
Heat Killed Lactobacillus reuteri GMNL-263 Reduces Fibrosis Effects on the Liver and Heart in High Fat Diet-Hamsters via TGF-β Suppression

 ,
,
Abstract
:
1. Introduction
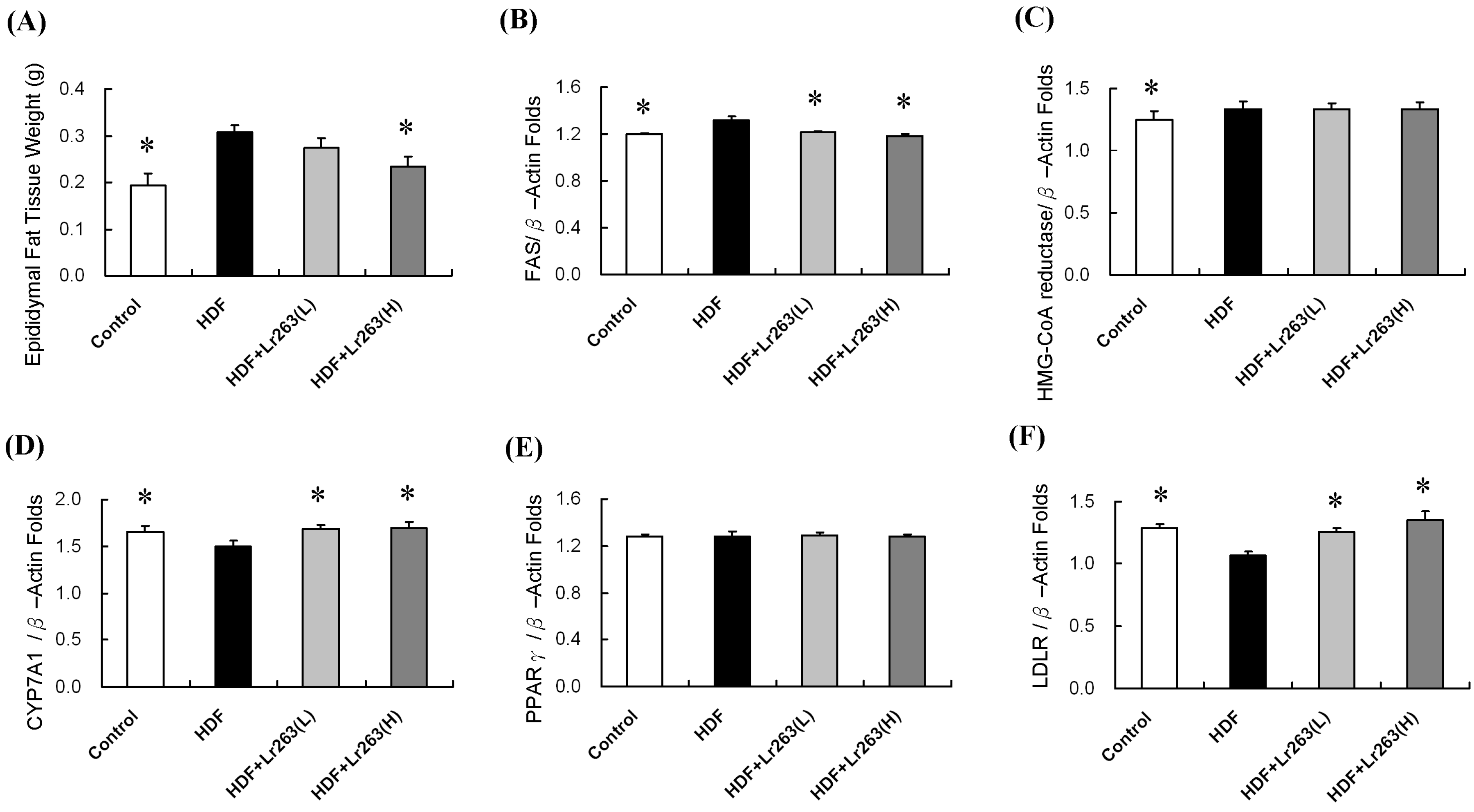
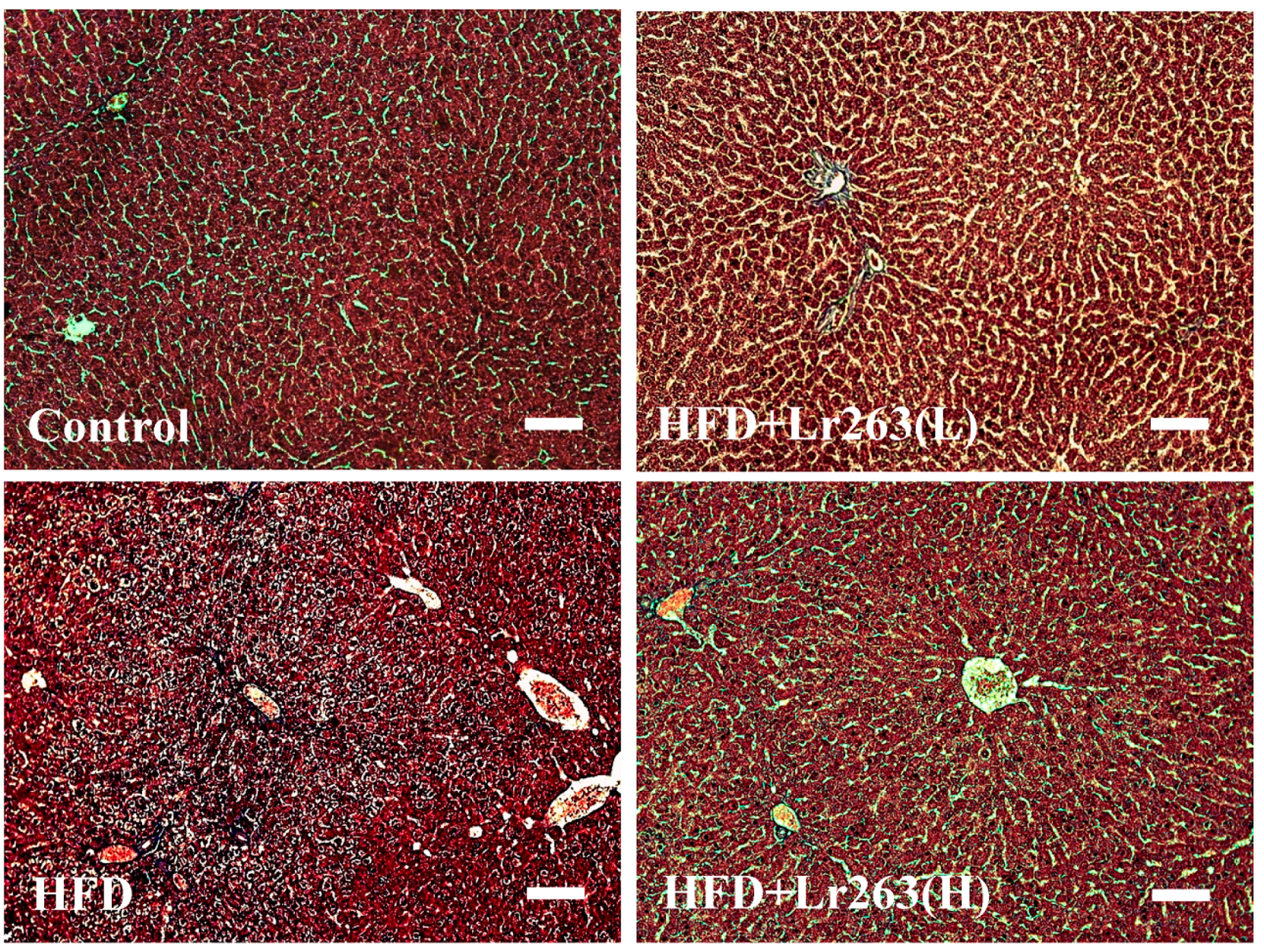
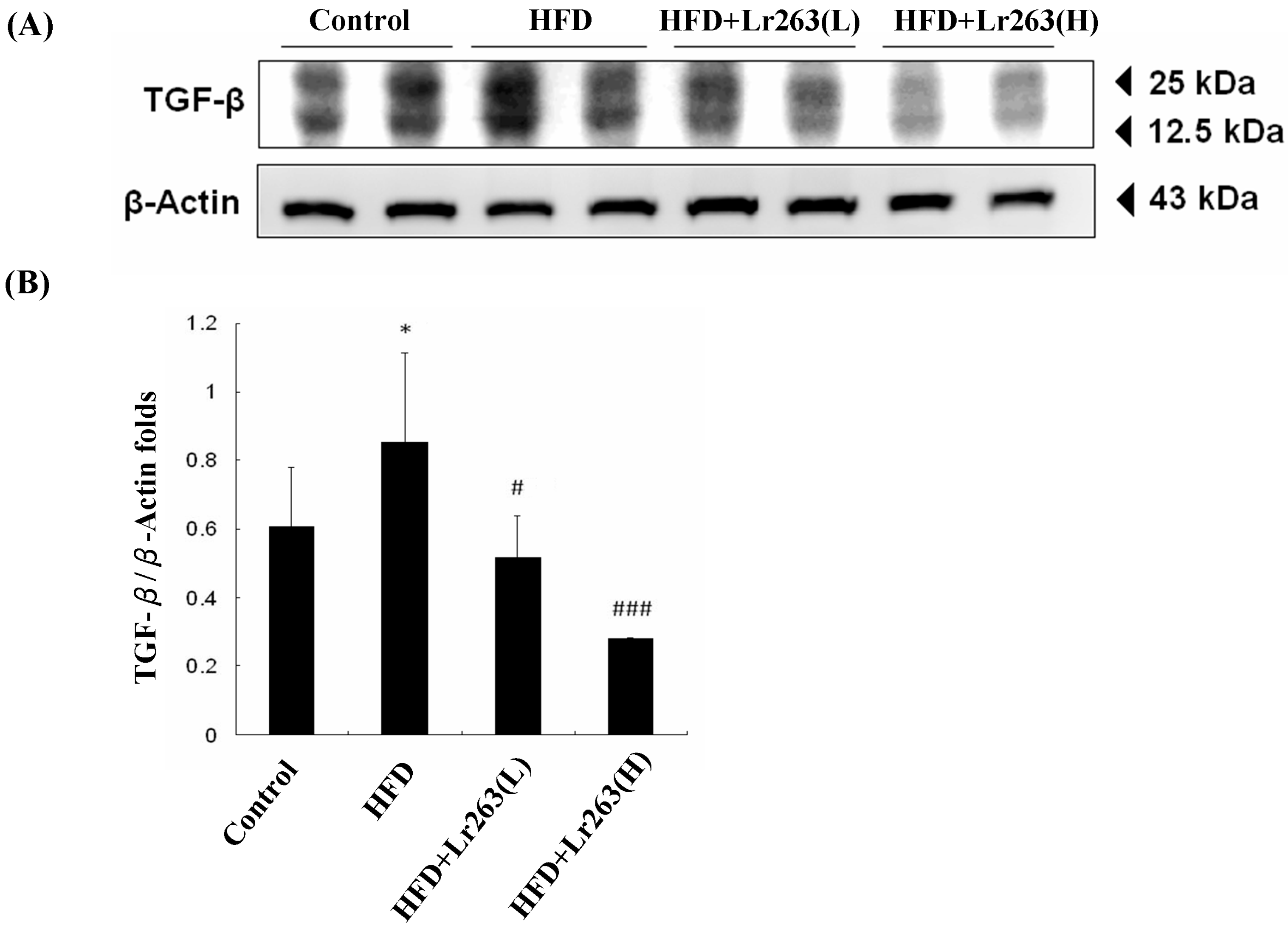
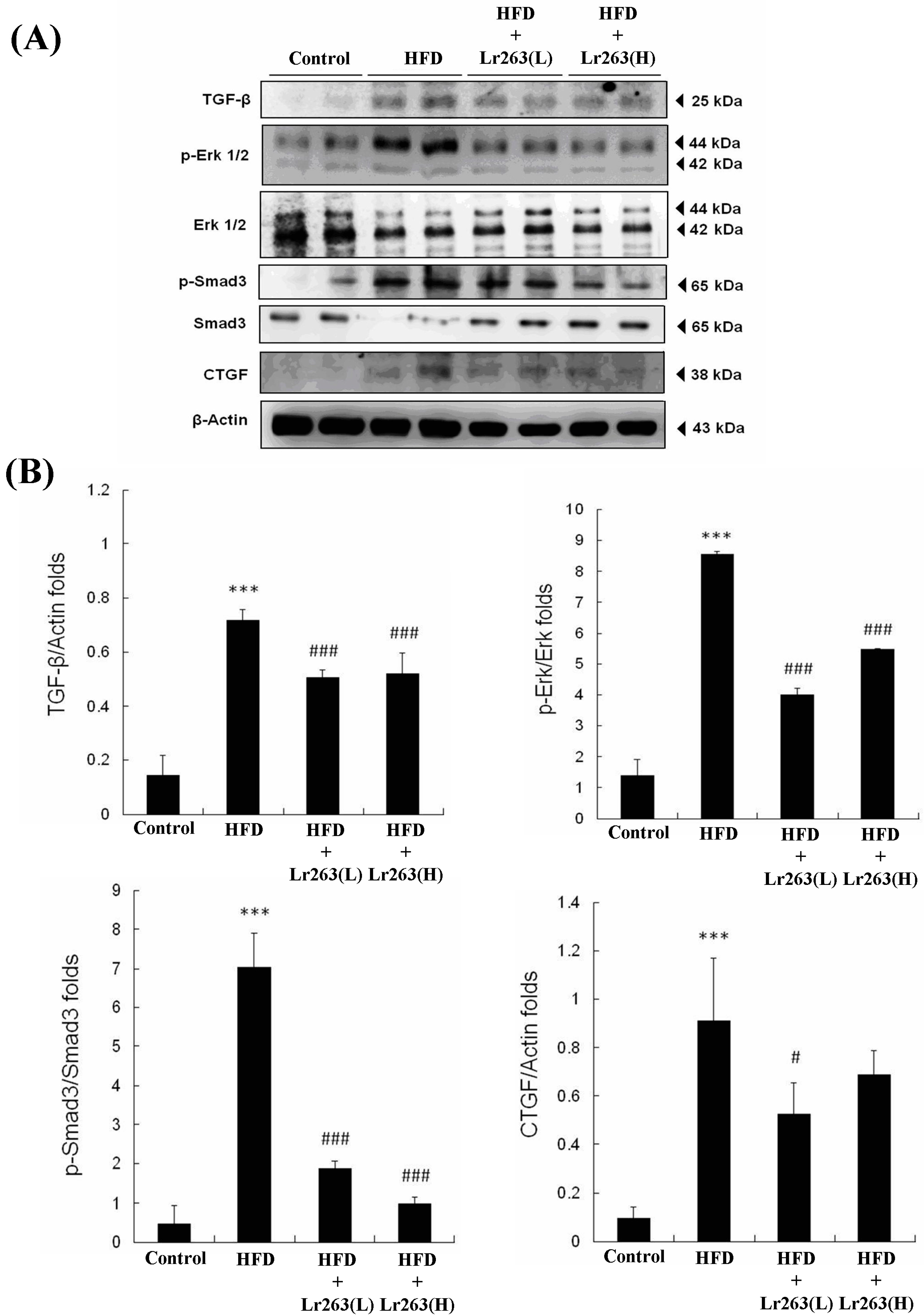
2. Results and Discussion




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Control | HFD | HFD + Lr-263(L) | HFD + Lr-263(H) |
|---|---|---|---|---|
| Parameter | ||||
| Body Weight (g) | 126.5 ± 10.6 * | 138.3 ± 5.9 | 136.7 ± 3.8 | 124.7 ± 4.0 * |
| Food Intake (g) | 8.1 ± 0.8 | 8.2 ± 0.9 | 7.9 ± 1.1 | 7.8 ± 0.5 |
| Liver | ||||
| TG (mg/g) | 85.0 ± 6.5 * | 93.3 ± 5.3 | 90.7 ± 3.4 | 84.0 ± 4.3 * |
| T-CHO (mg/g) | 89.7 ± 0.4 * | 130.0 ± 4.9 | 118.3 ± 12.8 | 127.3 ± 6.0 |
| MDA (μg/g) | 4.2 ± 0.4 * | 8.6 ± 0.7 | 6.7 ± 1.1 | 5.5 ± 0.4 * |
| Fecal | ||||
| TG (mg/g) | 5.9 ± 0.7 | 4.7 ± 0.8 | 7.6 ± 0.9 * | 10.4 ± 1.7 * |
| T-CHO (mg/g) | 6.7 ± 0.7 | 8.5 ± 1.4 * | 8.5 ± 1.4 | 11.0 ± 0.8 * |



3. Experimental Section
3.1. Preparing the Probiotic Suspensions
3.2. Animals
| Components (g/kg) | Normal Diet | High-Fat Diet |
|---|---|---|
| Casein | 200 | 232 |
| l-Cystine | 3.0 | 3.0 |
| d,l-Methionine | N.D. | 3.5 |
| Corn Starch | 397.48 | 137 |
| Maltodextrin | 132 | 150 |
| Sucrose | 100 | 162.58 |
| Cellulose | 50 | 50 |
| Cholesterol | N.D. | 1.9 |
| Mineral Mix (AIN-93) | 35 | 40.60 |
| Calcium phosphate dibasic | N.D. | 4.64 |
| Vitamin Mix (AIN-93) | 10 | 16.24 |
| Choline Bitartrate | 2.5 | 5 |
| Tert-butylhydroquinone | 0.014 | 0.04 |
| Soybean oil | 70 | 40 |
| Lard | N.D. | 153.5 |
3.3. Cardiac Echocardiography
3.4. Masson’s Trichrome Staining
3.5. RNA Extraction and RT-PCR
3.6. Tissue Protein Extraction
3.7. Western Blot Assay
3.8. Liver and Fecal Lipid Profile Analysis
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Casavalle, P.L.; Lifshitz, F.; Romano, L.S.; Pandolfo, M.; Caamaño, A.; Boyer, P.M.; Rodríguez, P.N.; Friedman, S.M. Prevalence of dyslipidemia and metabolic syndrome risk factor in overweight and obese children. Pediatr. Endocrinol. Rev. 2014, 12, 213–223. [Google Scholar] [PubMed]
- Cortez, M.; Singleton, J.R.; Smith, A.G. Glucose intolerance, metabolic syndrome, and neuropathy. Handb. Clin. Neurol. 2014, 126, 109–122. [Google Scholar] [PubMed]
- El-Bassossy, H.M.; Shaltout, H.A. Allopurinol alleviates hypertension and proteinuria in high fructose, high salt and high fat induced model of metabolic syndrome. Transl. Res. 2014, 165, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Geliebter, A.; Ochner, C.N.; Dambkowski, C.L.; Hashim, S.A. Obesity-related hormones and metabolic risk factors: A randomized trial of diet plus either strength or aerobic training versus diet alone in overweight participants. J. Diabetes Obes. 2014, 1, 1–7. [Google Scholar] [PubMed]
- Suh, S.; Baek, J.; Bae, J.C.; Kim, K.N.; Park, M.K.; Kim, D.K.; Cho, N.H.; Lee, M.K. Sex factors in the metabolic syndrome as a predictor of cardiovascular disease. Endocrinol. Metab. 2014, 29, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Moh, M.C.; Sum, C.F.; Lam, B.C.; Ng, X.W.; Su, C.; Tavintharan, S.; Yeoh, L.Y.; Wong, M.D.; Lee, S.B.; Tang, W.E.; et al. Evaluation of body adiposity index as a predictor of aortic stiffness in multi-ethnic Asian population with type 2 diabetes. Diabetes Vasc. Dis. Res. 2014, 12, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Ruff, R.R.; Akhund, A.; Adjoian, T.; Kansagra, S.M. Calorie intake, sugar-sweetened beverage consumption, and obesity among New York City adults: Findings from a 2013 population study using dietary recalls. J. Community Health. 2014, 39, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.C.; Lee, L.T.; Chen, C.Y.; Sung, P.K. All-cause and cardiovascular disease mortality increased with metabolic syndrome in Taiwanese. Obesity 2008, 16, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Q.; Ding, J.; Zhao, H.; Liu, C.M. Puerarin attenuates carbon tetrachloride-induced liver oxidative stress and hyperlipidaemia in mouse by JNK/c-Jun/CYP7A1 pathway. Basic Clin. Pharmacol. Toxicol. 2014, 115, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Teratani, T.; Suzuki, T.; Shimizu, M.; Sato, H.; Narimatsu, K.; Usui, S.; Furuhashi, H.; Kimura, A.; Nishiyama, K.; et al. Acyl-CoA: Cholesterol acyltransferase 1 mediates liver fibrosis by regulating free cholesterol accumulation in hepatic stellate cells. J. Hepatol. 2014, 61, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Fede, G.; Privitera, G.; Tomaselli, T.; Spadaro, L.; Purrello, F. Cardiovascular dysfunction in patients with liver cirrhosis. Ann. Gastroenterol. 2015, 28, 31–40. [Google Scholar] [PubMed]
- Yang, C.H.; Ting, W.J.; Day, C.H.; Ju, D.T.; Yeh, Y.L.; Chung, L.C.; Tsai, F.J.; Tsai, C.H.; Tsai, Y.; Huang, C.Y. SHSST cyclodextrin complex prevents the fibrosis effect on CCl₄-induced cirrhotic cardiomyopathy in rats through TGF-β pathway inhibition effects. Int. J. Mol. Sci. 2014, 15, 8037–8048. [Google Scholar] [CrossRef] [PubMed]
- Owens, A.P., III; Passam, F.H.; Antoniak, S.; Marshall, S.M.; McDaniel, A.L.; Rudel, L.; Williams, J.C.; Hubbard, B.K.; Dutton, J.A.; Wang, J.; et al. Monocyte tissue factor-dependent activation of coagulation in hypercholesterolemic mice and monkeys is inhibited by simvastatin. J. Clin. Investig. 2012, 122, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Ramspoth, T.; Roehl, A.B.; Macko, S.; Heidenhain, C.; Junge, K.; Binnebösel, M.; Schmeding, M.; Neumann, U.P.; Rossaint, R.; Hein, M. Risk factors for coagulopathy after liver resection. J. Clin. Anesth. 2014, 26, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Dobaczewski, M.; Chen, W.; Frangogiannis, N.G. Transforming growth factor (TGF)-β signaling in cardiac remodeling. J. Mol. Cell. Cardiol. 2011, 51, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Leask, A. Potential therapeutic targets for cardiac fibrosis: TGFβ, angiotensin, endothelin, CCN2, and PDGF, partners in fibroblast activation. Circ. Res. 2010, 106, 1675–1680. [Google Scholar] [CrossRef] [PubMed]
- Dobaczewski, M.; Bujak, M.; Li, N.; Gonzalez-Quesada, C.; Mendoza, L.H.; Wang, X.F.; Frangogiannis, N.G. Smad3 signaling critically regulates fibroblast phenotype and function in healing myocardial infarction. Circ. Res. 2010, 107, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Panek, A.N.; Posch, M.G.; Alenina, N.; Ghadge, S.K.; Erdmann, B.; Popova, E.; Perrot, A.; Geier, C.; Dietz, R.; Morano, I.; et al. Connective tissue growth factor overexpression in cardiomyocytes promotes cardiac hypertrophy and protection against pressure overload. PLoS ONE 2009, 4, e6743. [Google Scholar] [PubMed]
- Ettinger, G.; MacDonald, K.; Reid, G.; Burton, J.P. The influence of the human microbiome and probiotics on cardiovascular health. Gut Microbes 2014, 5, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.P.; Hsieh, Y.M.; Kuo, W.W.; Lin, Y.M.; Yeh, Y.L.; Lin, C.C.; Tsai, F.J.; Tsai, C.H.; Huang, C.Y.; Tsai, C.C. Probiotic-fermented purple sweet potato yogurt activates compensatory IGF-IR/PI3K/Akt survival pathways and attenuates cardiac apoptosis in the hearts of spontaneously hypertensive rats. Int. J. Mol. Med. 2013, 32, 1319–1328. [Google Scholar] [PubMed]
- Wang, H.F.; Lin, P.P.; Chen, C.H.; Yeh, Y.L.; Huang, C.C.; Huang, C.Y.; Tsai, C.C. Effects of lactic acid bacteria on cardiac apoptosis are mediated by activation of the phosphatidylinositol-3 kinase/AKT survival-signalling pathway in rats fed a high-fat diet. Int. J. Mol. Med. 2015, 35, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E. Probiotics: Definition, sources, selection, and uses. Clin. Infect. Dis. 2008, 2, S58–S61. [Google Scholar] [CrossRef] [PubMed]
- Elkins, C.A.; Moser, S.A.; Savage, D.C. Genes encoding bile salt hydrolases and conjugated bile salt transporters in Lactobacillus johnsonii 100-100 and other Lactobacillus species. Microbiology 2001, 147, 3403–3412. [Google Scholar] [CrossRef] [PubMed]
- Joyce, S.A.; MacSharry, J.; Casey, P.G.; Kinsella, M.; Murphy, E.F.; Shanahan, F.; Hill, C.; Gahan, C.G. Regulation of host weight gain and lipid metabolism by bacterial bile acid modification in the gut. Proc. Natl. Acad. Sci. USA 2014, 111, 7421–7426. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp. Diabetes Res. 2012, 2012, 902917. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.C.; Lin, P.P.; Hsieh, Y.M.; Zhang, Z.Y.; Wu, H.C.; Huang, C.C. Cholesterol-lowering potentials of lactic Acid bacteria based on bile-salt hydrolase activity and effect of potent strains on cholesterol metabolism in vitro and in vivo. Sci. World J. 2014, 2014, 690752. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, F.C.; Lee, C.L.; Chai, C.Y.; Chen, W.T.; Lu, Y.C.; Wu, C.S. Oral administration of Lactobacillus reuteri GMNL-263 improves insulin resistance and ameliorates hepatic steatosis in high fructose-fed rats. Nutr. Metab. 2013, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Yin, L.T.; Chang, W.T.; Huang, J.S. Effect of Lactobacillus reuteri GMNL-263 treatment on renal fibrosis in diabetic rats. J. Biosci. Bioeng. 2010, 110, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Ting, W.J.; Kuo, W.W.; Kuo, C.H.; Yeh, Y.L.; Shen, C.Y.; Chen, Y.H.; Ho, T.J.; Viswanadha, V.P.; Chen, Y.H.; Huang, C.Y. Supplementary heat-killed Lactobacillus reuteri GMNL-263 ameliorates hyperlipidaemic and cardiac apoptosis in high-fat diet-fed hamsters to maintain cardiovascular function. Br. J. Nutr. 2015, 114, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Miller, B.; Matthan, N.R.; Goktas, Z.; Wu, D.; Reed, D.B.; Yin, X.; Grammas, P.; Moustaid-Moussa, N.; Shen, C.L.; et al. Aortic cholesterol accumulation correlates with systemic inflammation but not hepatic and gonadal adipose tissue inflammation in low-density lipoprotein receptor null mice. Nutr. Res. 2013, 33, 1072–1082. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Shin, H.S.; Choi, H.S.; Park, J.W.; Jo, I.; Oh, E.S.; Lee, K.Y.; Lee, B.H.; Johnson, R.J.; Kang, D.H. Uric acid induces fat accumulation via generation of endoplasmic reticulum stress and SREBP-1c activation in hepatocytes. Lab. Investig. 2014, 94, 1114–1125. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Hou, S.; Zhang, D.; Xia, H.; Wang, Y.C.; Jiang, J.; Yin, H.; Ying, H. Regulation of fatty acid composition and lipid storage by thyroid hormone in mouse liver. Cell Biosci. 2014, 4, 38. [Google Scholar] [CrossRef] [PubMed]
- Yiu, W.F.; Kwan, P.L.; Wong, C.Y.; Kam, T.S.; Chiu, S.M.; Chan, S.W.; Chan, R. Attenuation of fatty liver and prevention of hypercholesterolemia by extract of Curcuma longa through regulating the expression of CYP7A1, LDL-receptor, HO-1, and HMG-CoA reductase. J. Food Sci. 2011, 76, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, S.; Yerian, L.; Kohli, R.; Xanthakos, S.; Angulo, P.; Ling, S.; Lopez, R.; Christine, C.K.; Feldstein, A.E.; Alkhouri, N. The evaluation of hepatic fibrosis scores in children with nonalcoholic fatty liver disease. Dig. Dis. Sci. 2015, 60, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, W.R.; Kim, H.J.; Therneau, T.M. Association between noninvasive fibrosis markers and mortality among adults with nonalcoholic fatty liver disease in the United States. Hepatology 2013, 57, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Treeprasertsuk, S.; Bjornsson, E.; Enders, F.; Suwanwalaikorn, S.; Lindor, K.D. NAFLD fibrosis score: A prognostic predictor for mortality and liver complications among NAFLD patients. World J. Gastroenterol. 2013, 19, 1219–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, Y.; Kurt, R.; Yonal, O.; Polat, N.; Celikel, C.A.; Gurdal, A.; Oflaz, H.; Ozdogan, O.; Imeryuz, N.; Kalayci, C.; et al. Coronary flow reserve is impaired in patients with nonalcoholic fatty liver disease: Association with liver fibrosis. Atherosclerosis 2010, 211, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.S.; Kim, H.N.; Park, H.J.; Lee, J.E.; Yeo, S.Y.; Yang, J.S.; Park, S.Y.; Yoon, H.S.; Cho, G.S.; Franz, C.M.; et al. Modulation of the murine microbiome with a concomitant anti-obesity effect by Lactobacillus rhamnosus GG and Lactobacillus sakei NR28. Benef. Microbes 2012, 3, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Tomaro-Duchesneau, C.; Jones, M.L.; Shah, D.; Jain, P.; Saha, S.; Prakash, S. Cholesterol assimilation by Lactobacillus probiotic bacteria: An in vitro investigation. CholesteBiomed. Res. Int. 2014, 2014, 380316. [Google Scholar]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740–745. [Google Scholar] [PubMed]
- Jung, J.Y.; Shin, J.S.; Lee, S.G.; Rhee, Y.K.; Cho, C.W.; Hong, H.D.; Lee, K.T. Lactobacillus sakei K040706 evokes immunostimulatory effects on macrophages through TLR 2-mediated activation. Int. Immunopharmacol. 2015, 28, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Fusaru, A.M.; Stănciulescu, C.E.; Surlin, V.; Taisescu, C.; Bold, A.; Pop, O.T.; Baniţă, I.M.; Crăiţoiu, S.; Pisoschi, C.G. Role of innate immune receptors TLR2 and TLR4 as mediators of the inflammatory reaction in human visceral adipose tissue. Rom. J. Morphol. Embryol. 2012, 53, 693–701. [Google Scholar] [PubMed]
- Ikeuchi, M.; Tsutsui, H.; Shiomi, T.; Matsusaka, H.; Matsushima, S.; Wen, J.; Kubota, T.; Takeshita, A. Inhibition of TGF-β signaling exacerbates early cardiac dysfunction but prevents late remodeling after infarction. Cardiovasc. Res. 2004, 64, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Frantz, S.; Hu, K.; Adamek, A.; Wolf, J.; Sallam, A.; Maier, S.K.; Lonning, S.; Ling, H.; Ertl, G.; Bauersachs, J. Transforming growth factor beta inhibition increases mortality and left ventricular dilatation after myocardial infarction. Basic Res. Cardiol. 2008, 103, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Bujak, M.; Ren, G.; Kweon, H.J.; Dobaczewski, M.; Reddy, A.; Taffet, G.; Wang, X.F.; Frangogiannis, N.G. Essential role of Smad3 in infarct healing and in the pathogenesis of cardiac remodeling. Circulation 2007, 116, 2127–2138. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ting, W.-J.; Kuo, W.-W.; Hsieh, D.J.-Y.; Yeh, Y.-L.; Day, C.-H.; Chen, Y.-H.; Chen, R.-J.; Padma, V.V.; Chen, Y.-H.; Huang, C.-Y. Heat Killed Lactobacillus reuteri GMNL-263 Reduces Fibrosis Effects on the Liver and Heart in High Fat Diet-Hamsters via TGF-β Suppression. Int. J. Mol. Sci. 2015, 16, 25881-25896. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161025881
Ting W-J, Kuo W-W, Hsieh DJ-Y, Yeh Y-L, Day C-H, Chen Y-H, Chen R-J, Padma VV, Chen Y-H, Huang C-Y. Heat Killed Lactobacillus reuteri GMNL-263 Reduces Fibrosis Effects on the Liver and Heart in High Fat Diet-Hamsters via TGF-β Suppression. International Journal of Molecular Sciences. 2015; 16(10):25881-25896. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161025881
Chicago/Turabian StyleTing, Wei-Jen, Wei-Wen Kuo, Dennis Jine-Yuan Hsieh, Yu-Lan Yeh, Cecilia-Hsuan Day, Ya-Hui Chen, Ray-Jade Chen, Viswanadha Vijaya Padma, Yi-Hsing Chen, and Chih-Yang Huang. 2015. "Heat Killed Lactobacillus reuteri GMNL-263 Reduces Fibrosis Effects on the Liver and Heart in High Fat Diet-Hamsters via TGF-β Suppression" International Journal of Molecular Sciences 16, no. 10: 25881-25896. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161025881