
Increased Uptake of Chelated Copper Ions by Lolium perenne Attributed to Amplified Membrane and Endodermal Damage

Abstract
:
1. Introduction
2. Results and Discussion
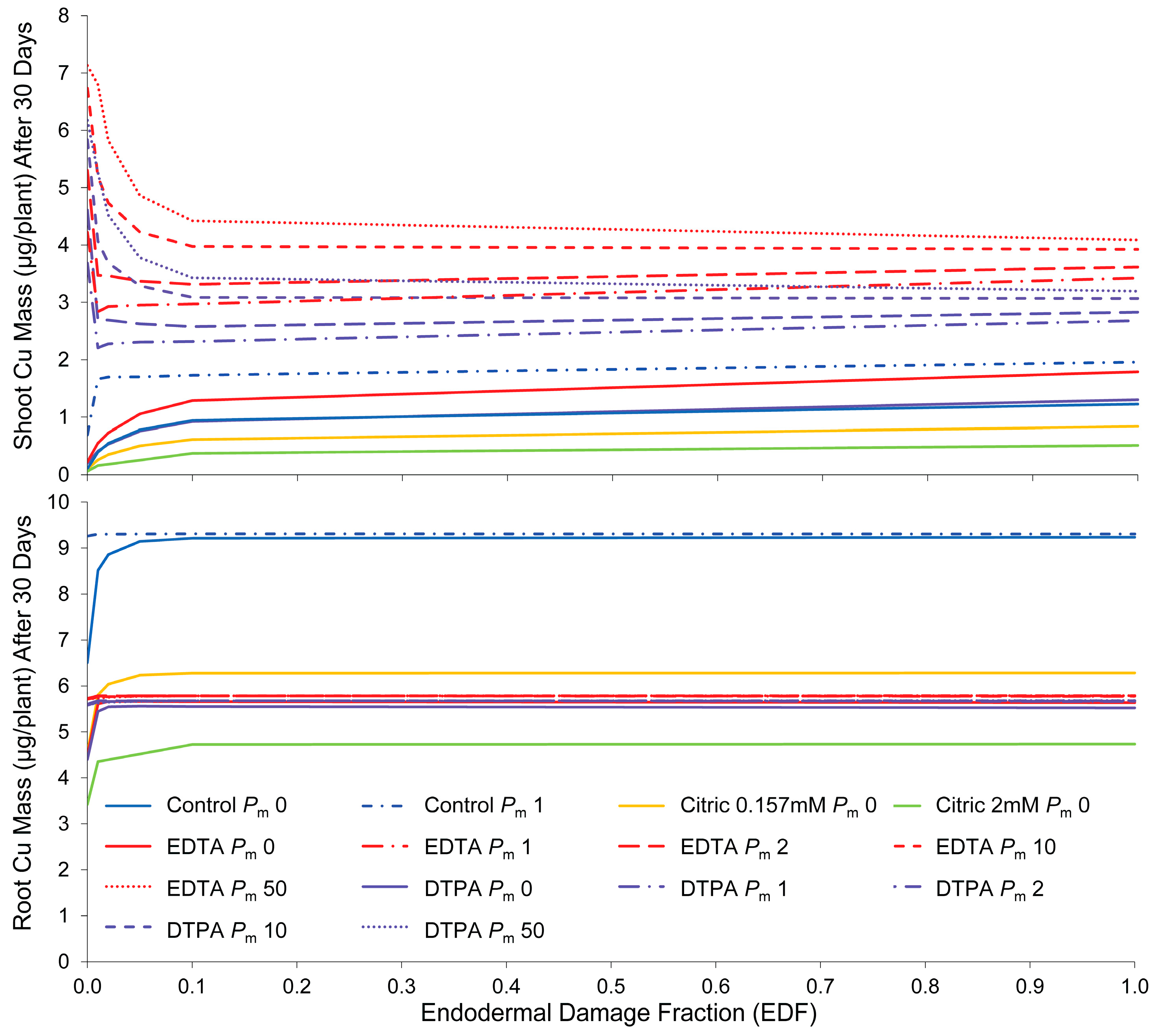
2.1. Identification of Critical Parameters Using Sensitivity Analysis
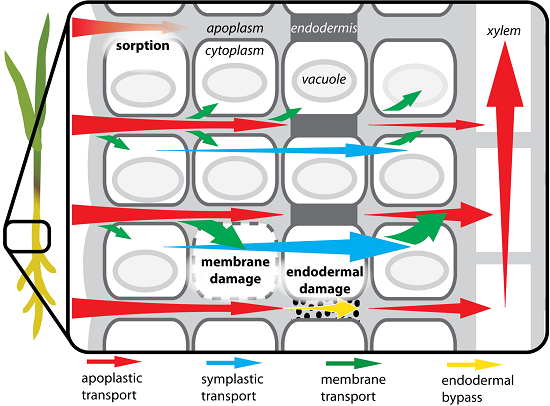
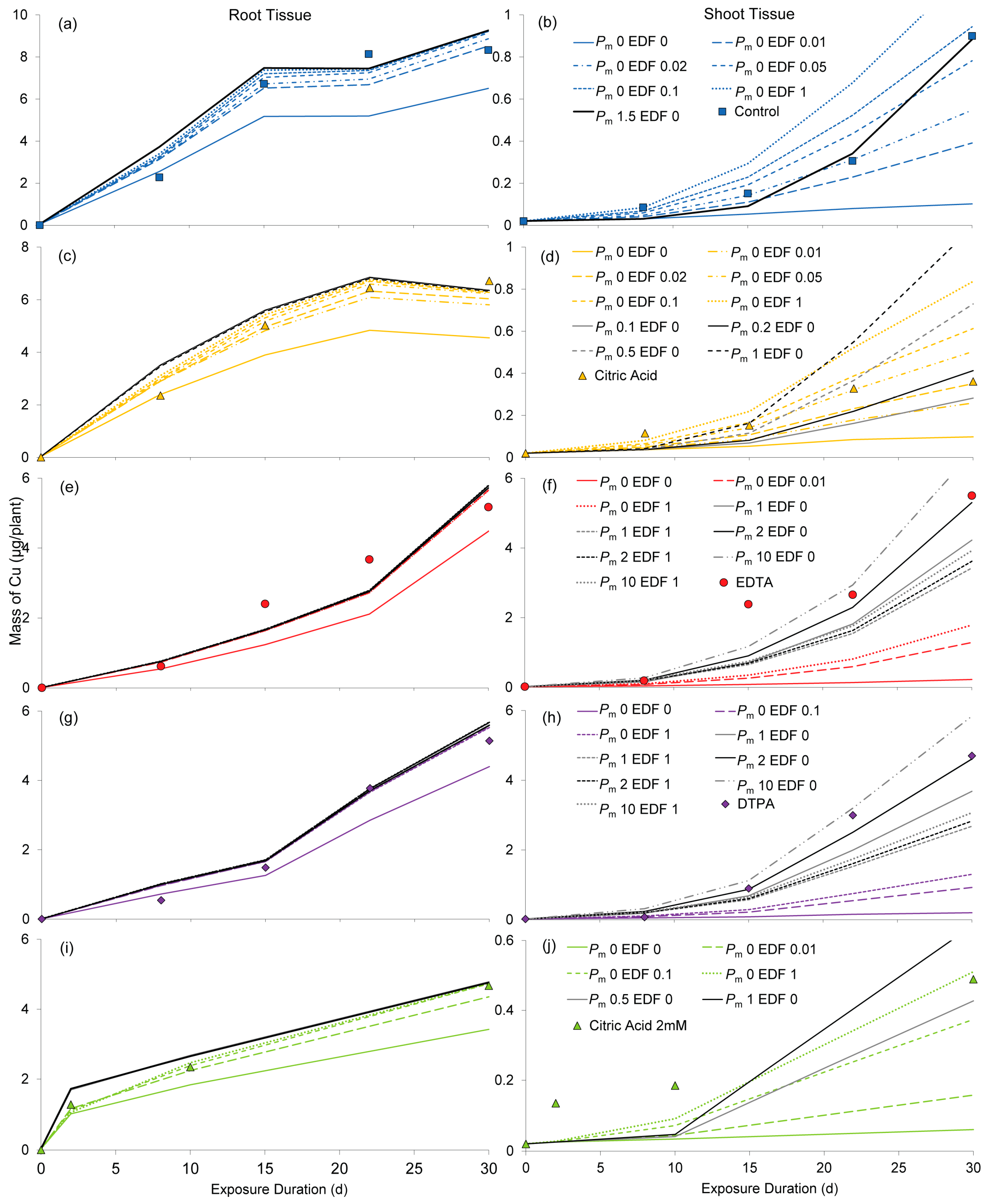
2.2. Simulations


{kind=link}
{kind=link}
{kind=link}
{kind=link}
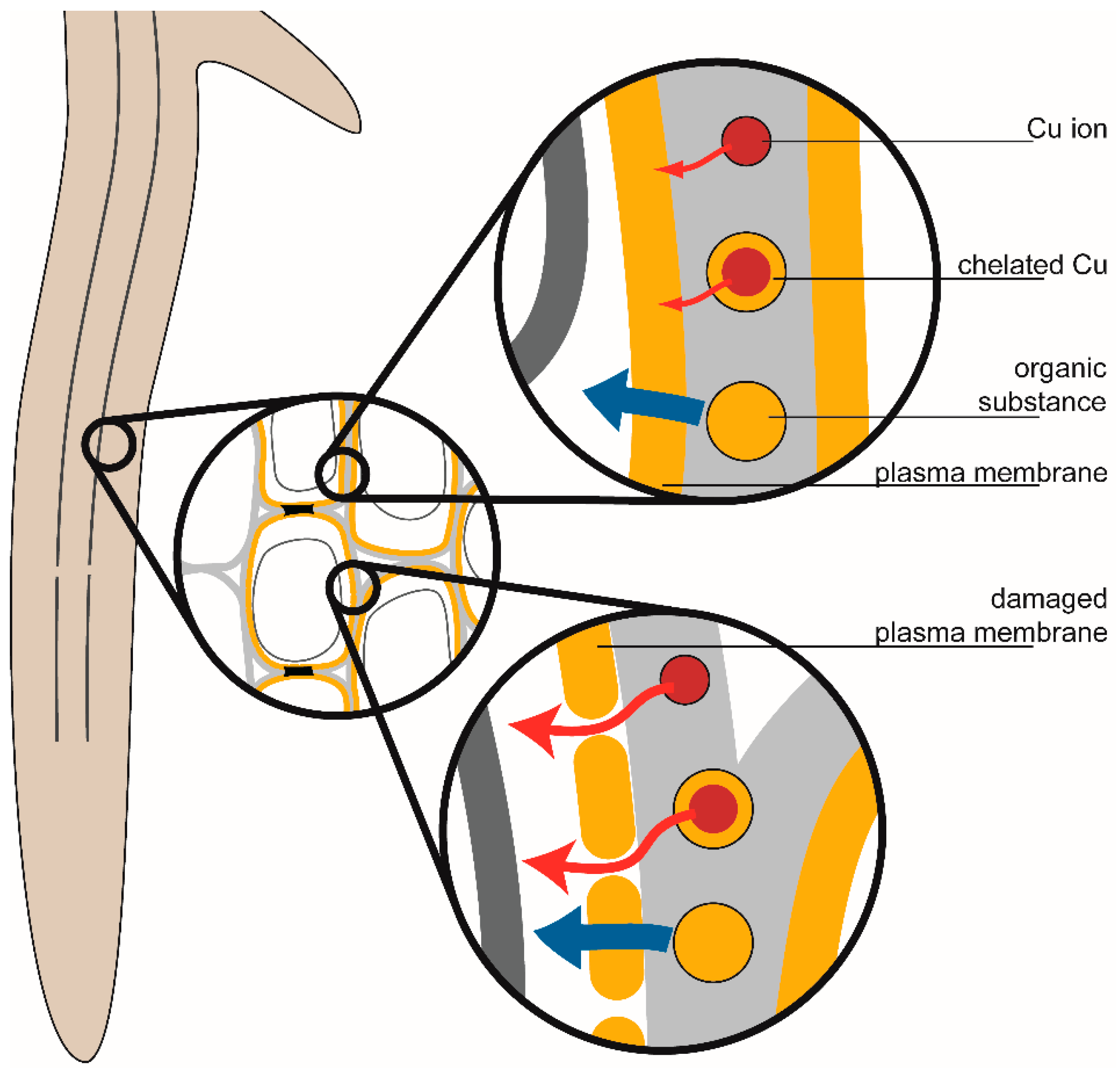{kind=link}
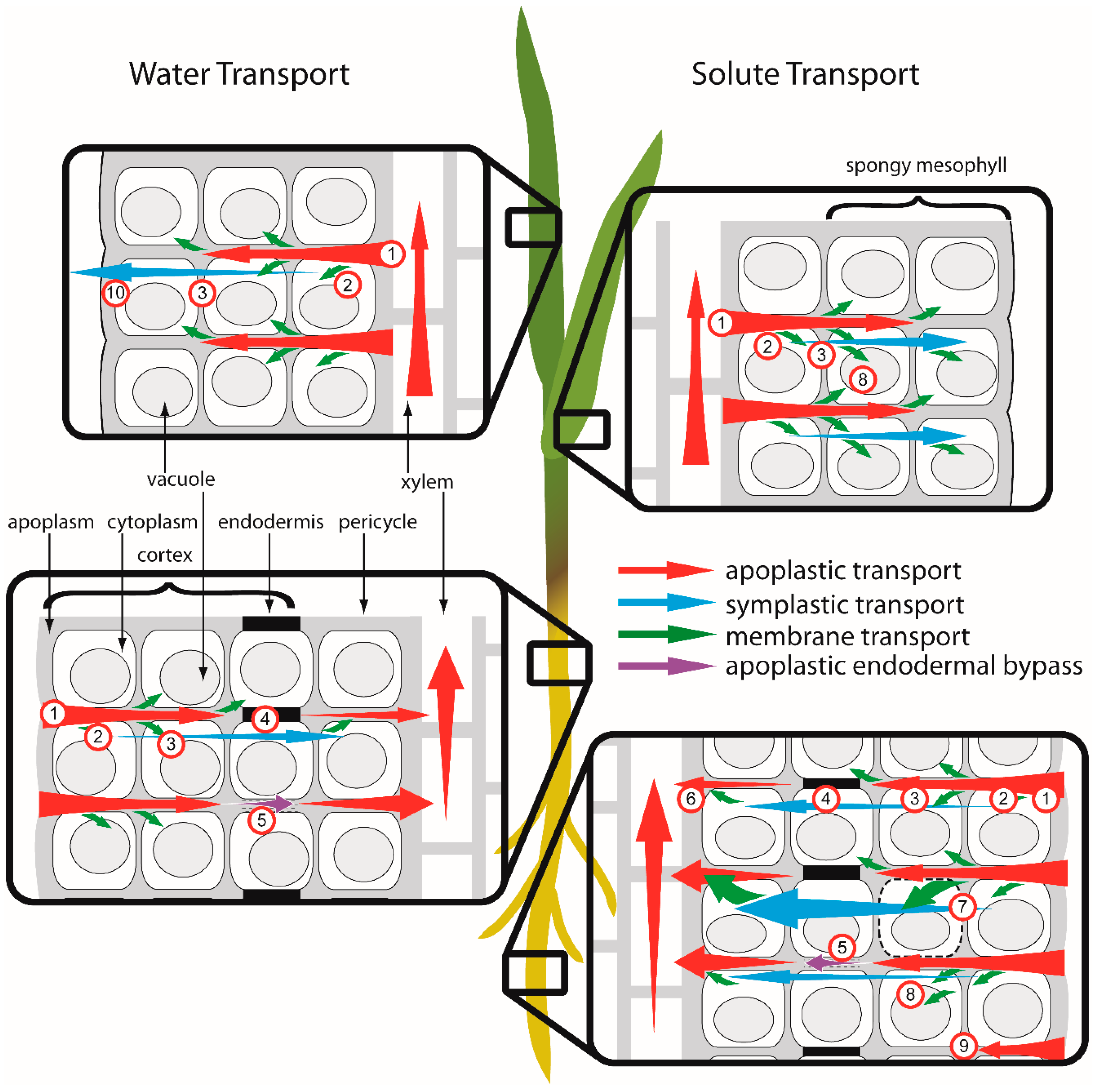{kind=link}
| Treatment | Pathway | Fitted Parameters | |||
|---|---|---|---|---|---|
| Da (m2/day) | KL (m3/g) | EDF | Pm (m/day) | ||
| Control | Apoplast | 3.1 × 10−5 | 0.0015 | 0.05 | 0 |
| Symplast | 0 | 1.5 | |||
| Citric acid (0.157 mM) | Apoplast | 1.7 × 10−5 | 0.00085 | 0.05 | 0 |
| Symplast | 0 | 0.4 (0.2–0.5) | |||
| Citric acid (2 mM) | Apoplast | 1.4 × 10−5 | 0.0009 | 1 | 0 |
| Symplast | 0 | 0.6 (0.5–1) | |||
| EDTA | Symplast | 1.2 × 10−5 | 0.0002 | 0 | 2 |
| DTPA | Symplast | 0.9 × 10−5 | 0.0002 | 0 | 2 |
2.3. Evaluation of Simulated Transport Mechanisms
2.3.1. Sorption
2.3.2. Endodermal Damage

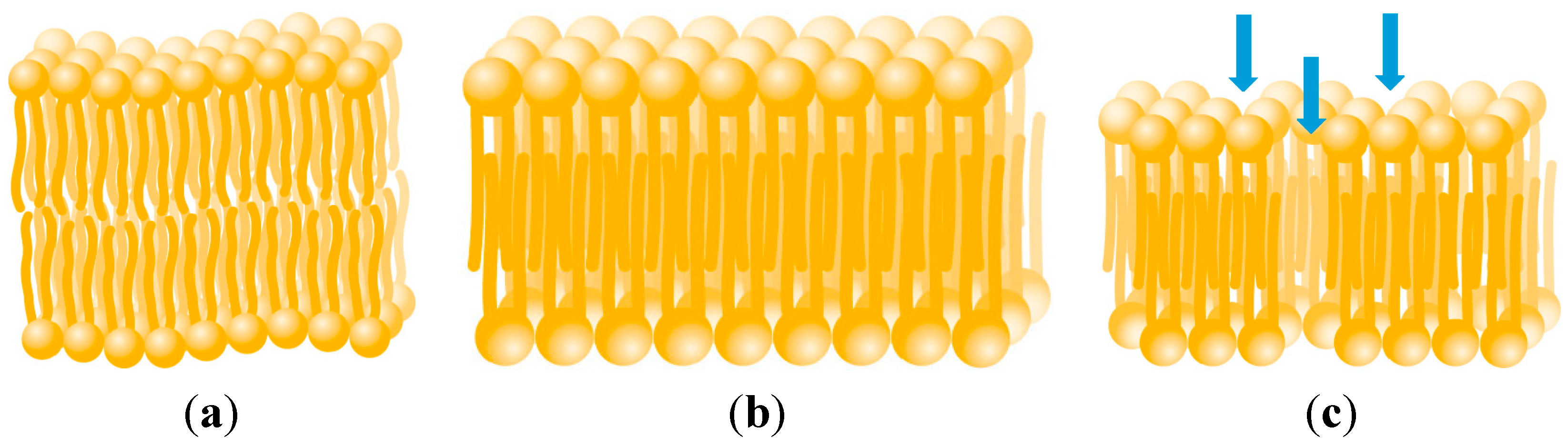
2.3.3. Membrane Permeability


2.3.4. Diffusion
2.4. Evaluation of Transport Pathways
2.4.1. Apoplastic Transport of Cu2+
2.4.2. Symplastic Transport of Cu Chelates
2.4.3. Dual Transport Pathways for Partial Cu Chelation
3. Experimental Section
3.1. Model Conceptualization

3.2. Model Simulations
| Treatment | Ds (×10−6 m2/day) | Da (×10−6 m2/day) | KL (m3/g) | Relative Pm |
|---|---|---|---|---|
| Control | 59 | 31 | 0.004 | 1 |
| Citric acid (0.157 mM) | 49 | 17 | 0.0016 | 0.20 |
| EDTA | 43 | 12 | 0.00016 | 0.006 |
| DTPA | 36 | 9 | 0.00015 | 0.05 |
| Citric acid (2 mM) | 47 | 14 | 0.00058 | – |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix
| Comparison Set | Parameter † | Ccortex | Cendodermis | Cxylem | Cleaf | Translocation |
|---|---|---|---|---|---|---|
| Global Parameters | Da | 0.346 *** | 0.057 *** | 0.175 *** | 0.207 ** | 0.183 * |
| Ds | 0 | 0.004 | 0.004 | 0.035 | 0.035 | |
| vfactor | 0 | 0.001 | 0.019 | 0.001 | 0 | |
| KL | 0.261 *** | 0.076 ** | 0.011 | 0.242* | 0.357 * | |
| α | 0.009 | 0.003 | 0 | 0.006 | 0.006 | |
| Pm | 0.280 *** | 0.823 *** | 0.756 *** | 0.508 *** | 0.416 *** | |
| EDF | 0.105 ** | 0.034 | 0.035 | 0.001 | 0.002 | |
| Parameter Groups | Kpm | 0.444 ** | 0.433 ** | 0.432 ** | 0.325 | 0.325 |
| Hydraulic conductivity: intact endodermis | Kxl | 0.049 | 0.049 | 0.048 | 0.141 | 0.141 |
| Ksym | 0.014 | 0.014 | 0.014 | 0 | 0 | |
| Kcw | 0.01 | 0.01 | 0.01 | 0.012 | 0.012 | |
| Kcb | 0.483 | 0.494 | 0.495 | 0.522 | 0.522 | |
| Hydraulic conductivity: damaged endodermis | Kpm | 0.019 *** | 0.026 | 0.026 | 0.162 | 0.162 |
| Kxl | 0.081 | 0.076 | 0.076 | 0.246 * | 0.244 * | |
| Ksym | 0.058 | 0.064 * | 0.062 | 0.09 | 0.09 | |
| Kcw | 0.000 *** | 0.002 | 0.002 | 0.08 | 0.077 | |
| EDF | 0.841 *** | 0.831 *** | 0.834 *** | 0.422 *** | 0.427 *** | |
| Da | Dacortex | 0.950 *** | 0.931 *** | 0.491 *** | 0.430 ** | 0.183 |
| Daendo | 0.006 | 0.002 | 0.070 * | 0.046 | 0.083 | |
| Daxylem | 0.005 | 0.043 | 0.054 | 0.118 | 0.099 | |
| Daleaf | 0.004 * | 0.007 | 0.009 | 0.036 | 0.061 | |
| Ds | 0.036 ** | 0.017 * | 0.376 ** | 0.370 * | 0.575 * | |
| Ds | Dscortex | 0.004 *** | 0.009 *** | 0.206 *** | 0.158 *** | 0.505 *** |
| Dsendo | 0.090 * | 0.199 | 0.157 | 0.154 | 0.059 | |
| Dsleaf | 0.017 | 0.023 * | 0.000 * | 0 | 0.001 | |
| Dacortex | 0.882 *** | 0.757 *** | 0.634 *** | 0.656 *** | 0.366 *** | |
| Daxylem | 0.007 ** | 0.012 *** | 0.002 ** | 0.033 * | 0.069 | |
| KL | KLcortex | 0.963 *** | 0.905 *** | 0.893 *** | 0.322 *** | 0.678 |
| KLendo | 0.006 | 0.06 | 0.023 | 0.163 | 0.082 | |
| KLxylem | 0.007 | 0.024 | 0.043 *** | 0.227 * | 0.074 ** | |
| KLleaf | 0.005 | 0.003 | 0.039 | 0.037 ** | 0.018 *** | |
| vfactor | 0.02 | 0.008 | 0.002 ** | 0.251 * | 0.147 * | |
| Permeability and dispersivity | Pmembrane | 0.883 *** | 0.931 *** | 0.732 *** | 0.724 *** | 0.734 *** |
| Ptonoplast | 0.005 * | 0.005 | 0.008 | 0.008 * | 0.008 | |
| αcortex | 0.001 | 0.005 | 0.004 | 0.005 * | 0.004 | |
| αxylem | 0.01 | 0.002 | 0.04 | 0.041 ** | 0.039 | |
| αleaf | 0.101 | 0.057 | 0.217 | 0.222 | 0.215 | |
| Specific Parameters | Dacortex | 0.398 *** | 0.115 | 0.046 *** | 0.006 | 0.000 * |
| Dscortex | 0.077 | 0.146 | 0.24 | 0.293 | 0.281 | |
| KLcortex | 0.242 *** | 0.006 | 0.056 | 0.055 | 0.018 * | |
| EDF | 0.02 | 0.001 | 0.000 *** | 0.004 | 0.006 | |
| Pmembrane | 0.263 ** | 0.733 *** | 0.658 *** | 0.643 *** | 0.696 *** |
| Treatment | [Cu2+] | [CuHPO4 (aq)] | [Cu-Citrate−] | [CuEDTA2−] | [CuDTPA3−] + [CuHDTPA2−] + [Cu2DTPA−] |
|---|---|---|---|---|---|
| Control | 68.7 | 20.4 | 1.1 | n.a. | n.a. |
| Citric acid (0.157 mM) | 20.4 | 5.5 | 70.23 | n.a. | n.a. |
| Citric acid (2 mM) | 0.5 | 0.17 | 96.6 | n.a. | n.a. |
| EDTA (0.157 mM) | 0.64 | 0.18 | 0.087 | 98.8 | n.a. |
| DTPA (0.157 mM) | 0.012 | n.a. | n.a. | n.a. | 86.5 + 7.5 + 6 |
| Species | Log Kow | Relative Pm (Kow Basis) * | Log Koc † | Relative Pm (Koc Basis) * | Log KDMPC/water ‡ | Relative Pm (KDMPC/water Basis) * |
|---|---|---|---|---|---|---|
| Cu | −1.11 a | 1.00000 | 0.77 | 1.000 | −1.111 | 1.0000 |
| Citric acid | −1.64 b | 0.28684 | 0.48 | 0.500 | −3.0164 | 0.0121 |
| CuCitric | −2.19 a | 0.06774 | 0.18 | 0.211 | −3.5694 | 0.0029 |
| EDTA | −3.86 c | 0.00157 | −0.72 | 0.028 | −2.052 | 0.1007 |
| CuEDTA | −5.03 a | 0.00009 | −1.36 | 0.006 | −3.225 | 0.0058 |
| DTPA | −4.9 b | – | −1.29 | – | −1.909 | – |
| CuDTPA | −5.22 a | 0.00005 | −1.46 | 0.004 | −2.219 | 0.0496 |
References
- Huang, J.W.; Chen, J.; Berti, W.R.; Cunningham, S.D. Phytoremediation of lead-contaminated soils: Role of synthetic chelates in lead phytoextraction. Environ. Sci. Technol. 1997, 31, 800–805. [Google Scholar] [CrossRef]
- Chiu, K.K.; Ye, Z.H.; Wong, M.H. Enhanced uptake of As, Zn, and Cu by Vetiveria zizanioides and Zea mays using chelating agents. Chemosphere 2005, 60, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Evangelou, M.W.H.; Ebel, M.; Schaeffer, A. Chelate assisted phytoextraction of heavy metals from soil. Effect, mechanism, toxicity, and fate of chelating agents. Chemosphere 2007, 68, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Kos, B.; Lestan, D. Chelator induced phytoextraction and in situ soil washing of Cu. Environ. Pollut. 2004, 132, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, M.D.; Leung, D.W.M. Chelated lead transport in Pinus radiata: An ultrastructural study. Environ. Exp. Bot. 2002, 48, 21–32. [Google Scholar] [CrossRef]
- Blaylock, M.J.; Dushenkov, S.; Zakharova, O.; Gussman, C.; Kapulnik, Y.; Ensley, B.D.; Salt, D.E.; Raskin, I. Enhanced accumulation of Pb in Indian mustard by soil-applied chelating agents. Environ. Sci. Technol. 1997, 31, 860–865. [Google Scholar] [CrossRef]
- Johnson, A.C.; Singhal, N. Influence of chelation on Cu distribution and barriers to translocation in Lolium perenne. Environ. Sci. Technol. 2013, 47, 7688–7695. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, M.; Lanzirotti, A. 241Am, 137Cs, Sr and Pb uptake by tobacco as influenced by application of Fe chelators to soil. J. Environ. Radioact. 2005, 82, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Vassil, A.D.; Kapulnik, Y.; Raskin, I.; Salt, D.E. The role of EDTA in lead transport and accumulation by Indian mustard. Plant Physiol. 1998, 117, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Turgut, C.; Pepe, M.K.; Cutright, T.J. The effect of EDTA and citric acid on phytoremediation of Cd, Cr, and Ni from soil using Helianthus annuus. Environ. Pollut. 2004, 131, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Tandy, S.; Schulin, R.; Nowack, B. The influence of EDDS on the uptake of heavy metals in hydroponically grown sunflowers. Chemosphere 2006, 62, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Shen, Z.; Luo, C.; Deng, Y.-E.; Wang, C. Accumulation mechanisms and subcellular distribution of Cu in maize grown on soil treated with [S,S]-ethylenediamine disuccinic acid. Plant Soil 2012, 351, 237–247. [Google Scholar] [CrossRef]
- Luo, C.L.; Shen, Z.G.; Li, X.D. Hot NTA application enhanced metal phytoextraction from contaminated soil. Water Air Soil Pollut. 2008, 188, 127–137. [Google Scholar] [CrossRef]
- Steudle, E.; Frensch, J. Water transport in plants: Role of the apoplast. Plant Soil 1996, 187, 67–79. [Google Scholar] [CrossRef]
- Luo, C.; Shen, Z.; Li, X.; Baker, A.J.M. The role of root damage in the chelate-enhanced accumulation of lead by Indian mustard plants. Int. J. Phytoremediat. 2006, 8, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.; Schulin, R.; Nowack, B.; Roulier, S.; Menon, M.; Clothier, B.; Green, S.; Mills, T. Phytoremediation for the management of metal flux in contaminated sites. Snow Landsc. Res. 2006, 80, 221–234. [Google Scholar]
- Novak, V.; Vidovic, J. Transpiration and nutrient uptake dynamics in maize (Zea mays L.). Ecol. Model. 2003, 166, 99–107. [Google Scholar] [CrossRef]
- Sawalha, M.F.; Peralta-Videa, J.R.; Romero-González, J.; Gardea-Torresdey, J. Biosorption of Cd(II), Cr(III), and Cr(VI) by saltbush (Atriplex canescens) biomass: Thermodynamic and isotherm studies. J. Colloid Interface Sci. 2006, 300, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Zhang, H.; Zhang, Q.; Jin, G.M.; Jiang, S.J.; Jiang, D.; He, Q.Y.; Li, Z.P. Copper-induced oxidative stress and responses of the antioxidant system in roots of Medicago sativa. J. Agron. Crop Sci. 2011, 197, 418–429. [Google Scholar] [CrossRef]
- Sibgatullin, T.; Vergeldt, F.; Gerkema, E.; van As, H. Quantitative permeability imaging of plant tissues. Eur. Biophys. J. 2010, 39, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Trapp, S. Modelling uptake into roots and subsequent translocation of neutral and ionisable organic compounds. Pest Manag. Sci. 2000, 56, 767–778. [Google Scholar] [CrossRef]
- Bushey, J.T.; Ebbs, S.D.; Dzombak, D.A. Development of a plant uptake model for cyanide. Int. J. Phytoremediat. 2006, 8, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Schoups, G.; Hopmans, J.W. Analytical model for vadose zone solute transport with root water and solute uptake. Vadose Zone J. 2002, 1, 158–171. [Google Scholar] [CrossRef]
- Sung, K.; Corapcioglu, M.Y.; Drew, M.C. Plant aided bioremediation in the vadose zone: Model development and applications. J. Contam. Hydrol. 2004, 73, 65–98. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y. Modeling the mechanisms for uptake and translocation of dioxane in a soil-plant ecosystem with STELLA. J. Contam. Hydrol. 2008, 95, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Maxted, A.P.; Black, C.R.; West, H.M.; Crout, N.M.J.; McGrath, S.P.; Young, S.D. Phytoextraction of cadmium and zinc from arable soils amended with sewage sludge using Thlaspi caerulescens: Development of a predictive model. Environ. Pollut. 2007, 150, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Kinraide, T.B. Ion fluxes considered in terms of membrane-surface electrical potentials. Aust. J. Plant Physiol. 2001, 28, 605–616. [Google Scholar]
- Somma, F.; Hopmans, J.W.; Clausnitzer, V. Transient three-dimensional modeling of soil water and solute transport with simultaneous root growth, root water and nutrient uptake. Plant Soil 1998, 202, 281–293. [Google Scholar] [CrossRef]
- Palm, V. A model for sorption, flux and plant uptake of cadmium in a soil profile: Model structure and sensitivity analysis. Water Air Soil Pollut. 1994, 77, 169–190. [Google Scholar]
- Skaggs, T.H.; Jarvis, N.J.; Pontedeiro, E.M.; van Genuchten, M.T.; Cotta, R.M. Analytical advection-dispersion model for transport and plant uptake of contaminants in the root zone. Vadose Zone J. 2007, 6, 890–898. [Google Scholar] [CrossRef]
- Thornton, B.; Macklon, A.E.S. Copper uptake by ryegrass seedlings; contribution of cell wall adsorption. J. Exp. Bot. 1989, 40, 1105–1111. [Google Scholar] [CrossRef]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology, 3th ed.; Elsevier Academic Press: San Diego, CA, USA, 2005; p. 567. [Google Scholar]
- Raven, J. Short and long distance transport of boric acid in plants. New Phytol. 1980, 84, 231–249. [Google Scholar] [CrossRef]
- Dordas, C.; Brown, P.H. Permeability of boric acid across lipid bilayers and factors affecting it. J. Membr. Biol. 2000, 175, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Steudle, E.; Tyerman, S.D. Determination of permeability coefficients, reflection coefficients, and hydraulic conductivity of Chara corallina using the pressure probe: Effects of solute concentrations. J. Membr. Biol. 1983, 75, 85–96. [Google Scholar] [CrossRef]
- Jeffries, R.L. The ionic relations of the seedlings of the halophyte, Triglochin maritima L. In Ion Transport in Plants; Anderson, W.P., Ed.; Academic Press: London, UK, 1973. [Google Scholar]
- Hope, A.B.; Walker, N.A. Ionic relations of cells of Chara australis. IV. Membrane potential differences and resistances. Aust. J. Biol. Sci. 1961, 14, 26–44. [Google Scholar]
- Diamond, J.M.; Solomon, A.K. Intracellular potassium compartments in Nitella axillaris. J. Gen. Physiol. 1959, 42, 1105–1121. [Google Scholar] [CrossRef] [PubMed]
- MacRobbie, E.A.C.; Dainty, J. Ion transport in Nitellopsis obtusa. J. Gen. Physiol. 1958, 42, 335–353. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, R.J. Energetic considerations of ion transport in Enteromorpha intestinalis (L.) Link. New Phytol. 1985, 100, 5–24. [Google Scholar] [CrossRef]
- Dainty, J. Ion transport and electrical potentials in plant cells. Annu. Rev. Plant Physiol. 1962, 13, 379–402. [Google Scholar] [CrossRef]
- Scott, B.; Gulline, H.; Pallaghy, C.K. The electrochemical state of cells of broad bean root. I. Investigations of elongating roots of young seedlings. Aust. J. Biol. Sci. 1968, 21, 185–200. [Google Scholar]
- Macklon, A.E.S. Cortical cell fluxes and transport to the stele in excised root segments of Allium cepa L. I. Potassium, sodium and chloride. Planta 1975, 122, 109–130. [Google Scholar] [CrossRef] [PubMed]
- Macklon, A.E.S. Cortical cell fluxes and transport to the stele in excised root segments of Allium cepa L. II. Calcium. Planta 1975, 122, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Gunawardana, W.B.; Singhal, N.; Johnson, A. Amendments and their combined application for enhanced copper, cadmium, lead uptake by Lolium perenne. Plant Soil 2010, 329, 283–294. [Google Scholar] [CrossRef]
- Calucci, L.; Pinzino, C.; Quartacci, M.F.; Navari-Izzo, F. Copper excess reduces the fluidity of plasma membrane lipids of wheat roots: A spin probe EPR study. J. Phys. Chem. B 2003, 107, 12021–12028. [Google Scholar] [CrossRef]
- Berglund, A.H.; Quartacci, M.F.; Calucci, L.; Navari-Izzo, F.; Pinzino, C.; Liljenberg, C. Alterations of wheat root plasma membrane lipid composition induced by copper stress result in changed physicochemical properties of plasma membrane lipid vesicles. Biochim. Biophys. Acta Biomembr. 2002, 1564, 466–472. [Google Scholar] [CrossRef]
- Lande, M.B.; Donovan, J.M.; Zeidel, M.L. The relationship between membrane fluidity and permeabilities to water, solutes, ammonia, and protons. J. Gen. Physiol. 1995, 106, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Wagatsuma, T. Plasma membrane permeability of root-tip cells following temporary exposure to Al ions is a rapid measure of Al tolerance among plant species. Plant Cell Physiol. 1998, 39, 516–525. [Google Scholar] [CrossRef]
- Azenha, M.; Vasconcelos, M.T.; Cabral, J.P.S. Organic ligands reduce copper toxicity in Pseudomonas syringae. Environ. Toxicol. Chem. 1995, 14, 369–373. [Google Scholar] [CrossRef]
- Suhayda, C.G.; Haug, A. Organic acids reduce aluminum toxicity in maize root membranes. Physiol. Plant. 1986, 68, 189–195. [Google Scholar] [CrossRef]
- Pellet, D.M.; Grunes, D.L.; Kochian, L.V. Organic acid exudation as an aluminum-tolerance mechanism in maize (Zea mays L.). Planta 1995, 196, 788–795. [Google Scholar] [CrossRef]
- Zhao, X.-J.; Sucoff, E.I.; Stadelmann, E.J. Al3+ and Ca2+ alteration of membrane permeability of Quercus rubra root cortex cells. Plant Physiol. 1987, 83, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Sucoff, E.I.; Stadelmann, E.J. Aluminum and temperature alteration of cell membrane permeability of Quercus rubra. Plant Physiol. 1991, 96, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Sokolik, A.; Yurin, V. Characteristics of non-specific permeability and H+-ATPase inhibition induced in the plasma membrane of Nitella flexilis by excessive Cu2+. Planta 2001, 212, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Rojanasakul, Y.; Robinson, J.R. The cytoskeleton of the cornea and its role in tight junction permeability. Int. J. Pharm. 1991, 68, 135–149. [Google Scholar] [CrossRef]
- Kikuchi, T.; Suzuki, M.; Kusai, A.; Iseki, K.; Sasaki, H. Synergistic effect of EDTA and boric acid on corneal penetration of CS-088. Int. J. Pharm. 2005, 290, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Suzuki, M.; Kusai, A.; Iseki, K.; Sasaki, H.; Nakashima, K. Mechanism of permeability-enhancing effect of EDTA and boric acid on the corneal penetration of 4-[1-hydroxy-1-methylethyl]-2-propyl-1-[4-[2-[tetrazole-5-yl]phenyl]phenyl]methylimidazole-5-carboxylic acid monohydrate (CS-088). Int. J. Pharm. 2005, 299, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Kaszuba, M.; Hunt, G.R.A. Protection against membrane damage: A 1H-NMR investigation of the effect of Zn2+ and Ca2+ on the permeability of phospholipid vesicles. J. Inorg. Biochem. 1990, 40, 217–225. [Google Scholar] [CrossRef]
- Pasternak, C. A novel role of Ca2+ and Zn2+: Protection of cells against membrane damage. Biosci. Rep. 1988, 8, 578–583. [Google Scholar] [CrossRef]
- Pasternak, C. A novel form of host defence: Membrane protection by Ca2+ and Zn2+. Biosci. Rep. 1987, 7, 81–91. [Google Scholar] [PubMed]
- Amro, N.; Kotra, L.; Mesthrige, K.; Bulychev, A.; Mobashery, S.; Liu, G. High resolution atomic force microscopy studies of the Escherichia coli outer membrane: Structural basis for permeability. Langmuir 2000, 16, 2789–2796. [Google Scholar] [CrossRef]
- Leive, L. Release of lipopolysaccharide by EDTA treatment of E. coli. Biochem. Biophys. Res. Commun. 1965, 21, 290–296. [Google Scholar] [CrossRef]
- Nikaido, H.; Vaara, M. Molecular basis of bacterial outer membrane permeability. Microbiol. Rev. 1985, 49, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Geebelen, W.; Vangrosveld, J.; Adriano, D.C.; van Poucke, L.C.; Clijsters, H. Effects of Pb–EDTA and EDTA on oxidative stress reactions and mineral uptake in Phaseolus vulgaris. Physiol. Plant. 2002, 115, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Prachayasittikul, V.; Isarankura-Na-Ayudhya, C.; Tantimongcolwat, T.; Nantasenamat, C.; Galla, H.-J. EDTA-induced membrane fluidization and destabilization: Biophysical studies on artificial lipid membranes. Acta Biochim. Biophys. Sin. 2007, 39, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Alakomi, H.L.; Paananen, A.; Suihko, M.L.; Helander, I.M.; Saarela, M. Weakening effect of cell permeabilizers on gram-negative bacteria causing biodeterioration. Appl. Environ. Microbiol. 2006, 72, 4695–4703. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Wong, P.G.W. Compounds which increase the permeability of the Pseudomonas aeruginosa outer membrane. Antimicrob. Agents Chemother. 1984, 26, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Li, L.; Hu, X.; Wang, Z.; Zhao, G. Increase in the permeability of tonoplast of garlic (Allium sativum) by monocarboxylic acids. J. Agric. Food Chem. 2006, 54, 8103–8107. [Google Scholar] [CrossRef] [PubMed]
- Oparka, K.J.; Prior, D.A.M.; Crawford, J.W. Behaviour of plasma membrane, cortical ER and plasmodesmata during plasmolysis of onion epidermal cells. Plant Cell Environ. 1994, 17, 163–171. [Google Scholar] [CrossRef]
- Jarvis, M.D.; Leung, D.W.M. Chelated lead transport in Chamaecytisus proliferus (L.f.) link ssp. proliferus var. palmensis (H. Christ): An ultrastructural study. Plant Sci. 2001, 161, 433–441. [Google Scholar] [CrossRef]
- Niu, L.; Shen, Z.; Wang, C. Sites, pathways, and mechanism of absorption of Cu-EDDS complex in primary roots of maize (Zea Mays L.): Anatomical, chemical and histochemical analysis. Plant Soil 2011, 343, 303–312. [Google Scholar] [CrossRef]
- Hopmans, J.W.; Bristow, K.L. Current capabilities and future needs of root water and nutrient uptake modeling. Adv. Agron. 2002, 77, 103–175. [Google Scholar]
- Dixon, H.H.; Joly, J. On the ascent of sap. Philos. Trans. R. Soc. London Ser. B 1895, 186, 563–576. [Google Scholar] [CrossRef]
- Tsuda, M.; Tyree, M.T. Plant hydraulic conductance measured by the high pressure flow meter in crop plants. J. Exp. Bot. 2000, 51, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Aumann, C.A.; Ford, E.D. Modeling tree water flow as an unsaturated flow through a porous medium. J. Theor. Biol. 2002, 219, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Früh, T.; Kurth, W. The hydraulic system of trees: Theoretical framework and numerical simulations. J. Theor. Biol. 1999, 201, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Joly, R.J. Effects of sodium chloride on the hydraulic conductivity of soybean root systems. Plant Physiol. 1989, 91, 1262–1265. [Google Scholar] [CrossRef] [PubMed]
- Roose, T.; Fowler, A.C. A mathematical model for water and nutrient uptake by plant root systems. J. Theor. Biol. 2004, 228, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Nye, P.H.; Brewster, J.L.; Bhat, K.K.S. The possibility of predicting solute uptake and plant growth response from independently measured soil and plant characteristics. I. The theoretical basis of the experiments. Plant Soil 1975, 42, 161–170. [Google Scholar]
- Albrecht, A.; Schultze, U.; Liedgens, M.; Fluhler, H.; Frossard, E. Incorporating soil structure and root distribution into plant uptake models for radionuclides: Toward a more physically based transfer model. J. Environ. Radioact. 2002, 59, 329–350. [Google Scholar] [PubMed]
- Wiegers, B.S.; Cheer, A.Y.; Silk, W.K. Modeling the hydraulics of root growth in three dimensions with phloem water sources. Plant Physiol. 2009, 150, 2092–2103. [Google Scholar] [CrossRef] [PubMed]
- Schnepf, A.; Schrefl, T.; Wenzel, W.W. The suitability of pde-solvers in rhizosphere modelling, exemplified by three mechanistic rhizosphere models. J. Plant Nutr. Soil Sci. 2002, 165, 713–718. [Google Scholar] [CrossRef]
- Lehto, N.; Davison, W.; Zhang, H.; Tych, W. Analysis of micro-nutrient behaviour in the rhizosphere using a DGT parameterised dynamic plant uptake model. Plant Soil 2006, 282, 227–238. [Google Scholar] [CrossRef]
- Puschenreiter, M.; Schnepf, A.; Molina Millán, I.; Fitz, W.J.; Horak, O.; Klepp, J.; Schrefl, T.; Lombi, E.; Wenzel, W.W. Changes of Ni biogeochemistry in the rhizosphere of the hyperaccumulator Thlaspi goesingense. Plant Soil 2005, 271, 205–218. [Google Scholar] [CrossRef]
- Johnson, A.C. Modelling Chelate Enhanced Phytoextraction Mechanisms. Ph.D. Thesis, The University of Auckland, Auckland, New Zealand, 2012. [Google Scholar]
- Bell, P.F.; Chaney, R.L.; Angle, J.S. Free metal activity and total metal concentrations as indices of micronutrient availability to barley [Hordeum vulgare (L.) “Klages”]. Plant Soil 1991, 130, 51–62. [Google Scholar] [CrossRef]
- Lyman, W.J.; Reehl, W.F.; Rosenblatt, D.H. Handbook of Chemical Property Estimation Methods: Environmental Behavior of Organic Compounds; American Chemical Society: Washington, DC, USA, 1990; p. 530. [Google Scholar]
- Vaes, W.H.J.; Urrestarazu Ramos, E.A.; Verhaar, H.J.M.; Cramer, C.J.; Hermens, J.L.M. Understanding and estimating membrane/water partition coefficients: Approaches to derive quantitative structure property relationships. Chem. Res. Toxicol. 1998, 11, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Avdeef, A.; Box, K.; Comer, J.; Hibbert, C.; Tam, K. Measuring log P using liposomes; a possible tool to study drug-membrane interactions. J. Pharm. Pharmacol. 1997, 49, 203. [Google Scholar]
- Meylan, W.M.; Howard, P.H. Atom/fragment contribution method for estimating octanol-water partition coefficients. J. Pharm. Sci. 1995, 84, 83–92. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, A.; Singhal, N. Increased Uptake of Chelated Copper Ions by Lolium perenne Attributed to Amplified Membrane and Endodermal Damage. Int. J. Mol. Sci. 2015, 16, 25264-25284. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161025264
Johnson A, Singhal N. Increased Uptake of Chelated Copper Ions by Lolium perenne Attributed to Amplified Membrane and Endodermal Damage. International Journal of Molecular Sciences. 2015; 16(10):25264-25284. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161025264
Chicago/Turabian StyleJohnson, Anthea, and Naresh Singhal. 2015. "Increased Uptake of Chelated Copper Ions by Lolium perenne Attributed to Amplified Membrane and Endodermal Damage" International Journal of Molecular Sciences 16, no. 10: 25264-25284. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161025264