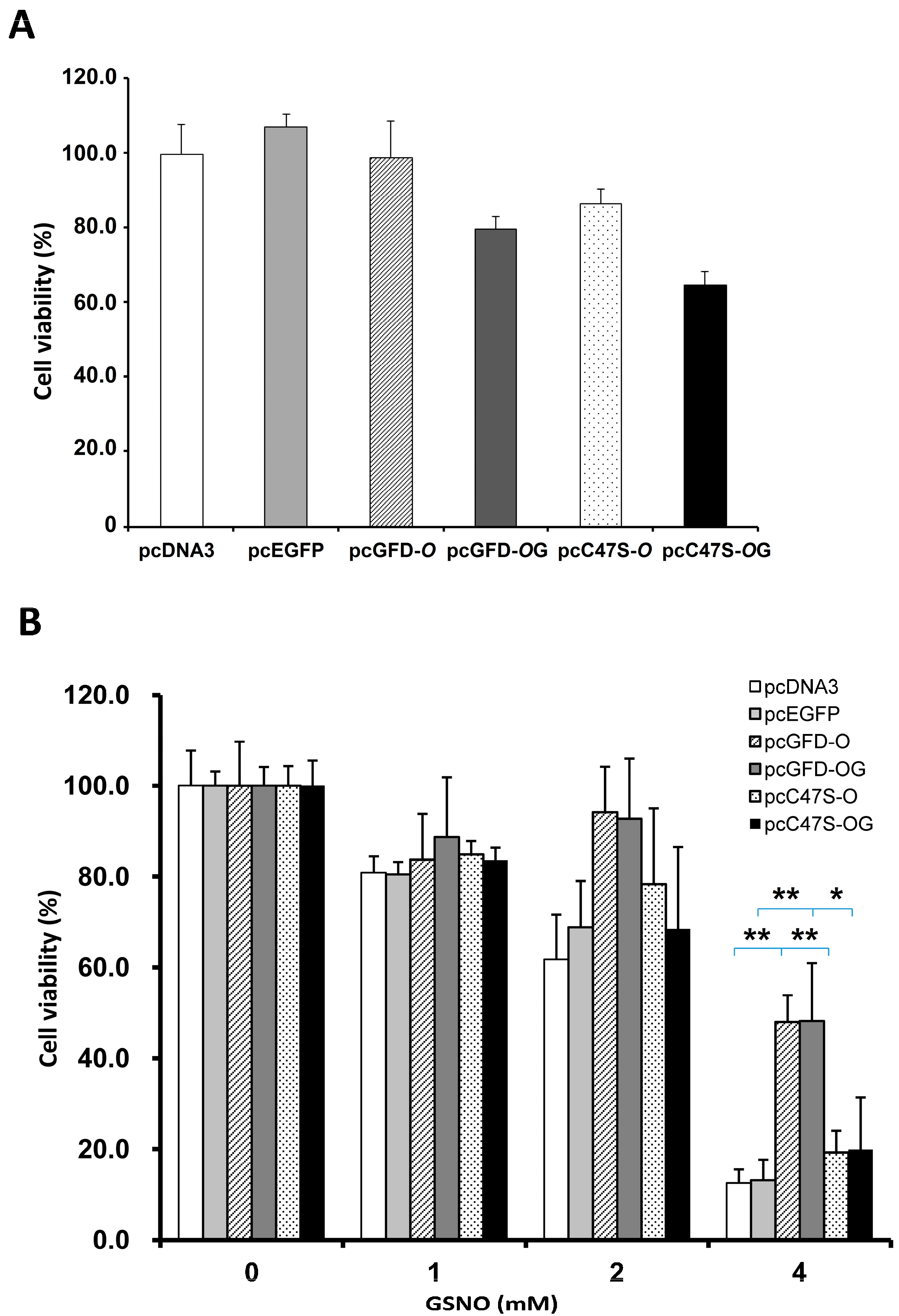
2.5. Effect of Tc-GFD Protein on S-Nitrosoglutathione (GSNO)-Induced 293T Cell Viability


2.6. Effect of Mutation at C47S of GFD-O on GSNO-Induced 293T Cell Viability

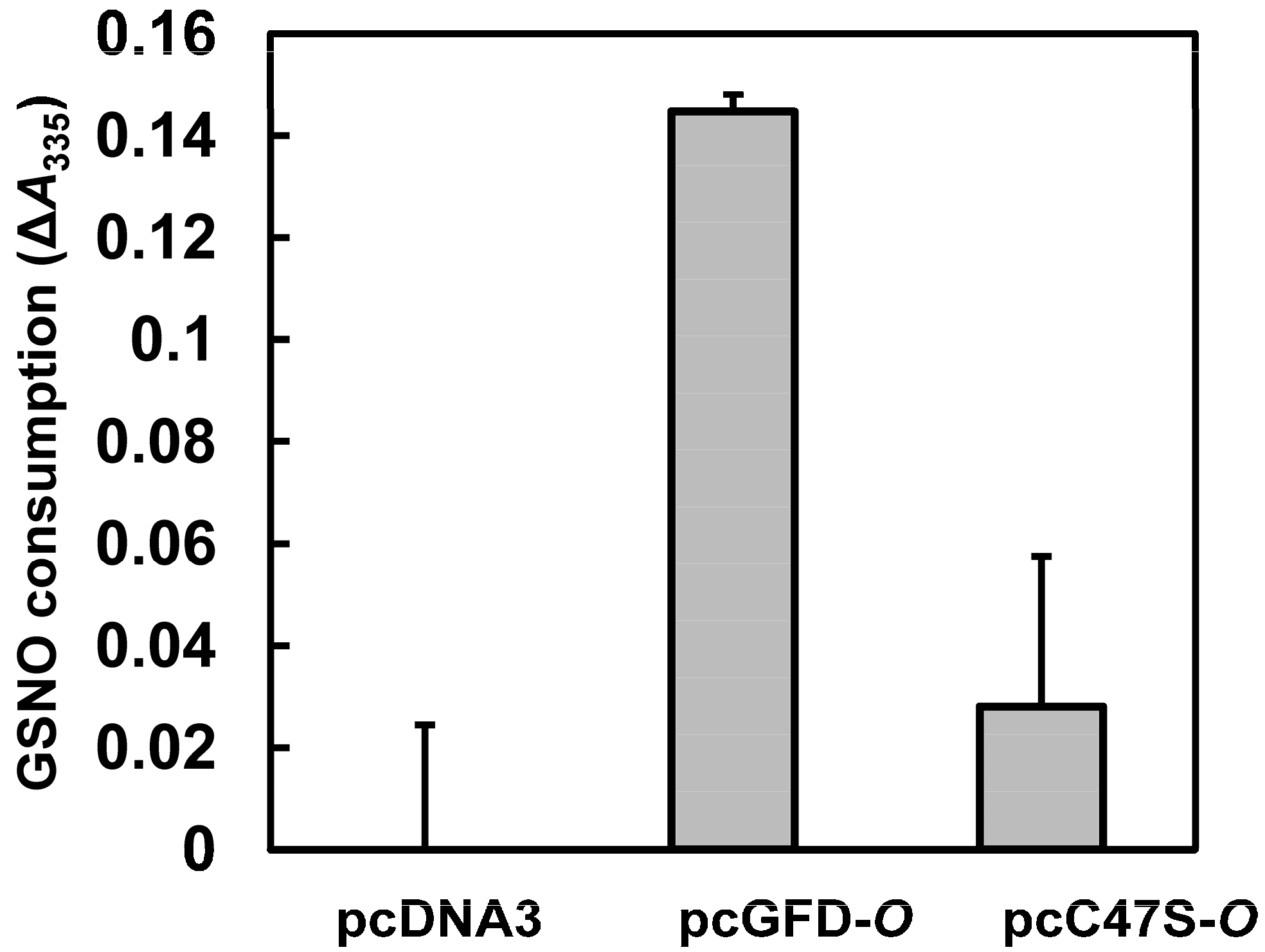
2.7. Effect of Tc-GFD Protein on GSNO Consumption in Transfected 293T Cells

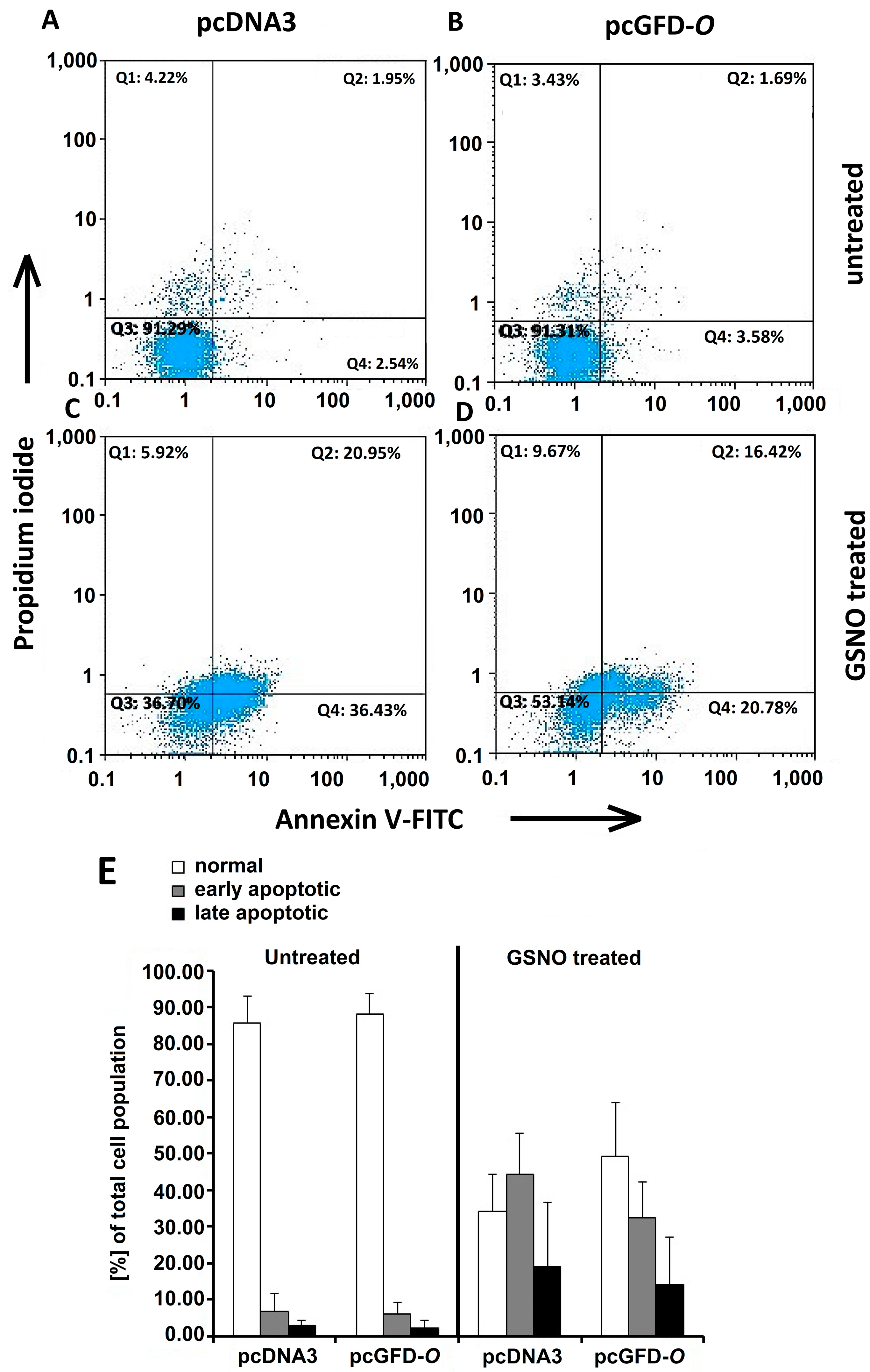
2.8. Effect of GFD on GSNO-Induced Apoptosis in 293T Cells

3. Materials and Methods
3.1. Subcloning of Wild-Type Tc-GFD and Codon-Optimized GFD cDNA (Tc-GFD-O) into Mammalian Expression Vectors
3.2. Site-Directed Mutagenesis of Cys 47 to Ser in Tc-GFD-O Subclones
3.3. Cell Culture and Transfection of 293T Cells with Tc-GFD-O
3.4. Cell Viability Assay
3.5. Expression of Tc-GFD Protein in 293T Cells
3.6. Induction of Oxidative/Nitrosative Stress in Parental and Transfected 293T Cells by S-Nitrosoglutathione Treatment
3.7. Transfection Efficiency of GFD in 293T Cells
3.8. Effect of Tc-GFD Protein on GSNO-Induced 293T Cell Viability
3.9. Effect of pcGFD-O or pcC47S-O on GSNO Consumption
3.10. Effect of GFD on GSNO-Induced 293T Cell Apoptosis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Andersen, J.K. Oxidative stress in neurodegeneration: Cause or consequence? Nat. Med. 2004, 10, S18–S25. [Google Scholar] [CrossRef]
- Barnham, K.J.; Masters, C.L.; Bush, A.I. Neurodegenerative diseases and oxidative stress. Nat. Rev. Drug Discov. 2004, 3, 205–214. [Google Scholar] [CrossRef]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef]
- Banati, R.B.; Gehrmann, J.; Schubert, P.; Kreutzberg, G.W. Cytotoxicity of microglia. Glia 1993, 7, 111–118. [Google Scholar] [CrossRef]
- Sano, M.; Ernesto, C.; Thomas, R.G.; Klauber, M.R.; Schafer, K.; Grundman, M.; Woodbury, P.; Growdon, J.; Cotman, C.W.; Pfeiffer, E.; et al. A controlled trial of selegiline, α-tocopherol, or both as treatment for Alzheimer’s disease. The Alzheimer’s disease cooperative study. N. Engl. J. Med. 1997, 336, 1216–1222. [Google Scholar] [CrossRef]
- Prasad, K.N.; Hovland, A.R.; Cole, W.C.; Prasad, K.C.; Nahreini, P.; Edwards-Prasad, J.; Andreatta, C.P. Multiple antioxidants in the prevention and treatment of Alzheimer disease: Analysis of biologic rationale. Clin. Neuropharmacol. 2000, 23, 2–13. [Google Scholar] [CrossRef]
- Barber, R.D.; Donohue, T.J. Pathways for transcriptional activation of a glutathione-dependent formaldehyde dehdrogenase gene. J. Mol. Biol. 1998, 280, 775–784. [Google Scholar] [CrossRef]
- Jones, D.P.; Thor, H.; Andersson, B.; Orrenius, S. Detoxification reactions in isolated hepatocytes. Role of glutathione peroxidase, catalase, and formaldehyde dehydrogenase in reactions relating to N-demethylation by the cytochrome P-450 system. J. Biol. Chem. 1978, 253, 6031–6037. [Google Scholar]
- Goenrich, M.; Bartoschek, S.; Hagemeier, C.H.; Griesinger, C.; Vorholt, J.A. A glutathione-dependent formaldehyde-activating enzyme (Gfa) from Paracoccus denitrificans detected and purified via two-dimensional proton exchange NMR spectroscopy. J. Biol. Chem. 2002, 277, 3069–3072. [Google Scholar]
- Mason, R.P.; Sanders, J.K.; Crawford, A.; Hunter, B.K. Formaldehyde metabolism by Escherichia coli. Detection by in vivo 13C NMR spectroscopy of S-(hydroxymethyl)glutathione as a transient intracellular intermediate. Biochemistry 1986, 25, 4504–4507. [Google Scholar] [CrossRef]
- Hoog, J.O.; Hedberg, J.J.; Stromberg, P.; Svensson, S. Mammalian alcohol dehydrogenase-functional and structural implications. J. Biomed. Sci. 2001, 8, 71–76. [Google Scholar]
- Harms, N.; Ras, J.; Reijnders, W.N.; van Spanning, R.J.; Stouthamer, A.H. S-Formylglutathione hydrolase of Paracoccus denitrificans is homologous to human esterase D: A universal pathway for formaldehyde detoxification? J. Bacteriol. 1996, 178, 6296–6299. [Google Scholar]
- Liu, L.; Hausladen, A.; Zeng, M.; Que, L.; Heitman, J.; Stamler, J.S. A metabolic enzyme for S-nitrosothiol conserved from bacteria to humans. Nature 2001, 410, 490–494. [Google Scholar] [CrossRef]
- Liu, L.; Yan, Y.; Zeng, M.; Zhang, J.; Hanes, M.A.; Ahearn, G.; McMahon, T.J.; Dickfeld, T.; Marshall, H.E.; Que, L.G.; et al. Essential roles of S-nitrosothiols in vascular homeostasis and endotoxic shock. Cell 2004, 116, 617–628. [Google Scholar] [CrossRef]
- Hess, D.T.; Matsumoto, A.; Kim, S.O.; Marshall, H.E.; Stamler, J.S. Protein S-nitrosylation: Purview and parameters. Nat. Rev. Mol. Cell Biol. 2005, 6, 150–166. [Google Scholar] [CrossRef]
- Godoy, L.; Gonzàlez-Duarte, R.; Albalat, R. S-nitrosogluthathione reductase activity of amphioxus ADH3: Insights into the nitric oxide metabolism. Int. J. Biol. Sci. 2006, 2, 117–124. [Google Scholar]
- Leterrier, M.; Chaki, M.; Airaki, M., Valderrama; Palma, J.M.; Barroso, J.B.; Corpas, F.J. Function of S-nitrosoglutathione reductase (GSNOR) in plant development and under biotic/abiotic stress. Plant Signal. Behav. 2011, 6, 789–793. [Google Scholar] [CrossRef]
- Hsiao, G.; Shen, M.Y.; Lin, K.H.; Lan, M.H.; Wu, L.Y.; Chou, D.S.; Lin, C.H.; Su, C.H.; Sheu, J.R. Antioxidative and hepatoprotective effects of Antrodia camphorata extract. J. Agric. Food Chem. 2003, 51, 3302–3308. [Google Scholar]
- Shen, Y.C.; Chou, C.J.; Wang, Y.H.; Chen, C.F.; Chou, Y.C.; Lu, M.K. Anti-inflammatory activity of the extracts from mycelia of Antrodia camphorata cultured with water-soluble fractions from five different Cinnamomum species. FEMS Microbiol. Lett. 2004, 231, 137–143. [Google Scholar] [CrossRef]
- Liau, Y.J.; Wen, L.; Shaw, J.F.; Lin, C.T. A highly stable cambialistic-superoxide dismutase from Antrodia camphorata: Expression in yeast and enzyme properties. J. Biotechnol. 2007, 131, 84–91. [Google Scholar] [CrossRef]
- Ken, C.F.; Chen, H.T.; Chang, R.C.; Lin, C.T. Biochemical characterization of a catalase from Antrodia camphorata : Expression in Escherichia coli and enzyme properties. Bot. Stud. 2008, 49, 119–125. [Google Scholar]
- Huang, C.Y.; Ken, C.F.; Wen, L.; Lin, C.T. An enzyme possessing both glutathione-dependent formaldehyde dehydrogenase and S-nitrosoglutathione reductase from Antrodia camphorata. Food Chem. 2009, 112, 795–802. [Google Scholar] [CrossRef]
- Liau, Y.J.; Chen, Y.T.; Lin, C.Y.; Huang, J.K.; Lin, C.T. Characterization of 2-Cys peroxiredoxin isozyme (Prx1) from Taiwanofungus camphorata (Niu-changchih): Expression and enzyme properties. Food Chem. 2010, 119, 154–160. [Google Scholar] [CrossRef]
- Chen, H.T.; Lin, C.Y.; Ken, C.F.; Wen, L.; Lin, C.T. Putative phospholipid hydroperoxide glutathione peroxidase from Antrodia camphorata. Food Chem. 2009, 115, 476–482. [Google Scholar] [CrossRef]
- Vistica, D.T.; Skehan, P.; Scudiero, D.; Monks, A.; Pittman, A.; Boyd, M.R. Tetrazolium-based assays for cellular viability: A critical examination of selected parameters affecting formazan production. Cancer Res. 1991, 51, 2515–2520. [Google Scholar]
- Lozinsky, O.V.; Lushchak, O.V.; Kryshchuk, N.I.; Shchypanska, N.Y.; Riabkina, A.H.; Skarbek, S.V.; Maksymiv, I.V.; Storey, J.M.; Storey, K.B.; Lushchak, V.I. S-Nitrosoglutathione-induced toxicity in Drosophila melanogaster: Delayed pupation and induced mild oxidative/nitrosative stress in eclosed flies. Comp. Biochem. Physiol. A 2013, 164, 162–70. [Google Scholar] [CrossRef]
- Mathews, W.R.; Kerr, S.W. Biological activity of S-nitrosothiols: The role of nitric oxide. J. Pharm. Exp. Ther. 1993, 267, 1529–1537. [Google Scholar]
- Van Engeland, M.; Nieland, L.J.; Ramaekers, F.C.; Schutte, B.; Reutelingsperger, C.P. Annexin V-affinity assay: A review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry 1998, 31, 1–9. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ken, C.-F.; Huang, C.-Y.; Wen, L.; Huang, J.-K.; Lin, C.-T. Modulation of Nitrosative Stress via Glutathione-Dependent Formaldehyde Dehydrogenase and S-Nitrosoglutathione Reductase. Int. J. Mol. Sci. 2014, 15, 14166-14179. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms150814166
Ken C-F, Huang C-Y, Wen L, Huang J-K, Lin C-T. Modulation of Nitrosative Stress via Glutathione-Dependent Formaldehyde Dehydrogenase and S-Nitrosoglutathione Reductase. International Journal of Molecular Sciences. 2014; 15(8):14166-14179. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms150814166
Chicago/Turabian StyleKen, Chuian-Fu, Chih-Yu Huang, Lisa Wen, Jenq-Kuen Huang, and Chi-Tsai Lin. 2014. "Modulation of Nitrosative Stress via Glutathione-Dependent Formaldehyde Dehydrogenase and S-Nitrosoglutathione Reductase" International Journal of Molecular Sciences 15, no. 8: 14166-14179. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms150814166