
Ferulic Acid Exerts Anti-Angiogenic and Anti-Tumor Activity by Targeting Fibroblast Growth Factor Receptor 1-Mediated Angiogenesis

Abstract
:1. Introduction
2. Results and Discussion
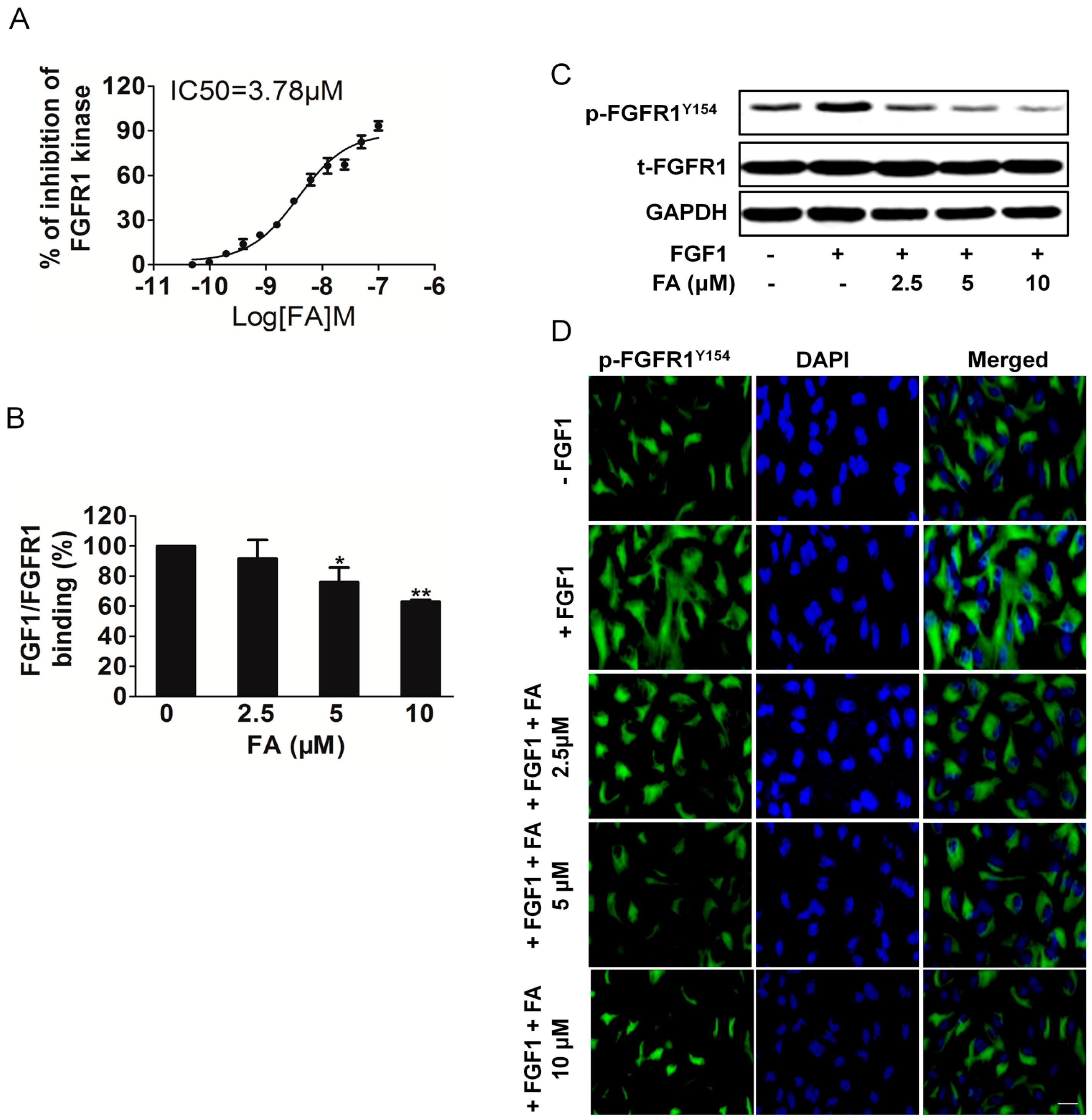
2.1. Kinase Inhibition Profile of Ferulic Acid (FA)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinase | Inhibition Rate at 1 µM (%) |
|---|---|
| FGFR1 | 92 ± 3 |
| FGFR2 | 64 ± 1 |
| VEGFR2 | 4 ± 0 |
| Flt3 | 8 ± 2 |
| PDGFR-α | 1 ± 0 |
| PDGFR-β | 2 ± 0 |
| c-Kit | 6 ± 1 |
| Aurora-A | −2 ± 2 |
| Haspin | 7 ± 2 |
| ErbB4 | 10 ± 2 |
| IKKβ | −5 ± 2 |
| c-Met | 11 ± 0 |
| CDK2 | −13 ± 2 |
| PI3K | 20 ± 1 |
| EGFR | 4 ± 1 |
| JNK | −9 ± 0 |
| mTOR | 5 ± 2 |
| GSK3β | 4 ± 0 |
| c-RAF | 18 ± 4 |
| JAK | 5 ± 1 |
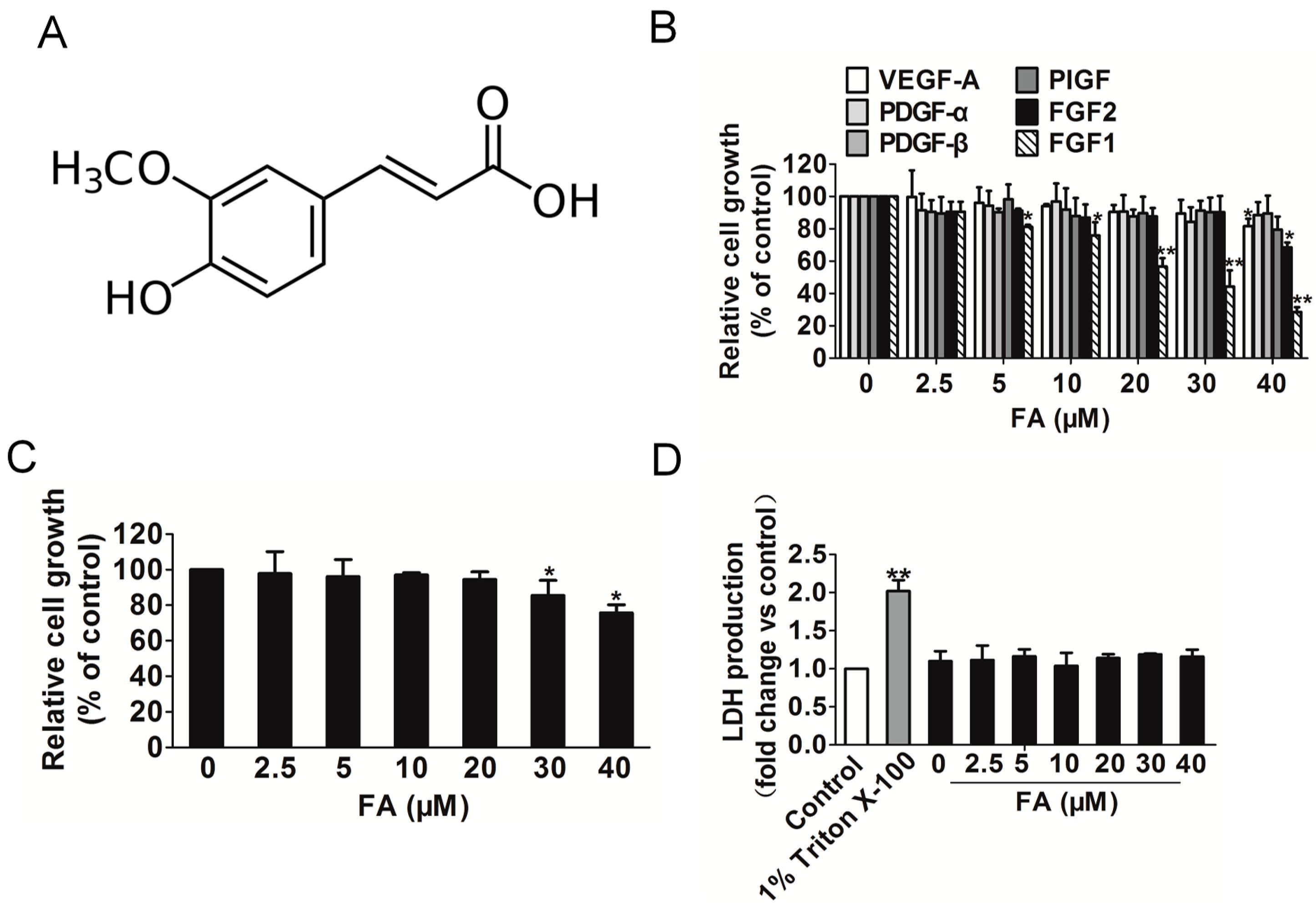
2.2. FA Inhibits Fibroblast Growth Factor 1 (FGF1) and Induces Human Umbilical Vein Endothelial Cells (HUVEC) Growth

2.3. FA Inhibits FGFR1 Kinase Activity in HUVEC

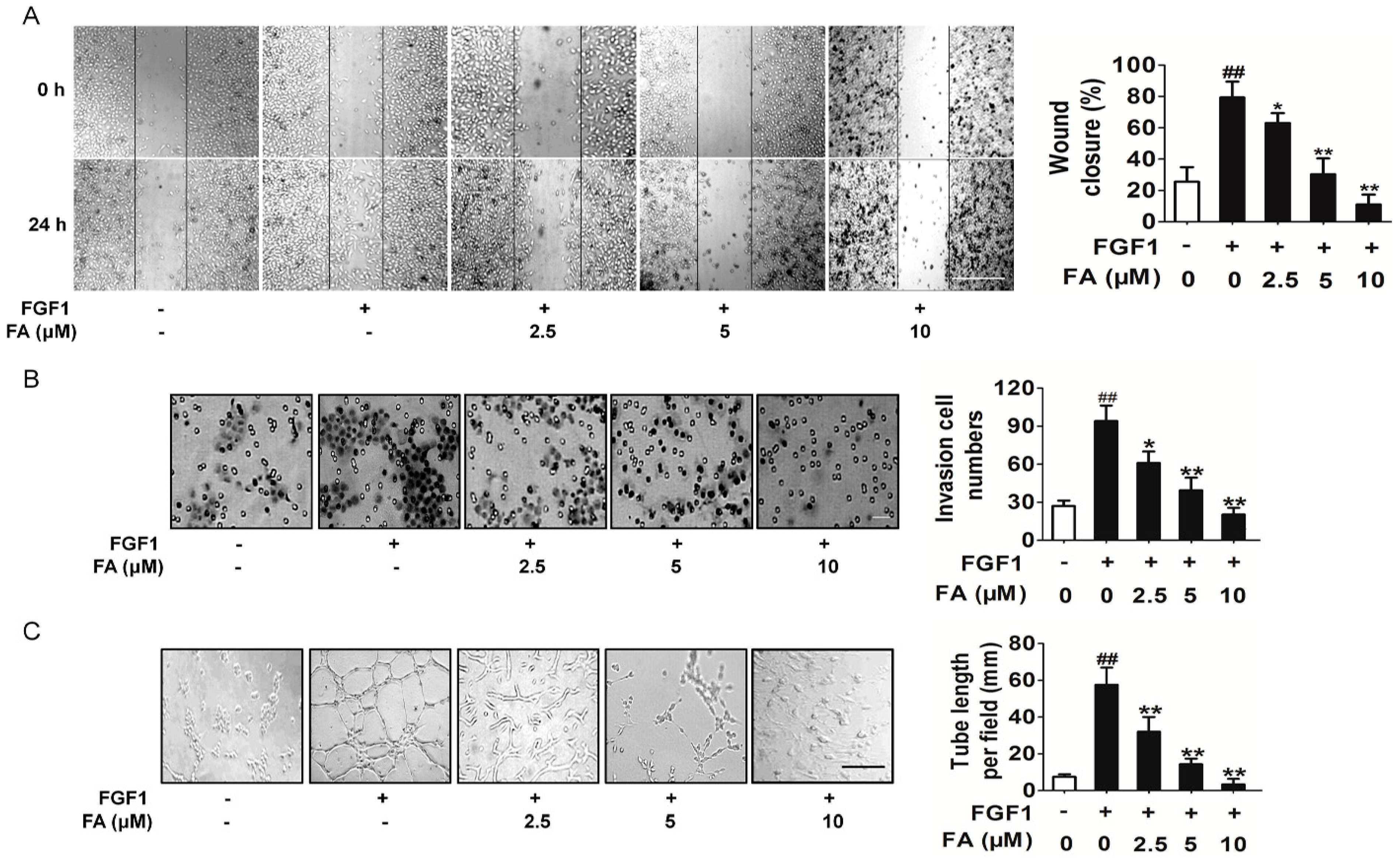
2.4. FA Inhibits FGF1-Induced Migration, Invasion and Tubular Structure Formation of HUVEC

2.5. FA Inhibits Angiogenesis ex Vivo and in Vivo

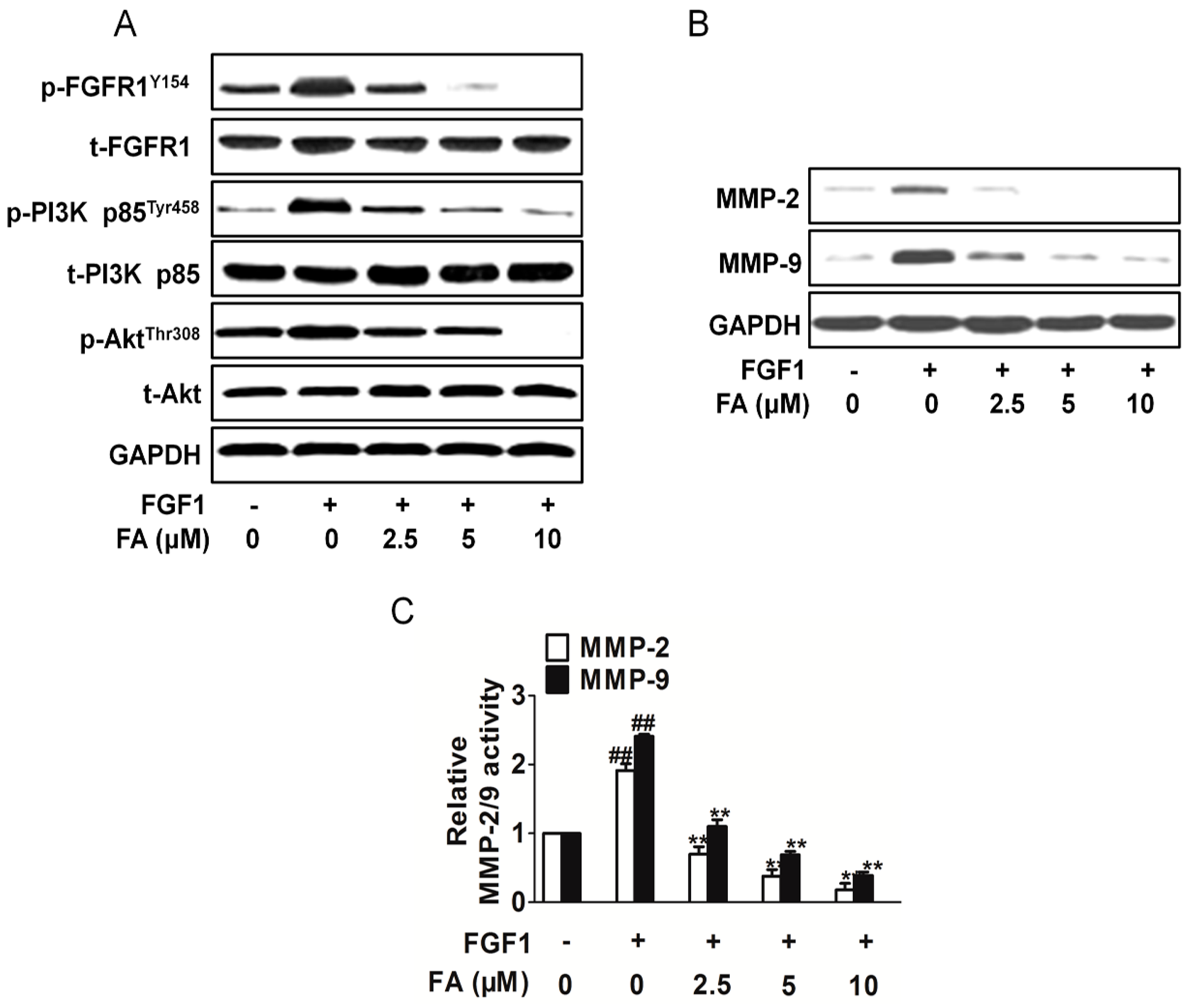
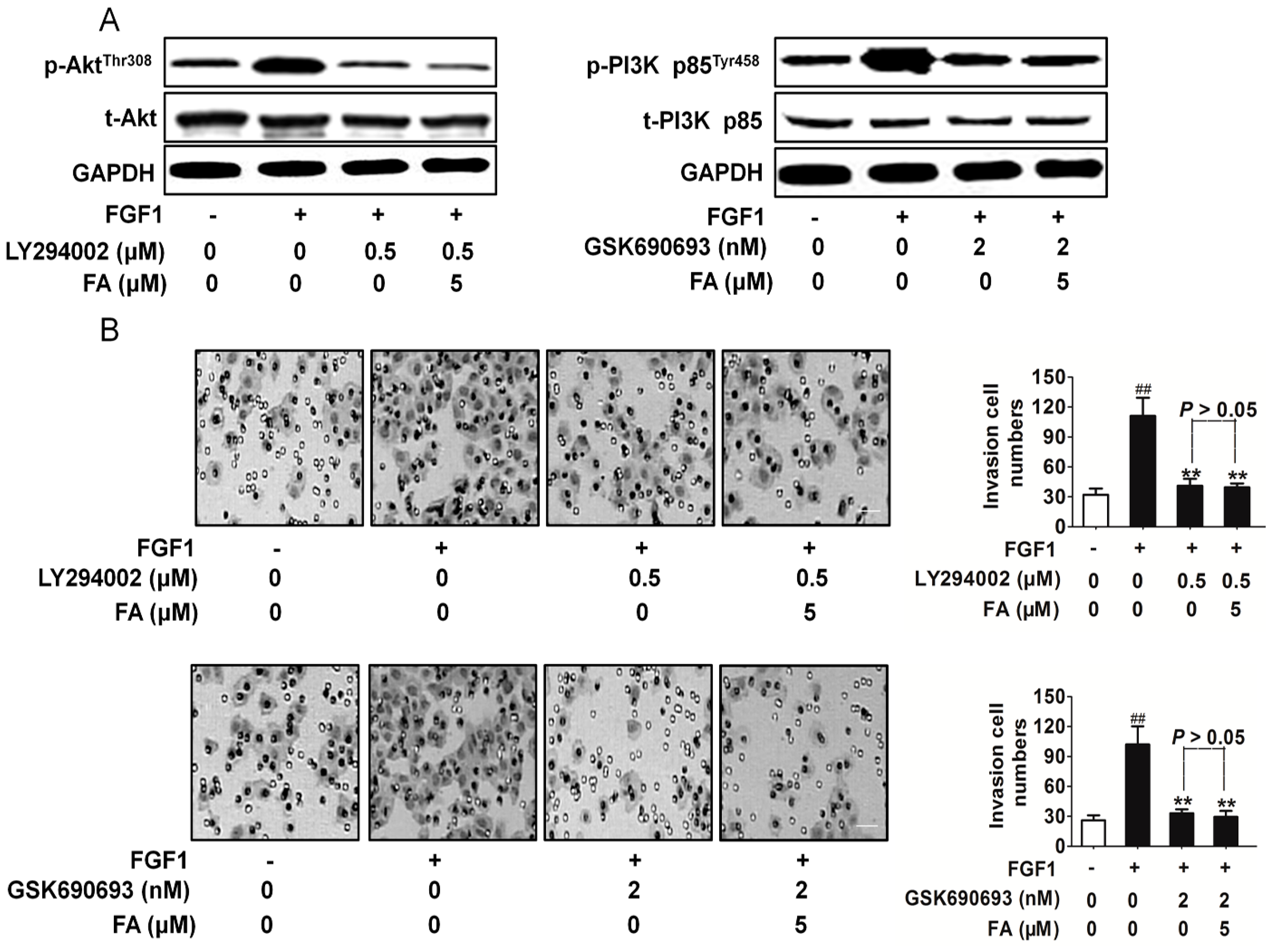
2.6. FA Inhibits Activation of PI3K/Akt Signaling Induced by FGF1 in HUVEC

2.7. FA Inhibits HUVEC Invasion Dependent on PI3K/Akt Signaling

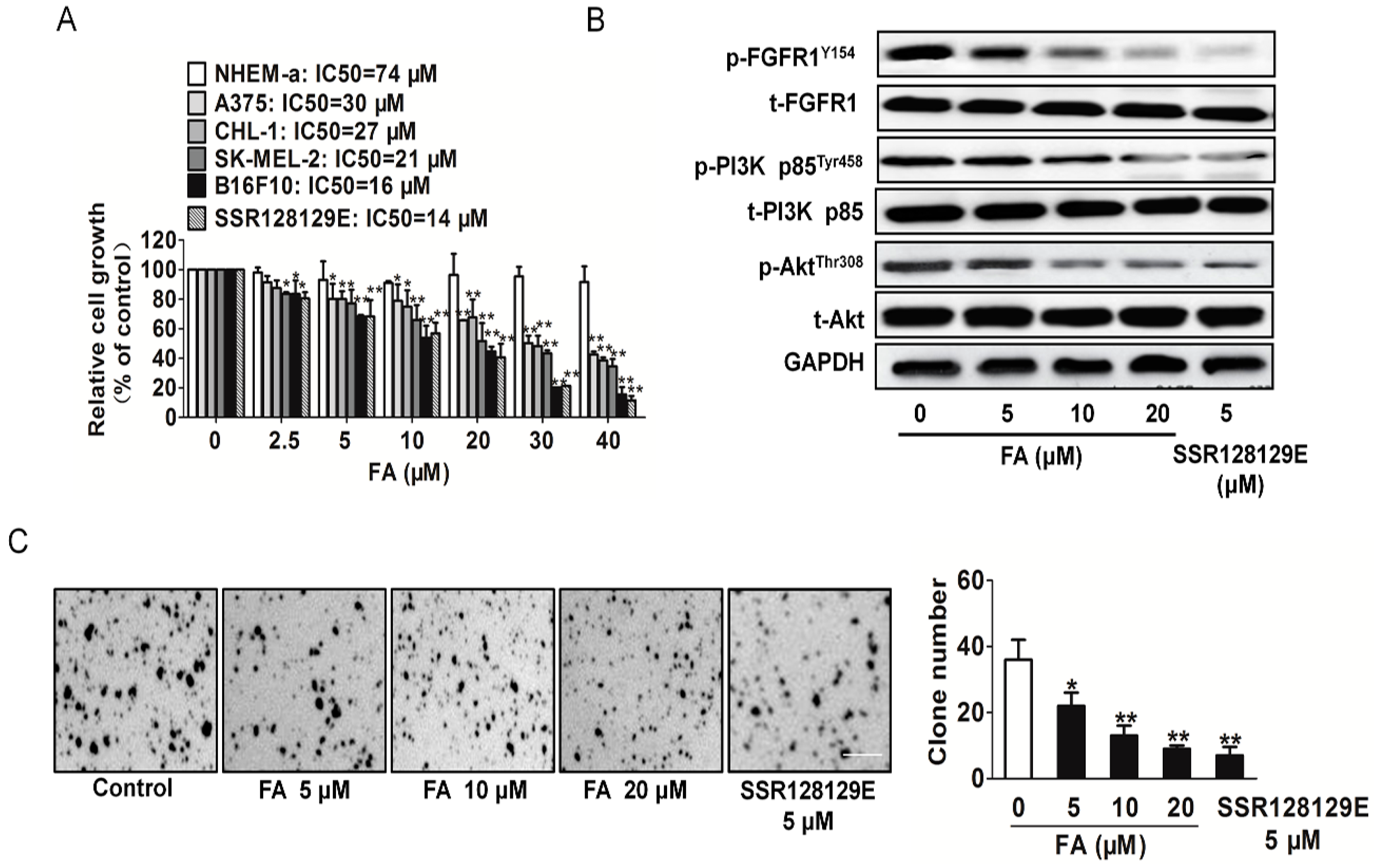
2.8. FA Inhibits Melanoma Cell Proliferation and the FGFR1 Downstream Signaling Pathway

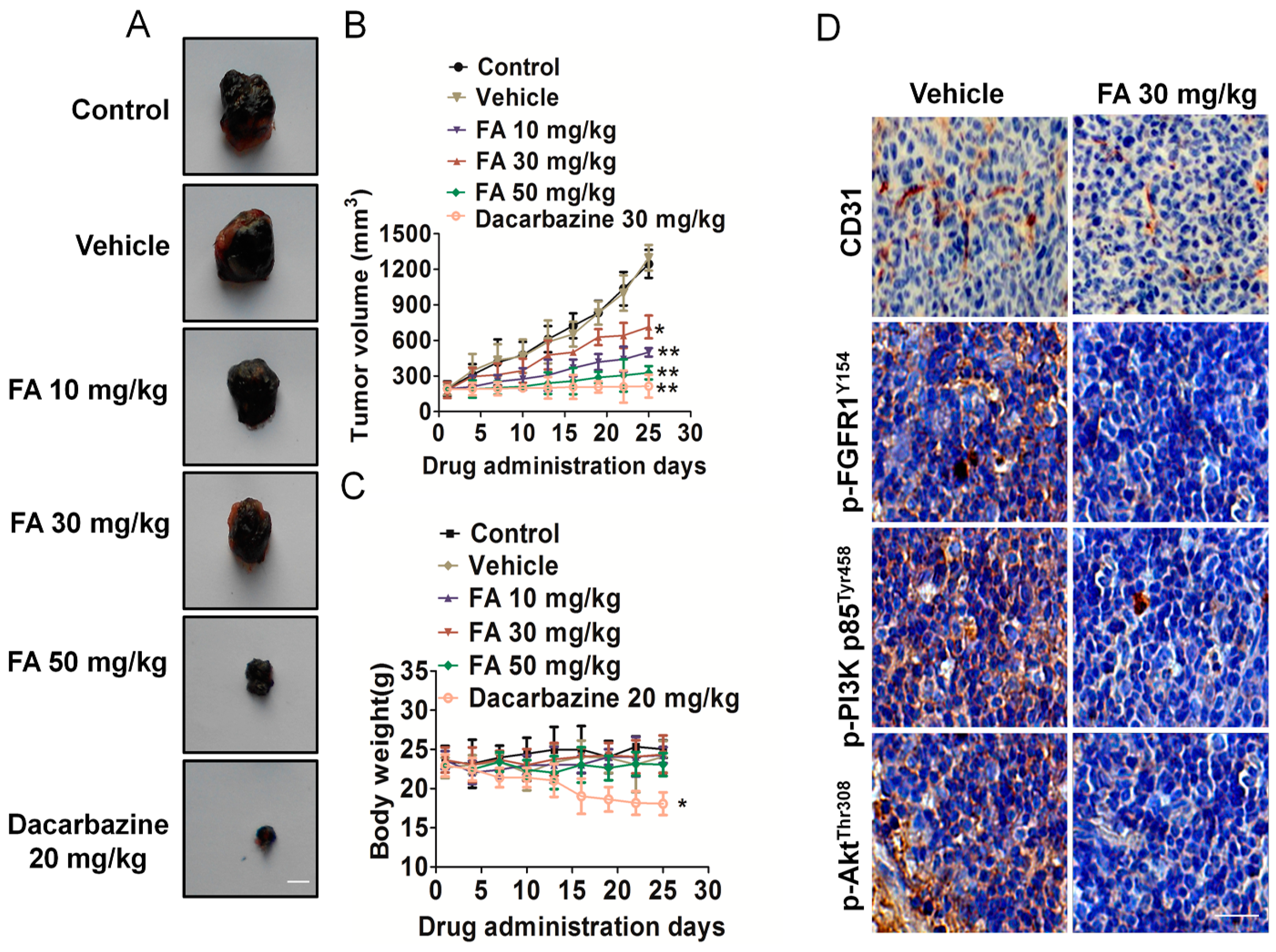
2.9. FA Inhibits Tumor Growth and Angiogenesis in a B16F10 Cell Xenograft Model

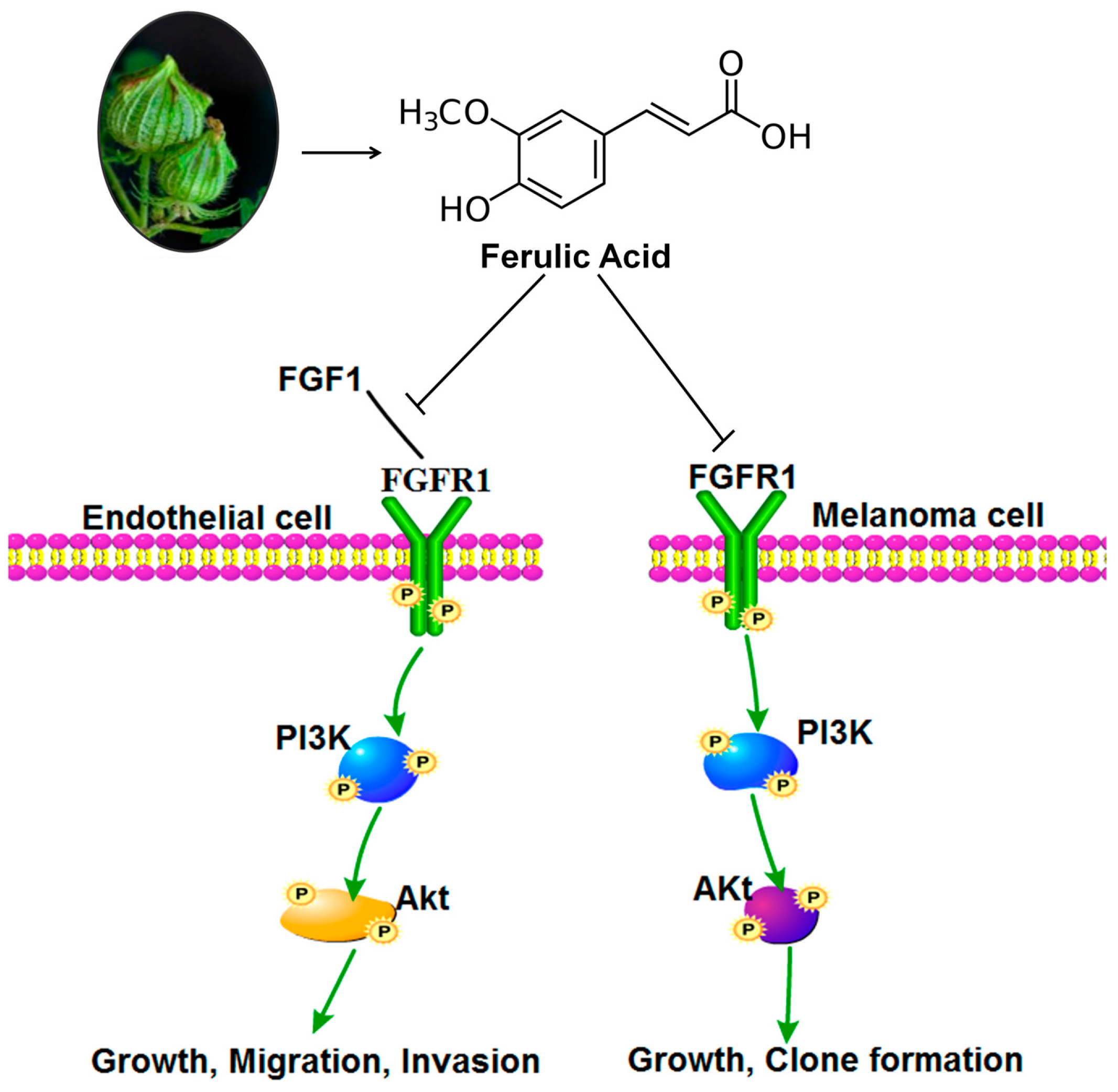
2.10. Discussion

3. Experimental Section
3.1. Cell Culture and Reagents
3.2. MTT Proliferation Assay
3.3. Lactate Dehydrogenase Toxicity Assay
3.4. Wound Healing
3.5. Invasion Assay
3.6. Anchorage-Independent Growth Assay
3.7. Tube Formation Assay
3.8. Rat Aortic Ring Assay
3.9. Chick Chorioallantoic Membrane Assay
3.10. Western Blotting Assay
3.11. FGFR1 Kinase Inhibition Assay
3.12. FGFR Binding Assay
3.13. Matrix Metalloproteins Activity Assay
3.14. Immunofluorescence Analysis
3.15. Xenograft Models and Immunohistochemistry Detections
3.16. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
Abbreviations
References
- Tchernev, G.; Chokoeva, A.A.; Wollina, U.; Patterson, J.W.; Lotti, T. Melanoma imitators-how to prevent the aggressive approach? J. Biol. Regul. Homeost. Agents 2015, 29, 75–80. [Google Scholar] [PubMed]
- Welti, J.; Loges, S.; Dimmeler, S.; Carmeliet, P. Recent molecular discoveries in angiogenesis and antiangiogenic therapies in cancer. J. Clin. Investig. 2013, 123, 3190–3200. [Google Scholar] [CrossRef] [PubMed]
- Meierjohann, S. Hypoxia-independent drivers of melanoma angiogenesis. Front. Oncol. 2015, 5, 102. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Koo, B.S.; Park, S.Y.; Kim, Y.M. Anti-angiogenic effects of resveratrol in combination with 5-fluorouracil on B16 murine melanoma cells. Mol. Med. Rep. 2015, 12, 2777–2783. [Google Scholar] [CrossRef] [PubMed]
- De Brot, S.; Ntekim, A.; Cardenas, R.; James, V.; Allegrucci, C.; Heery, D.M.; Bates, D.O.; Ødum, N.; Persson, J.L.; Mongan, N.P. Regulation of vascular endothelial growth factor in prostate cancer. Endocr. Relat. Cancer 2015, 22, R107–R123. [Google Scholar]
- Alfaidy, N.; Hoffmann, P.; Boufettal, H.; Samouh, N.; Aboussaouira, T.; Benharouga, M.; Feige, J.J.; Brouillet, S. The multiple roles of EG-VEGF/PROK1 in normal and pathological placental angiogenesis. Biomed. Res. Int. 2014, 2014, 451906. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Katoh, M. Cross-talk of WNT and FGF signaling pathways at GSK3β to regulate β-catenin and SNAIL signaling cascades. Cancer Biol. Ther. 2006, 5, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.J.; Weylie, B.; Hu, C.; Zhu, J.; Forough, R. FGFR1/PI3K/AKT signaling pathway is a novel target for antiangiogenic effects of the cancer drug fumagillin (TNP-470). J. Cell. Biochem. 2007, 101, 1492–1504. [Google Scholar] [CrossRef] [PubMed]
- Vara, J.A.F.; Casado, E.; de Castro, J.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wang, X.; Jin, H. EGFR-TKI resistance in NSCLC patients: Mechanisms and strategies. Am. J. Cancer Res. 2014, 4, 411–435. [Google Scholar] [PubMed]
- Choi, Y.E.; Park, E. Ferulic acid in combination with PARP inhibitor sensitizes breast cancer cells as chemotherapeutic strategy. Biochem. Biophys. Res. Commun. 2015, 458, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.C.; Chyau, C.C.; Wang, H.E.; Chang, C.H.; Chen, K.C.; Chou, K.Y.; Peng, R.Y. Cytotoxicity of ferulic acid on T24 cell line differentiated by different microenvironments. Biomed. Res. Int. 2013, 2013, 579859. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Ho, C.T. Chemopreventive effects of natural dietary compounds on cancer development. Chem. Soc. Rev. 2008, 37, 2558–2574. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Ryu, K.; Kang, J.H.; Choi, A.J.; Kim, T.I.; Oh, J.M. Anticancer activity of ferulic acid-inorganic nanohybrids synthesized via two different hybridization routes, reconstruction and exfoliation-reassembly. Sci. World J. 2013, 2013, 421967. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.M.; Chiu, J.H.; Wu, I.H.; Wang, B.W.; Pan, C.M.; Chen, Y.H. Ferulic acid augments angiogenesis via VEGF, PDGF and HIF-1α. J. Nutr. Biochem. 2010, 21, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yuan, Z.; Zhao, H.; Ju, D.; Chen, Y.; Chen, X.; Zhang, J. Ferulic acid promotes endothelial cells proliferation through up-regulating cyclin D1 and VEGF. J. Ethnopharmacol. 2011, 137, 992–997. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Yang, J.; Zhao, G.; Yuan, Y. Ferulic acid inhibits endothelial cell proliferation through NO down-regulating ERK1/2 pathway. J. Cell. Biochem. 2004, 93, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Dol-Gleizes, F.; Delesque-Touchard, N.; Marès, A.M.; Nestor, A.L.; Schaeffer, P.; Bono, F. A new synthetic FGF receptor antagonist inhibits arteriosclerosis in a mouse vein graft model and atherosclerosis in apolipoprotein E-deficient mice. PLoS ONE 2013, 8, e80027. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Kim, J.Y.; Kim, H.J.; Park, K.G.; Harris, R.A.; Cho, W.J.; Lee, J.T.; Lee, I.K. Scoparone exerts anti-tumor activity against DU145 prostate cancer cells via inhibition of STAT3 activity. PLoS ONE 2013, 8, e80391. [Google Scholar] [CrossRef] [PubMed]
- Mbengue, A.; Bhattacharjee, S.; Pandharkar, T.; Liu, H.; Estiu, G.; Stahelin, R.V.; Rizk, S.S.; Njimoh, D.L.; Ryan, Y.; Chotivanich, K.; et al. A molecular mechanism of artemisinin resistance in Plasmodium falciparum malaria. Nature 2015, 520, 683–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salt, M.B.; Bandyopadhyay, S.; McCormick, F. Epithelial-to-mesenchymal transition rewires the molecular path to PI3K-dependent proliferation. Cancer Discov. 2014, 4, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Nakagama, H. FGF receptors: Cancer biology and therapeutics. Med. Res. Rev. 2014, 34, 280–300. [Google Scholar] [CrossRef] [PubMed]
- Rapisarda, A.; Melillo, G. Role of the VEGF/VEGFR axis in cancer biology and therapy. Adv. Cancer Res. 2012, 114, 237–267. [Google Scholar] [PubMed]
- Dey, J.H.; Bianchi, F.; Voshol, J.; Bonenfant, D.; Oakeley, E.J.; Hynes, N.E. Targeting fibroblast growth factor receptors blocks PI3K/AKT signaling, induces apoptosis, and impairs mammary tumor outgrowth and metastasis. Cancer Res. 2010, 70, 4151–4162. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Yan, S.; Xue, H.; Zhang, P.; Sun, J.; Li, G. JSI-124 suppresses invasion and angiogenesis of glioblastoma cells in vitro. PLoS ONE 2015, 10, e0118894. [Google Scholar] [CrossRef] [PubMed]
- Lau, M.T.; So, W.K.; Leung, P.C. Fibroblast growth factor 2 induces E-cadherin down-regulation via PI3K/Akt/mTOR and MAPK/ERK signaling in ovarian cancer cells. PLoS ONE 2013, 8, e59083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xie, Q.; Shan, S.J.; Tabor, M.K.; Junkins-Hopkins, J.M. Reduced expression of RECK protein may help differentiate cutaneous malignant melanoma from melanocytic nevus. Anticancer Res. 2015, 35, 3185–3191. [Google Scholar] [PubMed]
- Gong, M.; Yang, J.T.; Liu, Y.Q.; Tang, L.H.; Wang, Y.; Wang, L.J.; Zhang, F.J.; Yan, M. Irradiation can selectively kill tumor cells while preserving erythrocyte viability in a co-culture system. PLoS ONE 2015, 10, e0127181. [Google Scholar] [CrossRef] [PubMed]
- Adaramoye, O.; Erguen, B.; Oyebode, O.; Nitzsche, B.; Höpfner, M.; Jung, K.; Rabien, A. Antioxidant, antiangiogenic and antiproliferative activities of root methanol extract of Calliandra portoricensis in human prostate cancer cells. J. Integr. Med. 2015, 13, 185–193. [Google Scholar] [CrossRef]
- Khoury, N.; El-Hayek, S.; Tarras, O.; El-Sabban, M.; El-Sibai, M.; Rizk, S. Kefir exhibits anti-proliferative and pro-apoptotic effects on colon adenocarcinoma cells with no significant effects on cell migration and invasion. Int. J. Oncol. 2014, 45, 2117–2127. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Zhou, X.; Mak, N.K.; Leung, K.N. Tryptanthrin inhibits angiogenesis by targeting the VEGFR2-mediated ERK1/2 signalling pathway. PLoS ONE 2013, 8, e82294. [Google Scholar] [CrossRef] [PubMed]
- Otjacques, E.; Binsfeld, M.; Rocks, N.; Blacher, S.; Vanderkerken, K.; Noel, A.; Beguin, Y.; Cataldo, D.; Caers, J. Mithramycin exerts an anti-myeloma effect and displays anti-angiogenic effects through up-regulation of anti-angiogenic factors. PLoS ONE 2013, 8, e62818. [Google Scholar] [CrossRef] [PubMed]
- Kue, C.S.; Tan, K.Y.; Lam, M.L.; Lee, H.B. Chick embryo chorioallantoic membrane (CAM): An alternative predictive model in acute toxicological studies for anti-cancer drugs. Exp. Anim. 2015, 64, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Saraswati, S.; Kanaujia, P.K.; Kumar, S.; Kumar, R.; Alhaider, A.A. Tylophorine, a phenanthraindolizidine alkaloid isolated from Tylophora indica exerts antiangiogenic and antitumor activity by targeting vascular endothelial growth factor receptor 2-mediated angiogenesis. Mol. Cancer 2013, 12, 82. [Google Scholar] [CrossRef] [PubMed]
- Campton, D.E.; Ramirez, A.B.; Nordberg, J.J.; Drovetto, N.; Clein, A.C.; Varshavskaya, P.; Friemel, B.H.; Quarre, S.; Breman, A.; Dorschner, M.; et al. High-recovery visual identification and single-cell retrieval of circulating tumor cells for genomic analysis using a dual-technology platform integrated with automated immunofluorescence staining. BMC Cancer 2015, 15, 360. [Google Scholar] [CrossRef] [PubMed]
- Yeh, M.Y.; Wu, M.F.; Shang, H.S.; Chang, J.B.; Shih, Y.L.; Chen, Y.L.; Hung, H.F.; Lu, H.F.; Yeh, C.; Wood, W.G.; et al. Effects of chitosan on xenograft models of melanoma in C57BL/6 mice and hepatoma formation in SCID mice. Anticancer Res. 2013, 33, 4867–4873. [Google Scholar] [PubMed]
- Theophanous, E.; Petraki, C.; Scorilas, A.; Komborozos, V.; Veloudis, G.; Varga, J.L.; Zarandi, M.; Schally, A.V.; Koutsilieris, M. The immunohistochemical expression of growth hormone-releasing hormone receptor splice variant 1 is a favorable prognostic marker in colorectal cancer. Mol. Med. 2009, 15, 242–247. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.-W.; Jiang, J.-S.; Lu, W.-Q. Ferulic Acid Exerts Anti-Angiogenic and Anti-Tumor Activity by Targeting Fibroblast Growth Factor Receptor 1-Mediated Angiogenesis. Int. J. Mol. Sci. 2015, 16, 24011-24031. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161024011
Yang G-W, Jiang J-S, Lu W-Q. Ferulic Acid Exerts Anti-Angiogenic and Anti-Tumor Activity by Targeting Fibroblast Growth Factor Receptor 1-Mediated Angiogenesis. International Journal of Molecular Sciences. 2015; 16(10):24011-24031. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161024011
Chicago/Turabian StyleYang, Guang-Wei, Jin-Song Jiang, and Wei-Qin Lu. 2015. "Ferulic Acid Exerts Anti-Angiogenic and Anti-Tumor Activity by Targeting Fibroblast Growth Factor Receptor 1-Mediated Angiogenesis" International Journal of Molecular Sciences 16, no. 10: 24011-24031. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161024011