
A PPO Promoter from Betalain-Producing Red Swiss Chard, Directs Petiole- and Root-Preferential Expression of Foreign Gene in Anthocyanins-Producing Plants

Abstract
:1. Introduction
2. Results
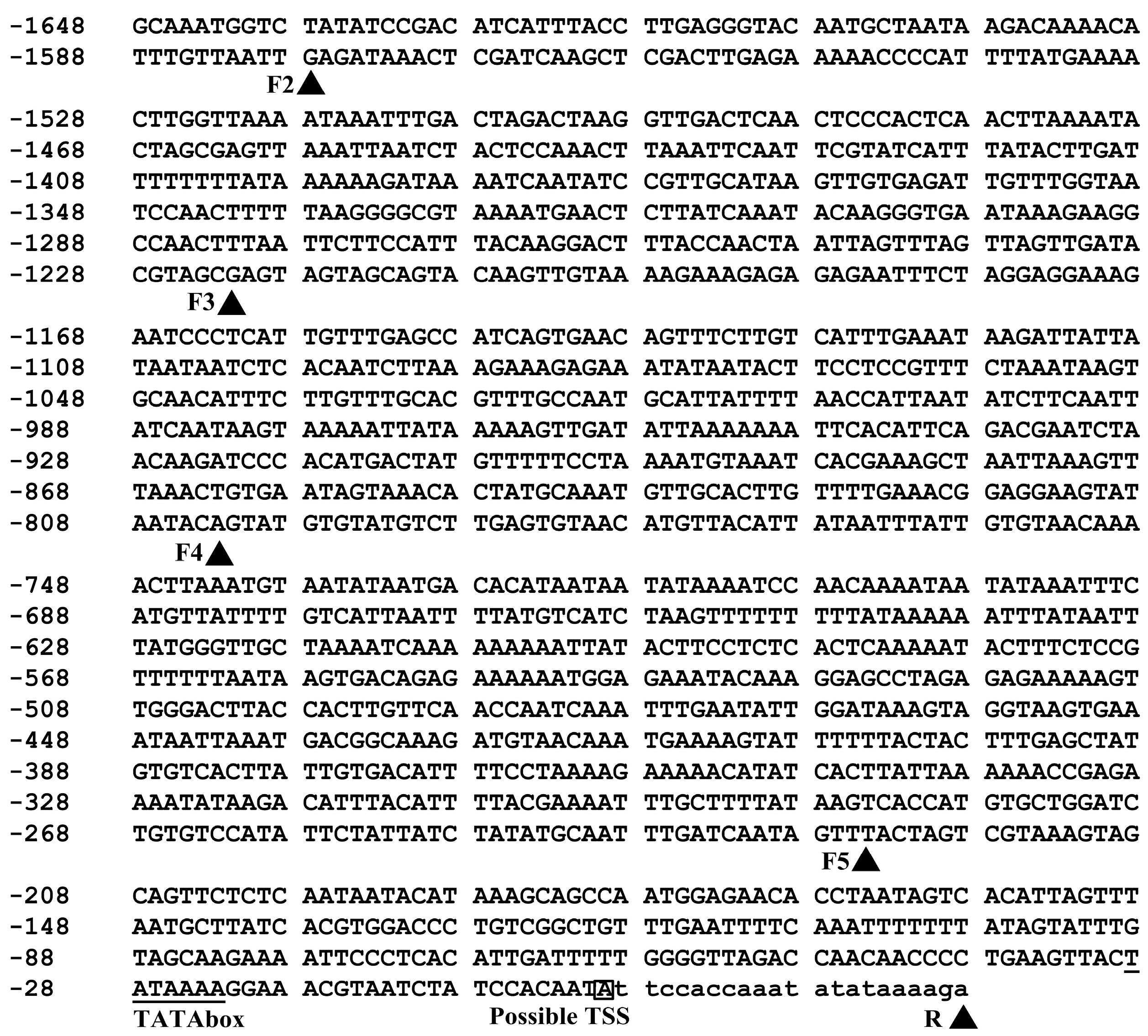
2.1. Cloning and Sequence Analysis of the BvcPPOP Promoter

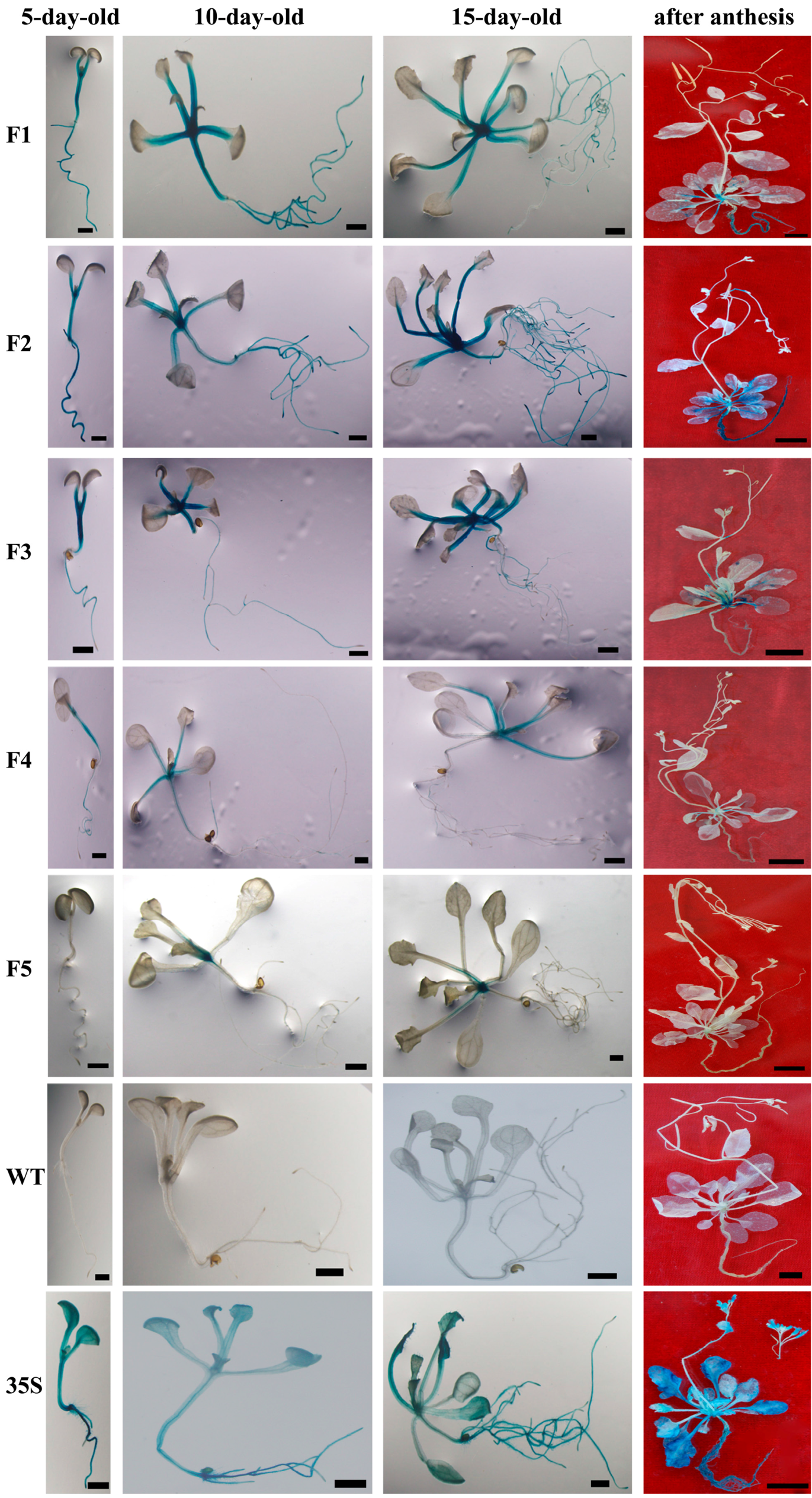
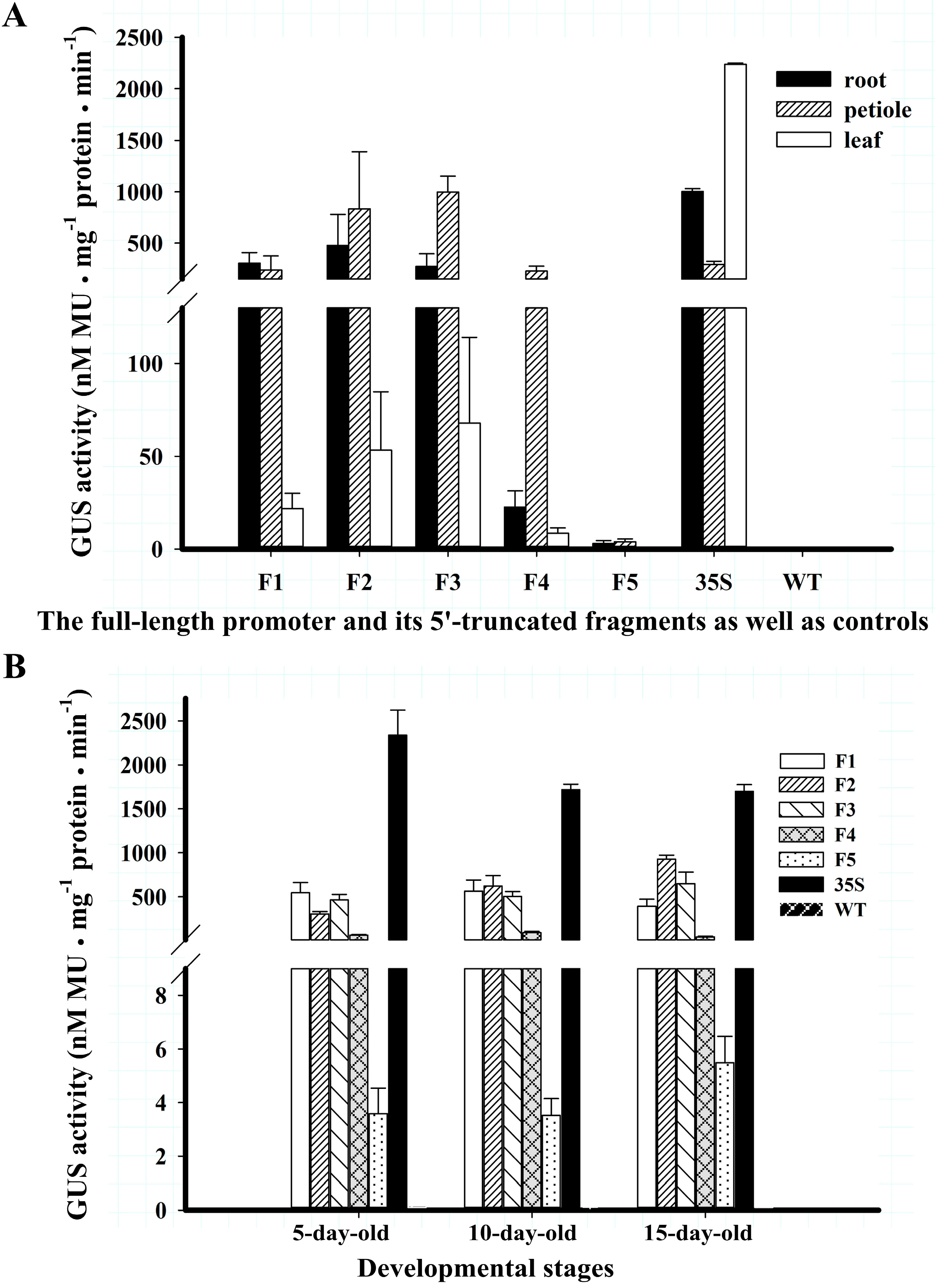
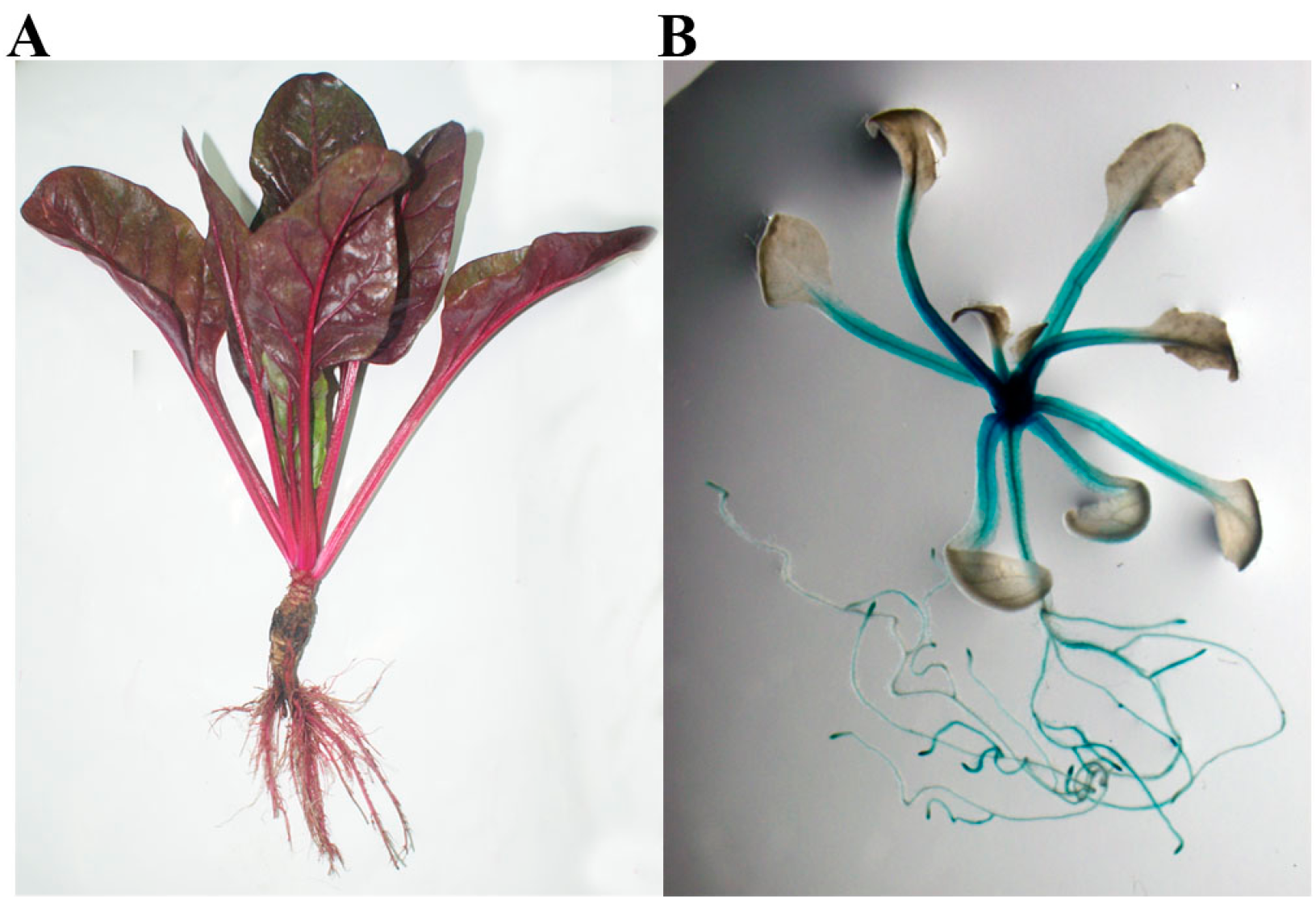
2.2. BvcPPOP Directed Petiole- and Root-Preferential Expression of GUS Gene at the Vegetative Stage of Transgenic Arabidopsis

2.3. BvcPPOP-Directed GUS Expression Is Regulated by Developmental Stages

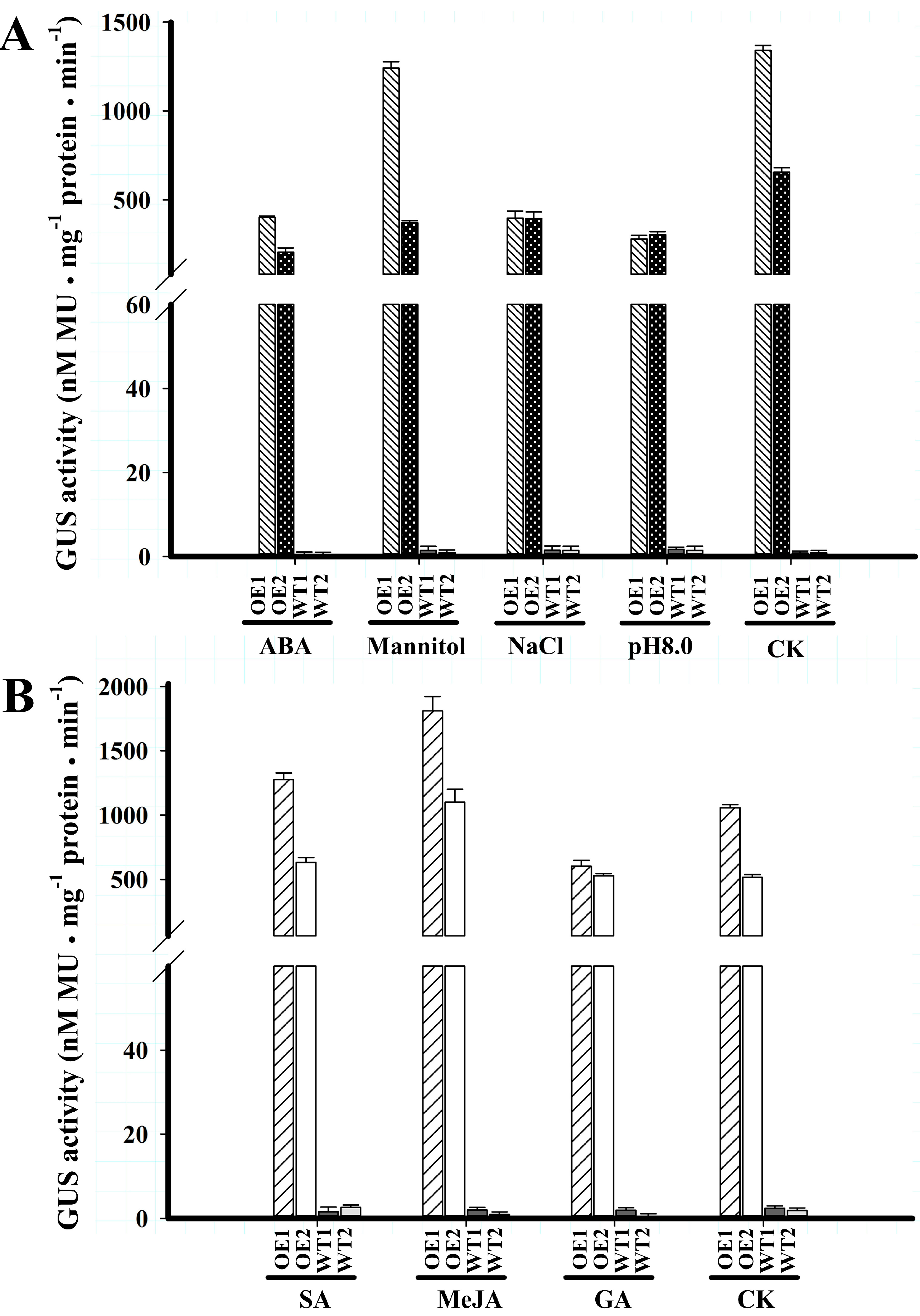
2.4. BvcPPOP-Directed GUS Expression Is Differently Regulated by Abiotic Stressors

2.5. 5′-Truncated BvcPPOPs Display Various Expression Patterns and/or Levels in Transgenic Arabidopsis
3. Discussion

4. Experimental Section
4.1. Plant Materials and Growth Conditions
4.2. Isolation of the Promoter and Its Deletion Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′ to 3′) * |
|---|---|
| Forward | |
| FW1 | AAGCTTGCAAATGGTCTATATCCGAC |
| FW2 | AAGCTTGAGATAAACTCGATCAAGCTCG |
| FW3 | AAGCTTGAGTAGTAGCAGTACAAGTTG |
| FW4 | AAGCTTAGTATGTGTATGTCTTGAGTG |
| FW5 | AAGCTTTACTAGTCGTAAAGTAGCAGTTCTC |
| Reverse | |
| RW | GGATCCTCTTTTATATATTTGGTGGA |
4.3. Plant Expression Vector Construction and Agrobacterium-Mediated Transformation
4.4. Analysis of GUS Expression
4.5. Abiotic Stress Treatment
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thipyapong, P.; Stout, M.J.; Attajarusit, J. Functional analysis of polyphenol oxidases by antisense/sense technology. Molecules 2007, 12, 1569–1595. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.L. Beyond brown: Polyphenol oxidases as enzymes of plant specialized metabolism. Front. Plant Sci. 2014, 5, 783. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Steffens, J.C. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta 2002, 215, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Constabel, C.P. Polyphenol oxidase overexpression in transgenic populus enhances resistance to herbivory by forest tent caterpillar (Malacosoma disstria). Planta 2004, 220, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Thipyapong, P.; Hunt, M.D.; Steffens, J.C. Antisense downregulation of polyphenol oxidase results in enhanced disease susceptibility. Planta 2004, 220, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Van Ranst, G.; Lee, M.R.F.; Fievez, V. Red clover polyphenol oxidase and lipid metabolism. Animal 2011, 5, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Wahler, D.; Gronover, C.S.; Richter, C.; Foucu, F.; Twyman, R.M.; Moerschbacher, B.M.; Fischer, R.; Muth, J.; Prufer, D. Polyphenoloxidase silencing affects latex coagulation in taraxacum species. Plant Physiol. 2009, 151, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Taketa, S.; Matsuki, K.; Amano, S.; Saisho, D.; Himi, E.; Shitsukawa, N.; Yuo, T.; Noda, K.; Takeda, K. Duplicate polyphenol oxidase genes on barley chromosome 2H and their functional differentiation in the phenol reaction of spikes and grains. J. Exp. Bot. 2010, 61, 3983–3993. [Google Scholar] [CrossRef] [PubMed]
- Thipyapong, P.; Steffens, J.C. Tomato polyphenol oxidase (differential response of the polyphenol oxidase F promoter to injuries and wound signals). Plant Physiol. 1997, 115, 409–418. [Google Scholar] [PubMed]
- Newman, S.M.; Tantasawat, P.; Steffens, J.C. Tomato polyphenol oxidase B is spatially and temporally regulated during development and in response to ethylene. Molecules 2011, 16, 493–517. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.M.; Chandrashekar, A.; Venkatesh, Y.P. Promoter analyses and transcriptional profiling of eggplant polyphenol oxidase 1 gene (SmePPO1) reveal differential response to exogenous methyl jasmonate and salicylic acid. J. Plant Physiol. 2012, 169, 718–730. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.; Dirks, M.E.; Gronover, C.S.; Prufer, D.; Moerschbacher, B.M. Silencing and heterologous expression of PPO-2 indicate a specific function of a single polyphenol oxidase isoform in resistance of dandelion (Taraxacum officinale) against Pseudomonas syringae pv. tomato. Mol. Plant Microbe Interact. 2012, 25, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; O’Hare, T.J.; Jobin-Decor, M.; Underhill, S.J.; Wills, R.B.; Graham, M.W. Transcriptional regulation of a pineapple polyphenol oxidase gene and its relationship to blackheart. Plant Biotechnol. J. 2003, 1, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Naveed, I.; Mahmood, T. In silico characterization of rice polyphenol oxidase gene promoters. Arch. Sci. 2012, 65, 297–304. [Google Scholar]
- Quarta, A.; Mita, G.; Durante, M.; Arlorio, M.; de Paolis, A. Isolation of a polyphenol oxidase (PPO) cDNA from artichoke and expression analysis in wounded artichoke heads. Plant Physiol. Biochem. 2013, 68, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Deng, Z.; Liu, C.; Zhao, M.; Guo, H.; Xia, Z.; Liu, H. Molecular cloning, expression profiles, and characterization of a novel polyphenol oxidase (PPO) gene in Hevea brasiliensis. Biosci. Biotechnol. Biochem. 2014, 78, 1648–1655. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Yonekura-Sakakibara, K.; Sato, T.; Kikuchi, S.; Fukui, Y.; Fukuchi-Mizutani, M.; Ueda, T.; Nakao, M.; Tanaka, Y.; Kusumi, T.; et al. Aureusidin synthase: A polyphenol oxidase homolog responsible for flower coloration. Science 2000, 290, 1163–1166. [Google Scholar] [CrossRef] [PubMed]
- Kaintz, C.; Molitor, C.; Thill, J.; Kampatsikas, I.; Michael, C.; Halbwirth, H.; Rompel, A. Cloning and functional expression in E. coli of a polyphenol oxidase transcript from Coreopsis grandiflora involved in aurone formation. FEBS Lett. 2014, 588, 3417–3426. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.H.; Moinuddin, S.G.A.; Helms, G.L.; Hishiyama, S.; Eichinger, D.; Davin, L.B.; Lewis, N.G. (+)-Larreatricin hydroxylase, an enantio-specific polyphenol oxidase from the creosote bush (Larrea tridentata). Proc. Natl. Acad. Sci. USA 2003, 100, 10641–10646. [Google Scholar] [CrossRef] [PubMed]
- Araji, S.; Grammer, T.A.; Gertzen, R.; Anderson, S.D.; Mikulic-Petkovsek, M.; Veberic, R.; Phu, M.L.; Solar, A.; Leslie, C.A.; Dandekar, A.M.; et al. Novel roles for the polyphenol oxidase enzyme in secondary metabolism and the regulation of cell death in walnut. Plant Physiol. 2014, 164, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Clement, J.; Mabry, T. Pigment evolution in the caryophyllales: A systematic overview. Bot. Acta 1996, 109, 360–367. [Google Scholar] [CrossRef]
- Mueller, L.A.; Hinz, U.; Zrÿd, J.-P. Characterization of a tyrosinase from Amanita muscaria involved in betalain biosynthesis. Phytochemistry 1996, 42, 1511–1515. [Google Scholar] [CrossRef]
- Steiner, U.; Schliemann, W.; Strack, D. Assay for tyrosine hydroxylation activity of tyrosinase from betalain-forming plants and cell cultures. Anal. Biochem. 1996, 238, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Strack, D.; Vogt, T.; Schliemann, W. Recent advances in betalain research. Phytochemistry 2003, 62, 247–269. [Google Scholar] [CrossRef]
- Moreno, D.A.; García-Viguera, C.; Gil, J.I.; Gil-Izquierdo, A. Betalains in the era of global agri-food science, technology and nutritional health. Phytochem. Rev. 2008, 7, 261–280. [Google Scholar] [CrossRef]
- Han, X.H.; Gao, Z.J.; Xiao, X.G. Enzymes and genes involved in the betalain biosynthesis in higher plants. Afr. J. Biotechnol. 2009, 8, 6735–6744. [Google Scholar]
- Brockington, S.F.; Walker, R.H.; Glover, B.J.; Soltis, P.S.; Soltis, D.E. Complex pigment evolution in the caryophyllales. New Phytol. 2011, 190, 854–864. [Google Scholar] [CrossRef] [PubMed]
- Gandia-Herrero, F.; Garcia-Carmona, F. Biosynthesis of betalains: Yellow and violet plant pigments. Trends Plant Sci. 2013, 18, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Giridhar, P. Plant betalains: Chemistry and biochemistry. Phytochemistry 2015, 117, 267–295. [Google Scholar] [CrossRef] [PubMed]
- Joy, R.W.T.; Sugiyama, M.; Fukuda, H.; Komamine, A. Cloning and characterization of polyphenol oxidase cDNAs of Phytolacca americana. Plant Physiol. 1995, 107, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, H.; Matsui, T.; Tomiyasu, R.; Kuroda, Y.; Higashimoto, Y.; Oda, T.; Miyasaka, H.; Okuhata, H.; Tanaka, S.; Harada, K. Isolation of polyphenol oxidase genes from Portulaca oleracea and evaluation of their ability to metabolize endocrine-disrupting chemicals. Plant Biotechnol. 2012, 29, 351–357. [Google Scholar] [CrossRef]
- Gao, Z.J.; Han, X.H.; Xiao, X.G. Purification and characterisation of polyphenol oxidase from red swiss chard (Beta vulgaris subspecies cicla) leaves. Food Chem. 2009, 117, 342–348. [Google Scholar] [CrossRef]
- Xiao, X.G.; Gao, Z.J.; Han, X.H.; Liu, J.B.; Nie, X.L. Tyrosinase and its encoding gene in betalain-producing plants and their use thereof. China patent ZL201010263562.7, 2010. [Google Scholar]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (place) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Hatlestad, G.J.; Sunnadeniya, R.M.; Akhavan, N.A.; Gonzalez, A.; Goldman, I.L.; McGrath, J.M.; Lloyd, A.M. The beet R locus encodes a new cytochrome p450 required for red betalain production. Nat. Genet. 2012, 44, 816–820. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M. Polyphenol oxidases in plants and fungi: Going places? A review. Phytochemistry 2006, 67, 2318–2331. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Paterson, A.H.; Brubaker, C.L.; Wendel, J.F. A rapid method for extraction of cotton (Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis. Plant Mol. Biol. Rep. 1993, 11, 122–127. [Google Scholar] [CrossRef]
- Chen, H.; Nelson, R.; Sherwood, J. Enhanced recovery of transformants of Agrobacterium tumefaciens after freeze-thaw transformation and drug selection. Biotechniques 1994, 16, 664–668, 670. [Google Scholar] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: β-Glucuronidase as a sensitive and versatile gene fusion marker in higher-plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [PubMed]
- Han, L.; Han, Y.-N.; Xiao, X.-G. Truncated cotton subtilase promoter directs guard cell-specific expression of foreign genes in tobacco and arabidopsis. PLoS ONE 2013, 8, e59802. [Google Scholar] [CrossRef] [PubMed]
- Geng, A.Q.; Zhao, Z.J.; Nie, X.L.; Xiao, X.G. Expression analysis of four flower-specific promoters of Brassica spp. in the heterogeneous host tobacco. Afr. J. Biotechnol. 2009, 8, 5193–5200. [Google Scholar]
- Chen, L.; Jiang, B.; Wu, C.; Sun, S.; Hou, W.; Han, T. GmPRP2 promoter drives root-preferential expression in transgenic arabidopsis and soybean hairy roots. BMC Plant Biol. 2014, 14, 245. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.-H.; Han, Y.-N.; Xiao, X.-G. A PPO Promoter from Betalain-Producing Red Swiss Chard, Directs Petiole- and Root-Preferential Expression of Foreign Gene in Anthocyanins-Producing Plants. Int. J. Mol. Sci. 2015, 16, 27032-27043. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161126011
Yu Z-H, Han Y-N, Xiao X-G. A PPO Promoter from Betalain-Producing Red Swiss Chard, Directs Petiole- and Root-Preferential Expression of Foreign Gene in Anthocyanins-Producing Plants. International Journal of Molecular Sciences. 2015; 16(11):27032-27043. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161126011
Chicago/Turabian StyleYu, Zhi-Hai, Ya-Nan Han, and Xing-Guo Xiao. 2015. "A PPO Promoter from Betalain-Producing Red Swiss Chard, Directs Petiole- and Root-Preferential Expression of Foreign Gene in Anthocyanins-Producing Plants" International Journal of Molecular Sciences 16, no. 11: 27032-27043. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161126011