
Identification of Arsenic Direct-Binding Proteins in Acute Promyelocytic Leukaemia Cells

Abstract
:1. Introduction
2. Results
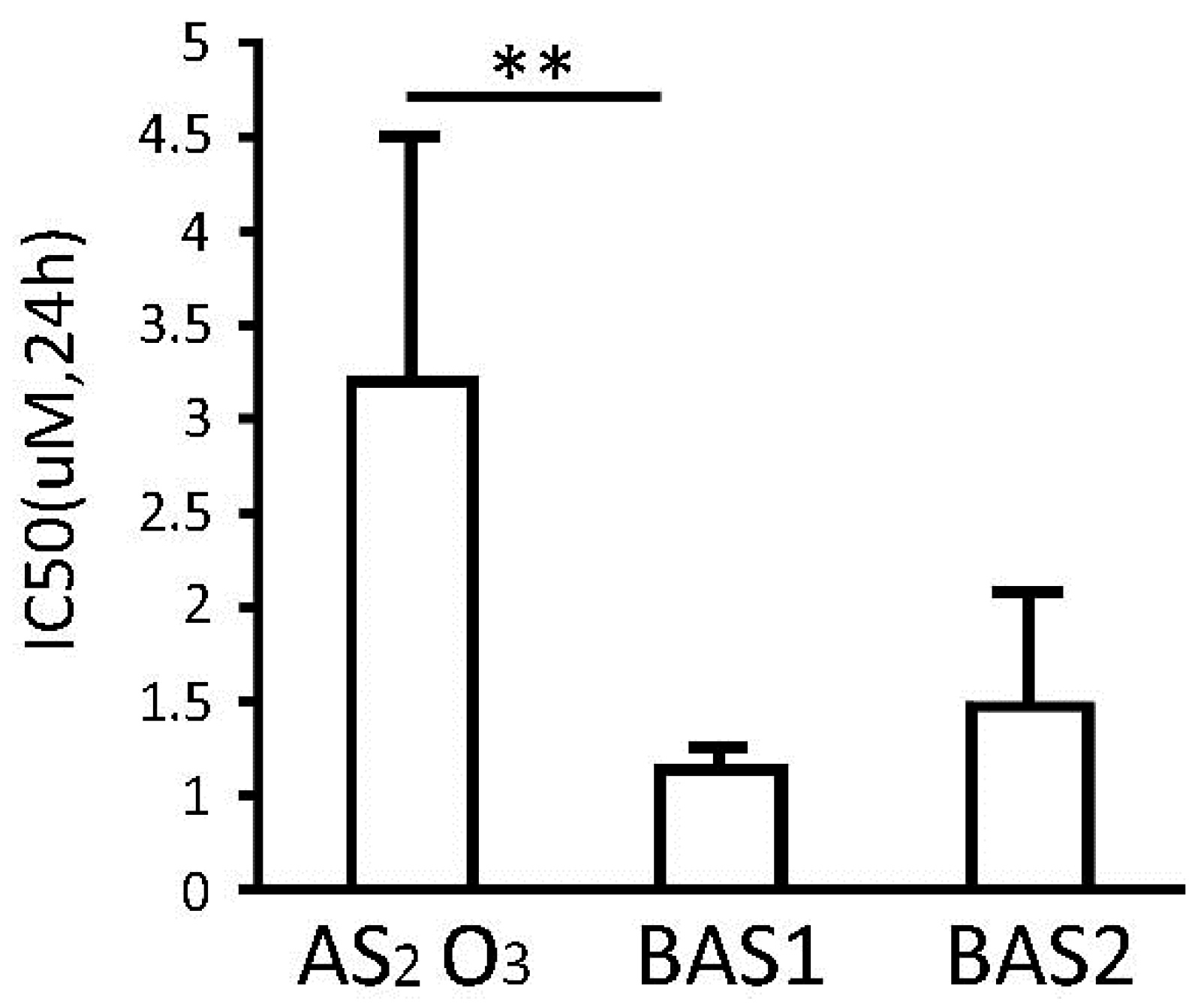
2.1. Arsenic-Biotin Inhibits APL Cell Proliferation

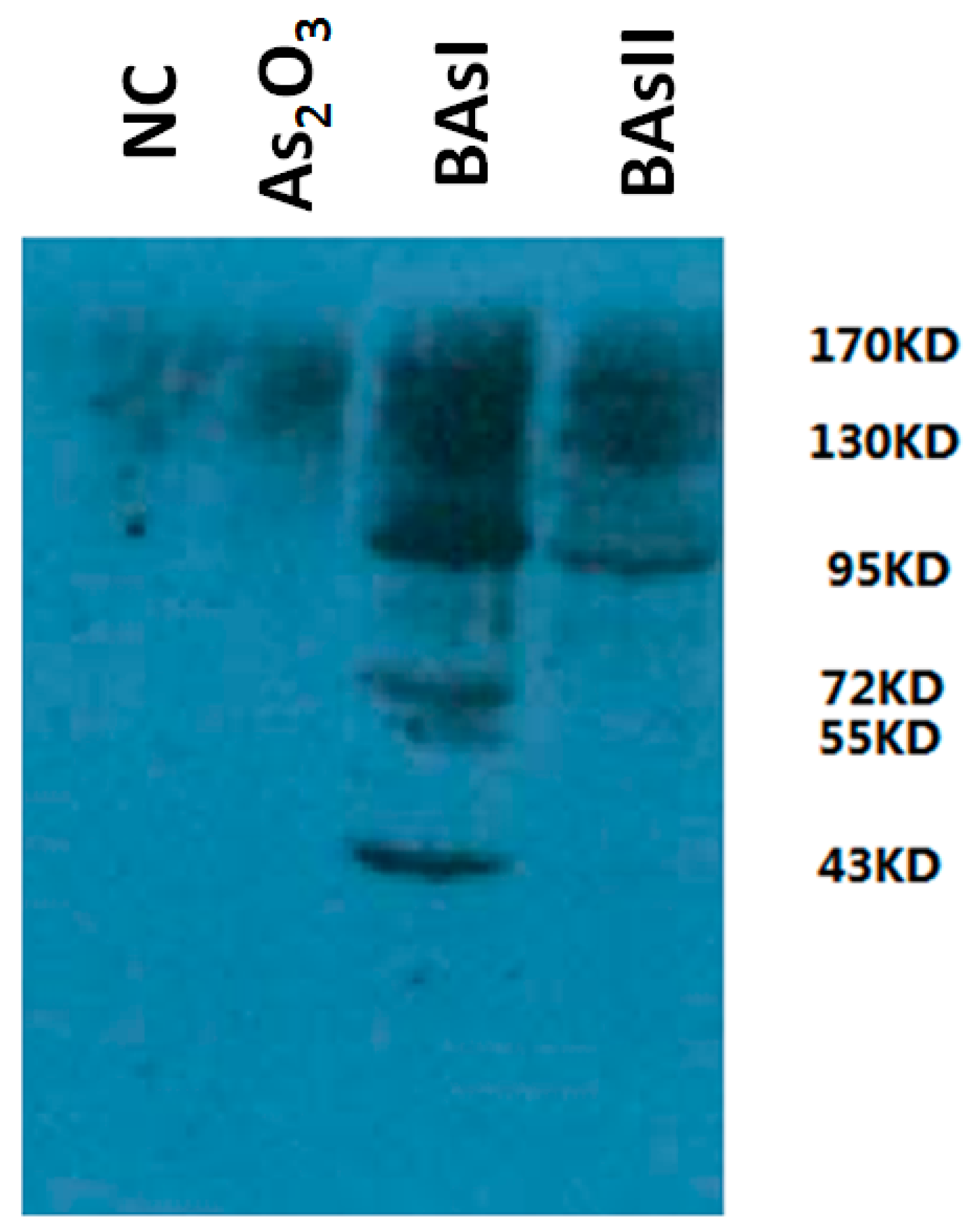
2.2. Measurement of Arsenic-Binding Proteins by Western Blot

2.3. Identification and Characterisation of Arsenic Binding Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Protein |
|---|---|
| Redox-related proteins | GSTP1, PKM2, HSPA9, LEG1 Galectin-1, AT8B4, XRRA1, GAPDH, LCE1B, TET2. |
| DNA-dependent transcription | RL12, Med29, DNA topoisomerase 1, DNA ligase 1, RL21, RS4X, RL23. |
| Regulation of glycometabolism and lipid metabolism | SGSM2, RREB1, NFYC, FBN1, AL1A3, haemoglobin, ACSM4. |
| G-protein coupled receptor family | LPAR1, GPCRs, RXFP2. |
| Inflammation response | Eosinophil peroxidase, NK-tumour recognition protein, RXFP2, DHX8. |
| Cell proliferation and cell cycle | DNLI1, UBP2, LAMB2, Galectin-1, HNRPR, PCDGI, HTRA1, Cytochrome P450. |
| Proteasome homeostasis | UBXN1, PIAS3, ML12, RING finger protein 144A-B. |
2.4. Confirmation of Binding of Redox-Related Proteins to Arsenic

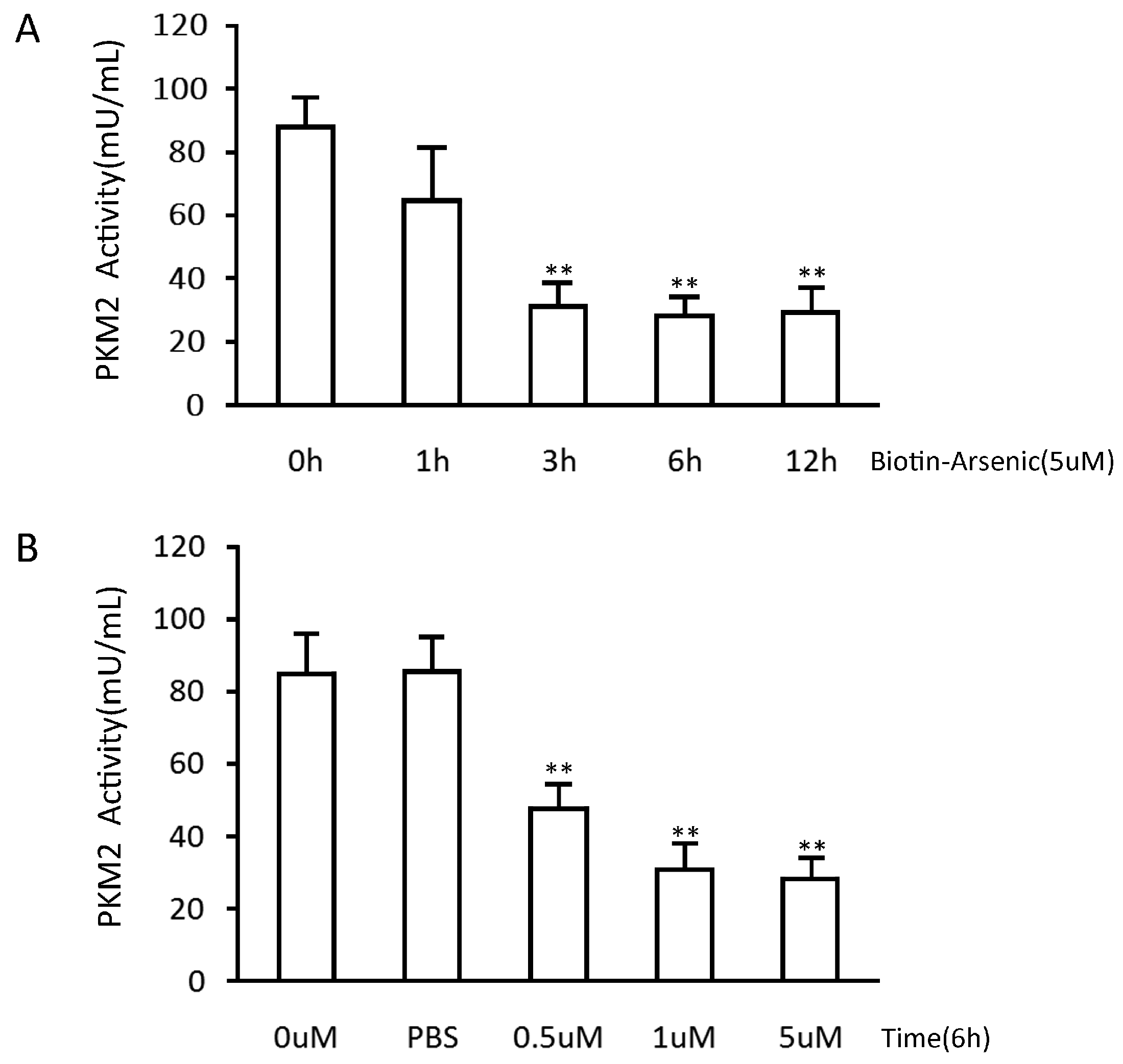
2.5. Arsenic-Biotin Suppressed PKM2 Activity

3. Discussion
4. Materials and Methods
4.1. Chemicals and Cells
4.2. MTT Assay
4.3. Identification of Arsenic Direct-Binding Proteins
4.4. Proteins Extraction and LC-MS/MS
4.5. Database Search
4.6. Arsenic-Binding Protein Analyses
4.7. Pyruvate Kinase Activity Assay
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, Z.Y.; Chen, Z. Acute promyelocytic leukemia: From highly fatal to highly curable. Blood 2008, 111, 2505–2515. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Yan, H.; Yu, T.; Sun, H.P.; Liu, J.X.; Li, X.S.; Wu, W.; Zhang, F.Q.; Chen, Y.; Zhou, L.; et al. Studies on treatment of acute promyelocytic leukemia with arsenic trioxide: Remission induction, follow-up, and molecular monitoring in 11 newly diagnosed and 47 relapsed acute promyelocytic leukemia patients. Blood 1999, 94, 3315–3324. [Google Scholar] [PubMed]
- Zhu, J.; Chen, Z.; Lallemand-Breitenbach, V.; de Thé, H. How acute promyelocytic leukaemia revived arsenic. Nat. Rev. Cancer 2002, 2, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Mathews, V.; George, B.; Lakshmi, K.M.; Viswabandya, A.; Bajel, A.; Balasubramanian, P.; Shaji, R.V.; Srivastava, V.M.; Srivastava, A.; Chandy, M. Single-agent arsenic trioxide in the treatment of newly diagnosed acute promyelocytic leukemia: Durable remissions with minimal toxicity. Blood 2006, 107, 2627–2632. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, B.S. Metalloid compounds as drugs. Res. Pharm. Sci. 2013, 8, 145–158. [Google Scholar] [PubMed]
- Salomoni, P.; Dvorkina, M.; Michod, D. Role of the promyelocytic leukaemia protein in cell death regulation. Cell Death Dis. 2012, 3, 247. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Koken, M.; Guignon, F.; ChelbiAlix, M.K.; Degos, L.; Wang, Z.Y.; Chen, Z.; de Thé, H. Arsenic-induced PML targeting onto nuclear bodies implications for the treatment of acute promyelocytic leukemia. Proc. Natl. Acad. Sci. USA 1997, 94, 3978–3983. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Wang, N.; Weinfeld, M.; Cullen, W.R.; Le, X.C. Identification of arsenic-binding proteins in human cells by affinity chromatography and mass spectrometry. Anal. Chem. 2009, 81, 4144–4152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, F.; Shim, J.Y.; Kirk, K.L.; Anderson, D.E.; Chen, X. Identification of arsenic-binding proteins in human breast cancer cells. Cancer Lett. 2007, 255, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Broome, J.D. Arsenic targets tubulins to induce apoptosis in myeloid leukemia cells. Cancer Res. 1999, 59, 776–780. [Google Scholar] [PubMed]
- Shi, W.; Dong, J.; Scott, R.A.; Ksenzenko, M.Y.; Rosen, B.P. The role of arsenic-thiol interactions in metalloregulation of the ars operon. J. Biol. Chem. 1996, 271, 9291–9297. [Google Scholar] [PubMed]
- Kitchin, K.T.; Wallace, K. Arsenite binding to synthetic peptides based on the Zn finger region and the estrogen binding region of the human estrogen receptor-alpha. Toxicol. Appl. Pharmacol. 2005, 206, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Kapahi, P.; Takahashi, T.; Natoli, G.; Adams, S.R.; Chen, Y.; Tsien, R.Y.; Karin, M. Inhibition of NF-κB activation by arsenite through reaction with a critical cysteine in the activation loop of IκB kinase. J. Biol. Chem. 2000, 275, 36062–36066. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Fanelli, M.; Ferrara, F.F.; Riccioni, R.; Rosenauer, A.; Davison, K.; Lamph, W.W.; Waxman, S.; Pelicci, P.G.; Coco, F.L.; et al. Arsenic trioxide as an inducer of apoptosis and loss of PML/RARα protein in acute promyelocytic leukemia cells. J. Natl. Cancer Inst. 1998, 90, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Lallemand-Breitenbach, V.; Zhu, J.; Puvion, F.; Koken, M.; Honoré, N.; Doubeikovsky, A.; Duprez, E.; Pandolfi, P.P.; Puvion, E.; Freemont, P.; et al. Role of promyelocytic leukemia (PML) SUMOylation in nuclear body formation, 11S proteasome recruitment, and As2O3-induced PML or PML/retinoic acid receptor α Degradation. J. Exp. Med. 2001, 193, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Shi, X.G.; Tang, W.; Xiong, S.M.; Zhu, J.; Cai, X.; Han, Z.G.; Ni, J.H.; Shi, G.Y.; Jia, P.M.; et al. Use of arsenic trioxide (As2O3) in the treatment of acute promyelocytic leukemia (APL): I. As2O3 exerts dose-dependent dual effects on APL cells. Blood 1997, 89, 3345–3353. [Google Scholar] [PubMed]
- Zhang, X.W.; Yan, X.J.; Zhou, Z.R.; Yang, F.F.; Wu, Z.Y.; Sun, H.B.; Liang, W.X.; Song, A.X.; Lallemand-Breitenbach, V.; Jeanne, M.; et al. Arsenic trioxide controls the fate of the PML-RARα oncoprotein by directly binding PML. Science 2010, 328, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; He, P.; Liu, F.; Zhou, N.; Cheng, X.; Shi, L.; Zhu, H.; Zhao, J.; Wang, Y.; Zhang, M. Tetra-arsenic tetra-sulfide (As4S4) promotes apoptosis in retinoid acid -resistant human acute promyelocytic leukemic NB4-R1 cells through downregulation of SET protein. Tumour Biol. 2014, 35, 3421–3430. [Google Scholar] [CrossRef] [PubMed]
- Koshiuka, K.; Elstner, E.; Williamson, E.; Said, J.W.; Tada, Y.; Koeffler, H.P. Novel therapeutic approach: Organic arsenical melarsoprol alone or with all-trans-retinoic acid markedly inhibit growth of human breast and prostate cancer cells in vitro and in vivo. Br. J. Cancer 2000, 82, 452–458. [Google Scholar] [PubMed]
- Charoensuk, V.; Gati, W.P.; Weinfeld, M.; Le, X.C. Differential cytotoxic effects of arsenic compounds in human acute promyelocytic leukemia cells. Toxicol. Appl. Pharmacol. 2009, 239, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Jing, Y.; Styblo, M.; Chen, Z.; Waxman, S. Glutathione-S-transferase π inhibits As2O3-induced apoptosis in lymphoma cells: Involvement of hydrogen peroxide catabolism. Blood 2005, 105, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, S.; Nuccetelli, M.; Noguera, N.I.; Bellincampi, L.; Lunghi, P.; Bonati, A.; Mann, K.; Miller, W.H., Jr.; Federici, G.; Lo Coco, F. Role of GSTP1–1 in mediating the effect of As2O3 in the Acute Promyelocytic Leukemia cell line NB4. Ann. Hematol. 2006, 85, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Ianaro, A.; Maffia, P.; Cuzzocrea, S.; Mazzon, E.; Santoro, M.G.; di Rosa, M.; Ialenti, A. 2-Cyclopenten-1-one and prostaglandin J2 reduce restenosis after balloon angioplasty in rats: Role of NF-κB. FEBS Lett. 2003, 553, 21–27. [Google Scholar] [CrossRef]
- Hulleman, E.; Kazemier, K.M.; Holleman, A.; VanderWeele, D.J.; Rudin, C.M.; Broekhuis, M.J.; Evans, W.E.; Pieters, R.; den Boer, M.L. Inhibition of glycolysis modulates prednisolone resistance in acute lymphoblastic leukemia cells. Blood 2009, 113, 2014–2021. [Google Scholar] [CrossRef] [PubMed]
- Prata, C.; Grasso, C.; Loizzo, S.; Sega, F.V.; Caliceti, C.; Zambonin, L.; Fiorentini, D.; Hakim, G.; Berridge, M.V.; Landi, L. Inhibition of trans-plasma membrane electron transport: A potential anti-leukemic strategy. Leuk Res. 2010, 34, 1630–1635. [Google Scholar] [CrossRef] [PubMed]
- Herst, P.M.; Howman, R.A.; Neeson, P.J.; Berridge, M.V.; Ritchie, D.S. The level of glycolytic metabolism in acute myeloid leukemia blasts at diagnosis is prognostic for clinical outcome. J. Leukoc. Biol. 2011, 89, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shi, Y.; Liu, S.; Cao, Y.; Wang, X.; Tao, Y. PKM2: The thread linking energy metabolism reprogramming with epigenetics in cancer. Int. J. Mol. Sci. 2014, 26, 11435–11445. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.; de Melo, J.; Tang, D. PKM2, a Central Point of Regulation in Cancer Metabolism. Int. J. Cell Biol. 2013, 2013, 242513. [Google Scholar] [CrossRef] [PubMed]
- Pacini, N.; Borziani, F. Cancer stem cell theory and the warburg effect, two sides of the same coin? Int. J. Mol. Sci. 2014, 15, 8893–8930. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Wellen, K.E.; Mazurek, S.; Bamezai, R.N. Pyruvate kinase M2: Regulatory circuits and potential for therapeutic intervention. Curr. Pharm. Des. 2014, 20, 2595–2606. [Google Scholar] [PubMed]
- Xie, H.; Hu, Z.; Chyna, B.; Horrigan, S.K.; Westbrook, C.A. Human mortalin (HSPA9): A candidate for the myeloid leukemia tumor suppressor gene on 5q31. Leukemia 2000, 14, 2128–2134. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Kambal, A.; Krysiak, K.; Walshauser, M.A.; Raju, G.; Tibbitts, J.F.; Walter, M.J. Knockdown of Hspa9, a del(5q31.2) gene, results in a decrease in hematopoietic progenitors in mice. Blood 2011, 117, 1530–1539. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Liao, C.L.; Lin, Y.P.; Chen, S.C.; Wang, D.L. S-nitrosoproteome in endothelial cells revealed by a modified biotin switch approach coupled with Western blot-based two-dimensional gel electrophoresis. J. Proteome Res. 2009, 8, 4835–4843. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Lu, H.; Li, W.; Hu, R.; Chen, Z. Identification of Arsenic Direct-Binding Proteins in Acute Promyelocytic Leukaemia Cells. Int. J. Mol. Sci. 2015, 16, 26871-26879. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161125994
Zhang T, Lu H, Li W, Hu R, Chen Z. Identification of Arsenic Direct-Binding Proteins in Acute Promyelocytic Leukaemia Cells. International Journal of Molecular Sciences. 2015; 16(11):26871-26879. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161125994
Chicago/Turabian StyleZhang, Tao, Haojie Lu, Weijun Li, Ronggui Hu, and Zi Chen. 2015. "Identification of Arsenic Direct-Binding Proteins in Acute Promyelocytic Leukaemia Cells" International Journal of Molecular Sciences 16, no. 11: 26871-26879. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161125994